
CENTRO DE DESARROLLO DE PRODUCTOS BIÓTICOStesis.ipn.mx/jspui/bitstream/123456789/3580/1/...a marzo...
85
INSTITUTO POLITÉCNICO NACIONAL CENTRO DE DESARROLLO DE PRODUCTOS BIÓTICOS DEPARTAMENTO DE BIOTECNOLOGÍA EFECTO DE LA ÉPOCA DE TRANSPLANTE SOBRE LA ACUMULACIÓN DE LUTEÍNA EN INFLORESCENCIAS DE CEMPAXÚCHIL (Tagetes erecta L.) P R E S E N T A: IBQ. PAUL MAURICIO SÁNCHEZ OCAMPO DIRECTORES DE TESIS DRA. MARTHA LUCÍA ARENAS OCAMPO DRA. MARÍA EUGENIA JARAMILLO FLORES Yautepec, Morelos, Diciembre de 2008 T E S I S QUE PARA OBTENER EL GRADO DE M A E S T R Í A E N C I E N C I A S EN DESARROLLO DE PRODUCTOS BIÓTICOS
Transcript of CENTRO DE DESARROLLO DE PRODUCTOS BIÓTICOStesis.ipn.mx/jspui/bitstream/123456789/3580/1/...a marzo...

INSTITUTO POLITÉCNICO NACIONAL
CENTRO DE DESARROLLO DE PRODUCTOS BIÓTICOS
DEPARTAMENTO DE BIOTECNOLOGÍA
EFECTO DE LA ÉPOCA DE TRANSPLANTE SOBRE LA ACUMULACIÓN DE LUTEÍNA EN INFLORESCENCIAS DE CEMPAXÚCHIL (Tagetes erecta L.)
P R E S E N T A:
IBQ. PAUL MAURICIO SÁNCHEZ OCAMPO
DIRECTORES DE TESIS
DRA. MARTHA LUCÍA ARENAS OCAMPO
DRA. MARÍA EUGENIA JARAMILLO FLORES
Yautepec, Morelos, Diciembre de 2008
T E S I SQUE PARA OBTENER EL GRADO DE
M A E S T R Í A E N C I E N C I A S EN DESARROLLO DE PRODUCTOS BIÓTICOS




El presente trabajo se llevó a cabo en el Departamento de
Biotecnología y campo experimental del Centro de
Desarrollo de Productos Bióticos del Instituto Politécnico
Nacional, bajo la dirección de la Dra. Martha Lucia Arenas
Ocampo. Así también, parte de este trabajo se desarrollo en
otras instituciones: Escuela Nacional de Ciencias
Biológicas de la Ciudad de México, D. F. con la
codirección de la Dra. María Eugenia Jaramillo Flores en
el Laboratorio de Enzimas del Departamento de Alimentos.
Para la realización de los estudios de Maestría, se contó
con el apoyo económico de las becas: CONACYT
(B061663), Programa Institucional de Formación de
Investigadores (PIFI) y Beca-Tesis IPN Institucional. La
investigación fue realizada con el financiamiento
económico de los proyectos COFAA, SIP (20080986), SIP
(20080336), CONACYT 61034.

DEDICATORIA
A MIS PADRES
Por su cariño, apoyo incondicional en la toma de mis decisiones
importantes en mi vida y por alentarme en los tiempos difíciles.
A MIS HERMANOS
Por creer en mí y por apoyarme durante todos mis estudios, gracias
por ayudarme a encontrar mí camino.

AGRADECIMIENTOS
Al comité tutorial y revisor:, Dra. María Eugenia Jaramillo Flores, Dr. Adrián Guillermo
Quintero Gutiérrez, Dra. Silvia Evangelista Lozano, Dr. Pablo E. Vanegas Espinoza y a la
Dra. Alma A. Del Villar Martínez por sus apreciables comentarios y correcciones que
permitieron enriquecer el trabajo.
A la Dra. Martha L. Arenas Ocampo, por su accesoria y dirección en la realización de este
trabajo de tesis, gracias por su paciencia, orientación y amistad.
Al Dr. Antonio Jiménez Aparicio, por apoyarme, en la realización de este trabajo y por su
amistad.
A la Dra. Silvia Evangelista Lozano, por darme las facilidades para llevar a cabo los
cultivos en campo.
A la Dra. María Eugenia Jaramillo Flores, por su asesoría, colaboración y apoyo para la
realización de este trabajo.
Al Dr. Miguel Ángel Serrato Cruz, por su asesoría y orientación en la realización de este
proyecto de tesis.
A Jacqueline Capataz Tafur, por su cariño, apoyo incondicional y por enseñarme a su
fortaleza ante los problemas de la vida.
A mis compañeros de la maestría y amigos por impulsarme a seguir adelante siempre,
gracias Dante, Mario Domínguez, Juan Torruco, Lino, Pamela, Vero Pérez, Nayeli, Galia,
Salvador y Margarita, Ximena, Norma Leticia, Rosa Elena, Denisse Guerra, mine, Susana,
Juan Carlos, Luz Behaney, María Luisa, y Rita por apoyarme siempre.

CONTENIDO
ÍNDICE DE FIGURAS………………………………………………………………... IÍNDICE DE CUADROS………………………………………………………………. IISIMBOLOS Y ABREVIATURAS……………………………………………………. IIIRESUMEN……………………………………………………………………………... IVABSTRACT……………………………………………………………………………. V1. INTRODUCCIÓN……………………………………………………………….. 12. ANTECEDENTES……………………………………………………………….. 3
2.1. Efecto de las condiciones ambientales en el crecimiento y desarrollo de las plantas……………………………………………………………………….
3
2.1.1. Influencia de la temperatura sobre las plantas……………………… 4 2.1.2. Efecto de la humedad relativa en las plantas……………………….. 5 2.1.3. Efecto de la luz en las plantas………………………………………. 6 2.1.4. Efecto del fotoperiodo en las plantas……………………………….. 7 2.2. Factores que inducen la floración…………………………………………... 7 2.2.1. Influencia de la Temperatura y fotoperiodo sobre la floración…….. 8 2.3. Producción de metabolitos secundarios en las plantas……………………… 9 2.3.1. Presencia de metabolitos secundarios en las plantas………………. 9 2.4. Cempaxúchil (Tagetes erecta L.) como modelo de estudio………………… 10 2.4.1. Clasificación botánica de Tagetes erecta L………………………… 11 2.4.2. Clasificación taxonómica…………………………………………... 12 2.4.3. Distribución geográfica…………………………………………….. 12 2.4.4. Producción de T. erecta en México………………………………… 13 2.4.5. Componentes bioactivos de Tagetes spp…………………………. 15 2.5. Carotenoides………………………………………………………………… 15 2.5.1. Biosíntesis de los carotenoides……………………………………... 21 2.5.2. Función de los carotenoides………………………………………... 24 2.5.2.1. Función fotoprotectora de los carotenoides……………… 24 2.5.2.2. Ciclo de las xantófilas……………………………………. 25 2.5.3. Factores que influyen en la acumulación de los carotenoides……… 27 2.5.4. Importancia económica del cempaxúchil y los carotenoides………. 28
3. JUSTIFICACIÓN………………………………………………………………... 294. HIPÓTESIS………………………………………………………………………. 305. OBJETIVOS……………………………………………………………………... 31 5.1. Objetivo general…………………………………………………………….. 31 5.2. Objetivos particulares……………………………………………………….. 31

6. MATERIALES Y MÉTODOS………………………………………………….. 32 6.1. Obtención de plántulas……………………………………………………… 32 6.1.1. Germinación de las semillas………………………………………... 33 6.1.1.1. Desinfección y germinación de semilla………………...... 33 6.1.2. Aclimatación de las plantas………………………………………… 33 6.1.2.1. Preparación de sustrato…………………………………... 33 6.1.2.2. Transplante a maceta…………………………………….. 34 6.1.3. Transplante a campo y medición de las condiciones ambientales…. 34 6.2. Evaluación del desarrollo vegetativo……………………………………….. 34 6.2.1. Altura de las plantas……………………………………………….. 34 6.2.2. Conteo del número de primordios florales……………………….... 34 6.3. Colecta de las inflorescencias………………………………………………. 35 6.4. Determinación del índice de color………………………………………….. 35
6.5. Evaluación del contenido de xantófilas totales por espectrofotometría UV-Visible……………………………………………………………………….
36
6.5.1. Extracción de carotenoides…………………………………………. 36
6.5.2. Cuantificación de xantofilas totales por espectrofotometría UV-Visible……………………………………………………………….
36
6.6. Identificación y cuantificación de carotenoides por HPLC…………........... 377. RESULTADOS Y DISCUSIÓN………………………………………………… 38
7.1. Monitoreo de las condiciones ambientales durante el periodo de los cultivos………………………………………………………………………
38
7.2. Evaluación del desarrollo vegetativo de los cultivos……………………….. 40 7.2.1. Evaluación de la altura de las plantas………………………………. 40 7.2.2. Evaluación del número de primordios florales……………………... 40 7.2.3. Evaluación del diámetro y peso fresco de las inflorescencias……… 41
7.3. Evaluación del índice de color de las inflorescencias de cempaxúchil por CIELAB……………………………………………………………………..
44
7.4. Evaluación del contenido de xantófilas totales por espectrofotometría UV-Visible……………………………………………………………………….
44
7.5. Identificación y cuantificación de luteína por HPLC……………………….. 46 7.6. Rendimientos de la producción de inflorescencias de cempaxúchil………... 49
8. CONCLUSIONES……………………………………………………………….. 519. PERSPECTIVAS………………………………………………………………… 5210. BIBLIOGRAFÍA………………………………………………………………… 5311. ANEXOS………………………………………………………………………….. 66

I
ÍNDICE DE FIGURAS
Pág.Figura 1. Distribución secundaria de las formas domesticas de cempaxúchil en
México. 12
Figura 2. Estructuras de los principales carotenoides acíclicos en orden ascendente de acuerdo al número de dobles enlaces (n).
17
Figura 3. Estructuras de los principales carotenoides cíclicos, los cuales poseen el anillo en uno o ambos extremos de la cadena hidrocarbonada.
18
Figura 4. Estructuras de carotenoides que en su esqueleto tienen grupos hidroxilo. 19Figura 5. Estructuras de carotenoides que en su esqueleto contienen grupos
epóxido. 20
Figura 6. Estructuras de apocarotenoides los cuales poseen en su esqueleto menos de 40 átomos de carbono.
21
Figura 7. Ruta de biosíntesis de los carotenoides. 23Figura 8. Ciclo de las xantófilas y sus efectos sobre la captura de luz y la
disipación de la energía dentro del aparato fotosintético. 26
Figura 9. Variación del peso fresco y peso seco del las flores de diferentes épocas de transplante a campo.
43
Figura 10. Variación de la acumulación de xantófilas totales para los diferentes épocas de transplante.
45
Figura 11. Perfil cromatográfico del HPLC del estándar de luteína. 46Figura 12. Perfil cromatográfico del HPLC obtenido de los extractos de las flores
de cempaxúchil del cultivo del mes de Julio. 47
Figura 13. Variación de la acumulación de luteína para las diferentes épocas de transplante.
48

II
ÍNDICE DE CUADROS
Pág.Cuadro 1. Producción de cempaxúchil a nivel nacional. 14Cuadro 2. Producción de cempaxúchil en el estado de Morelos. 14Cuadro 3. Variación de las condiciones ambientales durante los meses (julio 2007
a marzo 2008) del experimento en campo, desde el inicio del primer cultivo hasta la última colecta de inflorescencias.
39
Cuadro 4. Evaluación del desarrollo de las plantas a 12 días después del inicio de la floración de diferentes épocas de transplante a campo.
42
Cuadro 5. Valores del IC* de las inflorescencias de cempaxúchil de diferentes épocas de transplante a campo.
44
Cuadro 6. Rendimientos en la producción de inflorescencias de cempaxúchil de seis diferentes épocas de transplante a campo.
50

III
SÍMBOLOS Y ABREVIATURAS
% Porcentaje ° C Grado Centígrado ABA Ácido Absícico DOXP 1-deoxixilulosa-5-fosfato g Gramos GGPP Geranilgeranil difosfato GGPS Geranil difosfato sintasa h Hora HEAT Hexano: Etanol: Acetona: Tolueno HPLC Cromatografía Líquida de Alta Resolución HR (%) Porcentaje de Humedad Relativa IPP Isopentenil Pirofosfato KOH Hidróxido de Potasio LCY-β Licopeno-β-Ciclasa M Molar M Metros Ml Milílitro MVA Ácido Mevalónico Na2SO4 Sulfato de Sodio nm Nanómetro P/V Peso/Volumen PF Peso Fresco PS Peso Seco PSY Fitoeno Sintasa RFA Radiación Fotosintéticamente Activa UV/Vis Ultra Violeta / Visible V/V Volumen/Volumen VDE Violaxantina desepoxidasa VE Violaxantina epoxidasa μg Microgramo μl Microlitro

IV
RESUMEN
El cempaxúchil (Tagetes erecta L.) es una planta herbácea anual comercialmente cultivada para
la producción de carotenoides, cuyo cultivo está asociado a los meses de julio a octubre, durante
los cuales se presentan condiciones ambientales muy particulares. No obstante, también puede ser
cultivada durante cualquier otro periodo del año, bajo condiciones diferentes, que pueden jugar
un papel muy importante en el desarrollo vegetativo de la planta y la biosíntesis de los pigmentos
(carotenoides) en las inflorescencias. En este trabajo se evaluó el efecto de la época de
transplante a campo sobre el desarrollo de la planta y acumulación de luteína en inflorescencias
de T. erecta. Los experimentos fueron realizados en el campo experimental Emiliano Zapata del
Centro de Desarrollo de Productos Bióticos (Yautepec, Morelos). Para ello, plántulas de T.
erecta de porte bajo de 30 días de edad fueron transplantadas a campo entre los meses de julio a
diciembre de 2007. Los parámetros evaluados fueron: la altura de las plantas, el número de
primordios florales, el diámetro, los pesos fresco y seco e índice de color de las inflorescencias,
así como la acumulación e identificación de luteína en lígulas de cempaxúchil. Los resultados
obtenidos mostraron que las diferentes épocas de transplante presentaron diferencias
significativas en cuanto a la altura, número de primordios florales, diámetro, pesos fresco y seco
de las inflorescencias. Las plantas transplantadas en los meses de noviembre y diciembre
recibieron la menor radiación solar, temperatura, humedad relativa baja y fotoperiodo corto, lo
cual condujo a un mayor número de primordios florales (43±8.6 y 50.6±11.5 primordios/planta,
respectivamente), mayor acumulación de xantofilas totales (13.6 y 8.2 g/kg respectivamente) y
con un índice de color que ubicó a las flores de noviembre en el amarillo intenso, corroborando
que este cultivo en particular presentó la mayor acumulación de xantofilas (luteína). Estos
resultados sugieren que la época de transplante, en la cual se presentaron alta irradiación y mayor
tiempo de exposición, disminuyó el contenido de luteína, posiblemente debido a que son
moléculas que sufren transformaciones fotoquímicas.

V
ABSTRACT
Marigold (Tagetes erecta L.) is an annual herbaceous plant commercially grown to produce
carotenoids, crop which is associated with the months of july to october, during which
environmental conditions are very particular. However, it can also be cultivated during any other
period last year, under different conditions, which may play an important role in the development
of the vegetative plant and the biosynthesis of the pigments (carotenoids) in inflorescence. In this
study, was evaluated the effect of the time of transplantation to field on the development of the
plant and accumulation of lutein in inflorescences of T. erecta. The experiments were conducted
in the experimental field Emiliano Zapata´s Product Development Center Biotic (Yautepec,
Morelos). For that, dwarf seedlings´s marigold of 30 days of age was transplanted into the field
between the months of july to december 2007. Parameters were evaluated: plant height, the
number of floral primordia, diameter, fresh and dry weights and index of color of the
inflorescence, as well as the accumulation of lutein in ligules of marigold. The results showed
that different sowing dates were significantly different in terms of height, number of floral
primordia, diameter, fresh and dry weight of the inflorescence. The plants of the months of
november and december received the lower solar radiation, temperature, low relative humidity
and short photoperiod, which led to a greater number of floral primordia (43 ± 8.6 and 50.6 ±
11.5 primordia / plant, respectively), higher accumulation of total xanthophylls (13.6 and 8.2 g /
kg respectively) and the color index of november flowers was like intense yellow, corroborating
that this crop in particular had the highest accumulation of lutein. These results suggest that the
time of transplantation, which showed high radiation and increased exposure time, decreased the
content of lutein, possibly because they are molecules that are influenced by changes
photochemical.

1
1. INTRODUCCIÓN
Cempoaltxóchilt (Tagetes erecta L.) es un vocablo de origen náhuatl utilizado en la época
prehispánica para referirse de forma genérica a un grupo de plantas con características comunes:
flores vistosas por su forma y tamaño, diversidad aromática y colores llamativos como el
amarillo y del anaranjado al rojo. Los usos actuales y potenciales de esta planta por su
composición son numerosos, entre otros se mencionan los siguientes: para la obtención de
compuestos bioactivos antioxidantes, pigmentos utilizados en la industria alimentaria,
saborizantes y resinas; además de su utilización como ornamental (flor de corte, jardín y
maceta), insecticida, nematicida, larvicida, atrayente o repelente de insectos, abono verde y para
el control de malezas. En México, adicionalmente es relevante su empleo en la medicina
tradicional y en algunas festividades religiosas.
Cempoalxóchitl también conocida como cempaxúchil, productora importante de
carotenoides, es una planta que por la amplia diversidad de variedades presentes en nuestro país,
ha generado la iniciativa de proponer a México como su centro de origen (Serrato-Cruz, 2004).
Esta es una planta herbácea anual, cuyo cultivo está asociado a los meses de julio a octubre,
durante los cuales se presentan condiciones ambientales muy particulares, no obstante, también
puede ser cultivada durante cualquier otro periodo del año, bajo condiciones diferentes, que
pueden jugar un papel muy importante en el desarrollo vegetativo de la planta y la biosíntesis de
los pigmentos (carotenoides), en las inflorescencias.
En la naturaleza el ciclo, de vida de las plantas está sincronizado con los cambios
estacionales, los cuales afectan la transición en su desarrollo y la floración, entre otros aspectos.
Las fluctuaciones en la humedad y la temperatura proporcionan información que utilizan las
plantas para la sincronización, con las estaciones, de muchas de las respuestas fisiológicas
relacionadas con el desarrollo (Searle y Coupland, 2004). En este sentido, Dorais y col. (2001)
reportaron que la temperatura y la intensidad de la luz ejercen una influencia directa sobre los
atributos en la calidad de los frutos de tomate, como son la apariencia, la firmeza, textura, materia
seca y las propiedades sensoriales debido a que la variación en estas condiciones ambientales los
somete a varios tipos de estrés.

2
En la actualidad, un entendimiento detallado de la influencia de los factores ambientales y su
interacción con las prácticas agronómicas, que afectan la acumulación de metabolitos secundarios
durante el periodo de floración es aún desconocido, aunque en algunos casos, se ha observado el
efecto favorable de la exposición a la luz sobre la acumulación de la vitamina C en cultivos de
jitomate (Dumas y col., 2003).
En este trabajo se realizó una descripción mediante la observación del efecto de las
condiciones ambientales sobre la altura de las plantas, numero de botones florales, diámetro y
acumulación de luteína en inflorescencias de cempaxúchil en cultivos transplantados en
diferentes fechas.

3
2. ANTECEDENTES
2.1. Efecto de las condiciones ambientales en el crecimiento y desarrollo de las plantas.
Para entender qué factores son los que afectan el crecimiento y desarrollo en primera
instancia se tratará de definir el crecimiento, entendido como el aumento de tamaño. A medida
que crecen los organismos multicelulares a partir del cigoto, no solo aumentan de volumen, sino
también en peso, número de células, cantidad de protoplasma y complejidad.
El término desarrollo se refiere a un conjunto de procesos que comienzan con una célula
única, el cigoto, que crece y se desarrolla hasta llegar a ser un organismo multicelular. En este
proceso se da una síntesis continua de moléculas grandes y complejas a partir de iones y
moléculas más pequeñas, que son la materia prima del crecimiento. A través de la división celular
se generan células nuevas, muchas de las cuales llegan a ser más grandes y complejas. A este
proceso se le conoce como crecimiento y diferenciación de células para formar tejidos, órganos y
organismos. Otro término adecuando para este proceso es el de morfogénesis (del griego morfe,
forma, y génesis, origen) (Salisbury y Ross, 1992).
La medición del crecimiento tiene importancia en el estudio de los factores que afectan el
desarrollo de las plantas y se puede realizar midiendo una de las características de crecimiento
recién mencionadas, sin embargo, dos mediciones en particular pueden resultar de utilidad: el
incremento en el volumen (tamaño) puede ser estimada en forma aproximada, mediante la
determinación de la expansión en una o dos direcciones, como longitud (altura del tallo),
diámetro (de una rama) o área (de una hoja).
Hay muchos factores que afectan el crecimiento y desarrollo de las plantas en un ambiente
dado, definiendo éste como las circunstancias, objetos o condiciones que las rodean como pueden
ser: el complejo de factores climáticos, edáficos (del suelo) y bióticos que actúan sobre un
organismo o una comunidad ecológica y que finalmente determinan su forma y sobrevivencia.
Las condiciones ambientales durante el establecimiento de las plantas determinan su crecimiento

4
y desarrollo, debido a los efectos sobre la estructura y composición de la planta (Putnam y col.,
1993).
Por otro lado, Spomer (1973), definió el ambiente operacional aplicando conceptos de la
termodinámica, hizo notar que un factor ambiental interactúa con un organismo, solo cuando el
factor calienta o efectúa trabajo sobre el organismo o el organismo calienta o efectúa trabajo
sobre dicho factor. Si las ondas de radio pasan inalteradas a través de la planta, entonces la planta
también es inalterada y podría no haber interacción; entonces las ondas de radio no serían parte
del ambiente operacional de la planta. Los factores ambientales que se ajustan a la definición de
factores operacionales de Spomer incluyen la luz, calor, agua, potenciales eléctricos, diversos
gases, compuestos minerales y sustancias orgánicas.
Aunque Spomer no menciona a la fotoperiodicidad, esta también forma parte de los factores
ambientales operacionales de las plantas ya que si tienen efecto en su crecimiento y desarrollo, tal
descubrimiento fue hecho por Raven y Curtis (1975) con una variedad de soja (Glycine max)
denominada Bixoli.
Los estímulos ambientales incluyen a parámetros físicos como la luz, temperatura y la
gravedad, los cuales ejercen el mayor impacto en las plantas. Otros parámetros como el campo
magnético, el sonido y el viento (el cual es un estímulo físico) pueden tener efectos más sutiles,
aunque éstos pueden ser más difíciles de establecer experimentalmente. Otros factores
ambientales como la humedad del suelo, la humedad relativa y la nutrición pueden también
influenciar el desarrollo de las plantas en algunos casos. Recientemente se ha aportado evidencia
de que el aire y agua con algún agente contaminante pueden modificar significativamente los
patrones de desarrollo. De acuerdo con lo anterior, el ambiente es demasiado complejo para
describirlo, pero si nos limitamos a factores que son parte del ambiente operacional de la plantas,
la tarea puede ser menos compleja (Hopkins, 1999).
2.1.1. Influencia de la temperatura sobre las plantas.
Todos los organismos intercambian calor con su entorno en la dirección del gradiente
térmico. Por otro lado la variedad de regímenes térmicos sobre la Tierra, implica que unas plantas
deberán especializarse en disipar calor y otras en limitar las pérdidas. Las plantas han adaptado

5
sus ciclos vitales a los ciclos térmicos, es decir a la variabilidad cíclica de la temperatura; sin
embargo la aparición de valores extremos puede afectarlas. El intervalo de temperaturas
encontrado en la superficie de la tierra es favorable para que se lleven a cabo los procesos
biológicos que mantienen la vida. De esta manera, las especies y organismos manifiestan
crecimiento y sobrevivencia en algunas temperaturas cardinales (mínima, óptima y máxima) que
afectan las reacciones enzimáticas individuales o múltiples que catalizan las reacciones
metabólicas.
La temperatura en los climas tropicales es relativamente estable, las plantas crecen en
regiones moderadas (4 y 45° C) y las más cercanas a los polos están sujetas a variaciones en la
temperatura, predecibles diaria y estacionalmente. En este sentido las plantas tienen rutas para
incorporar esta información en su desarrollo y estrategias de sobrevivencia, además usan esta
información para asegurar la latencia de los brotes, tubérculos, y semillas, promoviendo
modificaciones en el comportamiento de la floración, lo cual parece ser clave para su
supervivencia en periodos de tiempo no favorables para un crecimiento y desarrollo normal
(Salisbury y Ross, 1992).
2.1.2. Efecto de la humedad relativa en las plantas.
La materia viva depende del agua, el funcionamiento bioquímico y fisiológico de todos los
organismos ocurre dentro del agua contenida en sus órganos, tejidos y células. Los organismos
terrestres viven en el aire, cuya concentración de agua es menor a la de los organismos acuáticos,
por ello pierden agua por transpiración y otros procesos.
El agua puede limitar el crecimiento y la productividad de un cultivo en cualquier sitio, ya
sea por periodos secos inesperados o por lluvia normalmente baja, lo cual hace necesario el
riesgo constante. La humedad relativa, como uno de los determinantes de la tasa de pérdida de
agua por evapotranspiración, es una condición ambiental importante para las plantas terrestres. El
estrés por deficiencia de agua es una amenaza persistente para la sobrevivencia de las plantas,
obligándolas a desarrollar modificaciones morfológicas y fisiológicas que les permita sobrevivir
en regiones con lluvia inadecuada y suelos con bajo contenido de humedad (Hopkins, 1999).

6
Los efectos de la humedad relativa resultan a menudo difíciles de separar de los de la
temperatura. Esto se debe simplemente a que un aumento de la temperatura conduce a un
incremento de la tasa de evaporación y a una disminución de la humedad de la planta. Cuanto
más elevada es la humedad relativa menores serán las pérdidas por evapotranspiración. Abdul y
col. (2007) realizaron un experimento en donde encontraron que una deficiencia de agua afecta el
crecimiento inicial de la planta, acumulación de materia seca y variación de pigmentos
fotosintéticos en dos variedades de Catharanthus roseus, esto es debido a que el estrés hídrico se
caracteriza por una reducción en el contenido de agua de las células, del número de estomas y la
turgencia.
2.1.3. Efecto de la luz en las plantas.
La radiación solar es la única fuente de energía natural disponible para la actividad
metabólica de las plantas verdes. La energía llega a la planta en forma de flujo o radiación
procedente del sol, tras haber sido difundida por la atmósfera, reflejada o transmitida por otros
objetos. Las cantidades relativas de radiación directa y difusa que llegan hasta una hoja dependen
de la cantidad de polvo existente en el aire y del grosor de la capa de aire dispersante que se
encuentra entre el sol y la planta (lo que depende de la latitud y la hora). La luz es
funcionalmente esencial para los organismos fotosintetizadores que transforman y almacenan la
energía solar en forma de compuestos de alta energía (Long y col., 1994).
La absorción excesiva de energía luminosa puede resultar en un daño de los complejos
proteína-pigmento de la membrana tilacoildal debido a la formación de especies reactivas de
oxígeno. Los complejos proteína-pigmento del fotosistema II son más sensibles al fotodaño
(Long y col., 1994; Korneev y col., 2005). Aunque la luz interviene directamente en la
fotosíntesis, también puede regular otros mecanismos metabólicos en el proceso del desarrollo de
las plantas (fotomorfogénesis). La relación de la luz con la fotomorfogénesis y los sistemas de
fitocromos ha sido estudiada intensivamente debido a la influencia directa que tienen sobre la
calidad y atributos de las plantas, (Hess, 1975; Dorais y col., 2001).
Un ejemplo evidente de la influencia de la luz, fue el experimento reportado por Ubaldo
(2007), en el cual se cultivó Tagetes erecta y se evaluaron dos ambientes de Radiación
Fotosintéticamente Activa (RFA); para esto algunas plantas se expusieron directamente a la luz

7
natural y otras recibieron la luz a través de una malla de media sombra. Observaron que el cultivo
que estuvo sin malla recibió mayor RFA, radiación UV y presentó una disminución en el
contenido de carotenoides (luteína y zeaxantina), en las inflorescencias, debido
fundamentalmente a que son moléculas que están sujetas a transformaciones fotoquímicas.
2.1.4. Efecto del fotoperiodo en las plantas.
El éxito de una plantación vegetal y la calidad de sus plantas están definidos por su
comportamiento final en el terreno, todo esto regulado por los atributos morfológicos y
fisiológicos de ellas, y por la interacción con el ambiente. Los viveristas evalúan diversos
atributos del comportamiento de las plantas, tales como frío resistencia y potencial de
crecimiento radicular (RGP). Además, evalúan atributos fisiológicos, entre los que destacan las
reservas de carbohidratos y el estatus nutricional de la planta. Diversos investigadores han
establecido que “la frío-resistencia” constituye una de las respuestas fisiológicas de la planta que
es controlada por el fotoperiodo, el cual es dependiente de la duración relativa del día y la noche
(Arévalo, 1994; Barceló y col., 1995).
Las fluctuaciones en la duración del día (fotoperiodo) proveen información que usan las
plantas para sincronizar su desarrollo con las estaciones. En este sentido, Sánchez-Olate y col.
(2003) desarrollaron cultivos de platas de Eucalyptus globulus en vivero para ver el efecto del
fotoperiodo en el desarrollo de las plantas, en donde evaluaron dos tratamientos uno de 7 h y un
control de 10 h de luz continua, durante 7 y 15 días y encontraron que la reducción del
fotoperiodo por 7 días disminuyó significativamente el contenido de carbohidratos solubles
totales, pero a medida que se aumentaron las horas del tratamiento también se incrementó el
contenido de CST en cantidades similares a los del tratamiento control. Las diferencias podrían
estar explicadas por la tasa fotosintética, lo que explicaría una mayor concentración de
carbohidratos solubles (Salisbury y Ross, 1992).
2.2. Factores que inducen la floración.
El proceso de floración puede dividirse en tres etapas: inducción, evocación y desarrollo. La
inducción involucra los eventos que señalan a la planta que debe alterar su desarrollo y orientarlo
hacia la reproducción. La evocación incluye los eventos que ocurren en los ápices después de la

8
inducción y que provocan la reorganización de las yemas para producir primordios florales en
lugar de foliares. El desarrollo de estos primordios florales da origen a las diferentes estructuras
florales y finalmente a la flor madura. Siendo la floración el primer paso hacia la reproducción
sexual, los factores que la promueven revisten especial importancia para la producción y
mejoramientos de la plantas de uso agrícola. Para regular su transición hacia el estado floral,
muchas plantas utilizan señales del medio ambiente (por ejemplo el fotoperiodo, temperatura y
disponibilidad de agua). Dichas señales promueven la floración sincronizada (en los mismos
periodos) para garantizar el éxito de la reproducción sexual, especialmente en el caso de las
plantas alógamas. Sin embargo, en otras plantas como los chicharos (Pisum sativum) por
ejemplo, el paso hacia el estado floral está fijado genéticamente, y por lo tanto ocurre
independientemente de la influencia de las condiciones ambientales (Guevara y Jiménez, 2006).
2.2.1. Influencia de la temperatura y fotoperiodo sobre la floración.
La temperatura y el fotoperiodo son factores que afectan los procesos indispensables para el
desarrollo de las plantas (Morris, 1957; Ormrood, 1978). La floración es cuantitativa o
cualitativamente dependiente de la exposición a bajas temperaturas por un período de tiempo, lo
cual es conocido como vernalización (Salisbury y Ross 1992).
Hay varios ejemplos de interacciones entre temperatura y fotoperiodo, particularmente con
respecto a la floración (Salisbury y Ross, 1992). Keeve y col., (2000) realizaron un estudio con
Lupinos albus bajo condiciones de campo. Ellos utilizaron cinco fenotipos diferentes (Esta,
Hantie, Tifwhite, Kiev y LAL 186) y encontraron que la fecha de siembra, el genotipo y la
interacción de la fecha de siembra con el fenotipo están relacionados con el período de floración,
sin embargo una disminución en la temperatura hasta 0° C promovió la presencia del fenómeno
de vernalización afectando a los fenotipos provocando un retraso en la floración. En este sentido
cuando ocurrió un aumento en la temperatura ambiental en los meses en que los días son más
largos, se presentó una floración más pronta en los fenotipos, pero esto pudo estar relacionado
con un efecto combinado del fotoperiodo.

9
2.3. Producción de metabolitos secundarios en las plantas.
Las plantas han sido utilizadas por el hombre a lo largo de muchos años como fuente para
elaborar medicinas, aromatizantes o pigmentos. Primero comenzaremos diciendo que los
metabolitos secundarios (MS) se caracterizan por la gran heterogeneidad de su estructura
química, distribución restringida, además de que se forman gracias a la acción de enzimas
codificadas por material genético especial. Su biosíntesis (también llamado metabolismo
secundario) tiene un control estricto, por medio de la regulación de la cantidad de enzimas
involucradas y de la actividad de estas, de modo que existe una compartamentalización de
enzimas, precursores, intermediarios y productos involucrados en su biosíntesis, almacenamiento
y desintegración (Wink, 1999).
Al respecto con la producción de los principios activos, o sea, de los metabolitos secundarios
elaborados por los vegetales y que son usados por poseer actividad farmacológica, se ha
reportado que además de los componentes hereditarios, el medio ambiente es un factor que
determina la expresión de los genes que dirigen la elaboración de dichas sustancias; es decir, que
su producción por la planta tanto cualitativa como cuantitativa, está condicionada a algunos
agentes externos, como la vegetación circundante, la presencia de determinado parásito, la
influencia de los elementos del tiempo atmosféricos (fuerza del viento), intensidad y variación
diaria de horas luz (fotoperiodo), el clima en lo general y el microclima (la topografía,
temperatura y humedad del aire), la disponibilidad de agua su exceso o limitación y la
abundancia o escasez de algún nutrimento en el suelo (Waizel, 2006).
2.3.1. Presencia de metabolitos secundarios en plantas.
Existe una gran cantidad de metabolitos secundarios producidos por las plantas y se pueden
clasificar según la presencia o no de nitrógeno en su composición, no obstante, los tres grupos de
metabolitos secundarios más importantes son los terpenoides (isoprenoides), fenilpropanoides
(compuestos fenólicos) y los alcaloides (llevan nitrógeno en su estructura). Los alcaloides
contienen uno o más átomos de nitrógeno y derivan principalmente de aminoácidos, de ellos se
conocen unos 12,000. Mientras que los aproximadamente 8,000 fenilpropanoides provienen de
las llamadas vías biosintéticas del shikimato o del malato/acetato (Valle-Piqueras, 2004).

10
Por su parte los terpenoides derivan del isopentenil difosfato (IPP) y se conocen unos 25,000.
Se clasifican por el número de unidades de isopreno que los componen. Como por ejemplo el
isopreno (hemiterpenos) el cual es el terpenoide mas simple, le siguen los monoterpenos
(formados por dos moléculas de isopreno), sesquiterpenos (formados por tres moléculas de
isopreno), diterpenos (formados por cuatro moléculas de isopreno), triterpenos (formados por seis
moléculas de isopreno), tetraterpenos (formados por ocho moléculas de isopreno) y politerpenos
(formados por más de ocho moléculas de isopreno). Al grupo de los tetraterpenos pertenecen los
carotenoides, que son pigmentos que poseen funciones importantes en la fotosíntesis (Valle-
Piqueras, 2004).
2.4. Cempaxúchil (Tagetes erecta L.) como modelo de estudio.
Cempoalxóchitl es un vocablo de origen náhuatl que se usó en la época prehispánica para
referirse de forma genérica a un grupo de plantas con características comunes: flores vistosas por
su forma y tamaño, con diversidad aromática y de colores llamativos; tales peculiaridades le
facilitaron al hombre prehispánico la identificación y clasificación en variedades: cempoalxóchitl,
macuilxóchitl, tepecempoalxóchitl, oquichtli, tlapalcozatli, zacaxochitlcoztic y tlapaltecacayactli.
Algunos grupos indígenas reconocen actualmente diversos tipos de cempoalxóchitl (Serrato-
Cruz, 2004).
Por otro lado, también tienen algunos nombres comunes en otros idiomas indígenas de
México como son; Apátsicua (lengua tarasca), guie'biguá, guie'coba, picoa, quiepi-goa (lengua
zapoteca), kalhpu'xa'm (lengua totonaca), ita-cuaan (lengua mixteca), jondri (lengua otomí),
musá, musajoyó (lengua zoque), cempaxúchil y xumpatsunchitl (Martínez, 1979).
Por otro lado, también tienen algunos nombres comunes en otros idiomas indígenas de
México como son; Apátsicua (lengua tarasca), guie'biguá, guie'coba, picoa, quiepi-goa (lengua
zapoteca), kalhpu'xa'm (lengua totonaca), ita-cuaan (lengua mixteca), jondri (lengua otomí),
musá, musajoyó (lengua zoque), cempaxúchil y xumpatsunchitl (Martínez, 1979).
México es considerado centro de radiación primaria de varias tribus, subtribus, géneros y
especies que integran a la familia Asteraceae. El género Tagetes es de origen americano y más
del 30% de las especies que lo integran encuentran su asentamiento en México. Las especies

11
Tagetes erecta L., T. patula L., T. tenuifolia Cav. y T. minuta, son reconocidas por la UPOV
(Unión Internacional para la Protección de Variedades) como plantas ornamentales. Las
principales formas de uso de estas especies corresponden a macetas, bancales en jardinería, flor
cortada en racimos y flores para uso agroindustrial. Las tres primeras especies se distribuyen
desde México hasta Guatemala, mientras que T. minuta se encuentra dispersa en América del Sur.
Las especies más importantes o más conocidas como ornamentales son T. erecta y T. patula
debido al gran tamaño y color (rojo, amarillo, anaranjado y combinaciones) de la inflorescencia,
atributos por las cuales estas especies fueron utilizadas por los grupos indígenas mesoamericanos,
antes de la llegada de los españoles a América (Serrato-Cruz, 2003).
Debido a que las especies cultivadas T. erecta y T. patula, conocidas como plantas
cempoalxóchitl, acumulan carotenoides en sus inflorescencias y las cuales presentan una amplia
variabilidad en México, ha sido suficiente razón para considerar a estas plantas como oriundas
de este país (Serrato-Cruz, 2003).
2.4.1. Clasificación botánica de T. erecta L.
El cempaxúchil es una planta herbácea anual o perenne, de días cortos. La raíz es cilíndrica,
pivotante, con un sistema ramificado fibroso y poco profundo. El tallo es estriado a veces
acostillado, glabro o ligeramente pubescente, cilíndrico, ovalado y de herbáceo a ligeramente
maderable, con canales de resina en la corteza, son aromáticos al estrujarse. La hojas opuestas en
la parte inferior, alternas en la parte superior; de hasta 20 cm de largo, pinnadas, de 11 a 17
foliolos, lanceolados a linear-lanceolados, de hasta 5 cm de largo y 1.5 cm de ancho, agudos a
acuminados, aserrados a subenteros, los inferiores de cada hoja frecuentemente setiformes (en
forma de hilos), los superiores reducidos, a veces completamente setiformes; con glándulas
redondas abundantes. (Vázquez-García, y col., 2002; Rzedowski y Rzedowski, 2005).
La principal característica de estas flores es que están agrupadas en cabezuelas o en
inflorescencias solitarias, sobre pedúnculos de hasta 15 cm de largo, son liguladas de colores
amarillos a rojo. En las flores del disco: 150 a 250 en las cabezuelas sencillas, en las "dobles"
muestra diferentes grados de transformación en lígulas, corolas amarillas a anaranjadas, de 8 a 10
mm de largo. Los frutos y semillas son: aquenios lineares de 7 a 10 mm de largo, glabros o
hispídulos en los ángulos (Mondragón y col., 2007).

12
2.4.2. Clasificación taxonómica.
Reino: Plantae.
Subreino: Traqueobionta (plantas vasculares)
Superdivisión: Spermatophyta (plantas con semillas)
División: Magnoliophyta (plantas con flor)
Clase: Magnoliopsida (dicotiledóneas)
Subclase: Archichlamydeae
Orden: Asterales
Familia: Asterareae
Subfamilia: Asteroideae
Género: Tagetes
Especie: Tagetes erecta L.
2.4.3. Distribución geográfica.
El área de origen de Tagetes erecta es Mesoamérica (México y Centroamérica), en donde las
formas más silvestres se encuentran en la cuenca del Balsas y el occidente de México y habita en
diversos tipos de ecosistemas, como selvas tropicales de hoja caduca, bosques espinosos, bosques
de niebla y bosques de pino-encino (Alvarado, 2004).
REPÚBLICA MEXICANA
Figura 1. Distribución secundaria de las formas domesticas de cempaxúchil en México.

13
La distribución secundaria de formas asilvestradas de las plantas domesticadas se encuentran
en los trópicos a nivel mundial a una altitud de 2,450 metros sobre el nivel del mar,
específicamente su distribución en México (Figura 1) se ha registrado en Aguascalientes,
Campeche, Chiapas, Coahuila, Distrito Federal, Durango, Guerrero, Hidalgo, Jalisco, estado de
México, Michoacán, Morelos, Oaxaca, Puebla, Querétaro, Quintana Roo, San Luis Potosí,
Sinaloa, Sonora, Tabasco, Tamaulipas, Tlaxcala, Veracruz, Yucatán y Zacatecas (Villaseñor-
Ríos y Espinosa-García, 1998).
2.4.4. Producción de T. erecta en México.
El uso de las plantas de cempaxúchil (Tagetes erecta L.) para jardinería y maceta es cada vez
más aceptado en México. La venta de esta planta en maceta durante la festividad de Día de
Muertos se ha incrementado desde el 2000 con un consumo de 10 000 plantas, aumentando a 500
000 en el 2004, y se estimaba un volumen de venta de 700 000 para 2005; en 15 años el consumo
nacional de esta especie para maceta alcanzará los 4 millones de plantas y su empleo fuera de esta
festividad cada vez será mayor (Empresa Plántulas de Tetela, Cuernavaca, Morelos;
comunicación personal, 2005). Empresas semilleras como Sakata, Seeds, Ball Seeds,
Goldschmith Seeds entre otras cubren, actualmente, el abasto de semillas para este fin, mediante
importaciones que solicitan viveristas y floricultores (Serrato-Cruz, 2005).
Por otro lado la superficie sembrada para la producción de cempaxúchil de temporal en
México es de aproximadamente 740 hectáreas, el de riego es de 1295 ha en el 2003 a 8849 ha en
el 2006, con una producción de hasta 10.93 t/ha (Delgado, 1997; SIACON SAGARPA, 2006). A
continuación se muestra un resumen nacional de la producción de cempaxúchil Según datos del
SIACON (Sistema de Información Agroalimentario de Consulta), SAGARPA de los años 2003-
2006 (Cuadro 1).

14
Cuadro 1. Producción de cempaxúchil a nivel nacional.
Descripción 2003 2004 2005 2006
Superficie sembrada total (ha) 2035 5384 5331 8849
Volumen de Producción (Ton) 22243 13677.2 10005.1 5374
Rendimiento (t/ha) 10.93 2.54 1.88 1.64
Datos tomados del Sistema de Información Agroalimentaria de consulta (SIACON SAGARPA,
2006).
Para el estado de Morelos, la superficie sembrada solo se registró un valor (riego más
temporal) teniendo un máximo de 8.8 hectáreas en el 2005 (Cuadro 2).
Cuadro 2. Producción de cempaxúchil en el estado de Morelos.
Descripción 2003 2004 2005 2006
Superficie sembrada total (ha) 2 2.1 8.8 5
Volumen de Producción (Ton) 18 21.2 80.1 50
Rendimiento (t/ha) 9 10.095 9.102 10
Datos tomados del Sistema de Información Agroalimentaria de consulta (SIACON SAGARPA,
2006).
Aunque en México existe una amplia diversidad de germoplasma de T. erecta asociada a
procesos de domesticación milenarios realizados por grupos indígenas mesoamericanos y por ser
el centro-sur de México el lugar de origen de este taxa, no se ha hecho trabajo de mejoramiento
en nuestro país para la selección de portes bajos con inflorescencias dobles para su
aprovechamiento ornamental en maceta o en jardinería, lo cual va relacionado con la falta de
estudio sistemático de este recurso filogenético: no se tiene banco de germoplasma ni se conoce
su variación (pigmentos, colores y tipos de flor, porte de la planta, aroma, ramificación, tamaño
de flor, resistencia a plagas y enfermedades, duración de la floración) por regiones, asociada con
grupos humanos y diversidad ambiental (Serrato-Cruz, 2005).

15
2.4.5. Componentes bioactivos de Tagetes spp.
Benk y col., (1976), reportaron que un extracto de Tagetes erecta contiene aproximadamente
27% de carotenoides con 4% de β-caroteno, 1.5% esteres de criptoxantina y 86.1% de esteres de
xantófilas. Este último es el principal carotenoide constituyente de los pétalos de T. erecta donde
se encuentran en mayor cantidad que en las semillas o los sépalos, y 200 veces más que el maíz
amarillo.
T. erecta merece mención especial debido a que presenta los tonos más intensos del color
anaranjado en las inflorescencias y es conocida como la especie más popular ya que aporta varios
beneficios, entre otros: pigmentos utilizados como colorantes vegetales en alimentos para
humanos (huevos con yemas amarillas, coloración de sopas de pasta, piel y grasa amarilla de aves
y reses) y carotenoides de uso terapéutico como las capsulas de luteína (Serrato-Cruz, 2003).
Su gran diversidad de tonalidades está relacionada con un mayor contenido de carotenoides.
Debido a esto, es importante conocer si la tonalidad de las flores de Tagetes van cambiando con
respecto al tiempo de siembra de las plantas, por lo que el índice de color (IC*) por sus
características de variación se ha utilizado como variable de control de la calidad organoléptica
de alimentos (Martínez-Peña y Cortés-Cuevas, 2004). Empresas mexicanas como Bioquimex y
PIVEG tienen una amplia experiencia en el mejoramiento genético para la obtención de plantas
con alta cantidad de carotenoides. De igual manera, en el Centro de Investigaciones Científicas
de Yucatán y en el Cinvestav-Irapuato, también se está trabajando en forma decidida para
desentrañar los mecanismos de regulación de la expresión o síntesis de carotenoides (Manzanero
y col., 2002).
2.5. Los carotenoides.
Los carotenoides son importantes fitoquímicos que tienen una gran diversidad estructural y
biológica (DellaPenna, 1999). Son los responsables de impartir el color rojo, anaranjado y
amarillo de las frutas, y también contribuyen a darle brillo al color de flores y frutos (Zariphep y
Erdma, 2002). Su presencia en diversas estructuras de plantas y en gran variedad de animales,
algas, hongos y bacterias se ha descrito desde hace décadas (Karrer y Jucket, 1948; Goodwin,

16
1980; Britton, 1983; Goodwin, 1984). En la naturaleza encontramos más de 600 carotenoides de
los cuales 40 se consumen en la dieta humana (Bendich, 1993).
Se puede afirmar que entre todos los pigmentos presentes en los organismos vivos, no hay
duda de que los carotenoides son, después de las clorofilas, los más ampliamente distribuidos en
la naturaleza. Éstos son compuestos liposolubles que se localizan en las células vegetales, en el
interior de orgánulos especializados, cloroplastos y cromoplastos (Kopsell y Kopsell, 2006); en
los primeros acompañan a las clorofilas. En el caso de los frutos maduros, se acumulan en los
plastoglóbulos de los cromoplastos en forma masiva y es donde la diversidad estructural alcanza
un mayor grado.
La presencia del extenso sistema de dobles enlaces conjugados de la cadena poliénica de los
carotenoides forma un cromóforo (parte de la estructura responsable de la absorción de luz
visible y por tanto del color del compuesto) cuya capacidad de absorción de luz da lugar a los
llamativos y característicos colores de estos pigmentos. El número de dobles enlaces conjugados
y la presencia de diferentes grupos funcionales determinarán en última instancia las
características espectroscópicas propias de cada pigmento (Gandul y Mínguez, 1996).
Químicamente la mayoría de los carotenoides son tetraterpenoides, compuestos de 40 átomos
de carbono formados por ocho unidades isoprenoides unidas de forma que la secuencia se
invierte en el centro de la molécula. Es decir, la unión de dichas unidades es “cabeza-cola”,
excepto en el centro de la molécula, donde es “cabeza-cabeza”. Debido a ello, los dos grupos
metilo centrales de la cadena poliénica están separados por seis átomos de carbono, mientras que
el resto están separados por cinco. Algunos carotenoides son acíclicos (Figura 2), aunque la
mayoría contienen anillos en uno o ambos extremos de la cadena (Figura 3). Considerando los
elementos químicos presentes en sus moléculas, los carotenoides pueden dividirse en dos grandes
grupos: carotenos que son hidrocarburos y xantófilas que contienen átomos de oxígeno
(Meléndez-Martínez y col., 2007).

17
Figura 2. Estructuras de los principales carotenoides acíclicos en orden ascendente de
acuerdo al número de dobles enlaces (n). (Adaptado de Rodríguez-Amaya, 2001).

18
Figura 3. Estructuras de los principales carotenoides cíclicos, los cuales poseen el anillo en
uno o ambos extremos de la cadena hidrocarbonada (Adaptado de Rodríguez-Amaya, 2001).
Los carotenoides además de contener átomos de carbono e hidrógeno contienen oxígeno, el
cual conforma otros dos grandes grupos, los hidroxicarotenoides (Figura 4) y los
epoxicarotenoides (Figura 5), los cuales son denominados xantófilas.

19
Las xantófilas hidroxílicas pueden existir en la naturaleza en estado libre o esterificadas con
ácidos grasos (palmítico, linoleico, linolénico, esteárico, mirístico, láurico, oleico, etc.) en chiles
y derivados, papas, mango, cítricos, etc, (Phillip, 1973; Breithaupt y Bamedi, 2001; Pott y col.,
2003; Ishida y col., 2004).
Figura 4. Estructuras de carotenoides que en su esqueleto tienen grupos hidroxilo (Adaptado de
Rodríguez-Amaya, 2001).

20
Figura 5. Estructuras de carotenoides que en su esqueleto contienen grupos epóxido. (Adaptado
de Rodríguez-Amaya, 2001).
No todos los carotenoides constan de ocho unidades isoprenoides, ya que algunos,
denominados apocarotenoides, poseen un esqueleto de menos de 40 átomos de carbono, debido
probablemente a escisiones en uno o ambos extremos de la molécula, como por ejemplo el β-apo-
8’-carotenal (Figura 6), pigmento presente en el níspero y en cítricos (Gross y col., 1973) o la
crocetina, pigmento característico del azafrán (Carmona y col., 2006).

21
Figura 6. Estructuras de apocarotenoides los cuales poseen en su esqueleto menos de 40 átomos
de carbono (Adaptado de Meléndez-Martínez y col., 2007)
2.5.1. Biosíntesis de los carotenoides
Los terpenos son un grupo diverso de metabolitos secundarios vegetales, formados por la vía
de la acetil–CoA (vía del acido mevalónico) o de intermediarios glicolíticos (vía metileritriol
fosfato). La vía básica de biosíntesis de los carotenoides fue descubierta a mediados de los 1960
(Fraser y Bramley, 2004) y actualmente la mayoría de los genes que codifican para las enzimas
que participan en la vía han sido clonados (Kopsell y Kopsell, 2006).
La síntesis de los carotenoides se lleva a cabo en los plastidios y son derivados del isopentil
difosfato (IPP) que es sintetizado por dos vías metabólicas en las plantas (Lichtenthaler, 1999).
La primera vía se desarrolla en el compartimiento citoplasmático a partir del ácido mevalónico y
produce compuestos como: esteroles y citocininas, y la segunda es la responsable de la
biosíntesis de las giberelinas (GA), carotenoides, y ácido abscísico (ABA) que también
contribuye a la biosíntesis de tocoferoles conocidos como clorofila a y b (Laule y col., 2003).
En el primer paso de la biosíntesis de carotenoides el IPP que es un isómero del dimetil
alilpirofosfato (DMAPP), se condensa con su propio isómero, DMAPP para producir

22
inicialmente el geranil pirofosfato (GPP), posteriormente el farnesil pirofosfato (FPP) y
finalmente el geranilgeranil pirofosfato (GGPP) (Delgado-Vargas y col., 2000) (Figura. 7). Las
unidades se van uniendo conforme a un patrón cabeza-cola hasta antes de la formación de GGPP,
la cual se origina de la unión cola-cola de dos moléculas de GGPP para producir un compuesto de
40 carbonos con el orden invertido en el centro (Delgado-Vargas y Paredes-López, 2003), el cual
da origen al fitoeno, primer carotenoide producido en la ruta, que es incoloro. La insaturación
posterior de la cadena hidrocarbonada produce dobles enlaces conjugados, lo que le da a la
molécula la capacidad de absorber luz de ciertas longitudes de onda y provocar la pigmentación
de los organismos o tejidos que los contienen.
El licopeno es el precursor biosintético de la mayoría de los carotenoides cíclicos; la
conversión de licopeno a β-caroteno se debe a la acción de la enzima β-licopeno ciclasa (lcy.b),
enzima que cataliza la formación de anillos β-ionona en los extremos de la molécula
(Cunningham y col., 1996).
La ciclación del licopeno se ha planteado como un punto de regulación importante en la ruta
de biosíntesis de los carotenoides. La formación del ciclo en ambos extremos de la molécula de
licopeno (Figura 7) da como resultado los α-, β- ó ε-caroteno. La reacción de ciclación es
catalizada por las enzimas β- o ε-licopeno ciclasa (Bartley y Scolnik, 1995).
Cuando la enzima β-licopeno ciclasa actúa en ambos lados de la molécula ocurre la
formación de β-caroteno, el cual es precursor de xantófilas como la zeaxantina, violaxantina y la
anteraxantina (Figura 7). Las xantófilas se producen cuando uno o más átomos de oxígeno en
grupos ceto, hidroxi, epoxi, etc. se introducen a la molécula del caroteno (Cunningham y col.,
1996; Ronen y col., 1999). La enzima ε-licopeno ciclasa puede actuar en ambos extremos de la
molécula y producir el ε-caroteno; este compuesto se ha localizado en muy pocos organismos y
en cantidades muy bajas (Cunningham y Gantt, 2001). Los anillos ε difieren de los β en la
posición del doble enlace dentro del ciclohexano; en cualquier caso, en plantas es común
encontrar anillos modificados (Pecker y col., 1996).
Finalmente, otro grupo de carotenoides bicíclicos tiene un anillo β y un anillo ε en su
estructura. Estos anillos son comunes en plantas y animales además son precursores de varias

23
xantófilas, de las que la luteína (Figura 7) es el ejemplo principal en muchas plantas y algas
(Bartley y Scolnik, 1995; Chappell, 1995; Pogson y col., 1996).
Figura 7. Ruta de biosíntesis de los carotenoides (Adaptado de Bungard y col., 1999;
Lichtenthaler, 1999 y Buchanan y col., 2000).

24
2.5.2. Función de los carotenoides.
La principal función biológica de los carotenoides es la de servir como pigmentos accesorios
en la recolección de la luz en el proceso fotosintético, y como sustancias fotoprotectoras,
inhibiendo la propagación de especies reactivas de oxígeno y otros radicales libres, por tanto
impidiendo la reacción nociva de estos a nivel celular (Mínguez y Homero, 1993). Además son
estructuras que determinan la pigmentación de plastidios y del complejo proteico, así en tejidos
fotosintéticos realizan funciones de fotoprotección y en no fotosintéticos determinan el color de
las flores y frutos (Bartley y Scolnick, 1995).
2.5.2.1. Función fotoprotectora de los carotenoides.
Los procesos de fotoprotección actúan disipando la energía de excitación que llega a la
molécula de clorofila y que no se puede utilizar en las reacciones fotoquímicas. En las plantas,
altos niveles de radiación, provocan fotoinhibición de la fotosíntesis y destrucción fotooxidativa
del aparato fotosintético (Osmond, 1994) caracterizado por una reducción en la eficiencia de la
utilización de la luz (Krause y Weis, 1991).
La fotoinhibición es provocada por la pérdida del funcionamiento de los fotosistemas II
(FSII) que se manifiesta como una disminución, transitoria o permanente, en la eficiencia
cuántica de la fotosíntesis (mol de CO2 fijado por mol de fotones absorbido) en niveles de luz de
baja intensidad (Osmond, 1994).
La “fotoprotección" en la cual la excesiva energía de excitación es reorientada y disipada
principalmente como calor, logra obtener un balance entre la energía recibida por los FSII y la
capacidad para utilizarla (Baker 1991; Demmig-Adams y Adams 1992; Aro y col., 1993; Long y
col., 1994). Esto forma parte de un mecanismo que la planta pone en marcha para disipar el
exceso de energía, cuando no todos los fotones absorbidos por las clorofilas pueden ser utilizados
en el proceso fotoquímico, para evitar el daño de las membranas fotosintéticas y la propia
clorofila.
Por lo general y en condiciones normales las plantas están bien adaptadas a los cambios de
luz diurnos, estacionales y momentáneos, siempre y cuando la heterogeneidad en el

25
comportamiento lumínico muestre una cierta pauta más o menos predecible a lo largo del
período. En este sentido puede llegar a suceder, cuando los cambios lumínicos se suman a la
acción de otros factores ambientales negativos, como las bajas o muy altas temperaturas
ocasionales, sequías prolongadas o deficiencia de nutrientes, que haya un punto de inflexión que
ponga a prueba todos los mecanismos de defensa de la planta llevándola incluso a la muerte, si la
acción del factor se prolonga durante un tiempo suficiente. De modo que los carotenoides como
transductores de energía hacia los centros de reacción no son muy eficientes (30 al 40 % de
eficiencia); pero como disipadores de la energía absorbida en exceso por la clorofila son
altamente eficientes. Debido a lo anterior el caroteno más importante en las plantas es el β-
caroteno y la luteína es la principal xantófila, pero hay además otras xantófilas que aun no
estando en tan alta concentración como los anteriores, juegan un papel decisivo en la disipación
de la energía excedente, son estas: la violaxantina, la anteraxantina y la zeaxantina (Manrique-
Reol, 2003).
2.5.2.2. Ciclo de las xantófilas.
El ciclo de las xantofilas está ampliamente distribuido en la naturaleza y se localiza en las
membranas tilacoidales de plantas superiores, hongos y algas. Existen dos variantes del ciclo, el
de la violaxantina, que es el más común en las plantas superiores y el ciclo de la diadinoxantina
en algunos tipos de algas (Adamska, 1997).
La importancia del ciclo de las xantófilas es que tiene una función fotoprotectora la cual es
“atrapar” el exceso de energía presente en la antena fotosintética y disiparla en forma de calor
(Eskling y col., 1997). Otras funciones que se le han asignado a este ciclo han sido la de protector
contra el estrés oxidativo de los lípidos de la membrana (Havaux y col., 1991), modulación de la
fluidez de la membrana (Havaux y Gruszecki, 1993) y regulación de la síntesis del ácido
abscísico (Marín y col., 1996).
La exposición de las plantas a una radiación superior a la que puede ser usada en el proceso
de la fotosíntesis induce cambios en el ciclo de las xantófilas, caracterizado por la conversión de
violaxantina en anteraxantina y posteriormente en zeaxantina, mediante una serie de
epoxidaciones inducidas por la luz. El ciclo normalmente se orienta hacia la formación de
violaxantina, que es la que predomina en las membranas fotosintéticas. Cuando la energía

26
absorbida por la clorofila resulta excesiva, bien por un aumento de la intensidad de radiación o
porque el aparato fotosintético haya sufrido algún cambio y no esté operando al 100%, de tal
manera que el pH del lumen disminuye hasta 5 (valor óptimo de la enzima violaxantina
desepoxidasa, produciéndose la conversión en anteraxantina y posteriormente en zeaxantina. Se
ha sugerido que ambos pigmentos, zeaxantina y anteraxantina, actúan como intermediarios en el
proceso de fotoprotección del aparato fotosintético (Verhoeven y col., 1999 a,b).
Cuando las condiciones se revierten y la energía recibida no excede a la que puede ser
utilizada en la fotosíntesis, la zeaxantina vuelve a convertirse en anteraxantina y violaxantina por
la acción de la enzima zeaxantina oxidasa (Adams y Demmig-Adams, 1994).
El ácido ascórbico juega un papel de cofactor de la enzima violaxantina-desepoxidasa
(VDE). En plantas aclimatadas a altas irradiaciones el ácido ascórbico es necesario para sostener
una adecuada actividad de VDE. La necesidad de ácido ascórbico sugiere un mecanismo (para la
desepoxidación) en el cual ocurre reducción de la VDE seguido de una deshidratación, pero se
carece de evidencia directa de este mecanismo (Figura 8). Durante el proceso de fotoxidación de
la zeaxantina a la violaxantina se consume O2 y en el proceso de fotorreducción se libera O2
(Yamamoto y col., 1999).
Figura 8. Ciclo de las xantófilas y sus efectos sobre la captura de luz y la disipación de la energía
dentro del aparato fotosintético (Adaptado de Ubaldo, 2007).

27
Durante los últimos años, se ha prestado mucha atención a la función protectora que tiene la
zeaxantina. El papel que se ha dado a este pigmento se basa en las medidas de “quenching” de
fluorescencia (disipación de la energía en forma de calor) y en la gran correlación que se ha
encontrado entre el quenching no fotoquímico y la concentración de zeaxantina y anteraxantina
(Demmig-Adams y Adams, 1996). El cual está regulado por la magnitud del gradiente de pH que
se genera a través de la membrana del tilacoide, y en especial por los pigmentos del ciclo de las
xantófilas. Por último, Gruszecki y col., (1996) encontraron que la conversión de violaxantina en
zeaxantina, tiene como resultado un aumento del transporte electrónico cíclico alrededor del PSII,
disminuyendo el flujo electrónico vectorial que comienza con la fotolisis del agua.
2.5.3. Factores que influyen en la acumulación de carotenoides.
La acumulación de carotenoides en tejidos específicos de las plantas está relacionada con las
cualidades fisiológicas, genéticas y bioquímicas de cada especie, así como con los factores
ambientales, como son la luz (el fotoperiodo y la irradiación), la temperatura y la fertilidad de los
suelos (Hochmuth y col., 1999; Kopsell y col., 2004; Chenard, 2005; Lefsrud y col., 2006 a y b).
Así mismo, la composición de los carotenoides contenidos en los vegetales puede ser afectada
por factores como la variedad o el cultivar, la parte de la planta utilizada, el clima o la geografía
del sitio de producción, la cosecha, el manejo poscosecha, el proceso y el almacenamiento
(Rodríguez-Amaya 1999).
Un experimento en donde se refleja la influencia de la luz sobre la acumulación de
carotenoides, es el que realizaron Sánchez-Millán y col., (2006), en donde analizaron el efecto de
la RFA (Radiación Fotosinteticamente Activa) en modelos in vivo e in vitro, sobre soluciones de
luteína el cual es el carotenoide más abundante en Tagetes erecta, este experimento lo realizaron
en cultivos mantenidos bajo dos ambientes de luz solar (un invernadero de plástico e invernadero
de vidrio) y concluyeron que la biosíntesis de carotenoides en Tagetes erecta puede ser afectada
drásticamente por la cantidad y calidad de la luz y por el tiempo de exposición, ya que se afectó
la estructura de las especies químicas de los carotenoides favoreciendo formación de carotenoides
del tipo cis.
Por otra parte, los cambios en la coloración de los vegetales asociados con la madurez,
ocasionan un aumento en la concentración de los carotenoides (Howard y col., 2000, Russo y

28
Howard, 2002; de Azevedo y Rodríguez-Amaya, 2005); sin embargo, se reducen con la
senescencia de tal manera que la manipulación de los tiempos de siembra y las condiciones de
crecimiento pueden afectar su concentración (Kopsell y Kopsell, 2006). Además, los pétalos
Tagetes erecta secados en horno causan más pérdida de carotenoides que el secado al aire libre y
que el congelamiento o las bajas temperaturas de almacenamiento, las cuales generalmente
preservan su concentración reduciendo el potencial de oxidación enzimática (Kopas-Lane y
Wathersen, 1995; Rodríguez-Amaya, 1999).
2.5.4. Importancia económica del cempaxúchil y los carotenoides
El cempaxúchil que se produce hoy en día, sólo sirven para atender una pequeña parte de la
demanda ornamental de las festividades del 1 y 2 de noviembre de cada año, por lo que no se
cultiva fuera de esta época. Sin embargo, el auténtico potencial de esta flor está en la industria
alimentaria, principalmente en la avicultura, debido a que son fuente importante de carotenoides
los cuales son de interés económico por su uso en los alimentos, farmacéutica y cosmetológica,
pero en la naturaleza su importancia va más allá de su rol como pigmentos. En forma creciente se
han atribuido a estos compuestos funciones y acciones biológicas. De hecho, por mucho tiempo
se ha sabido de la actividad de provitamina A de los carotenoides. En el área de la biotecnología
existe interés en las propiedades nutricionales y anticancerígenas de los carotenoides. En el año
de 1996 la demanda fue de 240 millones de dólares la cual fue incrementando, para el año 2002
ésta fue de 1000 millones de dólares, esto ha dado la pauta para el desarrollo de la investigación
relacionada con la producción de estos pigmentos (Delgado-Vargas y Paredes-López, 2003).

29
3. JUSTIFICACIÓN
El cempaxúchil es una fuente importante de carotenoides, principalmente de luteína, los
cuales son altamente cotizados en el mercado. Existe poca información en estudios relacionados
con su cultivo en diferente época de transplante y los factores ambientales para producción de
estos pigmentos en el estado de Morelos.
Aunque se identifica a los meses de julio a octubre con la producción de flores de
cempaxúchil en el país, para usos asociados con patrones culturales, actualmente no se cuenta
con suficiente evidencia para promover el cultivo cempaxúchil en diferentes fechas de transplante
para efectos de rendimiento en la producción de los carotenoides.
Por lo tanto, el estudio o seguimiento de los cultivos iniciados en diferentes época de
transplante y su relación con la producción de carotenoides (luteína), permitirá generar el
conocimiento necesario para hacer más eficiente el uso del campo en la producción de flores de
cempaxúchil, aprovechando el intervalo de tiempo en el cual se genere una mayor producción de
los metabolitos de interés, en el estado de Morelos.

30
4. HIPÓTESIS
La acumulación de luteína en inflorescencias de cempaxúchil varía de acuerdo a la época de
transplante a campo.

31
5. OBJETIVOS
5.1. Objetivo General
Determinar el efecto de la época de transplante a campo sobre el desarrollo de la planta y
acumulación de luteína en inflorescencias de cempaxúchil (Tagetes erecta L.).
5.2. Objetivos Específicos
Evaluar la altura de las plantas, número de primordios florales, diámetro, peso fresco y
seco de las inflorescencias de cultivos del cempaxúchil en diferente época de transplante a
campo.
Evaluar el color de las inflorescencias de cempaxúchil cultivadas en diferentes épocas de
transplante.
Cuantificar la luteína obtenida de extractos de lígulas de cempaxúchil cultivadas en
diferentes épocas de transplante.

32
6. MATERIALES Y MÉTODOS
6.1. Obtención de las plantas
El material utilizado en el presente estudio, fueron semillas de Tagetes erecta del fenotipo
porte bajo con dos retro cruzamientos, proporcionadas por el Dr. Miguel Ángel Serrato Cruz
Profesor Investigador de la Universidad Autónoma Chapingo.
Para determinar el efecto de la época de transplante sobre la altura, número de primordios
florales, diámetro, peso fresco, seco y en la acumulación de los carotenoides en inflorescencias
se germinaron semillas cada mes, obteniendo plántulas que fueron transplantadas a maceta a los
diez días después de la germinación y mantenidas en invernadero por 20 días; posteriormente se
pasaron a campo obteniendo cultivos a los que se les dio seguimiento a través de su crecimiento y
desarrollo.
El experimento se realizó en el campo experimental Emiliano Zapata del Centro de
Desarrollo de Productos Bióticos ubicado en la población de San Isidro, municipio de Yautepec,
Morelos. El municipio se localiza en la parte norte del estado, colinda con los municipios de
Cuautla y Atlatlahucan al este, al sur con los municipios de Ayala, Tlaltizapán y Emiliano
Zapata; al oeste colinda con los municipios de Jiutepec y Tepoztlán y finalmente al norte colinda
con el municipio de Tlayacapan. Su localización geográfica es 18° 53´ de latitud norte y 99° 04´
de longitud este con una altura a nivel del mar de 1.210 metros, donde la temperatura media anual
es de 22.7 °C y el tipo de clima es cálido sub-húmedo con lluvias en verano; la precipitación
pluvial es de 945.7 milímetros anuales (INEGI, 2007).
El estudio general fue realizado durante el periodo comprendido del 25 de junio de 2007 al
25 marzo de 2008, en los laboratorios de Biotecnología del CEPROBI y la cuantificación de los
pigmentos se realizó en el Laboratorio de Enzimas del Departamento de Alimentos en la Escuela
Nacional de Ciencias Biológicas del Instituto Politécnico Nacional, Unidad Santo Tomás.

33
6.1.1. Germinación de semillas
La germinación de semillas se llevó a cabo por el método descrito por Ubaldo (2007). En
donde ellos evaluaron el efecto de la calidad de la luz en el porcentaje de germinación en semillas
de cempaxúchil y salud de los brotes.
6.1.1.1. Desinfestación y germinación de las semillas
Se lavaron las charolas utilizando hipoclorito de sodio al 10% para su desinfestación y se
dejaron secar. El área de trabajo se limpió con una solución de hipoclorito de sodio al 2%. Se
escogieron las semillas que tuvieron las siguientes características: tamaño uniforme, estructura
rígida y que no estuvieran pálidas o con color amarillento; después se procedió a la
desinfestación, para lo cual: primero se sumergieron las semillas en etanol al 100% durante un
minuto, después en etanol al 75% durante cinco minutos, posteriormente en hipoclorito de sodio
al 2% durante 15 y por último se sumergen en hipoclorito de sodio al 1% durante 15 minutos.
Entre cada etapa se realizaron lavados de un minuto con agua desionizada estéril, para que no
quedaran remanentes de etanol o de hipoclorito, todo se realizo manteniendo siempre agitación
constante (Vanegas-Espinoza, 2003).
Una vez lavadas las semillas se pusieron a germinar en cámara húmeda. Las charolas se
etiquetaron y mantuvieron a: humedad 80+10% y temperatura 25+4° C, en un cuarto de cultivo
(Ubaldo, 2007).
6.1.2. Aclimatación de las plántulas
La aclimatación de las plántulas, se realizó en invernadero de vidrio, en donde se utilizaron
una mezcla de turba para su transplante a maceta el cual favorece el desarrollo de las plántulas en
invernadero (Serrato-Cruz, 2005).
6.1.2.1. Preparación de sustrato.
El material utilizado como sustrato consistió en una mezcla de sustratos que como son: turba,
vermiculita y agrolita en una relación 3:1:1. Para su preparación, primero se tomaron 100 ml de
agua en un recipiente, para posteriormente mezclarlos con 100 g de la combinación de sustratos;

34
después se homogenizaron los componentes de la mezcla, se ajustó el pH a 5.8±0.2 con una
porosidad al 86% (Evangelista-Lozano y col., 2005).
6.1.2.2 Transplante a maceta
Las plántulas se pasaron a macetas con sustrato orgánico estéril a los 10 días después de
haber germinado, posteriormente las macetas fueron llevadas al invernadero en donde se
mantuvieron hasta los 30 días de edad, para posterior traspaso a campo.
6.1.3. Transplante a campo y mediciones de las condiciones ambientales
Las plántulas se traspasaron a campo (Anexo I) con un tamaño de 12±2.0 cm y con más de
tres hojas verdaderas, a una distancia de 40 cm entre plantas, dándole seguimiento durante su
periodo de crecimiento y desarrollo, en donde se evaluaron las condiciones ambientales
imperantes en los meses del desarrollo del cultivo, las condiciones ambientales evaluadas fueron
la temperatura (T, °C) y humedad relativa (% HR) con un higrotermómetro de la marca Taylor
modelo 06A00, la Radiación solar (RadS, µmol/mes.m2) se midió con un cuantómetro de la
marca Apogee Instruments Inc. Modelo QMSW-55 de la serie 1232 y el fotoperiodo (F, h) horas
de luz disponible durante el día, estas condiciones se registraron diariamente durante todo el ciclo
de los cultivos.
6.2. Evaluación del desarrollo vegetativo.
6.2.1. Altura de las plantas
La altura de las plantas, se determinó midiendo (cm) desde el cuello del tallo hasta la última
yema terminal con una cinta métrica esta medición se inició cuando los cultivos comenzaron la
floración (apertura de los primeros primordios florales) (Ubaldo, 2007).
6.2.2. Número de primordios florales
Se registró la fecha en que aparecieron los primordios florales después del transplante a
campo y se contabilizó el número de primordios florales; así también, comenzando el periodo de
floración, una vez que las inflorescencias abrieron completamente se registró el diámetro (cm) de

35
estas y se pesaron (g); este último dato se utilizó para conocer el peso fresco y peso seco de las
flores, el peso seco de las flores so obtuvo deshidratándolas en una estufa eléctrica marca
SQUAROID (LAB-Line) estufa al vacio Modelo 3608 (De la Cruz y col., 1994).
6.3. Colecta de las inflorescencias
Las inflorescencias fueron colectadas mediante un muestreo completamente al azar con tres
repeticiones, en el día 12 después de haber iniciado la floración (apertura de los primeros
primordios florales), ya que en este día es cuando se presenta una floración más uniforme en los
cultivos (Serrato-Cruz, comunicación personal). Las inflorescencias fueron cortadas al nivel del
borde del receptáculo, en las primeras horas de la mañana y colocadas en bolsas polietileno de
15x25 cm, para su posterior análisis.
6.4. Determinación del índice de color
El color se evaluó por el Sistema CIELAB, mediante la determinación del índice de color
IC* obtenido por la expresión (1) donde “L” proporciona un valor de la iluminación o brillo de
la muestra “a” indica la zona de variación entre el rojo y el verde del espectro y el parámetro
“b” se refiere a la zona de variación entre el amarillo y el azul del espectro.
Las flores frescas fueron analizadas mediante un colorímetro (Milton Roy (Diano Color
Products de la serie 4FB4186006), el cual se calibró previamente utilizando el área pequeña de
observación y con especular incluido (iluminador estándar D65 Observador 10°), en donde las
flores fueron colocadas de forma frontal en el área de observación, tomando tres lecturas y su
promedio de los parámetros “L, a, b”.
bL1000aIC*×
×= (1)
Debido a que la acumulación de carotenoides está relacionada por la variación de color, el
IC* es una medida válida a utilizar para determinar si hay variación de tonalidades en las flores
obtenidas de las diferentes épocas de transplante (Anexo II).

36
6.5. Evaluación del contenido de xantófilas totales por espectrofotometría UV-Visible
6.5.1. Extracción de carotenoides
Una vez colectadas las flores se pulverizaron para realizar la extracción de los pigmentos. En
matraces de 250 ml se pesaron 0.2 g de tejido fresco y se adicionaron 15 ml de una mezcla de
disolventes compuesta por hexano:etanol:acetona:tolueno (10:6:7:7 v/v/v/v) y se agregaron 2 ml
de KOH al 40% en metanol al 80%. Los matraces se cubrieron con papel aluminio y se incubaron
a 20 °C, a 200 rpm durante 16 horas en oscuridad. La mezcla se transfirió a un matraz
volumétrico de 100 ml y se adicionaron 15 ml de hexano, se agitaron los matraces y se aforaron a
100 ml con Na2SO4 al 10% y se dejaron reposar por una hora. Se separó la fase orgánica y se
pasó por una columna preparada con Na2SO4 anhidro y en la parte terminal se colocó una
membrana de 0.2 µm (Millipore, Molsheim, Francia) (Anexo III). El volumen recuperado se
concentró a sequedad bajo una atmósfera de nitrógeno y las muestras se almacenaron a -70° C
hasta el momento de ser analizadas (modificado de Delgado-Vargas y Paredes-López, 1996).
6.5.2. Cuantificación de xantófilas totales por espectrofotometría UV-Visible
Una vez secas las muestras se resuspendieron en 2.5 ml de acetona para leer las absorbancias
de cada una de las muestras en el espectrofotómetro UV/ Visible de la marca Shimadzu Modelo
UV-16OA previamente calibrado, a una longitud de 474 nm usando acetona como blanco.
Finalmente se reportó el contenido en g de xantófilas por kg de peso fresco (AOAC, 1984).
gMFDAbsfresco peso de Kg totales/ s xantófilade g ××
= (2)
Donde:
Abs: Absorbancia a 474 nm F: Factor de extinción de la luteína
D: Factor de dilución gM: Gramos de la muestra

37
6.6. Identificación y cuantificación de carotenoides por HPLC
Los extractos (oleorresinas) de las flores de los cultivos fueron analizados mediante el uso de
un cromatógrafo líquido de alta resolución (Perkin Elmer de la serie 200) (Anexo IV), equipado
con una bomba Binaria (Perkin Elmer Binary LC pump 250) y un detector UV-Vis con arreglo de
diodos, el programa utilizado fue el Turbochrom Navigator versión 4.1. <2F12> Copyrigh Perkin
Elmer Corp. 1987-1995. La fase estacionaria fue una columna C18 de la marca Supelcosil LC-
18-DB de 5µm con 250 mm de longitud con un diámetro interno de 4.6 mm; para la fase móvil se
emplearon dos tipos disolventes constituyentes:
Disolvente A: Acetato de etilo al 100 %
Disolvente B: Acetonitrilo-Metanol 90:10 (v/v).
La elusión fue en gradiente, teniendo un tiempo total de corrida de 30 min, con un flujo
constante de 1.0 mL/min inyectándose 40 μl de muestra. La detección de los pigmentos se realizó
mediante con un detector de arreglo de diodos con un intervalo de 360 a 800 nm, con un registro
fijo a 450 nm.
La identificación de los pigmentos se realizó considerando los picos de cada uno de los
gráficos y el tiempo de retención de cada pigmento en el cromatograma comparado con un
estándar (luteína), además por las absorciones máximas de los espectros con tablas de absorción
reportados por diferentes autores: Mínguez y Homero, 1993; Razungles y col., 1996; Delgado-
Vargas y Paredes-López, 1997. La cuantificación se realizó integrando el área bajo la curva del
pico correspondiente al pigmento en el cromatograma.

38
7. RESULTADOS Y DISCUSIÓN
7.1. Monitoreo de las condiciones ambientales durante el periodo de los cultivos
La variación de las condiciones ambientales se va dando conforme al paso de los meses. El
trabajo se inició en el mes julio en el cual se presentó la mayor humedad relativa (82.55±1.59%)
y mayor precipitación pluvial (281.45 mm), mientras que la menor humedad relativa (41.4±3.5%)
ocurrió en el mes de marzo, cuando la precipitación pluvial fue nula, las plantas fueron regadas
diariamente por un sistema de tuberías manteniendo una humedad del suelo constante.
Adicionalmente a los cambios en la HR, se observó que 5 meses después de haber comenzado el
experimento en campo, se vieron afectados otros factores como la radiación solar, la cual afecta
directamente la calidad de las plantas (Dorais y col., 2001).
La temperatura también varió durante los meses del experimento, sin embrago solo se
obtuvieron diferencias estadísticamente significativas en las temperaturas mínimas, en los meses
de noviembre, diciembre y enero (Cuadro 3), en los cuales se observó un descenso de hasta
(8.5±1.6 °C). Las temperaturas máximas se presentaron en los meses de febrero y marzo.
El fotoperiodo más largo fue entre los meses de julio a septiembre, que corresponden a la
estación de verano y es la época más lluviosa en el país. A partir de noviembre, el fotoperiodo
disminuyó hasta el mes de enero (Cuadro 3). En este sentido, para muchas plantas que no son
influenciadas por este factor su medición carece de importancia, pero particularmente para el
caso del cempaxúchil si es importante debido a que responde a variaciones en el fotoperiodo
(Serrato-Cruz, 2003).

39
Cuadro 3. Variación de las condiciones ambientales durante los meses (Julio 2007 a Marzo 2008) del experimento en campo, desde el
inicio del primer cultivo hasta la última colecta de inflorescencias.
Condición Ambiental Julio Agosto Septiembre Octubre Noviembre Diciembre Enero Febrero Marzo
Temp. Max. (°C) 29.5±1.4a 28.9±1.9a 28±1.4a 30.4±1.7a 31±2.7ab 28.5±0.8a 27±1.5a 34.6±1.7bc 35.2±1.06c
Temp. Min. (°C) 12.9±1.2a 13.9±0.98a 12.7±0.94ac 11.4±1.02ab 10±1.2bc 9.5±1.8b 8.5±1.6b 11.8±0.7ab 12.8±1.15a
Temp. Prom. (°C) (NS) 19.9±8.9 20.2±8.2 19.1±8.2 19.6±10.2 19.2±11.4 17.7±10.2 16.6±9.9 21.9±12.2 22.6±12.0
HR Prom. (%) 82.55±1.59a 82.03±1.81a 79.12±1.16a 66.76±3.5ab 57.01±4.2ab 53.72±3.1ab 47.81±2.3bd 46.4±2.7b 41.4±3.5b
Rad. Solar (mol/mes.m2) 645.5±101a 684.8±18a 696.6±27a 561.6±15.9a 554.8±38ab 475.6±23b 487.8±8.26b 535.2±23.3ab 628.7±15.7a
Fotoperiodo (h) 13.12±0.12a 12.75±0.56a 12.29±0.34a 12.01±0.14a 10.17±0.16a 9.35±0.01ab 9.1±0.01b 8.96±0.09b 10.45±0.39a
Precip. Acum. (mm)* 281.45 236.4 202.2 41 0 0 0 0 0
Los datos son valores de medias (±desviación estándar), las diferentes letras indican que hay diferencias estadísticamente significativas
de las variaciones ambientales de los diferentes meses de acuerdo con la prueba de Tukey con P<0.001. (NS) No significativo. *Prueba
no realizada.

40
7.2. Evaluación del desarrollo vegetativo de los cultivos
7.2.1. Evaluación de la altura de las plantas
La mayor altura fue alcanzada por las plantas transplantadas a campo en el mes de julio (89.7
± 9.9 cm), seguidas en tamaño por las del mes de agosto (69.3±12.8 cm). Con los cultivos del
mes de octubre a diciembre, ocurrió lo contrario, en donde se observó la menor altura alcanzada
por los cultivos iniciado en diciembre (39 ± 6.2 cm), observando también que la HR que se
presentó durante el experimento en campo fue disminuyendo de 82.55±1.59% hasta 41.4±3.5%
(Cuadro 3), lo cual pudo afectar la elongación celular y por ende el crecimiento. Sugiriendo que
posiblemente, se debió a que uno de los primeros efectos que se observan por la baja humedad
relativa en los vegetales la cual es la elongación celular y por ende el crecimiento. Otra
consecuencia tendría que ver con cierre estomático que puede ocurrir bajo estas condiciones
humedad, el cual trae como consecuencia la disminución de la conductancia que se refleja en una
disminución de la tasa transpiratoria y un menor consumo de agua por la planta, produciendo una
caída en la asimilación de CO2 y diversos efectos asociados tales como la acumulación de poder
reductor, fotoinhibición y/o fotooxidación (Tambussi, 2004).
Estos resultados concuerdan con el experimento que realizó Mortensen (2000), en donde
observó el efecto de la humedad relativa (70, 81 y 93 % de HR) sobre el crecimiento de cuatro
especies de plantas de día corto en invernadero (Begonia, Chrysanthemum, Kalanchoe y
Poinsettia), obteniendo como resultado que a mayor HR, mayor era la altura de las plantas en los
cuatro cultivos, sugiriendo que esto se debe principalmente a la pérdida del control estomatal de
las hojas, ya que las variaciones en la HR afectan el funcionamiento de los estomas
manteniéndolos cerrados o constantemente abiertos, no permitiendo que se dé una adecuada
transpiración.
7.2.2. Evaluación del número de primordios florales
El mayor número de primordios florales se presentó en los cultivos de transplante en
noviembre y diciembre alcanzando 43±8.6 y 50.6±11.5 primordios por planta respectivamente
(Cuadro 4). Esto pudo ser ocasionado por las bajas temperaturas que se registraron durante estos

41
meses, causando en las plantas el efecto llamado vernalización, el cual induce la floración en las
plantas debido a la exposición a bajas temperaturas. (Salisbury y Ross, 1992).
Con relación a lo anterior, ya se ha mencionado que la floración se da en tres etapas:
inducción, evocación y desarrollo y es en la inducción en donde se generan las señales para que
se altere el desarrollo y se oriente hacia la reproducción. En este sentido, se han realizado
estudios en los que la iniciación de los brotes florales, ha sido promovida mediante la incubación
bajo largos periodos a bajas temperaturas, mostrando actividad mitótica (evocación) durante la
exposición a las temperaturas de inducción (Nebauer y col., 2006). En este caso la división
celular es considerada necesaria para la vernalización (Wellensiek, 1964).
Adicionalmente se ha demostrado que en la floración, la inducción también puede ser
afectada por el estrés hídrico, como lo reporta Nir y col., (1972). Por otro parte, también la
reducción en las horas luz puede influir una respuesta en la floración, ya que hay plantas de días
largos, cortos y las neutras las cuales no responden al influencia del fotoperiodo, al parecer el
cempaxúchil, responde a los periodos de días cortos, presentando un mayor número de
primordios florales como respuesta a esto, aunado a un posible efecto sinérgico con las bajas
temperaturas.
7.2.3. Evaluación del diámetro y peso de las inflorescencias
Las flores con mayor diámetro se obtuvieron en los meses julio (5.3±0.5 cm) con la mayor
HR (82.55±1.59%) y en el mes de noviembre (5.2±0.4 cm) donde se registró una baja de HR y la
radiación solar más baja (475.6±23 mol/mes.m2) y aunque no hay diferencia significativa con
respecto a los otros meses de transplante, si se pudo registrar que las flores de octubre tuvieron el
tamaño más pequeño (3.2±0.4 cm de diámetro) (Cuadro 4), en las plantas de los cultivos restantes
las flores fueron más homogéneas en tamaño. Esto probablemente se debió a que la humedad en
el suelo trató de mantenerse constante para asegurar el buen crecimiento y desarrollo de las
plantas. Cabe mencionar que cuando la HR es baja, el control estomático se puede ver afectado y
una excesiva pérdida de agua de las hojas puede no ser suficientemente compensada por el agua
que se encuentra en los tallos, la cual es tomada del suelo (Mortensen y Gislerod, 1999).

42
Con respecto a la intensidad de la luz, Ubaldo (2007) demostró que una disminución del
50% en la radiación fotosintéticamente activa (RFA) dio lugar a que las inflorescencias
presentaran un diámetro más grande, 0.4 veces más que las crecidas a pleno sol (100% de RFA)
concluyendo que una disminución de la RFA con malla favoreció la diferenciación de los
primordios florales.
En el presente trabajo se sugiere que HR que se presenta en los meses de julio a septiembre
favorecería el diámetro de las inflorescencias pero al mismo tiempo la radiación solar disminuye
dicho efecto. Por otra parte, en los meses de noviembre y diciembre HR baja podría afectar el
tamaño de las flores, sin embargo esta situación se compensa con la disminución de la radicación
solar y favorecería el tamaño, lo que hace pensar que puede ocurrir un equilibrio que se ve
reflejado en el diámetro de las flores, pero no así en el peso de las mismas.
Cuadro 4. Evaluación del desarrollo de las plantas a 12 días después del inicio de la floración de
diferentes épocas de transplante a campo.
Desarrollo vegetativo *Julio Agosto Septiembre Octubre Noviembre Diciembre
Altura de las plantas (cm) 89.7±9.9a 69.3±12.8b 49.2±12.2c 46.5±8.8c 48.6±12.1c 39.0±6.2c
No. de Botones/planta 34.7±8.1a 27.8±8ab 21.6±9.4b 20.0±8.4b 43.0±8.6c 50.6±11.5c
Diámetro de la Inflorescencia (cm) 5.3±0.5a 4.9±0.6a 4.9±0.2a 3.2±0.4b 5.2±0.4a 4.8±0.3a
Días hasta el inicio de la floración 39 44 46 47 56 72
Los datos son valores de medias (± desviación estándar). Las letras diferentes indican diferencias
significativas en el desarrollo vegetativo en los diferentes tiempos de siembra, de acuerdo con la
prueba de Tukey para una P≤0.001. (* Mes de siembra del cempaxúchil para motivos religiosos).
El mayor peso fresco (pf) y peso seco (ps) de las inflorescencias se obtuvo en las plantas
sembradas en los meses de julio y agosto con 7.5±1.5 g de pf y 2.14±0.42 g de ps (Figura 9). La
disminución en el peso de las flores estuvo de acuerdo con una reducción en la HR y RFA.
Dorais y col., (2001), reportaron que estos factores ejercen una influencia directa sobre la calidad
de las plantas, flores y los frutos: en su apariencia, firmeza, textura, materia seca y propiedades

43
sensoriales. En este caso, los cambios más dramáticos están relacionados con la HR la cual
disminuyó a lo largo del experimento.
Tomando en cuenta que el cempaxúchil es una planta C3, estas son consideradas menos
eficientes para la fijación de CO2 y se caracterizan por consumir grandes cantidades de agua y
requerir de una alta HR (más del 60%) para que puedan desarrollar su máximo potencial
(Vásquez-García y col., 2002).
Figura 9. Variación del peso fresco y peso seco del las flores de diferentes épocas de transplante
a campo. En este cuadro las diferentes letras indican diferencias estadísticamente significativas en
el peso de las flores de acuerdo con la prueba de Tukey (P<0.05). *Mes de siembra por motivos
religiosos.
Con respecto al inicio de la floración, las más tardías en florear fueron las del mes de
diciembre (cuadro 5), esto pudo deberse a que las plantas de este cultivo crecieron con la menor
intensidad luminosa, con menor fotoperíodo y que cuando comenzó la floración, estas estaban
expuestas a la menor humedad relativa del ambiente, pero principalmente a las temperatura más
bajas, sugiriendo que desarrollo de la planta fuera retrasándose, dándose su tiempo para florear
(Keeve y col., 2000).

44
7.3. Evaluación del índice de color de inflorescencias de cempaxúchil por CIELAB.
Con respecto al color, en las flores del cultivo transplantado en el mes de noviembre se
observó un amarillo más intenso de acuerdo con los valores del índice de color de IC*=1.98
(Cuadro 5) y que estuvieron en un intervalo de entre -2 a +2, lo cual las ubica en colores que van
desde el amarillo intenso al amarillo verdoso. Las flores procedentes de las fechas de siembra
restantes presentaron valores mayores de 2 lo que indica, de acuerdo con la nomenclatura del
índice de color (2-20), que los colores van desde el amarillo pálido al naranja intenso.
Cuadro 5. Valores del IC* de las inflorescencias de cempaxúchil de diferentes época de
transplante a campo.
Mes de transplante IC*
Julio 3.53
Agosto 5.03
Septiembre 4.09
Octubre 5.01
Noviembre 1.98
Diciembre 3.81
IC*=Índice de color
Lo anterior podría estar relacionado con el hecho de que cuando la radiación solar es alta, el
contenido de pigmentos es más bajo, por lo que las condiciones ambientales especialmente la luz
influencian la biosíntesis de los pigmentos y su formación son es controlada por factores externos
e internos que van a determinar su biosíntesis y degradación (Dymova y Golovko, 2007).
7.4. Evaluación del contenido de xantófilas totales por espectrofotometría UV-Visible
Se observó que las inflorescencias de los cultivos del mes de noviembre y diciembre
presentaron una mayor concentración de xantófilas totales con 13.6 y 8.2 g/kg de peso fresco
respectivamente (Figura 10).

45
Dorais y col. (2001) reportaron que la producción de estos compuestos se ve afectada
principalmente por la luz. En este trabajo, cuando las radiaciones solares fueron menores en los
meses de noviembre y diciembre (575 y 475 mol/m2.mes) se favoreció la acumulación de estos
compuestos y por el contrario en los meses de mayor radiación solar el contenido fue menor
debido posiblemente a que ésta puede estar dañando a los pigmentos oxidándolos y
transformándolos en grupos epóxido. Otra explicación es, que esta radiación probablemente
pudiera estar afectando directamente la expresión de los genes carotenogénicos (psy, pds y lcy-b)
en la ruta de biosíntesis (Bartley y Scolnik, 1995).
Por otra lado, el fotoperiodo es otro factor que presumiblemente puede estar afectando a la
concentración de dichos pigmentos. Debido a que las plantas de cempaxúchil son de fotoperiodo
corto, estas pueden ser afectadas cuando se exponen a periodos de luz más largos, como se
observa en los meses de julio a octubre obteniendo los rendimientos más bajos, sugiriendo que a
mayor tiempo de exposición de las flores a la luz solar los pigmentos pueden ser dañados.
Figura 10. Variación de la acumulación de xantófilas totales para las diferentes épocas de
transplante. Las letras diferentes indican diferencias estadísticamente significativas de acuerdo
con Tukey para una (P≤0.05). *Mes de siembra por motivos religiosos.
Los resultados mostrados (Figura 10) son similares a lo que reporta Ubaldo (2007), con
relación a que una disminución de la radiación favorece la acumulación de pigmentos

46
carotenoides, la cual fue mayor (4.39 ±1.25 g/kg de peso fresco) en plantas crecidas bajo una
malla con un RFA menor (50% de luz solar), que las crecidas a pleno luz solar (3.013±1.48 g/kg
por kilogramo de peso fresco) con un RFA mayor (100% de luz solar), sugiriendo que la malla
filtra los rayos UV reduciendo el daño por oxidación de los pigmentos.
7.5. Cuantificación de luteína por HPLC
De acuerdo con los resultados de los extractos completamente saponificados de
inflorescencias de cempaxúchil de las diferentes épocas de transplante (julio-diciembre de 2007),
estos exhibieron espectros de absorción, obtenidos por HPLC, con un perfil típico de xantófilas
al ser comparado con un estándar de luteína (Figura 11).
Figura 11. Perfil cromatográfico del HPLC del estándar de luteína.
Los cromatogramas de los extractos de las inflorescencias de cempaxúchil del cultivo de
julio (Figura 12) presentaron un perfil típico de xantófilas, con un pico principal a los 4.72 min el
cual correspondió a luteína comparado con el estándar correspondiente, también se observaron
otros picos adyacentes al principal, los cuales posiblemente sean esteres e isómeros de luteína
(Scott y Eldridge, 2004; Navarrete-Bolaños y col., 2005; Ubaldo, 2007).
Luteína

47
Figura 12. Perfil cromatográfico del HPLC obtenido de los extractos de las flores de
cempaxúchil del cultivo del mes de Julio. Los números indican los posibles carotenoides
presentes en el cromatograma: (1) luteína, (2) isómero de luteína, (3) esteres.
El comportamiento del perfil cromatográfico de los extractos de las flores en los cultivos
trasplantados en el mes de agosto y hasta el mes de diciembre (Anexos V al IX) fue muy similar
al del mes de julio (Figura 12) y al estándar de luteína (Figura 11), indicando que no hay
variación entre los pigmentos obtenidos de las inflorescencias de las diferentes épocas de
trasplante. Esta similitud en los cromatogramas podría estar indicando que efectivamente las
inflorescencias de cempaxúchil contienen células especializadas acumuladoras de xantófilas,
principalmente de luteína. Por otro lado, debido a que las condiciones ambientales presentaron
una variación estadísticamente significativa para los cultivos transplantados en los meses de
noviembre y diciembre, se pudo inferir que como respuesta a esto las flores acumularon un
mayor contenido de luteína (Figura 13).
El contenido de luteína en las lígulas de las inflorescencias de los cultivos de las diferentes
épocas de transplante osciló entre los 2882±247 y los 6105±417 μg/g de tejido fresco (Figura 13),
cuantificado a partir de los cromatogramas obtenidos y mediante una curva de calibración para
luteína (Anexo X); estos datos corresponden con lo reportado por Del Villar-Martínez (2003),
con relación a que los niveles de luteína encontrados en flores cultivadas de cuatro variedades
diferentes de cempaxúchil oscilaron entre los 400 y 6000 μg de luteína por gramo de tejido
fresco.
(1)
(2)(3)
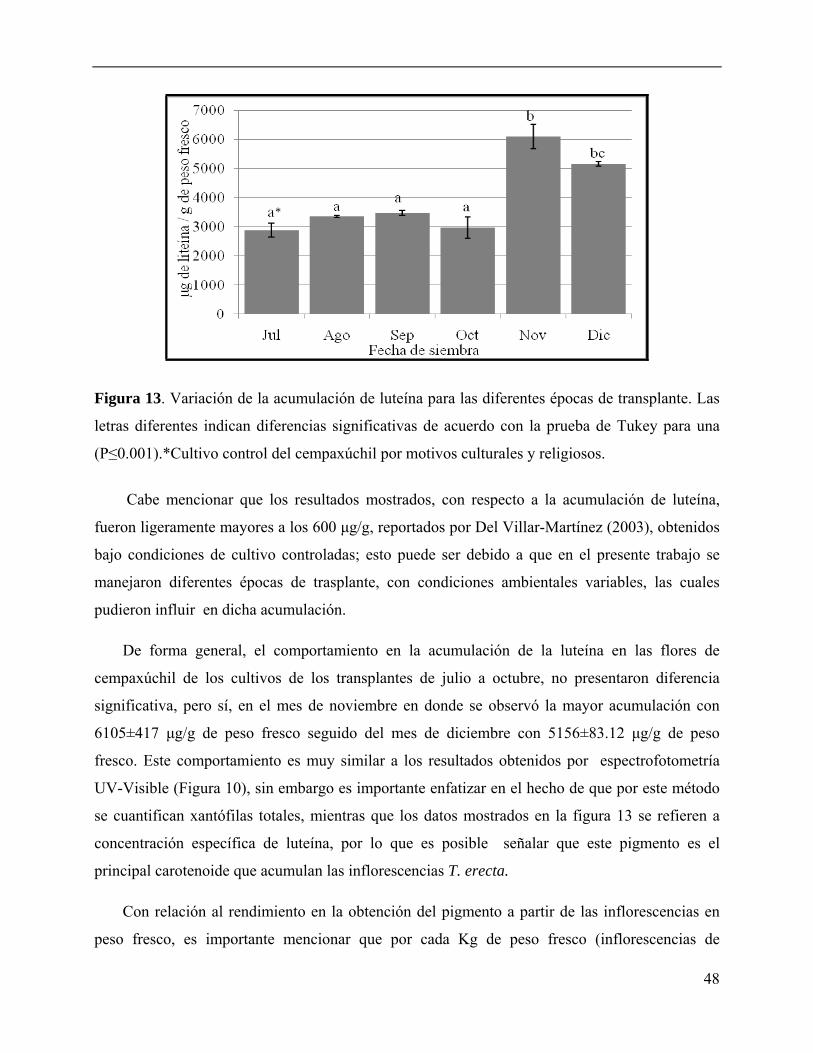
48
Figura 13. Variación de la acumulación de luteína para las diferentes épocas de transplante. Las
letras diferentes indican diferencias significativas de acuerdo con la prueba de Tukey para una
(P≤0.001).*Cultivo control del cempaxúchil por motivos culturales y religiosos.
Cabe mencionar que los resultados mostrados, con respecto a la acumulación de luteína,
fueron ligeramente mayores a los 600 μg/g, reportados por Del Villar-Martínez (2003), obtenidos
bajo condiciones de cultivo controladas; esto puede ser debido a que en el presente trabajo se
manejaron diferentes épocas de trasplante, con condiciones ambientales variables, las cuales
pudieron influir en dicha acumulación.
De forma general, el comportamiento en la acumulación de la luteína en las flores de
cempaxúchil de los cultivos de los transplantes de julio a octubre, no presentaron diferencia
significativa, pero sí, en el mes de noviembre en donde se observó la mayor acumulación con
6105±417 μg/g de peso fresco seguido del mes de diciembre con 5156±83.12 μg/g de peso
fresco. Este comportamiento es muy similar a los resultados obtenidos por espectrofotometría
UV-Visible (Figura 10), sin embargo es importante enfatizar en el hecho de que por este método
se cuantifican xantófilas totales, mientras que los datos mostrados en la figura 13 se refieren a
concentración específica de luteína, por lo que es posible señalar que este pigmento es el
principal carotenoide que acumulan las inflorescencias T. erecta.
Con relación al rendimiento en la obtención del pigmento a partir de las inflorescencias en
peso fresco, es importante mencionar que por cada Kg de peso fresco (inflorescencias de

49
cempaxúchil), se recuperó el 10% como extracto seco y de éste, el 1.07% son xantofilas totales.
Así mismo, el rendimiento como luteína es de 0.65% a partir del 1.07% de xantofilas.
Observado en paralelo los resultados del IC*, se vio que la acumulación de carotenoides por
UV-Vis y de luteína por HPLC, fue mayor en los cultivos transplantados en el mes de noviembre,
mientras que el IC* de estas muestras estuvo dentro de un intervalo de -2 a +2 , el cual indica que
los colores de estas flores se encontraban entre el amarillo intenso al verdoso. En los cultivos
transplantados en otras épocas, los cuales mostraron un contenido menor de carotenoides, el IC*
estuvo en un intervalo entre +2 a +20, lo que corresponde a colores que van desde el amarillo
pálido al naranja intenso.
7.6. Rendimientos en la producción de flores de cempaxúchil.
El mayor rendimiento en la producción de flores de cempaxúchil se observó en el cultivo del
mes de diciembre, con una producción de 13.2 ton/ha (Cuadro 6), seguidos de los cultivos de
transplante en el mes de julio, agosto y noviembre. Estos resultados al ser comparados con la
producción de cempaxúchil a nivel nacional (10.9 ton/ha) en el años 2003 (Cuadro 1), los
rendimientos fueron ligeramente superiores con hasta 2.3 ton/ha. Por otro lado al compararlos
con los rendimientos de la producción de cempaxúchil en el estado de Morelos, la cual alcanzó 9
ton/ha en el mismo tiempo (Cuadro 2), los resultados fueron superiores con 3.3 ton/ha (Fuente
SIACON SAGARPA, 2006). Esto se sugiere que se debió probablemente a que los cultivos
crecieron con las temperaturas mínimas registradas, aunado a que la radiación solar y el
fotoperiodo también se habían reducido, provocaron una respuesta en las plantas, dando como
resultado un mayor número de primordios florales y como consecuencia un mayor número de
inflorescencias por planta (Cuadro 6).
El cultivo transplantado en el mes de octubre presentó los rendimientos más bajos en
producción de inflorescencias, esto podría deberse principalmente a que a estas plantas en
particular, crecieron en la transición entre una época lluviosa y la sequía, provocando estrés
hídrico que se reflejó en la altura, el numero de primordios florales, la acumulación de xantófilas
y luteína y en el rendimiento en la producción de flores.

50
Cuadro 6. Rendimientos en la producción de inflorescencias de cempaxúchil de seis diferentes
épocas de transplante a campo.
Época de
transplante
No. de inflorescencias
por planta
Rendimientos
(Ton/ha)
Julio* 20.7±3.4a 12.3
Agosto 22.2±3.9a 10.7
Septiembre 19.4±3.4a 8.5
Octubre 16.9±2.3b 5.8
Noviembre 33.2±5.5c 10.6
Diciembre 37.3±8.5c 13.2
Las letras diferentes indican diferencias estadísticamente significativas de acuerdo con Tukey
para una (P≤0.05). *Mes de siembra por motivos religiosos.
La producción de cempaxúchil en el país está asociada a las festividades religiosas,
principalmente al día de muertos, por lo que el cultivo se inicia a finales del mes de Julio y
principios de Agosto; en este tiempo la temporada de lluvias en México ha iniciado, provocando
las condiciones ambientales en temperatura, humedad y luz para su producción. El estado de
Morelos, es uno de los estados que goza de condiciones ambientales estables, lo que trae como
consecuencia que sea un lugar adecuado para la producción de cultivos como el cempaxúchil,. De
acuerdo con los resultados obtenidos su producción en este estado podría darse, no solo en la
temporada destinada a la festividad mencionada, sino que puede ser cultivado en otra época, lo
cual repercute en la producción de pigmentos de alto valor comercial, a lo largo de la mayor
parte del año y por ende en el bienestar en los campesinos sus familias.

51
8. CONCLUSIONES
El cultivo transplantado en el mes de julio presentó mayor altura, diámetro de las flores, peso
fresco y seco de las inflorescencias. Adicionalmente, el número de primordios florales fue
importante en los transplantes de los meses de noviembre y diciembre. Siendo este ultimo el
cultivo mas tardado en florear.
El análisis del IC* de las inflorescencias de cempaxúchil de diferentes épocas de transplante,
mostraron diferencias en cuanto a la variación del color, el cual se observó desde el amarillo
intenso al amarillo pálido. Las inflorescencias de las plantas del mes de noviembre presentaron
un color amarillo intenso y esto coincidió con una mayor acumulación de xantofilas.
Las inflorescencias provenientes de los cultivos del mes de noviembre, mostraron el mayor
contenido de xantófilas totales, posiblemente esta acumulación fue favorecida por las bajas
radiaciones solares y temperaturas registradas en esta época.
Las xantófilas identificadas fueron luteína e isómeros de luteína, mientras que solo fue
cuantificada la luteína, la cual fue la de mayor acumulación en las inflorescencias cultivadas en
los meses de noviembre y diciembre.
En general los cultivos trasplantados en los meses de noviembre y diciembre proporcionaron
los mejores resultados en acumulación de carotenoides y los mejores rendimientos en producción
de flores de cempaxúchil. El mes de julio proporcionó el mejor desarrollo vegetativo.

52
9. PERSPECTIVAS
Continuar con los estudios acerca de las variaciones ocasionadas por la fecha de transplante a
campo del cempaxúchil T.erecta, principalmente en el estado de Morelos, en los meses que no se
incluyeron en el presente trabajo, ya que en los meses de abril y mayo se presentan las
condiciones ambientales con mas variación durante el año, principalmente en cuanto a la
intensidad luminosa y el fotoperiodo.

53
10. BIBLIOGRAFÍA
Abdul C., Gopi R., Manivannan P y Panneerselvam R. 2007. Responses of antioxidant defense
system of Catharanthus roseus (L.) G. Don to paclobutrazol treatment under salinity. Acta
Physiologiae Plantarum 29: 205-209.
Adams WW y Demmig-Adams B. 1994. Carotenoid composition and down regulation of
photosystem II in three conifer species during the winter. Physiologia Plantarum 92: 451-
458.
Adamska I. 199. ELIPs-light-induced stress proteins. Physiology Plantarum 100: 794-805.
Alvarado ZA. 2004. Cempoalxóchitl, la flor de los veinte pétalos. Correo del Maestro. 102. [En
línea] Disponible: http://www.correodelmaestro.com, Noviembre de 2004.
AOAC. 1984. Official Methods of the Association of Official Analytical Chemists (14th Ed).
Washington, DC.
Arévalo J. 1994. Efecto del manejo de raíces y tallo en plantas de Eucalyptus globulus Labill en
vivero, en la resistencia a bajas temperaturas. Tesis de Grado. Facultad de Ciencias
Forestales, Universidad de Concepción, Concepción, Chile. 56 p.
Aro EM., I Virgin y B Anderson. 1993. Photoinhibition of photosystem II. Inactivation, protein
damage and turnover. Biochimica et Biophysica Acta 1143: 113-134.
Baker NR. 1991. A possible role for photosystem II in environmental perturbations of
photosynthesis. Physiologia Plantarum 81: 563-570.
Barceló J., Nicolas G., Sabater B y Sanchez R. 1995. Fisiología Vegetal, Ciencia y Técnica.
Editorial Pirámide, S.A., Madrid, España. 566 p.
Bartley GE y Scolnick PA. 1995. Plant carotenoids: Pigments for photoprotection, visual
attraction, and human health. The Plant Cell 7:1027-1038.

54
Bendich A. 1993. Biological functions of dietary carotenoids. Annual of the New York Academy
of Sciences 691 (1): 61-67.
Benk E., Trieber H y Bergmann R. 1976. Detection taw materials and beverages. Riechst Aromen
and Koerpen 26(10): 216-221.
Breithaupt DE y Bamedi A. 2001. Carotenoid esters in vegetables and fruits: a screening with
emphasis on β-cryptoxanthin esters. Journal of Agricultural Food Chemistry 49(4): 2064-
2070.
Britton G. 1983. The Biochemistry of Natural Pigments. Cambridge, United Kingdom:
Cambridge University Press. 366 p
Buchanan BB., Gruissem W y Jones, RL. 2000. Membrane structure and inheritance of cell
membranas. En: Biochemistry Molecular Biology of Plants. Ed. American Society of Plant
Physiologist. Rockville, Maryland.
Bungard RA., Ruban AV., Hibberd JM., Press MC., Horton P y Scholes JD. 1999. Unusual
carotenoid composition and new type of xanthophyll cycle in plants. Process Natural
Academic Science 96:1135-1139.
Carmona M., Zalacain A., Sánchez AM., Novella JL y Alonso GL. 2006. Crocetin esters,
picrocrocin and its related compounds present in Crocus sativus stigmas and Gardenia
jasminoides fruits. Tentative identification of seven new compounds by LCESI-MS.
Journal of Agricultural Food Chemistry 54: 973-979.
Chappell J. 1995. The biochemistry and molecular biology of isoprenoid metabolism. Plant
Physiology 107:1-6.
Chenard CH. 2005. Nitrogen concentration affects nutrient and carotenoid accumulation in
parsley. Journal of Plant Nutrition 28: 285–297.

55
Cunningham FX., Pogson B., Sun Z., McDonald K., DellaPenna D y Gantt E. 1996. Functional
analysis of the β- and ε- lycopene cyclase enzymes of Arabidopsis reveals a mechanism for
control of cyclic carotenoid formation. The Plant Cell 8:1613-1626.
Cunningham FX y Gantt E 2001. One ring or two? Determinating of ring number in carotenoids
by licopene ε-cyclases. Proceedings of the National Academy of Sciences USA 98:2905-
2910.
de Azevedo CH y Rodriguez-Amaya DB. 2005. Carotenoid composition of kale as influenced by
maturity, season and minimal processing. Journal of the Science of Food and
Agriculture.85: 591-597.
De La Cruz TE., M. Rubí A y T. Falcón B. 1994. pp. 101-112. Evaluación del desarrollo de
plantas de aguacate cv Hass sometidas a radiación gamma de 60Co. En: Memoria de la
Fundación Salvador Sánchez Colín CICTAMEX, S.C.
DellaPenna, D. 1999. Nutritional Genomics: Manipulating Plant Micronutrients to Improve
Human Health. Science 285 (5426):375-379
Del Villar-Martínez AA. 2003. Caracterización molecular y expresión in vitro de las β y ε-
licopeno ciclasas de cempaxúchil (Tagetes erecta). Tesis Doctoral. Centro de
investigaciones de estudios avanzados del IPN. Irapuato, Gto.
Delgado-Vargas F y Paredes-López O. 1996. Correlation of HPLC and AOAC methods to assess
the all-trans-lutein content in marigold flowers. Journal of the Science of Food and
Agriculture 72:283-290.
Delgado VF. 1997. Pigmentos de flor de cempasúchil (Tagetes erecta) Caracterización
fisicoquímica, procesamiento y eficiencia pigmentante. Tesis doctoral. Centro de
Investigación y de Estudios Avanzados del Instituto Politécnico Nacional. Irapuato,
Guanajuato.
Delgado-Vargas F y Paredes-López O. 1997. Enzymatic treatment to enhance carotenoid content
in dehydrated marigold flower meal. Plant Foods for Human Nutrition. 50: 163-169.

56
Delgado-Vargas F., Jiménez A R y Paredes-López O. 2000. Natural pigments: carotenoids,
anthocyanins, and betalains - characteristics, biosynthesis, processing, and stability. Critical
Reviews in Food Science and Nutrition 40:173-289.
Delgado-Vargas, F y Paredes-López O. 2003. pp: 257– 305. Chemicals and colorants as
nutraceuticals. En: Natural Colorants for Food and Nutraceuticals Uses. CRC. Press. Boca
Raton, Florida U.S.A.
Demmig-Adams B y Adams WW. 1992. Photoprotection and other responses of plants to high
light stress. Annual Review of Plant Physiology and Plant Molecular Biology 43: 599-626.
Demmig-Adams B y Adams WW. 1996b. The role of xanthophyll cycle carotenoids in the
protection of photosynthesis (Review). Trends in Plant Science 1(1): 21-26.
Dorais M., Papadopoulos AP y Gosselin A. 2001. Greenhouse tomato fruit quality. Horticultural
Reviews. 26: 239-319.
Dumas Y., Dadomo M., Di Lucca G y Grolier P. 2003. Effects of environmental factors and
agricultural techniques on antioxidants content of tomatoes. Journal of the Science of Food
and Agriculture 83: 369-382.
Dymova O y Golovko T. 2007. Pigment apparatus in Ajuga reptans plants as affected by
adaptation to light growth conditions Russian Journal of Plant Physiology Volume 54(1):
39-45
Eskling M., Arvidsson PO y Akerlund HE. 1997. The xanthophyll cycle, its regulation and
components. Physiologia Plantarum 100: 806-816.
Evangelista-Lozano S., Manzanares CY, Escobar AS y Ventura-Zapata E. 2005. Propagación de
tres cultivares de Bougainvillea glabra Choise mediante esquejes en diferentes sustratos.
Proceding of the Interamerican Society of Tropical Horticulture 48: 161-163.
Fraser PD y Bramley P. 2004. The biosynthesis and nutritional uses of carotenoids. Progress in
Lipid Research 43(3): 228-265.

57
Gandul RB y Mínguez MM. 1996. Chlorophyll and carotenoids composition in virgin olive oils
from various spanish olive varieties. Journal of the Science of Food and Agriculture 72: 31-
39.
Goodwin TW. 1980. The biochemistry of the carotenoids. Volume I. Plants. London, United
Kingdom: Chapman and Hall.
Goodwin TW. 1984. The biochemistry of the carotenoids. Volume II. Animals. London, United
Kingdom: Chapman and Hall.
Gross J., Gabai M., Lifshitz A y Sklarz B. 1973. Carotenoids of Eriobotrya japonica.
Phytochemistry 12: 1775-1782.
Gruszecki WI., Strzalka K., Bader KP., Radunz A y Schmid, G.H. 1996. Involvement of the
xanthophyll cycle in regulation of cyclic electron flow around photosystem II. Zeitschrift
fur Naturforschung. Section C, Biosciences 51 (1-2): 47-52.
Guevara E y Jiménez V. 2006. La reproducción de las plantas. Editorial Universidad de Costa
Rica, 82 p. Serie: Fisiología de la Producción de los Cultivos Tropicales (Costa Rica) v. 5.
San José (Costa Rica).
Havaux M., Greppin H y Strasser RT. 1991. Functioning of photosystems I and II in pea leaves
exposed to heat stress in the presence or absence of light. Analysis using in vivo
fluorescence, absorbance, oxygen and photoacustic measurements. Planta 186: 88-98.
Havaux M y Gruszecki WI. 1993. Heat-induced and light-induced chlorophyll-a fluorescence
changes in potato leaves containing high or low levels of the carotenoid zeaxanthin -
Indications of a regulatory effect of zeaxanthin on thylakoid membrane fluidity.
Photochemistry and Photobiology 58: 607-614.
Hess D. 1975. Plant Physiology: Molecular, Biochemical and Physiological Fundamentals of
Metabolism and Development. pp: 219-220. Springer-Verlag, New York.

58
Hochmuth GJ., Brecht J K y Bassett M. 1999. Nitrogen fertilization to maximize carrot yield and
quality on a sandy soil. HortScience 34: 641–645.
Hopkins WG. 1999. Introduction to Plant Physiology. pp. 415-435. 2nd ed., John Wiley and
Sons, New York.
Howard LR., Talcott ST., Brenes CH y Villalon B. 2000. Changes in phytochemical and
antioxidant activity of selected pepper cultivars (Capsicum species) as influenced by
maturity. Journal of Agricultural. Food Chemistry 48: 1713-1720.
Ishida BK., Turner C., Chapman MH y McKeon TA. 2004. Fatty acid and carotenoid
composition of gac (Momordica cochinchinensis Spreng) fruit. Journal of Agricutural Food
Chemistry 52: 274-279.
Karrer P y Jucket E. 1948. Carotenoids. Bassel, Switzerland: Birkhäuser
Keeve R., Loubser HL y Krüger GHJ. 2000. Effects of temperature and photoperiod on days to
flowering, yield and yield components of Lupinus albus (L.) under field conditions. Journal
of Agronomy and Crop Science 184 (3): 187-196.
Kopas-Lane LM y Warthesen JJ. 1995. Carotenoid photostability in raw spinach and carrots
during cold storage. Journal of Food Science 60: 773–776.
Kopsell DA., Kopsell DE y Lefsrud MG. 2004. Variation in lutein, β-carotene, and chlorophyll
concentrations among Brassica oleracea cultigens and seasons. HortScience 39: 361-364.
Kopsell DA y Kopsell DE. 2006. Accumulation and bioavailability of dietary carotenoids in
vegetable crops. Trends in Plants Science 11(10): 1360-1385.
Korneev DY., Logan BA y Kholadei, AS. 2005. Estimation of the rate of photosystem II
reparation under field conditions. Fiziol. Biokh. Kul’t. Rast. 37:17-22.
Krause GH y Weis E. 1991. Chlorophyll fluorescence and photosynthesis: the basics. Annual
Review of Plant Physiology and Plant Molecular Biology 42: 313-349.

59
Laule O., Fürholz A., Chang HS., Zhu T., Wang X., Heifetz, PB., Gruissem W y Lange BM.
2003. Crosstalk between cytosolic and plastidial pathways of isoprenoid biosynthesis in
Arabidopsis thaliana. Proceedings of the National Academy of Sciences USA 100: 6866-
6871.
Lefsrud MG., Kopsell DA., Auge RM y Both AJ. 2006a. Biomass production and pigment
accumulation in kale grown under increasing photoperiods. HortScience 41: 603-606.
Lefsrud MG., Kopsell DA., Kopsell DE y Curran-Celentano J. 2006b. Irradiance levels affect
growth parameters and carotenoid pigments in kale and spinach grown in a controlled
environment. Physiologia Plantarum 127: 624–631.
Levy J., Karas M., Amir H, y cols. 1996. Lycopene inhibits cancer cell proliferation by delaying
cell cycle progression in G1 phase. pp 93. Abstracts of 11th International Symposium on
Carotenoids. Leiden, Netherlands.
Lichtenthaler H. 1999. The 1-deoxy-D-xilulose-5-phosphate pathway of isoprenoids biosynthesis
in plants. Annual Review of Plant Physiology and Plant Molecular Biology. 50: 47-65.
Long SP., Humphries S y Falkowski PG. 1994. Photoinhibition of photosynthesis in nature.
Annual Review of Plant Physiology and Plant Molecular Biology 45: 633-662.
Manrique Reol E. 2003. Los pigmentos fotosintéticos, algo más que la captación de luz para la
fotosíntesis Ecosistemas 12 (1). 235-245. [En línea] Disponible:
http://www.revistaecosistemas.net/articulo.asp?Id=235.
Manzanero BH, Flores P y Rivera R. 2002. Cempasúchil: fuente importante de carotenoides.
Revista ciencia y desarrollo 165 (28): 20-35.
Marin E., Nussaume L., Quesada A., Gonneau M., Sotta B., Hugueney P., Frey A y Marionpoll
A. 1996. Molecular identification of zeaxanthin epoxidase of Nicotiana plumbaginifolia, a
gene involved in abscisic acid biosynthesis and corresponding to the abalocus of
Arabidopsis thaliana. The EMBO Journal 15 (10): 2331-2342

60
Martínez M. 1979. Catálogo de nombres vulgares y científicos de plantas mexicanas. Fondo de
Cultura Económica. México, D.F.
Martínez-Peña M., Cortés-Cuevas A. 2004. Evaluación de tres niveles de pigmento de flor de
cempasúchil (Tagetes erecta) sobre la pigmentación de la piel de pollos de engorda.
Técnica Pecuaria en México 42(1):105-111.
Meléndez-Martínez A., Vicario IM., Heredia FJ. 2007. Pigmentos carotenoides: Consideraciones
estructurales y fisicoquímicas. Archivos Latinoamericanos de Nutrición 57(2): 109-117.
Mínguez M y Homero D. 1993. Separation and quantification of the carotenoid pigments in red
pepper (Capsicum annum L.), paprika, and oleoresin by reversed-phase HPLC. Journal
Agricultural Food Chemistry. 41 (10): 1616-1620.
Mondragón PJ., Tenorio LP y Vibrans H. 2007. Malezas de México. Ficha informativa. [En
línea] Disponible: http://www.conabio.gob.mx/malezasdemexico/asteraceae/tagetes-
erecta/fichas/pagina1.htm, 18 de junio de 2007.
Morris LG. 1957. Some aspects of control of plant environment 11. The control of temperature
and humidity in artificially illuminated rooms. Journal of Agricultural Engineering
Research 2 (1): 30- 43.
Mortensen LM y Gislerod HR. 1999. Influency of air humidity and lighting period on growth,
vase life and water relation of 14 rose cultivars. Scientia Horticulturae 82: 289-259.
Mortensen LM. 2000. Effect of air humidity on growth, flowering, keeping quality and water
relations of four short-day greenhouse species. Science Horticulturae. 86: 299-3100.
Navarrete-Bolaños JL., Rangel-Cruz CL., Jiménez-Islas H., Botello-Álvarez E y Rico-Martínez
R. 2005. Pre-treatment effects on the extraction efficiency of xanthophylls from marigold
flower (Tagetes erecta) using hexane. Food Research International. 38: 159-165.
Nebauer SG., Avila C., García-Luis A y Guardiola JL 2006. Seasonal variation in the competence
of the buds of three cultivars from different Citrus species to flower. Trees 20:507-514.

61
Nir I., Goren R y Leshem B. 1972. Effect Of water stress, gibberellic acid and 2-chloroethyl
thryethilammonium chloride (CCC) on flower differentiation in Eureka lemon. Journal of
the American Society for Horticultural Science 97: 774-778.
Ormrood DP. 1978. Temperature pp. 45-56. En: R. W. Langhans (ed), A growth chamber
manual. Enviromnental control for plants. Cornell Univ.Press, London.
Osmond CB. 1994. What is photoinhibition? Some insights from comparisons of shade and sun
plants. En: Baker NR y Bowyer JR (eds) Photoinhibition of Photosynthesis: from
Molecular Mechanisms to the Field: 1-24. BIOS Scientific Publishers Limited, Oxford,
United Kingdom.
Pecker I., Gabbay R., Cunningham FX, Hirshberg J. 1996. Cloning and characterization of the
cDNA for lycopene beta cyclase from tomato reveals decrease in its expression during fruit
ripening. Plant Physiology 104:227-234.
Philip T. 1973. The nature of carotenoid esterification in citrus fruits. Journal of Agricultural
Food Chemistry 21(6): 964-966.
Pogson B., McDonald K., Truong M., Britton G y DellaPenna D. 1996. Arabidopsis carotenoid
mutants demonstrate that lutein is not essential for photosynthesis in higher plants. The
Plant Cell 8:1627-1639.
Pott I., Breithaupt DE y Carle R. 2003. Detection of unusual carotenoid esters in fresh mango
(Mangifera indica L. cv. ‘Kent’). Phytochemistry 64: 825-829.
Putman DH., Simmons SR y Hardman LL. 1993. Vernalization and seeding date effects on yield
and yield components of white lupin. Crop Science 33: 1076-1083.
Raven PH y Curtis H. 1975. Biología Vegetal. Ed. Omega. Barcelona.
Razungles A., Babic I., Sapis J y Bayonove C. 1996. Particular behavior of epoxy xanthophylls
during veraison and maturation of grape. Journal of Agricultural and Food Chemistry 44
(12): 3821-3825.

62
Rodríguez-Amaya, DB. 1999. Changes in carotenoids during processing and storage of foods.
Archivos Latinoamericanos de Nutición 49: 38S–47S.
Rodríguez-Amaya DB. 2001. A guide to carotenoid analysis in food. pp. 10. Ed ILSI Press.
Washington, USA.
Ronen G., Cohen M., Zamir D y Hirschberg J. 1999. Regulation of carotenoid biosynthesis
during tomato fruit development: expression of the gene for lycopene epsilon-cyclase is
down-regulated during ripening and is elevated in the mutant Delta. The Plant Journal
17:341-351.
Russo VM y Howard LR. 2002. Carotenoids in pungent and non-pungent peppers at various
developmental stages grown in the field and glasshouse. Journal of the Science of Food and
Agriculture. 82: 615–624.
Rzedowski J y Rzedowski GC. 2005. Flora Fanerogámica del Valle de México. Vol. II 1ª.
Edición. Escuela Nacional de Ciencias Biológicas, IPN e Instituto de Biología. México D.
F.
Salisbury F y Ross C. 1992. Plant Physiology. 4th edition. Wadsworth Publishing, California,
United States. 682 p.
Sánchez-Olate M., Zapata J., Ríos D., Pereira G y Escobar R. 2003. Efecto del fotoperíodo en el
desarrollo de plantas de Eucalyptus globulus Labill. ssp. globulus cultivadas en vivero.
Bosque (Valdivia) 24 (2): 23-28.
Sánchez-Millán JL., Arreguín-Espinosa R., Barradas VL., Anaya-Lang AL y García-Jiménez FA.
2006. Efecto de la Radiación Fotosintéticamente Activa (RFA) en carotenoides en
condiciones in Vitro e in Vivo. XXVI Congreso Nacional de la Sociedad Mexicana de
Bioquímica. Guanajuato, México.
Scott CE y Eldridge AL. 2004. Comparation of carotenoid content in fresh, frozen and canned
corn. Journal of Food Composition and Analysis. 18: 551-559.

63
Searle L y Coupland G. 2004. Induction of flowering by seasonal changes in photoperiod. The
Embo Journal 23: 1227-1222.
Serrato-Cruz MA. 2003. Estado actual del conocimiento sobre Tagetes como ornamental. pp:
189-198. En: Mejía M., José M y Espinosa FA. (Ed). Plantas Nativas de México con
Potencial Ornamental. Universidad Autónoma de Chapingo – Departamento de Fitotecnia.
México, DF.
Serrato-Cruz MA. 2004. Cempoalxochitl. Ciencia y Desarrollo en Internet. 2004/1. [En línea]
Disponible:
http://www.conacyt.mx/comunicacion/revista/181/articulos/pdf/Cempoalxochit.pdf
Serrato-Cruz MA. 2005. Colecta, caracterización y aprovechamiento de Tagetes erecta L. Como
ornamental; Avances. X Congreso Nacional y III Internacional de Horticultura Ornamental.
Universidad Michoacana de San Nicolás de Hidalgo. Michoacán, México.
SIACON SAGARPA, 2006. Sistema de Información Agroalimentaria de consulta, SAGARPA.
[En línea] Disponible: http://www.sagarpa.gob.mx .
Spomer GG. 1973. The Concepts of "Interaction" and "Operational Environment" in
Environmental Analyses. Ecology 54 (1): 200-204.
Tambussi EA. 2004. Fotosíntesis, fotoprotección, productividad y estrés abiótico: algunos casos
de estudio. Tesis Doctoral en Biología. Universidad de Barcelona. Barcelona, España.
Ubaldo D. 2007. Efecto de la luz en el desarrollo y en la acumulación de carotenoides en lígulas
de Tagetes erecta L. Tesis Maestría. Centro de Desarrollo de Productos Bióticos del
Instituto Politécnico Nacional. Yautepec, Morelos. Mexico.
Valle-Piqueras C. 2004. Metabolismo secundario. [En línea] Disponible en:
http://www.psicostasia.com/Rev07.htm

64
Vanegas-Espinoza PE. 2003. Establecimiento de un sistema de regeneración y transformación
genética de plantas de cempaxúchil. Tesis Doctoral. CINVESTAV-IPN. Unidad Irapuato,
México.
Vázquez-García LM., Víveros-Farfán IMG y Salomé-Castañeda E. 2002. Cempasúchil (Tagetes
spp.) Recursos Fitogenéticos Ornamentales de México. Universidad del Estado de México.
28-67.
Verhoeven AS., Adams WW y Demmig-Adams B. 1999a. The xanthophyll cycle and
acclimation of Pinus ponderosa and Malva neglecta to winter stress. Oecologia 118: 277-
287.
Verhoeven AS., Adams WW., Demmig-Adams B., Croce R y Bassi R. 1999b. Xanthophyll cycle
pigment localization and dynamics during exposure to low temperatures and light stress in
Vinca major. Plant Physiology 120: 727-737
Villaseñor-Ríos JL y Espinosa-García FJ. 1998. Catálogo de malezas de México. Universidad
Nacional Autónoma de México, Consejo Nacional Consultivo Fitosanitario y Fondo de
Cultura Económica, México, D.F.
Waizel BJ. 2006. El cultivo de las plantas medicinales. pp. 345-361. En: Waizel-Bucay J (ed.)
Las plantas medicinales y las ciencias, una visión multidiciplinaria. Primera Edición.
Instituto Politécnico Nacional. México.
Wellensiek SJ. 1964. Dividing cells as a prerequisite for vernalization. Plant Physiology 39: 832-
835.
Wink M. 1999. Functions of plant secondary metabolites and their exploitation in biotechnology.
Annual Plant Reviews, Volume 3. 362 pp. Sheffield Academic Press, Sheffield. ISBN 1-
8493-4086-1.
Yammamoto YH., Bugos CR y Hieber D. 1999. Biochemistry and Molecular Biology of the
Xanthophyll Cycle. pp: 293-299. En: Frank H.A., Young AJ., Briton G. y Cogdell, R.J
(Eds.) The Photochemistry of Carotenoids. Kluwer Academic Publishers. USA.

65
Zaripheh S y Erdma JW. 2002. Factors that influence the bioavailability of xanthophylls. Journal
of Nutrition 132: 531S–534S.

66
11. ANEXOS
Anexo I. Plántulas transplantadas a campo.
Cultivos a los 12 días Cultivos a los 24 días
Cultivos a los 36 días Cultivos a los 46 días

67
Anexo II. Cuadro de intervalos del índice de color (IC*)
IC* Relación con el color
(-40 a -20) Si IC* es negativo, su valor relaciona los colores que van desde el azul-
violeta al verde profundo.
(-20 a -2) Si IC* es negativo, su valor relaciona los colores que van del
verde profundo al verde amarillento.
(-2 a +2) Si IC* está entre, representa el amarillo intenso al verdoso.
(+2 a +20) Si IC* es positivo, se relaciona con los colores que van desde el amarillo
pálido al naranja intenso.
(+20 a +40) Si IC* es positivo, se relaciona con los colores que van desde el naranja
intenso al rojo profundo.
Anexo III. Extracción de carotenoides (Filtración por columna)

68
Anexo IV. Cromatógrafo de líquidos de alta resolución (HPLC)
Anexo V. Perfil cromatográfico de la época de Agosto
Cromatograma del HPLC obtenido de los extractos de inflorescencias de cempaxúchil del cultivo
del mes de agosto.

69
Anexo VI. Perfil cromatográfico de la época de Septiembre
Cromatograma del HPLC obtenido de los extractos de inflorescencias de cempaxúchil del cultivo
del mes de septiembre.
Anexo VII. Perfil cromatográfico de la época de octubre
Cromatograma del HPLC obtenido de extractos de inflorescencias del cultivo del mes de octubre.

70
Anexo VIII. Perfil cromatográfico de la época del mes de noviembre
Cromatograma del HPLC obtenido de extractos de inflorescencias del cultivo del mes de
noviembre.
Anexo IX. Perfil cromatográfico de la época de diciembre
Cromatograma del HPLC obtenido de extractos de inflorescencias del cultivo del mes de
diciembre.

71
Anexo X. Curva tipo de luteína


















