Celula, Estructura y Funcion, TEXTO
20
CÉLULA, ESTRUCTURA Y FUNCION Dra. en C. Martha Leticia Ornelas-Arana Est. Carolina Pérez-Ornelas Dr. en C. Guillermo Pérez-García Laboratorio de Bioquímica, DBMG, CUCS, UDG Servicio de Genética, Hospital Civil “Fray Antonio Alcalde” Cuerpo Académico UDG-CA-80 Enfermedades Metabólicas
-
Upload
carlos-acero -
Category
Documents
-
view
784 -
download
0
Transcript of Celula, Estructura y Funcion, TEXTO

CÉLULA, ESTRUCTURA Y FUNCION
Dra. en C. Martha Leticia Ornelas-AranaEst. Carolina Pérez-OrnelasDr. en C. Guillermo Pérez-García
Laboratorio de Bioquímica, DBMG, CUCS, UDGServicio de Genética, Hospital Civil “Fray Antonio Alcalde”Cuerpo Académico UDG-CA-80 Enfermedades Metabólicas

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
CÉLULA, ESTRUCTURA Y FUNCIÓN Martha Leticia Ornelas-Arana, Carolina Pérez-Ornelas, Guillermo Pérez-García
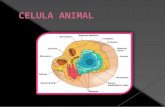
La palabra “célula” fue utilizada en el sentido biológico hace aproximadamente 300 años. Robert Hooke, utilizando un microscopio de su propia construcción, se dio cuenta que el corcho y los tejidos vegetales de otras plantas, estaban formados de pequeñas cavidades separadas por paredes denominándolas como “cella”, que en latín significa celda. En 1838, Matthias Schleiden, llegó a la conclusión de que todos los tejidos vegetales estaban formados por células. En el siguiente año, Theodor Schwann, extendió las observaciones de Schleiden a los tejidos animales y propuso la base celular para todas las formas vivientes. La célula es considerada como una unidad estructural y funcional fundamental, reproductora, viviente, que contiene subestructuras u organelos (Figura 1). Desde el punto de vista de complejidad bioquímico-morfológico se reconocen dos tipos generales de células: procariotas y eucariotas. Las procariotas son células relativamente simples, que llevan a cabo sus procesos metabólicos en su citoplasma y en consecuencia no presentan sus funciones compartamentalizadas en organelos, ejemplos de éstas células son las bacterias y espiroquetas, las algas verdeazules (reino monera), así como micoplasmas (parte del reino fungi). Las células eucariotes presentan compartimientos con funciones definidas, denominados organelos, limitados en su mayoría por membranas, aunque existen procesos metabólicos que ocurren en la matriz celular; éstas células están presentes en todos los organismos superiores, vegetales y animales, en hongos, protozoarios y la mayoría de las algas. Se ha estimado que ser humano tiene 1014 células.
Membranacelular
Lisosoma
Centriolo
Peroxisomas
Nucleolo
Vesículas
Núcleo
Polisomas
Aparato deGolgi
Microtúbulos y microfilamentosMitocondria
Retículo endoplásmicoliso
Retículo endoplásmicorugoso
Figura 1. Estructura general de la célula de mamíferos

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Membrana celular Está formada por una bicapa de fosfolípidos y proteínas globulares (incluidas en la membrana). Los principales fosfolípidos localizados en las membranas celulares son: fosfatidilcolina, fosfatidiletanolamina, fosfatidilinositol y fosfatidilserina; también existen ciertos lípidos neutros como el colesterol. Las proteínas globulares se localizan en la superficie de una u otra de las capas (proteínas extrínsecas), otras pueden atravesar total o parcialmente la membrana (proteínas intrínsecas). Las proteínas se desplazan bidimensionalmente en la bicapa lipídica, aunque algunas permanecen ancladas al citoesqueleto y funcionan como acarreadoras de moléculas a través de la membrana, o como receptores para ciertas hormonas, entre otras funciones (Figuras 2 y 3).
La membrana celular también presenta carbohidratos localizados en la superficie externa de la membrana, unidos a proteínas (glucoproteínas) o lípidos (glucolípidos). La capa formada en la superficie celular por dichos carbohidratos, recibe el nombre de glucocálix.
Fosfolípidos
Bicapa de fosfolípidos
Membranacelular
Colesterol
Carbohidratos
Intracelular
Extracelular
Proteínas periféricas
Proteínaintegral
Figura 2. Estructura de la membrana celular
Este modelo de membrana fue propuesto por Singer y Nicolson en 1972, al cual denominaron modelo de mosaico fluido. Este modelo de membrana también lo presentan todos los organelos a excepción de los ribosomas. Es importante señalar que el porcentaje y tipo de fosfolípidos, colesterol, proteínas y carbohidratos presentes en las membranas, es distinto para cada organelo y membrana celular, también existen diferencias entre tejidos y de acuerdo a la función que realizan.

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Bicapa de fosfolípidos
Fosfolípidos
Proteínaintegral
Carbohidratos
Intracelular
Extracelular
Proteínaperiférica
Membranacelular
Colesterol
Figura 3. Estructura tridimensional de la membrana celular
Mitocondria Generalmente hay de 1 a 1000 mitocondrias por célula, según su naturaleza y función. Habitualmente tienen forma elipsoide, filamentosa o esférica. En algunas células, las mitocondrias permanecen fijas, mientras que en otras se mueven o navegan por todo el citoplasma. Respecto a su origen evolutivo, se considera que hace aproximadamente uno o dos mil millones de años, cuando la tierra tenía escaso oxígeno, una célula primitiva anaeróbica se tragó a una bacteria más pequeña con la capacidad de respirar (síntesis de ATP), que al no ser eliminada, se desarrolló una simbiosis, que se supone, evolucionó hasta la célula actual. La mitocondria tiene una membrana externa y otra interna, como consecuencia hace que se distingan dos espacios: el intermembranoso y la matriz (Figura 4). Posiblemente debido a que las mitocondrias son descendientes de bacterias que vivían libremente, tiene su propio material genético y la maquinaria para expresarlo (DNA, RNAm, RNAt, RNAr y ribosomas. La principal función de la mitocondria es sintetizar ATP a través de la cadena respiratoria y fosforilación oxidativa. Las moléculas que forman parte de la cadena respiratoria se localizan en la membrana interna y la ATPasa también se localiza en la membrana interna (Figura 5). Una célula hepática típica, contiene desde 800 a más de 2500 mitocondrias, que ocupan más del 20% del volumen de la célula. Mientras que en un espermatozoide hay de 10 a 12 mitocondrias.
Lisosoma Es una vesícula rodeadas por una membrana del tipo del mosaico fluido, está
constituida al menos por 50 enzimas hidrolíticas (proteasas, nucleasas, glucosidasas, arilsulfatasas, lipasas, fosfolipasas y fosfatasas) (Figura 6).

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
La función del lisosoma en la célula es la de destruir las partículas (bacterias, virus, etc.) fagocitadas a través de sus enzimas hidrolasas ácidas. Cuando la célula fagocita, se forma una vesícula en el citoplasma, posteriormente el lisosoma se fusiona con la vesícula endocitada y se le llama fagolisosoma, de esta forma las enzimas lisosomales destruyen las partículas o bacterias endocitadas. El residuo de la digestión es exocitado como producto de excreción o permanece como cuerpo residual (Figura 7).
Espacio intermembranoso
Matriz
Genes mitocondriales
2 ARNr
22 ARNt
13 ARNm
12S 16S
Citocromo oxidasa C subunidad I Citocromo oxidasa C subunidad II Citocromo oxidasa C subunidad III ATPasa subunidad 6 ATPasa subunidad 8 Citocromo b NADH deshidrogenasas ( ND1, ND2, ND3, ND4, ND4L, ND5, ND6 )
ADN mitocondrial 16,569 pb
Membrana mitocondrial externa
Membrana mitocondrial interna
Figura 4. Estructura de la mitocondria y DNA mitocondrial
Reconocimiento de las enzimas lisosomales Las enzimas que entran al lisosoma para ser reconocidas por el receptor localizado en la membrana lisosomal deben tener cierto tipo de características bioquímicas. Las enzimas lisosomales que pueden entrar a la célula por pinocitosis son fosfoglucoproteínas.

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Más específicamente, un fosfomonoéster de manosa parece ser el marcador de reconocimiento para muchas enzimas lisosomales (Figura 8).
Membranamitocondrial
interna
matrizmitocondrial
Espaciointermembranoso
FMNH2
NADH + H+
2H+
2H+
2H+
2H+ 2H+
2H+
QH+
QH+
Q
1 +/ O + 2H2 2
H O2
NAD+
FeS
FeS b
Fo
F1
ADP + P1
QH2
ATP
1cc
a
a3 bCu
ND1ND2
ND3
ND4
ND4L
ND5
ND6
CoQCit b
Cit c
Cox I
Cox II
Cox III A6
A8
Complejo I
Matriz
2H+
2H+2H+
2H+
2H+
Espaciointermembranoso
Succinato Fumarato
Complejo II Complejo III
H 02
Complejo IV
O2
Complejo VNo de subunidadescodificadas por elDNA mitocondrial 7 0 1 3 2
No de subunidadescodificadas por elDNA nuclear ~39 4 10 10 ~14
ADP ATP
Figura 5. Las enzimas y moléculas que participan en la cadena respiratoria y fosforilación oxidativa se localizan en la membrana mitocondrial interna.
Dos enzimas están involucradas en la generación del monosacárido fosfomanosil en las hidrolasas ácidas lisosomales que sirven como marcadores de reconocimiento específico para captar estas enzimas por los lisosomas: la enzima, N-acetilglucosamina-1-fosfotransferasa (GlcNAc-1-P transferasa; GNPTA) y la enzima acetilglucosaminil

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
fosfodiesterasa. La enzima GlcNAc-1-P transferasa une una molécula de N-acetilglucosamina-1-P al monosacárido manosa de múltiples enzimas lisosomales. La diesterasa expone el marcador manosa-6-P.
El receptor para las enzimas lisosomales necesario para la transferencia de las enzimas a los lisosomas esta presente en todos los tejidos.
Génesis de los lisosomas Los lisosomas primarios se originan de vesículas que se desprenden del aparato de
Golgi. La función de los lisosomas es degradar material endocitado por fagocitosis o pinocitosis.
Lisosoma
Enzimaslisosomales
Figura 6. Estructura del lisosoma
Lisosoma
Fagolisosoma
Vesículas
Figura 6. Participación de los lisosomas en la fagocitosis

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Célula
-GlcNAc-1-P
enzima manosa GlcNAc-1-P transferasa
E M
receptor para enzimas lisosomales
lisosoma
Lisosoma
Figura 8. Reconocimiento de las enzimas lisosomales mediante receptores de membrana
Ribosomas Los ribosomas son organelos no membranosos. Están formados por dos subunidades
que se reconocen como la subunidad menor y la mayor. En los ribosomas de los eucariotes la subunidad menor es la 40S, la subunidad mayor la 60S y el ribosoma es 80S. En los procariotes la subunidad menor es la 30S, la mayor 50S y el ribosoma es 70S (Figura 9). El ribosoma de las bacterias se divide en dos dominios, el dominio traduccional y el dominio de salida y se reconocen varios centros activos. En la subunidad 30S se une el RNAm y el complejo factor de iniciación iniciador-tRNA, posteriormente se une a la subunidad 50S. El ribosoma 70S tiene los sitios A y P donde se une el RNAt. La peptidil transferasa se localiza en la subunidad 50S. El sitio EF-G, responsable de la translocación

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
del ribosoma se localiza en la subunidad 50S. En el dominio de salida se localiza el sitio de salida, en la subunidad 50S, este sitio de salida sirve para unir el ribosoma a la membrana del RER y es también el sitio por donde sale la proteína recién sintetizada y entra a través de la membrana hasta la luz del RER (Figura 10).
Los ribosomas están formados por proteínas ribosomales y RNA ribosomal (RNAr) (Tabla 1). Las subunidades del ribosoma se sintetizan en el nucleolo y llegan al citoplasma a través de los poros nucleares.
La función de los ribosomas es la síntesis de proteínas (traducción). Los ribosomas que se localizan en el retículo endoplásmico rugoso sintetizan proteínas de exportación, en cambio, los ribosomas del citoplasma (polirribosomas o polisomas) sintetizan proteínas para la propia célula y los ribosomas de las mitocondrias sintetizan proteínas codificadas por los genes mitocondriales para la mitocondria. Los ribosomas también se localizan en la membrana nuclear externa.
Eucariotes Procariotes
40S30S
60S 50S
80S 70S
Figura 9. Subunidades de los ribosomas de eucariotes y procariotes
Tabla 1. Estructura de los ribosomas Ribosomas Subunidades Tipos de
RNAr Número de proteínas
Mamíferos 80S 60% RNA
40S
60S
18S
28S 5.8S 5S
33
49
Bacterias 70S 66% RNA
30S
50S
16S
23S 5S
21
31

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Retículo endoplásmico rugoso El retículo endoplásmico rugoso (RER) es una estructura membranosa formada por cisternas interconectadas entre si, la membrana es del tipo del modelo del mosaico fluido. Contiene ribosomas, lo que le da el aspecto “rugoso” cuando se observa con el microscopio electrónico (Figura 11). El RER es continuación de la membrana nuclear externa. El RER en la superficie de la membrana contiene un receptor que se une a la subunidad mayor del ribosoma y un poro adyacente que permite que la proteína recién sintetizada entre al lumen del RER. La función del RER es la síntesis de proteínas de exportación. Las proteínas sintetizadas sufren cambios en su estructura (modificaciones postraducción) que consisten en la adición de carbohidratos y/o cortes en la estructura de la molécula por enzimas específicas. Estas modificaciones hacen que una proteína inmadura (no funcional) se convierta en una proteína madura (funcional). El proceso de maduración de la proteína ocurre en el RER y en las vesículas que se desprenden para fusionarse con el aparato de Golgi, donde continúa la maduración de la proteína.
Domino traduccional
Centros activos del ribosoma 70S
Domino de salida
Sitio desalida
Siti
oP
Siti
oA
Peptidiltransferasa
SitioRNAm
SitioEF-G
SitioEF-Tu
MembranaRER
Citoplasma
Luz del RER
Poro parala entrada de
preínas
Figura 10. Centros activos de los ribosomas de bacterias 70S

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
60S
40S
UAC
Met
Met
Fen
Ser
Asn
Tre
Met
Fen
SerAsn
Tre
Leu
Gli
Ser
His
Tre
-CGG-GGC-CGG-AUG-ACG-UUU-AACUAA-AAC
Retículo endoplásmico rugoso
RNAm
Ribosomas
RNAt
UAC
Met
U GC
Tre
Cadena polipeptídica
3’5’
N-terminal
C-terminal
Met
Fen
SerAsn
Tre
Leu
Gli
Ser
His
Tre
N-terminal
C-terminal
Met
Fen
SerAsn
Tre
Leu
Gli
Ser
His
Tre
N-terminal
C-terminal
Met
Fen
SerAsn
Tre
Leu
Gli
Ser
His
Tre
N-terminal
C-terminal
Figura 11. Biosíntesis de proteínas en el retículo endoplásmico rugoso
Aparato de Golgi El aparato de Golgi (AG) o complejo de Golgi, recibe este nombre por el biólogo celular italiano Conde Camilo Golgi, que lo describió por primera vez en 1898. Está formado por vesículas membranosas relativamente grandes y aplanadas, tiene una estructura en forma de “C”, no contiene ribosomas (Figura 12). La función del AG es la continuación de la maduración de las proteínas sintetizadas en el RER. El AG tiene dos extremos, el “cis” y el “trans”. Al extremo “cis” llegan las vesículas que se formaron en el RER y se fusionan con la membrana del AG. Las proteínas entran y continúan el proceso de maduración. Las vesículas se forman y fusionan en las distintas membranas del AG, hasta que finalmente se desprenden del AG y las vesículas se fusionan con la membrana celular, expulsando las proteínas maduras al espacio extracelular (exocitosis).

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Aparato de Golgi
Cis
Trans
Vesículas que derivan delretículo endoplásmico rugoso
VesículasVesículas
Vesículasque se fusionarán conla membrna plasmática
Membranacelular
IntracelularExtracelular
Secreción deproteínasmaduras
Vesículas
Figura 12. Estructura del aparato de Golgi, también llamado complejo de Golgi. Participa en la maduración de proteínas.
Retículo endoplásmico liso El retículo endoplásmico liso (REL) está formado por una serie de vesículas fusionadas unas con otras, semejante al RER, sin embargo el REL no contiene ribosomas, lo que le da el aspecto “liso” cuando se observa al microscopio electrónico. El REL es continuación del RER. La función del REL es la síntesis de lípidos, ya que en este organelo se encuentran algunas enzimas que participan en la síntesis de triglicéridos, colesterol o de hormonas esteroides. Participa también en el metabolismo de carbohidratos en el hepatocito ya que la enzima glucosa-6-fosfatasa que convierte la glucosa-P a glucosa se encuentra en el REL. Participa en la degradación de barbituratos o del alcohol ya que en su superficie se localiza el citocromo P450 que participa en el metabolismo de medicamentos. En la célula de Leydig que se encuentra en el testículo, se sintetiza la testosterona. El inicio de la síntesis de testosterona es a partir del colesterol. El colesterol entra a la mitocondria donde es convertido a pregnenolona por la enzima mitocondrial P450scc. La pregnenolona sale de la mitocondria y entra al REL donde las enzimas convierten la pregnenolona a testosterona (Figura 13).
Peroxisomas Son organelos esféricos u ovoides que se autorreplican, en las células de algunas especies, cuando se observan al microscopio electrónico, tienen un centro denso de depósitos cristalinos (Figura 14). Están muy asociados con el retículo endoplásmico. Las funciones mas importantes de los peroxisomas incluyen la beta oxidación de ácidos grasos

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
de cadena muy larga y sustancias relacionadas (proveen acetil-CoA para las reacciones de síntesis) y también para la síntesis de lípidos (especialmente los fosfolípidos del sistema nervioso central, corazón y el músculo, como los plasmalógenos), el colesterol y los ácidos biliares. Muchas reacciones oxígeno-dependientes también ocurren en los peroxisomas para proteger a la célula de los radicales de oxígeno, el H2O2 que se produce es metabolizado por una catalasa.
LH
AMPc
P450sccactivo
P450scc
ATP
Colesterol
Pregnenolona
Célula de Leydig
Mitocondria
AC
RL
H Pregnenolona
17-hidroxipregnenolona17-hidroxiprogesterona
Androstenediona
Testosterona
Testosterona
Dehidroepiandrosterona (DHEA)
Progesterona
2
34
1
6
910
19
5
A
A
A
A
B
B
B
B
O
O
O
O
C O
O
OH
CH3
C
C
D
D
7
8
1112
13
14 15
1617
18
B
C O
CH3
C D
A
O
OH
OH
C
C
D
D
A B
HO
C O
CH3
C D
A B
HO
OH
C O
CH3
C D
3- -HSD II(
3- -HSD II(
3- -HSD II(
17- -HSD(
Retículo endoplásmico liso
17- -Hidroxilasa(
17- -Hidroxilasa( 17- -Hidroxilasa(
17- -Hidroxilasa(
Síntesis de andrógenos en el testículo
Figura 13. Biosíntesis de hormonas esteroides (andrógenos) en el retículo endoplásmico liso de la célula de Leydig. Microtúbulos Los microtúbulos son organelos huecos, semirígidos, cilíndricos que semejan “popotes” al microscopio electrónico. De un diámetro uniforme (25 nm), no son ramificados y de longitud extremadamente variable. Se encuentran en todas las células, pero son especialmente abundantes en las neuronas, plaquetas, leucocitos y en las células que se dividen. Son los principales constituyentes de los flagelos y centriolos. Proveen la fuerza mecánica y forma de la célula como parte del citoesqueleto. Se encargan del transporte intracelular de los organelos (como las mitocondrias o vesículas citoplámicas), del movimiento de los cilios y flagelos y de la citocinesis durante la división celular. No tienen membrana, sus paredes se componen de polímeros lineales (protofilamentos) de la proteína globular llamada tubulina. Los 13 protofilamentos en cada microtúbulo están formados por subunidades alfa y beta alternas que dan lugar a un diseño helicoidal de heterodímeros de tubulina en la pared cilíndrica. Están en constante elongación por polimerización y acortamiento por despolimerización. Los microtúbulos interactúan con otras proteínas asociadas a los microtúbulos, que modulan su estabilidad, ensamblaje y

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
desensamblaje. Dos proteínas motor de los microtúbulos, la kinesina y la dineina se mueven a lo largo del microtúbulo. La kinesina hacia el extremo más y la dineina hacia el extremo menos. Los microtúbulos se originan de un centro organizador de microtúbulos llamado centrosoma (Figura 15).
Membranacelular
Intracelular
Peroxisomas
Extracelular
H O2 2
H O2
Catalasa
H O2 2
H O2
Catalasa
Beta oxidación deácidos grasos de cadena
muy larga
Beta oxidación deácidos grasos de cadena
muy larga
Metabolismo deácidos biliares
Metabolismo deácidos biliares
Metabolismo delácido oxálico
Metabolismo delácido oxálico
Metabolismo deletanol
Metabolismo deletanol
Síntesis de plasmalógenos
Síntesis de plasmalógenos
Figura 14. Los peroxisomas son estructuras esféricas que llevan a cabo el metabolismo de ácidos grasos de cadena muy larga, de ácidos biliares, alcohol, ácido oxálico y otros metabolitos. Filamentos El citoesqueleto de la mayoría de las células está formado por los microtúbulos y dos tipos de filamentos llamados intermedios y filamentos de actina. Estos organelos varían en diámetro, distribución, contenido de proteínas y propiedades mecánicas. Los filamentos intermedios, de 8-12 nm de diámetro, forman grupos ondulantes en una ramificación tridimensional. Están formados por una familia heterogénea de proteínas filamentos intermedios. Proveen principalmente soporte mecánico a la célula, son flexibles pero previenen de un estiramiento excesivo, interactúan con los filamentos de actina y los microtúbulos. Hay seis clases distintas de filamentos intermedios, con 50 genes que los codifican. Diferentes tipos de células expresan tipos específicos de filamentos intermedios. Las láminas nucleares, son las más ampliamente distribuidas, refuerzan la membrana nuclear interna y ayudan en la organización de la arquitectura cromosómica en interfase.

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Otros filamentos intermedios son: queratina, desmina, vimentina, neurofilamentos, filamentos gliales.
Los filamentos de actina, también llamados filamentos delgados o microfilamentos, tienen función de citoesqueleto y de movimiento. Con diámetros de 6 a 8 nm, están formados por la proteína fibrosa, actina. Son flexibles y resisten la deformación y transmiten la fuerza. Contribuyen al movimiento celular e interactúan con los filamentos gruesos (miosina) en las células musculares durante la contracción. Están dispersos en el citoplasma de las células no musculares y forman haces lineales. Se encuentran en las microvellosidades o justo entre la membrana plasmática, determinan la forma de la superficie celular y contribuyen a la locomoción celular, la citocinesis y la fagocitosis.
Centrosoma
Microtúbulos
Figura 15. Los microtúbulos se originan del centrosoma
Centrosoma y centriolos El centrosoma es el principal centro organizador de microtúbulos y es el sitio para la formación de nuevos microtúbulos y las fibras del huso mitótico. Generalmente está cercano al núcleo y rodeado del aparato de Golgi. El centrosoma esta formado por un par de centriolos (el diplosoma), que se encuentran orientados en ángulo recto u oblicuo uno a otro. Cada centriolo es un cilindro corto de 200 nm de diámetro y 500 a 700 nm de longitud (Figura 16). Cada anillo consiste de nueve tripletes fusionados de microtúbulos. En muchas células los microtúbulos irradian del centrosoma en forma de estrella y contribuyen para dar forma a la célula. Durante la mitosis los centriolos se separan y migran a polos opuestos de la célula y es el foco para la síntesis de microtúbulos necesarios para el movimiento de los cromosomas.

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Núcleo Nucleolo
Centrosoma
Centriolo Centriolo
Centrosoma
Figura 16. El centrosoma está formado por un par de centriolos, es el sitio generador de microtúbulos
Núcleo Es la estructura más grande en la célula que contiene el material genético. La forma
del núcleo es variable, puede ser esférico, ovoide o lobulado (como en los polimorfonucleares). La mayoría de las células tienen un núcleo, sin embargo, algunas células pueden ser binucleadas (hepatocito), otras multinucleadas (como el osteoclasto o las fibras musculares). El núcleo esta formado por el nucleolo, la cromatina, la matriz nuclear y la cubierta nuclear (Figura 17).
El nucleolo es una área densa, ovoide (no mayor de 1mm de diámetro) no rodeada de membrana (Figura 18). Su tamaño y número depende de la actividad funcional de la célula. El nucleolo es el sitio de transcripción del RNAr y producción de las subunidades de los ribosomas. El nucleolo contiene grandes cantidades de RNA motivo por el cual es intensamente basofílico y se tiñe con el colorante hematoxilina. El nucleolo muestra dos áreas, la parte granulosa (región periférica nucleolar) y la parte fibrosa (región central del nucleolo). La parte granulosa es el sitio de ensamblaje de RNAr y proteínas ribosomales para formar las subunidades del ribosoma, una vez formadas, salen del núcleo a través de los poros nucleares. La parte fibrosa consiste en filamentos finos, genes de RNAr y factores de transcripción.
La cromatina, esta formada por DNA, proteínas histonas (principalmente), proteínas no histonas y RNA. La cromatina tiene gran afinidad por el colorante básico hematoxilina. La cromatina nuclear existe en dos formas: la eucromatina y la heterocromatina. La eucromatina, que se tiñe más pálido que la heterocromatina, es transcripcionalmente activa y es prominente en las células que sintetizan proteínas. La heterocromatina es transcripcionalmente inactiva

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Núcleo NucleoloPoro nuclear
Membrana nuclearexterna Membrana nuclear
interna
Espacioperinuclear
Ribosoma
CromatinaEucromatina
Heterocromatina
Figura 17. Estructura del núcleo
La matriz nuclear es el área semejante a una esponja, situada entre la cromatina y el nucleolo, es rica en proteínas no histonas, también contiene filamentos intermedios como las láminas nucleares, la mayoría de las cuales se adhieren a la parte interna de la membrana nuclear interna.
La cubierta nuclear, encierra y separa el contenido nuclear del citoplasma en las células en interfase. Esta formada por dos membranas, separadas por un espacio estrecho (10-70 nm) llamado espacio perinuclear (cisterna). La membrana nuclear externa tiene ribosomas y es continuación del RER, el espacio perinuclear es continuo con la luz del RER. La membrana nuclear interna no tiene ribosomas y está en contacto con la heterocromatina.
El complejo poro nuclear
La membrana nuclear externa e interna se unen en algunos sitios formando una estructura llamada, complejo poro nuclear. En el centro de cada complejo hay un poro que provee de un canal hidrosoluble entre el núcleo y el citoplasma (Figura 19). Hay aproximadamente 3,000 complejos poro nuclear por núcleo en las células de mamíferos. Las proteínas que necesitan pasar del citoplasma al núcleo entran a través del complejo poro nuclear. Todos los RNA (RNAm, RNAt, RNAr y otros RNA pequeños) son sintetizados en el núcleo y pasan al citoplasma a través del complejo poro nuclear. Los

Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
poros nucleares son utilizados para importar y exportar moléculas. El complejo visto al microscopio electrónico parece una estructura en roseta, de ocho unidades simétricas y una central.
Núcleo Nucleolo
Nucleolo
Célula
Parte granulosaFormación de las subunidades
del ribosoma (40S y 60S)
Subunidad 60S Subunidad 40S
Parte fibrosaGenes RNAr, Filamentos finosFactores de transcripción
40S
40S
40S
40S
40S
40S
60S
60S
60S
60S
60S
60S
Figura 18. Estructura del nucleolo
Poro nuclear
Núcleo
Célula
Figura 19. El poro nuclear La capacidad de una molécula para pasar libremente por el poro nuclear depende de su tamaño. Las moléculas <5 kD pasan libremente el poro nuclear, motivo por el cual se ha concluido que los iones, nucleótidos y otra moléculas pequeñas son permeables y entran libremente a través del poro. Las proteínas entre 5-50 kD difunden en una proporción inversamente proporcional a su tamaño, por lo que se ha concluido que las proteínas pequeñas pueden entrar al núcleo por difusión pasiva (pero también pudieran entrar por transporte activo). Las proteínas >50 kD no entran al núcleo por difusión pasiva, requieren de un mecanismo de transporte activo para que entren al núcleo. Un ejemplo de una proteína grande que necesita entrar a través del poro nuclear es la proteína wernerina (WRNp) que se encuentra mutada en los pacientes con un tipo de envejecimiento prematuro llamado síndrome Werner (Figura 20). El gen WRN codifica para la proteína de 1432 aminoácidos denominada WRNp. En el extremo C-terminal de la WRNp se encuentra

una región denominada NLS o señal de localización nuclear, que es necesaria para que la proteína sea importada al núcleo. La proteína WRNp en el citoplasma se tiene que unir a un grupo de proteínas denominadas importinas. El complejo importinas-WRNp entra al núcleo a través del poro nuclear. Dentro del núcleo, la proteína se localiza en el nucléolo. La función normal de la WRNp no se conoce. Podría funcionar para desenrollar el DNA durante la replicación, reparación, transcripción o cualquier otra función del DNA que se requiera para desenrollar la cadena. La WRNp interactúa con otras proteínas involucradas en el metabolismo del DNA, incluyendo proteínas relacionadas con el metabolismo del telómero. La WRNp en conjunto con la proteína ligadora de telómero (TRF2) y la proteína de replicación A (RPA) podrían facilitar el desenrollamiento de segmentos largos teloméricos.
Gen WRN 8p12-p11.2
Efectos metabólicos
Membrana nuclearinterna
Membrana nuclearexterna
Cisterna
ARNm WRN
Proteína WRNp1432 aa
Sistema de importación
nuclear
WRNp
La WRNp a través del dominioNLS se une a las importinas
y es importada al núcleo
RAN GTP
RAN GTP
Importina 0
0
Importina 0
00
0
0
0
0
0
5’ 5’
3’3’RNasa
Helicasa
NLS
WRNp
WRNp
DNA
DNA Helicasa (WRNp)Sistema de reparación del DNAIniciación de la replicación del DNAMetabolismo del DNA que implique la helicasa
HRDC
0
0
Poronuclear
Figura 20. La proteína WRNp tiene varios dominios que son importantes para el funcionamiento adecuado. La WRNp se localiza en el núcleo y más específicamente se ha encontrado en el nucléolo. Uno de los dominios es denominado NLS “señal de localización nuclear” es necesario para que la WRNp pueda ser importada al núcleo por las proteínas importadoras, probablemente las importinas, a través del poro nuclear.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.

Bibliografía 1. Baird DM, Davis T, Rowson J, Jones CJ, Kipling J, Kipling D. Normal telomere erosion rates at the single cell level in Werner syndrome fibroblast cells. Hum Mol Genetics 2004;13:1515-24. 2. Colella S, Nardo T, Botta E, Lehmann AR, Stefanini M. Identical mutations in the CSB gene associated with either Cockayne syndrome or the DeSanctis-cacchione variant of xeroderma pigmentosum. Hum Mol Genet 2000;9:1171-5. 3. Colella S, Nardo T, Mallery D, Borrone C, Ricci R, Ruffa G, Lehmann AR, Stefanini M . Alterations in the CSB gene in three Italian patients with the severe form of Cockayne syndrome (CS) but without clinical photosensitivity. Hum Mol Genet 1999;8:935-1. 4. Gebhart E, Bauer R, Raub U, Schinzel M, Ruprecht KW, Jonas JB. Spontaneous and induced chromosomal instability in Werner syndrome. Hum Genet 1998;80: 135-9. 5. Gray MD, Shen J-C, Kamath-Loeb AS, Blank A, Sopher BL, Martin GM, Oshima J, Loeb LA. The Werner syndrome protein is a DNA helicase. Nature Genet 1997;17: 100-3. 6. Huang S, Li B, Gray MD, Oshima J, Mian IS, Campisi J. The premature ageing syndrome protein, WRN, is a 3-prime-5-prime exonuclease. Nature Genet 1998;20: 114-5. 7. Itoh M, Hayashi M, Shioda K, Minagawa M, Isa F, Tamagawa K, Morimatsu Y, Oda M. Neurodegeneration in hereditary nucleotide repair disorders. Brain Dev 1999;21:326-3. 8. Laat W, Jaspers N, Hoeijmakers. Molecular mechanism of nucleotide excision repair. Gen Develop 1999;13:768-85. 9. Lewin B (Ed.). Genes VII. Oxford University Press. Oxford, New York, Tokyo, 2000. 10. McKee T, McKee JR (Eds.). Bioquímica. La base molecular de la vida. McGraw-Hill. 3ª Ed. 2003. 11. Moser MJ, Bigbee WL, Grant SG, Emond MJ, Langlois RG, Jensen RH, Oshima J, Monnat RJ. Genetic instability and hematologic disease risk in Werner syndrome patients and heterozygotes. Cancer Research 2000;60:249-96. 12. Omary MB, Coulombe PA, McLean I. Intermediate filament proteins and their associated diseases. N Engl J Med 2004;351: 2087-00. 13. Ovalle WK, Nairney PC. Netter’s essential histology. Suanders Elsevier, 2008. 14. Schellenberg GD, Miki T, Yu C-E, Nakura J. Werner syndrome. In: Scriver CR, Beaudet AL, Sly WS. Valle D. Eds. The Metabolic and Molecular Bases of Inherited Disease. 8th ed. Mc Graw-Hill, pp. 785-797, 2001. 15. Wood R, Mitchell M, Sgouros J, Lindah T. Human DNA repair genes. Science 2001;291:1284-9. 16. Wyllie FS, Jones CJ, Skinner JW, Haughton MF, Wallis C, Wynford-Thomas D, Faragher RGA, Kipling D. Telomerase prevents the accelerated cell ageing of Werner syndrome fibroblast. Nature Genetics 2000;24: 16-7. 17. Zschocke J, Hoffmann GF. Vademecum metabolicum. Manual of metabolic paediatrics. Milupa. Schattauer. Stutgart, New York, 1999.
Ornelas-Arana ML. et al. Célula, Estructura y Función. Citogenética Básica.
Laboratorio de Bioquímica. Departamento de Biología Molecular y Genómica, CUCS, Universidad de Guadalajara.Servicio de Genética. Hospital Civil de Guadalajara “Fray Antonio Alcalde”. Guadalajara, Jalisco, México. Septiembre 2009.