
Clase 3 metab de h de c cadena respiratoria (2011)
77
LA OXIDACIÓN EN LOS SISTEMAS LA OXIDACIÓN EN LOS SISTEMAS BIOLÓGICOS BIOLÓGICOS CADENA RESPIRATORIA O TRANSPORTE ELECTRONICO y FOSFORILACION OXIDATIVA DR.MIGUEL WONG SERRANO
-
Upload
maximiliano-encino -
Category
Health & Medicine
-
view
2.153 -
download
2
Transcript of Clase 3 metab de h de c cadena respiratoria (2011)

LA OXIDACIÓN EN LOS SISTEMAS LA OXIDACIÓN EN LOS SISTEMAS BIOLÓGICOS BIOLÓGICOS
CADENA RESPIRATORIA
O
TRANSPORTE ELECTRONICO
y
FOSFORILACION OXIDATIVADR.MIGUEL WONG SERRANO

Desde el punto de vista químico
OXIDACIÓN
•Ganancia de oxígeno
•Pérdida de electrones
•Pérdida de hidrógeno
REDUCCIÓN
•Pérdida de oxígeno
•Ganancia de electrones
•Ganancia de hidrógeno (en compuestos orgánicos)
El uso principal del OXÍGENO es en la RESPIRACIÓN
Y ESTE ES EL PROCESO POR EL CUAL LAS CÉLULAS OBTIENEN ENERGÍA EN FORMA DE ATP
Este principio de OXIDO- REDUCCIÓN se aplica a los sistemas bioquímicos y es un concepto importante para la
comprensión de la naturaleza de las oxidaciones biológicas.

EN LOS SISTEMAS REDOX
LOS CAMBIOS DE ENERGÍA LIBRE
PUEDEN EXPRESARSE EN TÉRMINOS DEL
POTENCIAL DE
OXIDACIÓN – REDUCCIÓN
LAS ENZIMAS QUE INTERVIENEN EN LOS
PROCESOS REDOX
SE DENOMINAN
OXIDORREDUCTASASOXIDORREDUCTASAS

OxidorreductasasOxidorreductasas
Catalizan reacciones de oxido- reducción
Ared + Box Aox + Bred
A : es el reductor o dador electrónico; en el cursode la reacción se oxida (pierde electrones)
B : es el oxidante o aceptor electrónico; en el cursode la reacción se reduce (gana electrones)
En las reacciones redox, siempre tienen que estarpresentes a la vez el aceptor y el dador electrónico
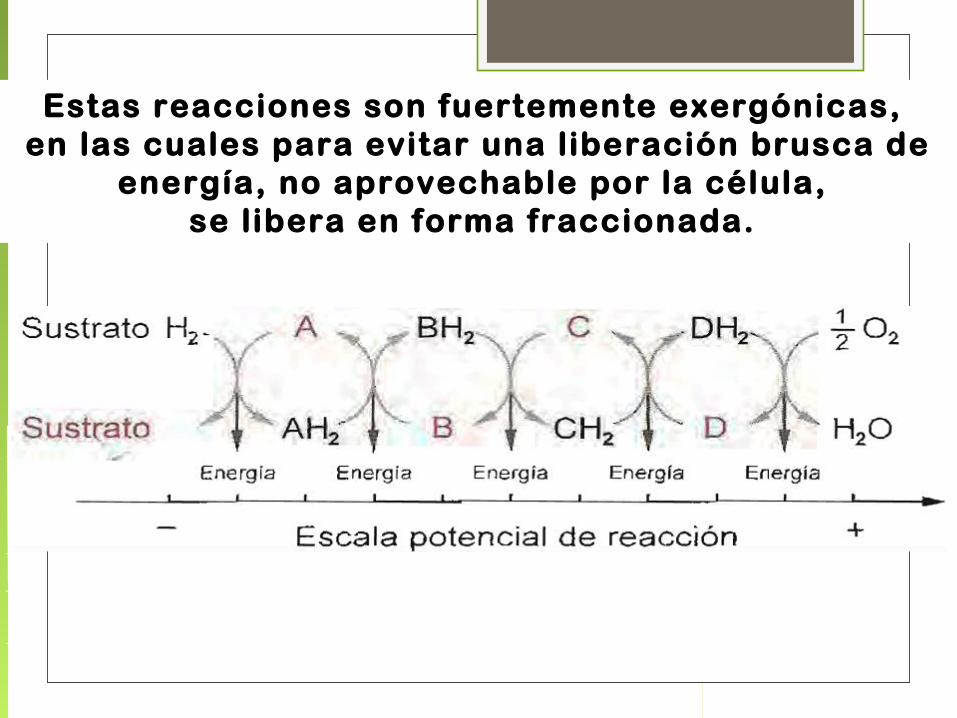
Estas reacciones son fuertemente exergónicas, en las cuales para evitar una liberación brusca de
energía, no aprovechable por la célula, se libera en forma fraccionada.

DISTINTAS FORMAS EN QUE LA CELULA PUEDE TRANSFERIR ELECTRONES
1.- Transferencia de 1 e-: Fe +++ Fe++
2.- Transferencia de un átomo de hidrógeno:
(H+ + e-): AH2 + B A + BH2
3.- Transferencia de un ion Hidruro (:H-)
AH2 + NAD+ → A + NADH + H+
4.- Transferencia de e- desde un reductor orgánico al oxígeno:
R-CH3 + ½ O2 RCH2-OH

• Gran parte de los sustratos oxidados en el organismo sufren deshidrogenación.
• Las reacciones de deshidrogenación son catalizadas por las ENZIMAS DESHIDROGENASAS.
• En estas reacciones el hidrógeno es captado por una coenzima.
• Las coenzimas pueden ser:
- Nicotinamida (NAD o NADP) - Flavina (FAD).
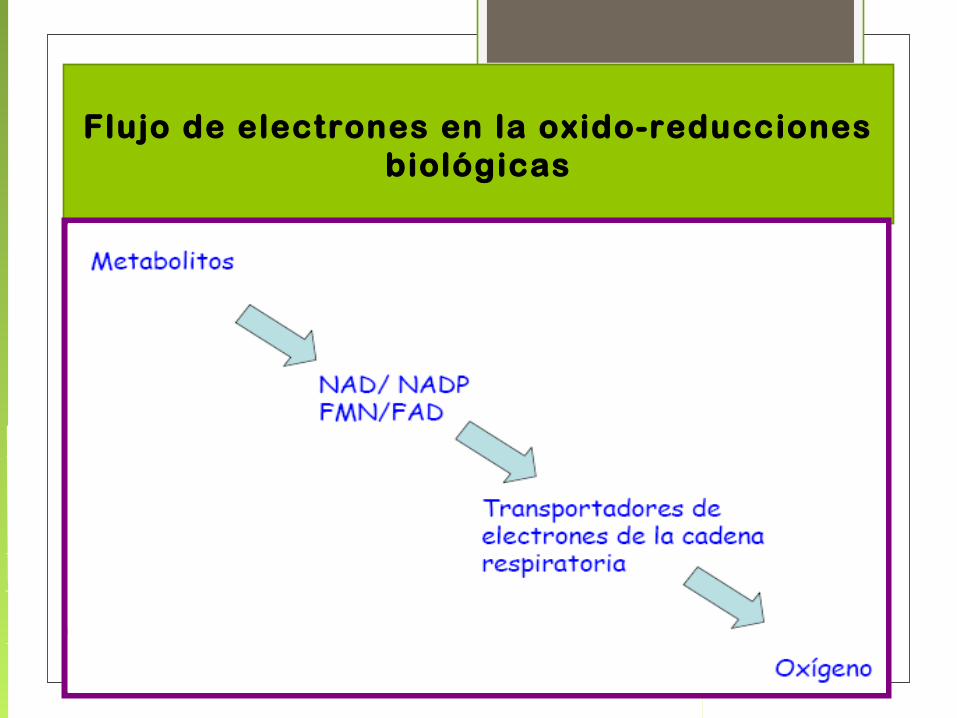
Flujo de electrones en la oxido-reducciones biológicas

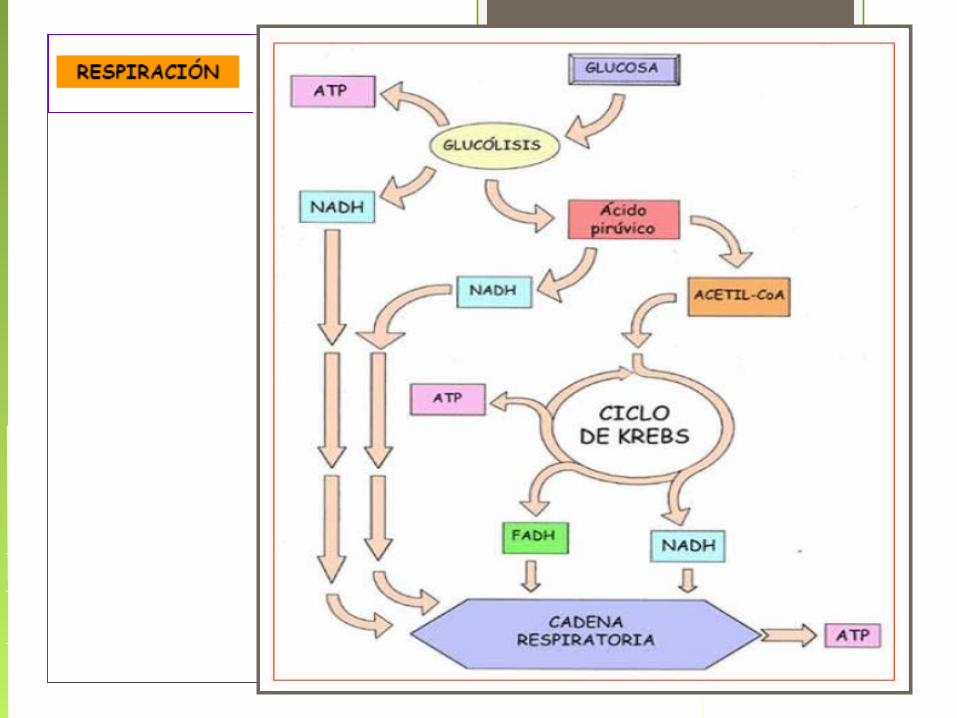
Respiración Celular
Es el conjunto de reacciones en las cuales el ác. pirúvico producido por la glucólisis se desdobla a CO2 y H2O y se producen 30 ATP.
En las células eucariontes la respiración se realiza en la mitocondria.

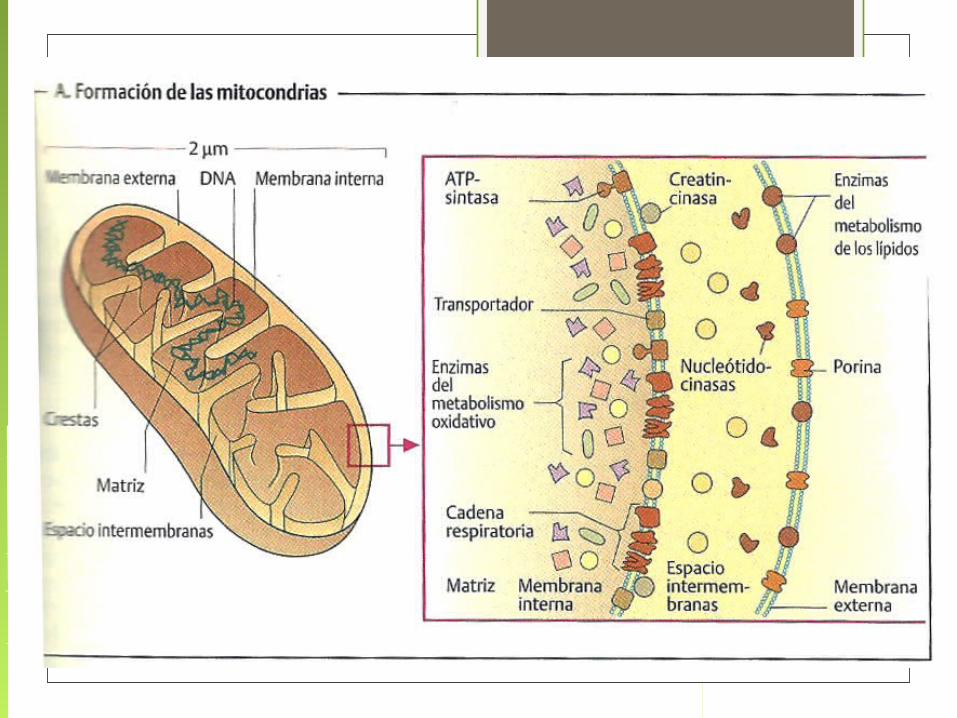
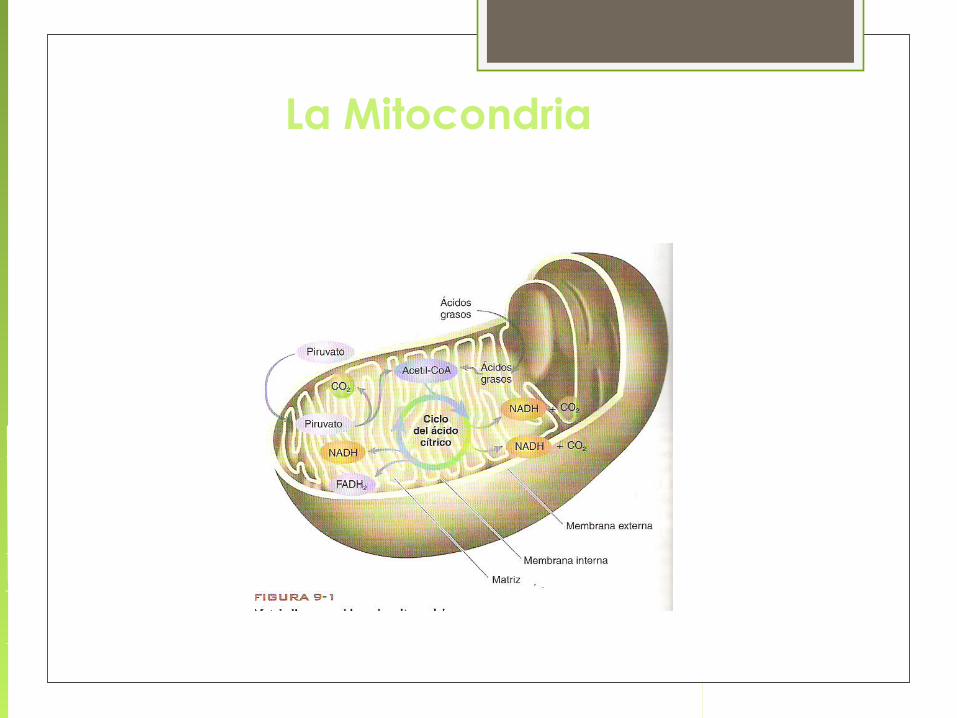
LA MITOCONDRIA
FÁBRICA DE ENERGÍA CELULAR
ES EL SITIO DONDE TIENEN LUGAR
EL TRANSPORTE ELECTRÓNICO Y
LA FOSFORILACIÓN OXIDATIVA

Localización
30ATP
32ATP

La cadena transportadora de electrones
El NADH+H y el FADH2, obtenidos en el ciclo de Krebs, van a entrar en una cadena transportadora de electrones o cadena respiratoria, donde pasan los electrones, de una molécula reducida a otra oxidada, hasta el aceptor final que será el oxígeno molecular, que al reducirse formará agua.

CADENA DE TRANSPORTE ELECTRONICO
•Los componentes de la cadena se encuentran en la membrana mitocondrial interna.
•Reciben equivalentes de reducción de NADH Y FADH2 producidos en la matriz.
•Los componentes actúan secuencialmente en orden creciente según sus potenciales de reducción.
•La energía que se libera durante la transferencia electrónica está acoplada a varios procesos endergónicos entre los que se destaca la síntesis de ATP.


Reacciones de la cadena de transporte de electrones
Con excepción de la coenzima Q, todos los miembros de esta cadena son
proteínas.
Pueden funcionar como enzimas como en el caso de varias deshidrogenasas.
Pueden contener hierro como parte de su centro hierro-azufre .
Los citocromos a y a3 contienen cobre.
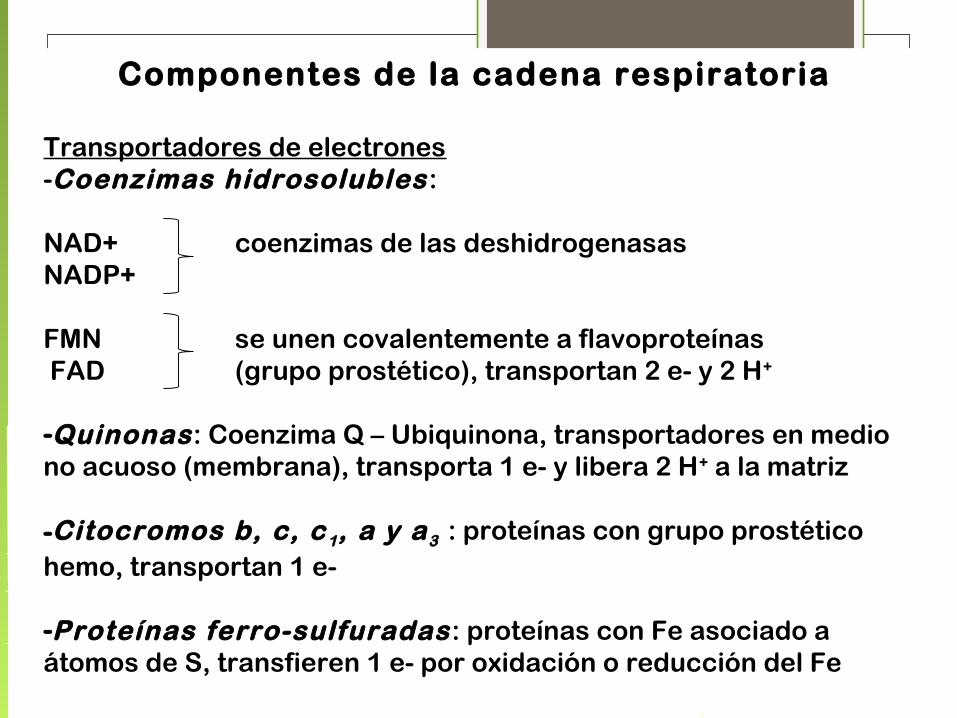
Componentes de la cadena respiratoria
Transportadores de electrones-Coenzimas hidrosolubles:
NAD+ coenzimas de las deshidrogenasasNADP+
FMN se unen covalentemente a flavoproteínas FAD (grupo prostético), transportan 2 e- y 2 H+
-Quinonas: Coenzima Q – Ubiquinona, transportadores en medio no acuoso (membrana), transporta 1 e- y libera 2 H+ a la matriz
-Citocromos b, c, c 1, a y a3 : proteínas con grupo prostético hemo, transportan 1 e-
-Proteínas ferro-sulfuradas: proteínas con Fe asociado a átomos de S, transfieren 1 e- por oxidación o reducción del Fe
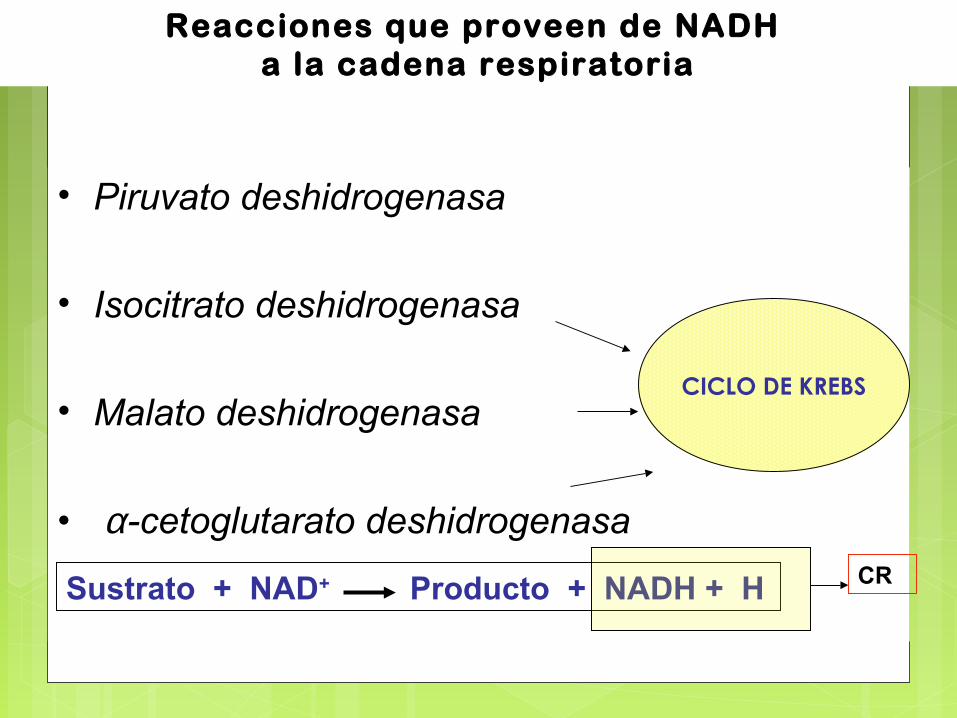
Reacciones que proveen de NADH a la cadena respiratoria
• Piruvato deshidrogenasa
• Isocitrato deshidrogenasa
• Malato deshidrogenasa
• α-cetoglutarato deshidrogenasa
CICLO DE KREBS
Sustrato + NAD+ Producto + NADH + H CR

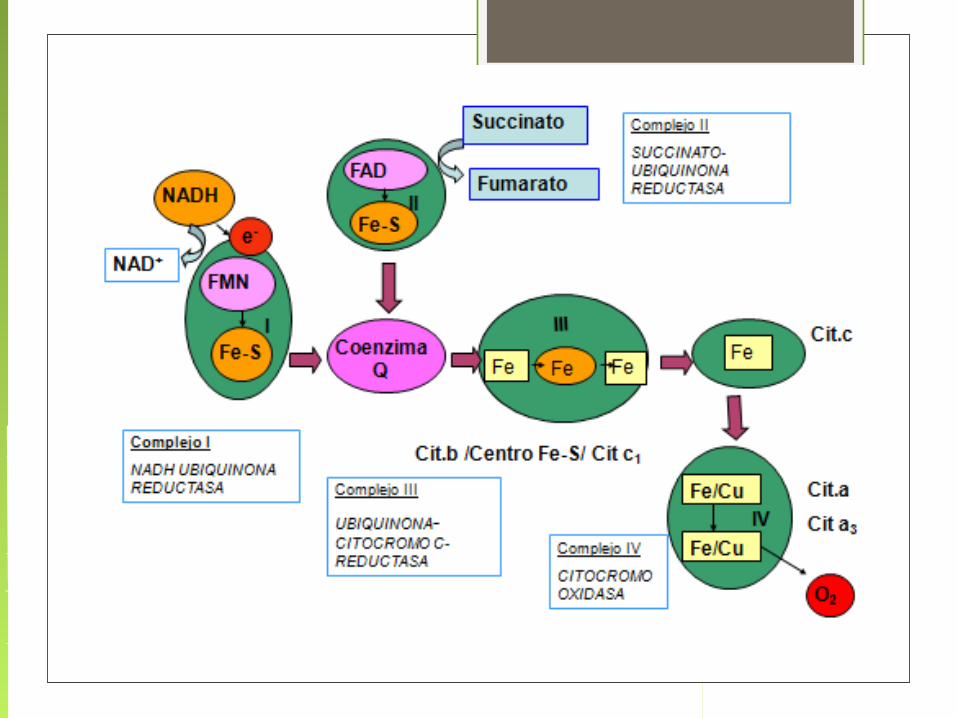
Componentes de la Cadena de transporte electrónico
Complejo enzimático Grupos prostéticos
Complejo I (NADH deshidrogenasa) FMN, FeS
Complejo II ( succinato deshidrogenasa) FAD, FeS
Complejo III (citocromo bc1) Hemo, FeS
Citocromo c Hemo
Complejo IV (citocromo oxidasa) Hemo, Cu
Complejo V (ATP sintasa)

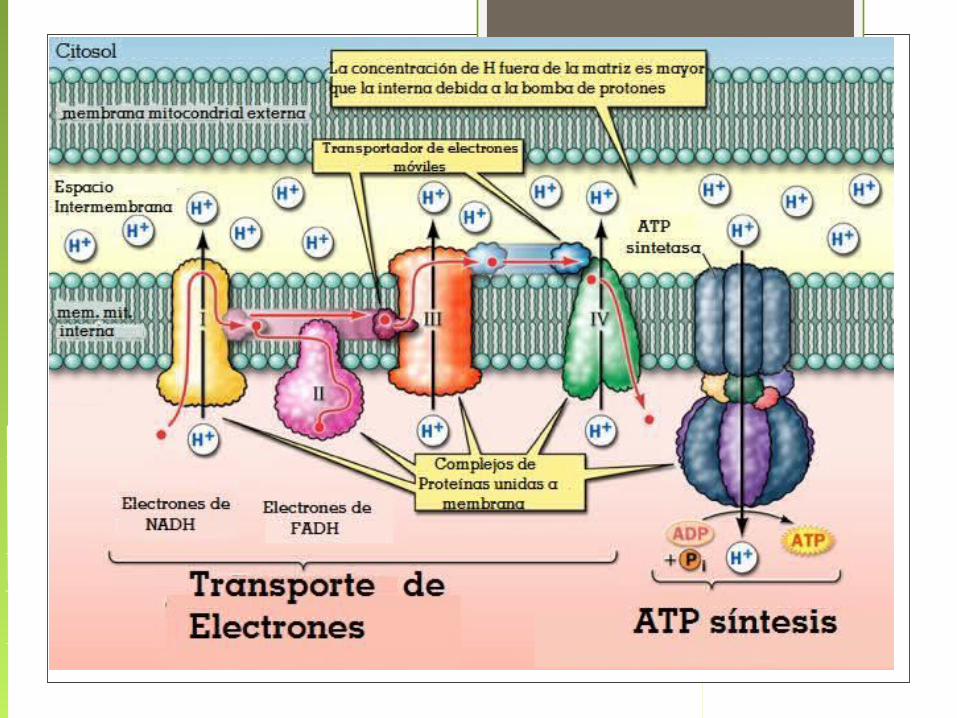
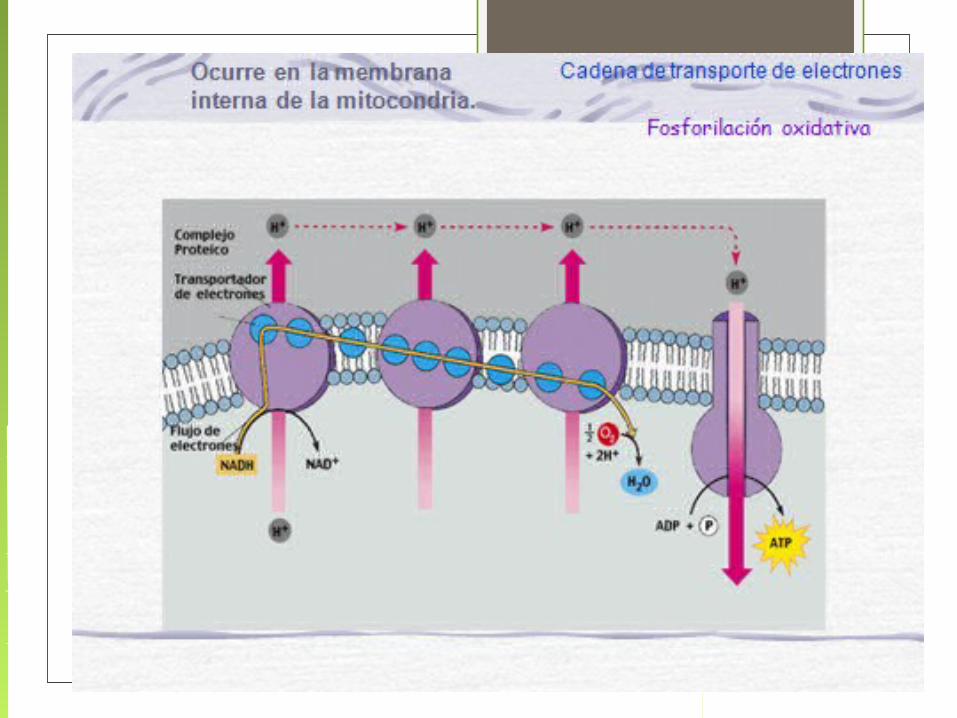
La Cadena de Transporte de Electrones comprende dos procesos:
1.- Los electrones son transportados a lo largo de la membrana, de un complejo de proteínas transportadoras a otro.
2. Los protones son translocados a través de la membrana, desde el interior o matriz hacia el espacio intermembrana de la mitocondria.
Esto constituye un gradiente de protones
El oxígeno es el aceptor terminal del electrón, combinándose con electrones e iones H + para producir
agua.

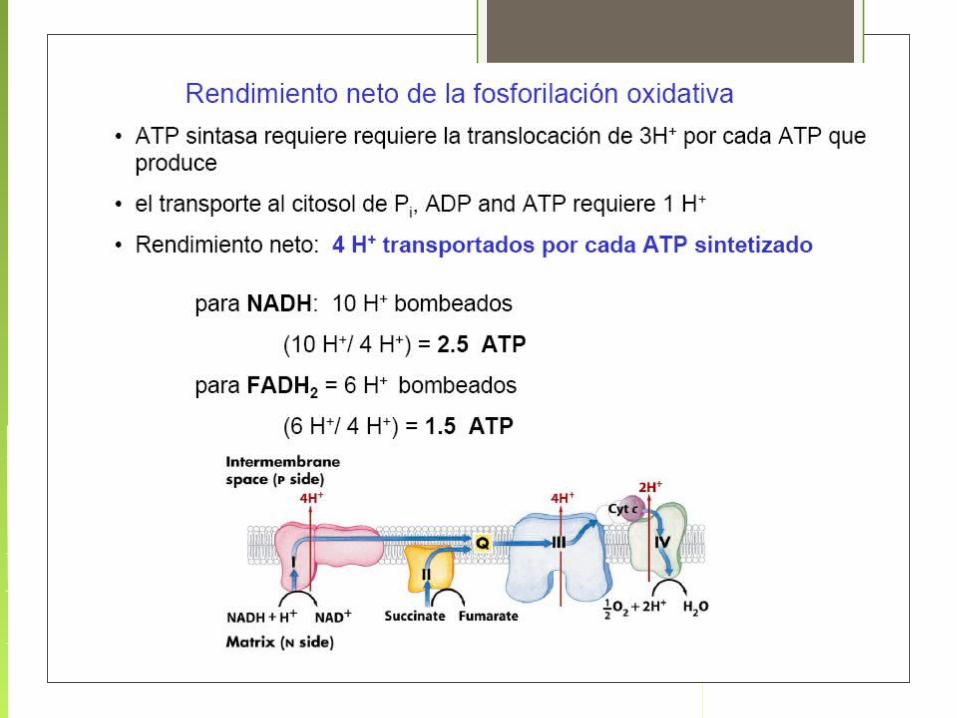
• La transferencia de electrones desde el NADH a través de la cadena respiratoria hasta el O 2 es un proceso altamente exergónico.
• La mayor parte de esa energía se emplea para bombear protones fuera de la matriz.
• Por cada par de electrones transferidos al O 2 los complejos I y III bombean 4 H + y 2 el complejo IV.
• El complejo II no transfiere H + ya que no atraviesa la membrana interna como los demás.
• Así esta energía electroquímica generada por el gradiente protónico impulsa la síntesis de ATP.
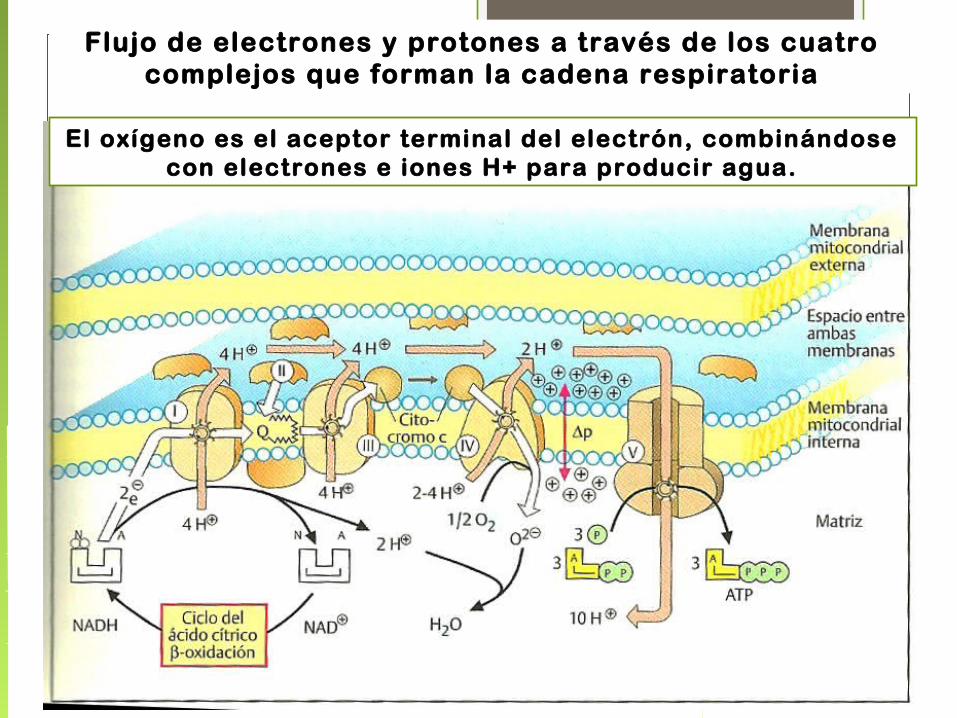
Flujo de electrones y protones a través de los cuatro complejos que forman la cadena respiratoria
El oxígeno es el aceptor terminal del electrón, combinándose con electrones e iones H+ para producir agua.

INHIBICION DEL TRANSPORTE ELECTRÓNICO
• Inhibidores del transporte electrónico Inhiben solamente el transporte de e-
• Inhibidores de la fosforilación Inhiben la síntesis de ATP, indirectamente el transporte
de e-
• Desacoplantes Impiden la síntesis de ATP pero no inhiben el transporte
de electrones. Actúan como ionóforos eliminando el gradiente de protones.
• Inhibidores de la translocasa Inhiben la entrada de ADP y la salida de ATP desde la
mitocondria
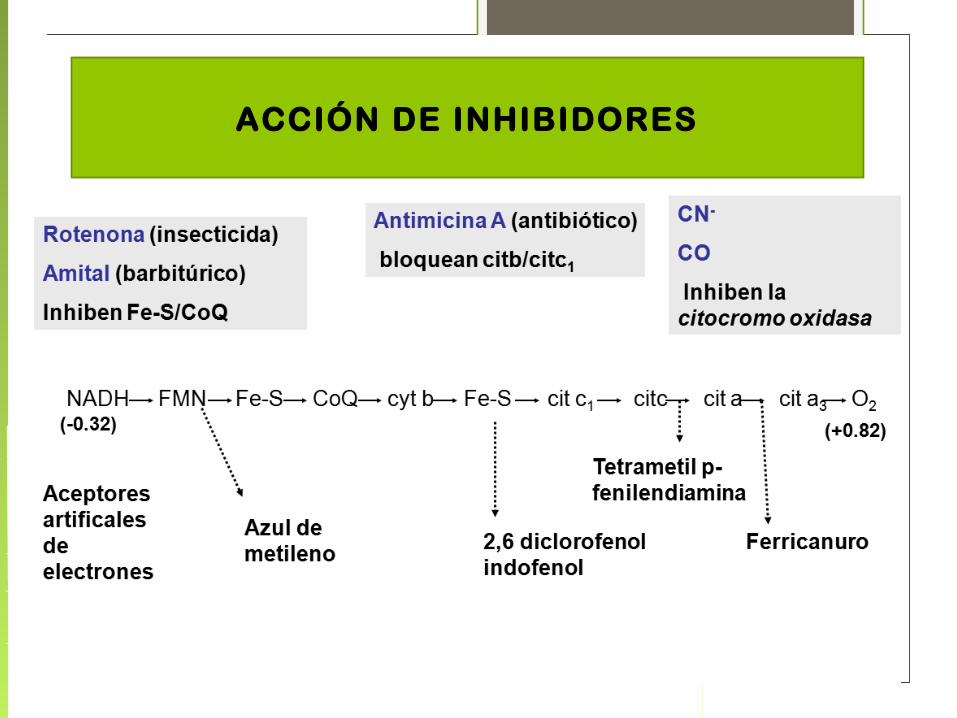
ACCIÓN DE INHIBIDORES

La cadena de transporte de electrones y la fosforilación oxidativa estuvieron separadas conceptualmente por mucho tiempo.
Las observaciones sobre la formación del ATP hacían pensar a los investigadores en un intermediario fosforilado de la reacción.
En 1961 Peter Mitchell propuso la Hipótesis Quimiosmótica:
“EL INTERMEDIARIO ENERGÉTICO NECESARIO PARA LA FORMACIÓN DEL ATP (O FOSFORILACIÓN DEL ADP), ES LA DIFERENCIA EN LA CONCENTRACIÓN DE PROTONES A TRAVÉS DE LA MEMBRANA”

PETER DENNIS MITCHELL (1920 - 1992)
-Interesado inicialmente en la penicilina, a partir de 1961 trabajó en el estudio sobre el almacenamiento de la energía en los seres vivos para ser posteriormente transportada a los puntos de utilización por medio de las moléculas de ATP.
-Así la energía liberada por el traslado de electrones en la cadena respiratoria se conserva mediante la fosforilación del ADP, que se convierte nuevamente en ATP, proceso denominado FOSFORILACIÓN OXIDATIVA.
-En 1978 fue galardonado con el Premio Nobel de Química por sus trabajos sobre el INTERCAMBIO DE ENERGÍA BIOLÓGICA MEDIANTE LA TEORÍA DE LA QUÍMICA OSMÓTICA.

POSTULADOS DE LA TEORIA QUIMIOSMOTICA
• Pasaje de H+ durante la transferencia de electrones desde la matriz al espacio intermembrana.
• Generación de un gradiente electroquímico : flujo electrónico acompañado de la transferencia de protones.
• Los protones acumulados en el espacio intermembrana crean una fuerza: «protón-motriz», por la tendencia de volver a pasar al interior para igualar el pH a ambos lados de la membrana.
• Esa fuerza es utilizada para el pasaje de los H + a través de Fo y así activan la ATP sintasa
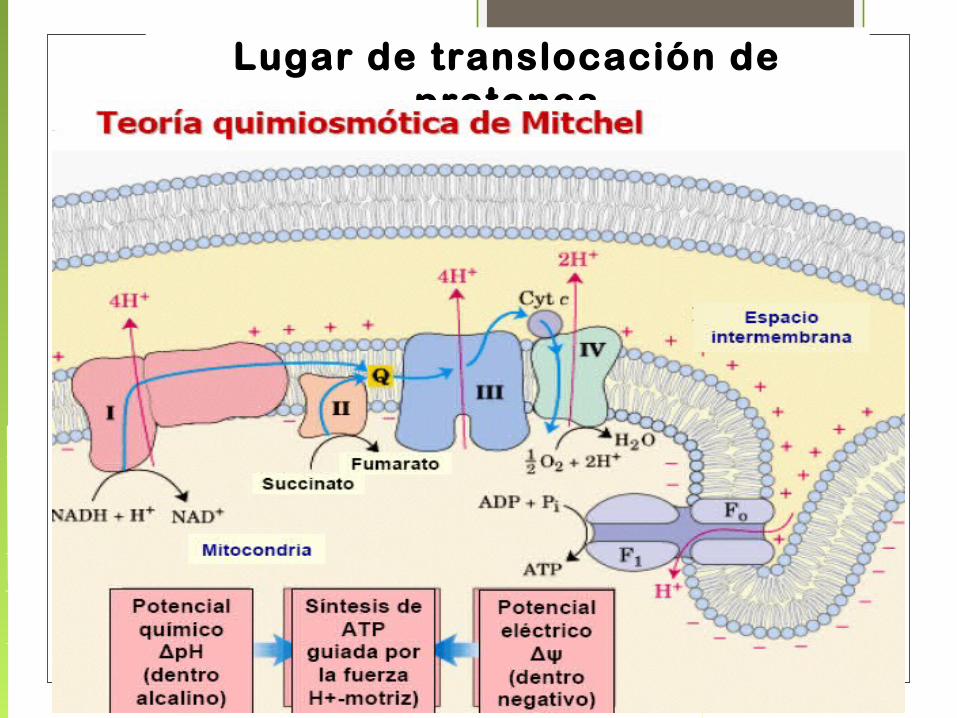
Lugar de translocación de protones
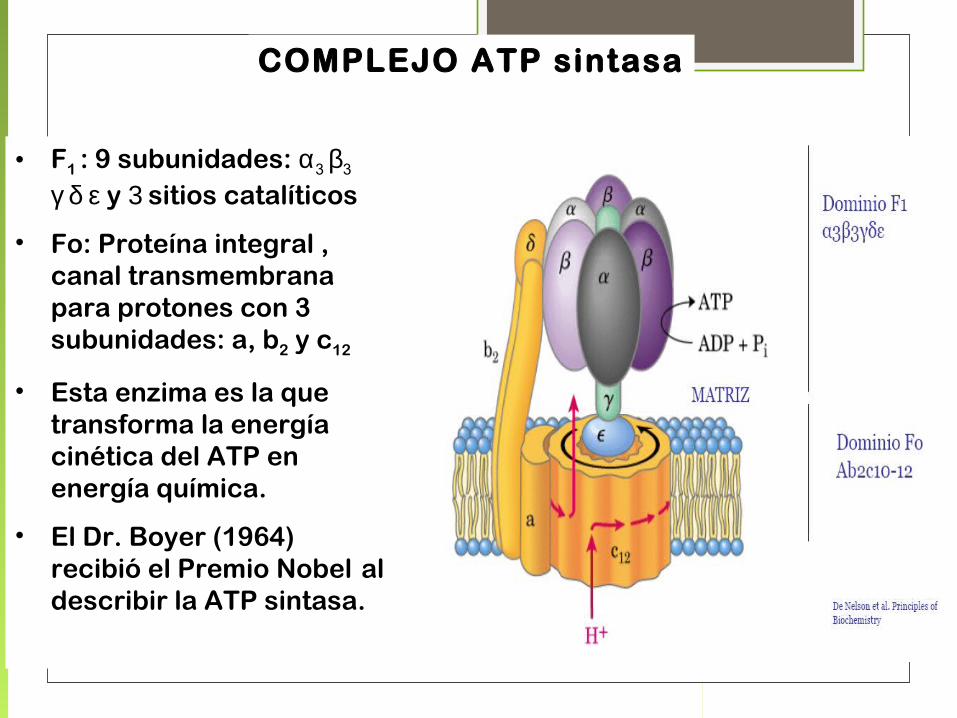
COMPLEJO ATP sintasa
• F1 : 9 subunidades: α3 β3
γ δ ε y 3 sitios catalíticos
• Fo: Proteína integral , canal transmembrana para protones con 3 subunidades: a, b2 y c12
• Esta enzima es la que transforma la energía cinética del ATP en energía química.
• El Dr. Boyer (1964) recibió el Premio Nobel al describir la ATP sintasa.
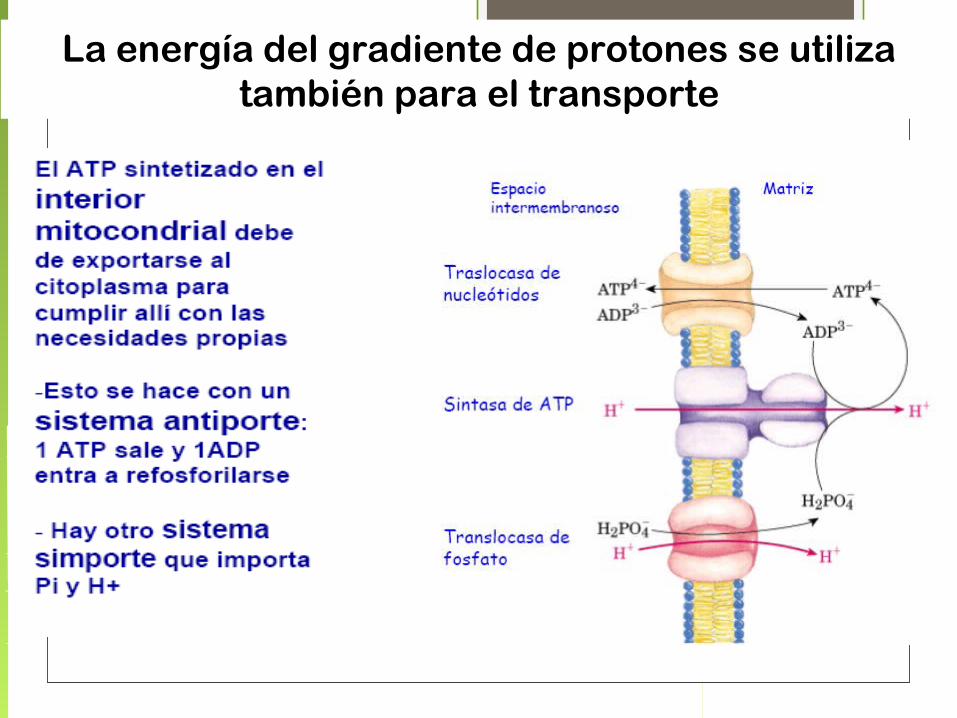
La energía del gradiente de protones se utiliza también para el transporte

• El control de la fosforilación oxidativa permite a la célula producir solo la cantidad de ATP que se requiere para el mantenimiento de sus actividades.
• El valor del cociente P/O, representa el número de moles de Pi que se consumen para que se reduzca cada átomo de O2 a H2O.
• El cociente máximo medido para la oxidación de NADH es 2,5 y para FADH2 es 1,5, para mayor practicidad se consideran 3 ATP y 2 ATP, respectivamente.
Control respiratorio por el aceptor:
• Las mitocondrias solo pueden oxidar al NADH y al FADH cuando hay una concentración suficiente de ADP y Pi.
• Cuando todo el ADP se transformó en ATP, disminuye el consumo de oxígeno y aumenta cuando se suministra ADP.

Inhibidores de la fosforilación
Oligomicina:
•Bloquea el flujo de protones a través de F0, impidiendo la fosforilación.•Se inhibe la síntesis de ATP•Se acumulan protones y se produce una fuerza inversa deteniéndose el transporte de electrones.
Desacoplantes: •Compuestos que impiden la síntesis de ATP, pero no bloquean el flujo de electrones, de esa manera desacoplan la cadena respiratoria de la fosforilación oxidativa.•El 2,3-dinitrofenol (DNF) transfiere iones hidrógeno desde el lado externo hacia la matriz y anula el gradiente de protones creado por la cadena respiratoria.

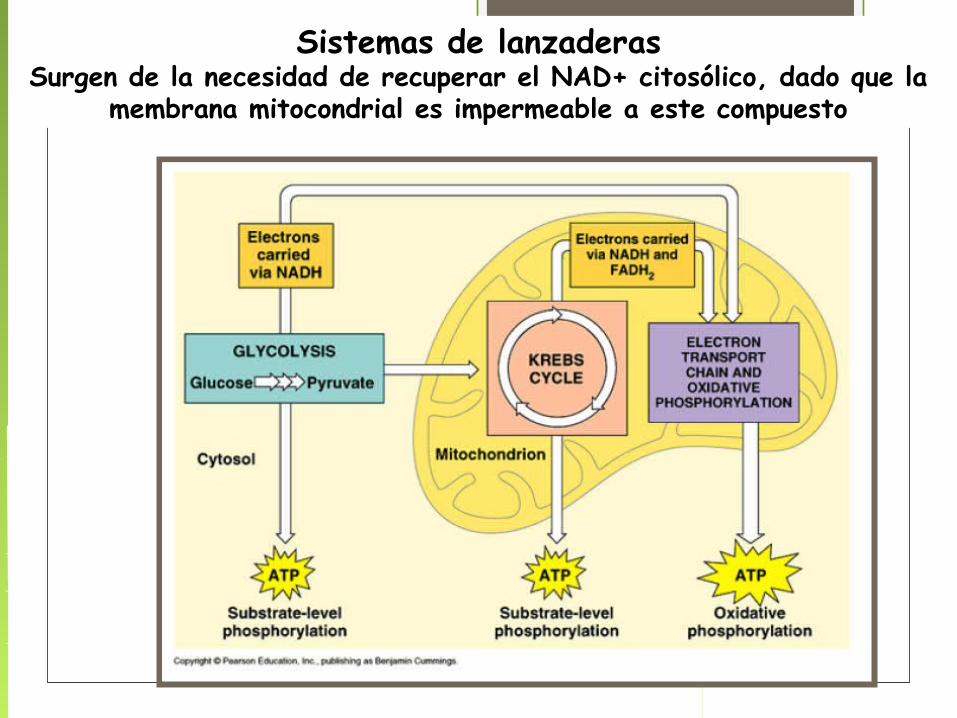
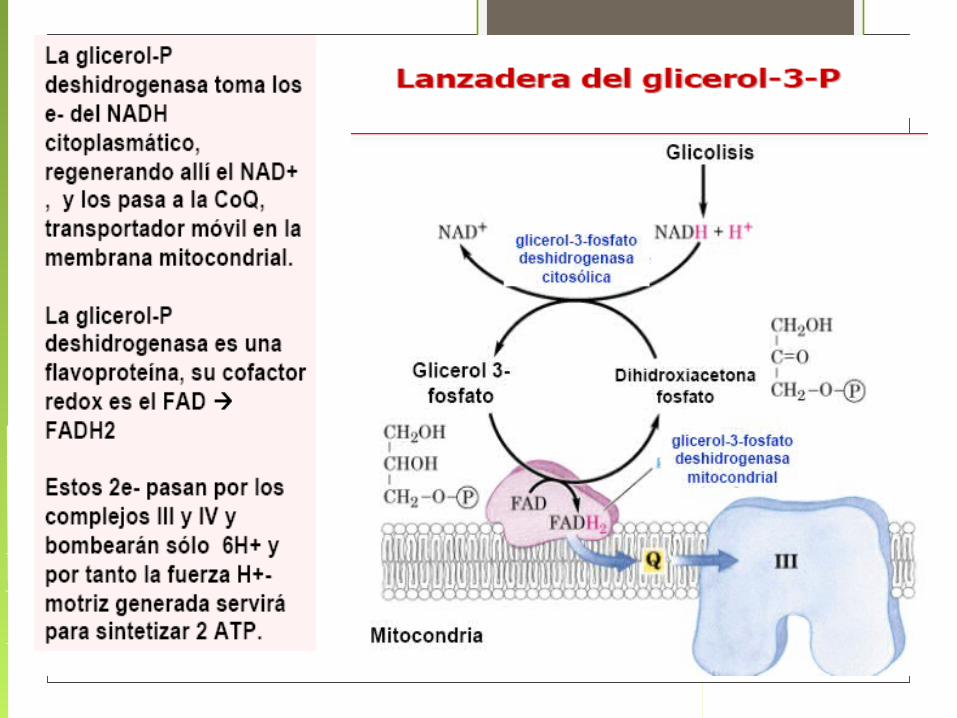
Sistemas de lanzaderasSurgen de la necesidad de recuperar el NAD+ citosólico, dado que la
membrana mitocondrial es impermeable a este compuesto
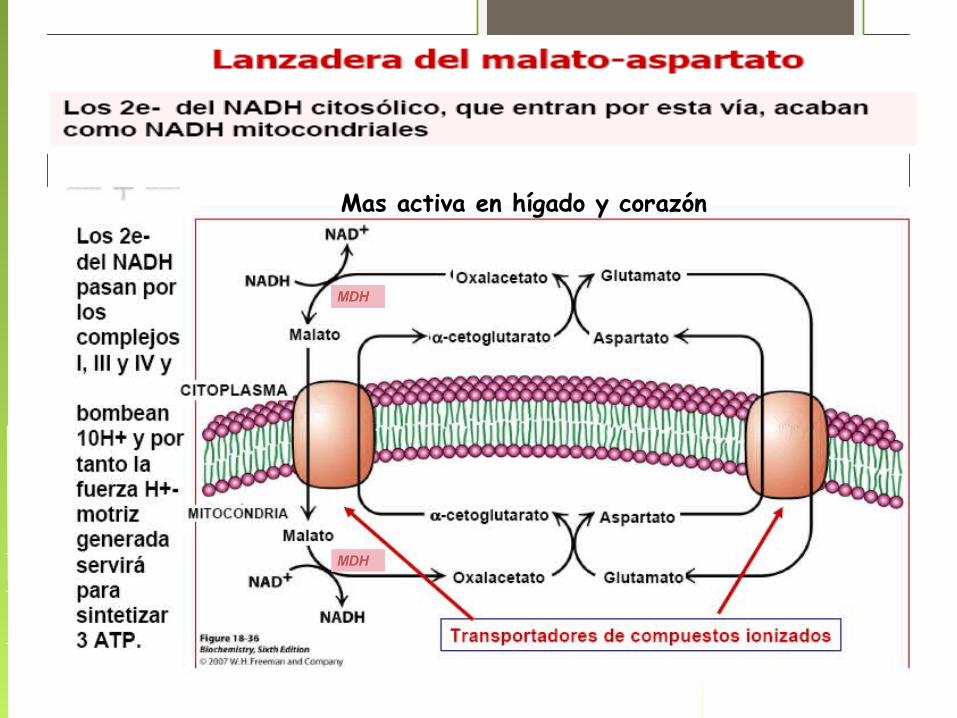
Mas activa en hígado y corazón


Ciclo del Ciclo del Acido CítricoAcido Cítrico

Ciclo de KrebsCiclo de Krebs
El ciclo de Krebs (también llamado ciclo del ácido cítrico o ciclo de los ácidos tricarboxílicos) es una serie de reacciones químicas de gran importancia, que forman parte de la respiración celular en todas las células aerobias, es decir que utilizan oxígeno.
En organismos aeróbicos el ciclo de Krebs es parte de la vía catabólica que realiza la oxidación de hidratos de carbono, ácidos grasos y aminoácidos hasta producir CO2 y agua, liberando energía en forma utilizable (poder reductor y ATP).
El ciclo de Krebs también proporciona precursores para muchas biomoléculas tales como ciertos aminoácidos. Por ello se considera una vía anfibólica, es decir, catabólica y anabólica al mismo tiempo.

Historia del Ciclo de Krebs El ciclo de Krebs recibe su nombre en honor
a su descubridor Sir Hans Krebs, quien propuso los elementos clave de esta vía en 1937.
La historia comienza a principios de la década de los 30´s con el descubrimiento de que al agregar succinato, fumarato y malato a músculos machacados incrementa la velocidad del consumo de Oxígeno.
El oxaloacetato se incorporó a la lista de ácidos dicarboxílicos cuando se descubrió que se podía formar en condiciones aeróbicas a partir del piruvato.

En 1935 A. Szent-Györgyi propuso que ciertos pares de ácidos dicarboxilicos eran interconvertidos por la acción de deshidrogenasas y que este proceso estaba relacionado con la respiración.
Carl Martius y Franz Knoop mostraron que el ácido cítrico es convertido en alfa-cetoglutarato por medio del isocitrato. Se supo también que el alfa-cetoglutarato puede ser oxidado a succinato.
La formación del citrato era la pieza faltante para poder armar completamente el rompecabezas metabólico.
El descubrimiento que resolvió este rompecabezas y unificó el metabolismo fue hecho en 1937 por Sir Hans Krebs y W.A. Johnson: ellos mostraron que el citrato es derivado del piruvato y del oxaloacetato completando lo que se conoce como el ciclo del ácido cítrico.

En condiciones anaerobias, las células animales reducen el piruvato a lactato, en las levaduras a etanol.
En condiciones aerobias, el piruvato ingresa a la matriz mitocondrial y es convertido a acetil-Coenzima A (AcCoA) para llevar estos Carbonos a su estado de oxidación total en el ciclo del ácido cítrico.
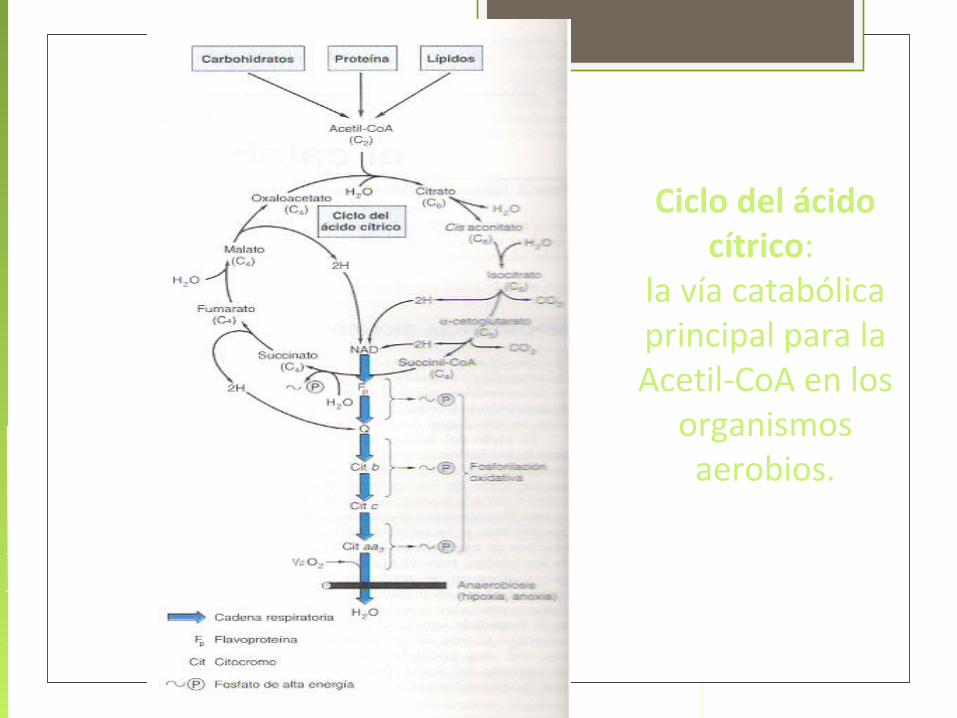
El ciclo del ácido cítrico, considerado el embudo del metabolismo, consiste ocho reacciones enzimáticas, todas ellas mitocondriales en los eucariontes.
El ciclo del ácido cítrico es la vía central del metabolismo aerobio: es la vía oxidativa final en el catabolismo de los carbohidratos, ácidos grasos y aminoácidos, además es una fuente importante de intermediarios de vías biosintéticas.

En muchas células la acción acoplada del ciclo del ácido cítrico y la cadena de transporte de electrones son responsables de la mayoría de la energía producida.
La oxidación del piruvato a Ac-CoA es catalizada por el complejo multienzimático de la piruvato deshidrogenasa (PDH), el proceso que es muy complicado, se resume en:
Piruvato + NAD+ + CoA ® Ac-CoA + NADH + H+ + CO2 DG°´= - 8.0kcal/mol
Esta reacción irreversible en tejidos animales, no forma parte del ciclo de Krebs, pero constituye un paso obligatorio para la incorporación de los glúcidos al ciclo.

El trabajo acoplado del ciclo del ácido cítrico y la cadena de transporte de electrones es la mayor fuente de energía metabólica.
El metabolismo aerobio del piruvato por el ciclo del ácido cítrico y la cadena de transporte de electrones produce mucha mas energía que la simple conversión aerobia del piruvato a lactato o etanol .
En condiciones aerobicas, el piruvato sufre una descarboxilacion oxidativa con la formación de AcCoA. El grupo acetilo del AcCoA es transferido al oxaloacetato para dar citrato

En reacciones subsecuentes, dos de los átomos de Carbono del citrato se oxidan a CO2 y el oxaloacetato es regenerado.
La reacción neta de ciclo del ácido cítrico también produce tres moléculas de NADH, una de FADH2 y una molécula del compuesto trifosfato de guanosina (GTP) altamente energético (en algunos organismos es directamente ATP) por cada molécula de AcCoA oxidada
Las moléculas de NADH y FADH2 son oxidadas en la cadena de transporte de electrones con la formación de ATP en la fosforilación oxidativa.
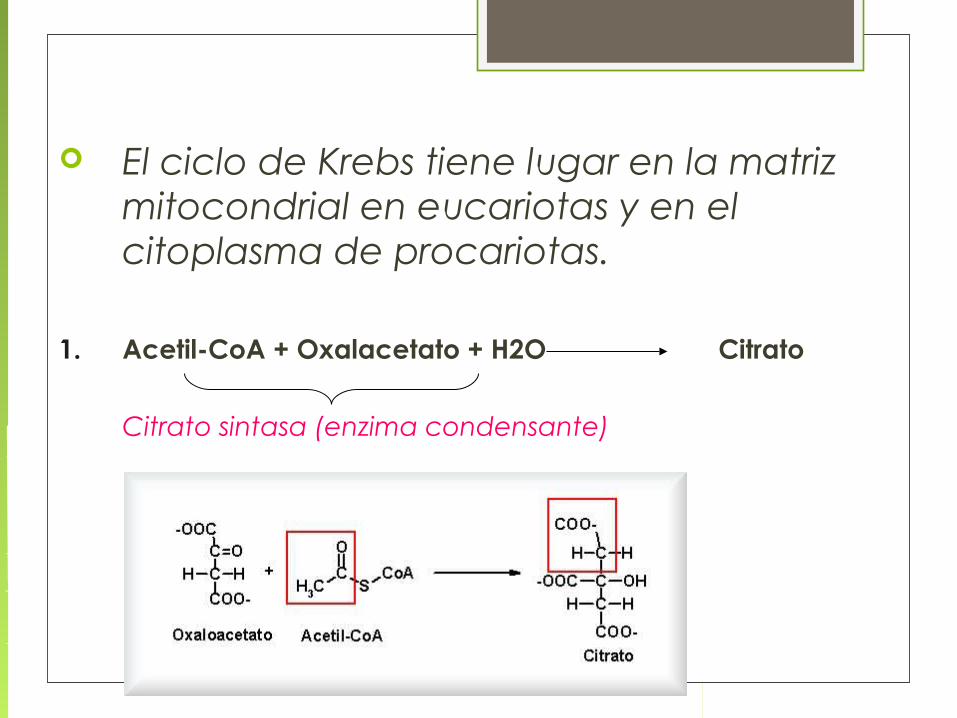
El ciclo de Krebs tiene lugar en la matriz mitocondrial en eucariotas y en el citoplasma de procariotas.
1. Acetil-CoA + Oxalacetato + H2O Citrato
Citrato sintasa (enzima condensante)

2. Citrato cis-aconitato + H2O Isocitrato
Aconitasa
3. Isocitrato + NAD α-cetoglutarato + CO2 + NADH primer CO2
Isocitrato deshidrogenasa
Enzima reguladora Descarboxilación oxidativa
Acoplada a la conversiónDe NAD NADH
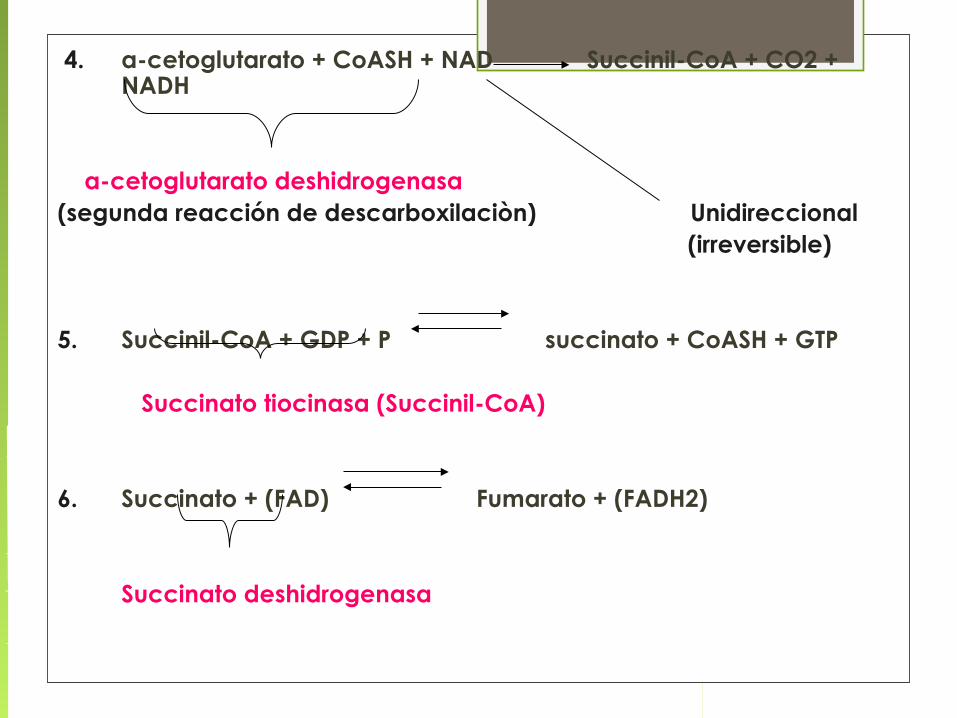
4. α-cetoglutarato + CoASH + NAD Succinil-CoA + CO2 + NADH
α-cetoglutarato deshidrogenasa(segunda reacción de descarboxilaciòn) Unidireccional (irreversible)
5. Succinil-CoA + GDP + P succinato + CoASH + GTP
Succinato tiocinasa (Succinil-CoA)
6. Succinato + (FAD) Fumarato + (FADH2)
Succinato deshidrogenasa
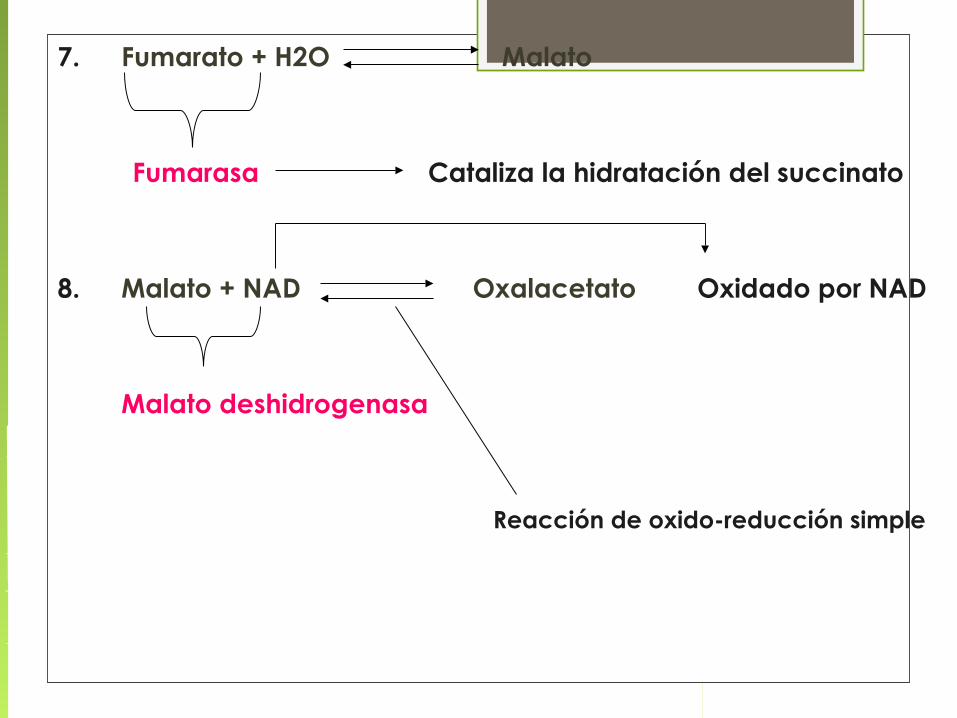
7. Fumarato + H2O Malato
Fumarasa Cataliza la hidratación del succinato
8. Malato + NAD Oxalacetato Oxidado por NAD
Malato deshidrogenasa
Reacción de oxido-reducción simple



Principales vías que convergen en el ciclo de Krebs
Las reacciones que forman intermediarios del ciclo se conocen como reacciones anapleróticas.
El ciclo de Krebs constituye la segunda etapa del catabolismo de carbohidratos. La glucólisis rompe la glucosa (6 carbonos) generando dos moléculas de piruvato (3 carbonos).
En eucariotas el piruvato se desplaza al interior de la mitocondria (gracias a un transportador específico de membrana interna). En la matriz mitocondrial produce acetil-CoA que entra en el ciclo de Krebs.

En el catabolismo de proteínas, los enlaces peptídicos de las proteínas son degradados por acción de enzimas proteasas en el tracto digestivo liberando sus constituyentes aminoacídicos. Estos aminoácidos penetran en las células, donde pueden ser empleados para la síntesis de proteínas o ser degradados para producir energía en el ciclo de Krebs.
En el catabolismo de grasas, los triglicéridos son hidrolizados liberando ácidos grasos y glicerol.
En muy diversos tejidos, especialmente en músculo cardiaco, los ácidos grasos son degradados en la matriz mitocondrial mediante sucesivos ciclos de beta oxidación que liberan unidades de acetil-CoA, que pueden incorporarse al ciclo de Krebs.

El ciclo de Krebs siempre es seguido por la fosforilación oxidativa.
Este proceso extrae la energía en forma de electrones de alto potencial de las moléculas de NADH y FADH2, regenerando NAD+ and FAD, gracias a lo cual el ciclo de Krebs puede continuar.
Los electrones son transferidos a moléculas de O2, rindiendo H2O.
Pero esta transferencia se realiza a través de una cadena transportadora de electrones capaz de aprovechar la energía potencial de los electrones para bombear protones al espacio intermembrana de la mitocondria.
De este modo el ciclo de Krebs no utiliza directamente O2, pero lo requiere al estar acoplado a la fosforilación oxidativa.
Por cada molécula de glucosa la energía obtenida mediante el metabolismo oxidativo, es decir, glucolisis seguida del ciclo de Krebs, equivale a unas 36 moléculas de ATP.

Ciclo de Krebs

Funciones del Ciclo de Krebs PRODUCE LA MAYOR PARTE DEL CO2 METABOLICO ES LA PRINCIPAL FUENTE DE COENZIMAS REDUCIDAS
(NADH+H, Y FADH2) QUE POSTERIORMENTE SE OXIDAN EN LA CADENA RESPIRATORIA PARA PRODUCIR ATP.
ES LA VIA COMÚN PARA LA DEGRADACIÓN METABÓLICA DE CARBOHIDRATOS, LIPIDOS Y PROTEÍNAS.
PROPORCIONA PRECURSORES PARA LA SINTESIS DE CARBOHIDRATOS, LIPIDOS Y PROTEINAS.
ES UNA ROTONDA DE TRÁFICO METABÓLICO (CHO A GRASAS, AMINOACIDOS A CARBOHIDRATOS)
RUTA PROHIBIDA: GRASAS A CARBOHIDRATOS REPRESENTA LA OXIDACIÓN METABÓLICA DEL
ACETATO: (como Acetil-CoA)
La Mitocondria
Ciclo del ácido cítrico:
la vía catabólica principal para la Acetil-CoA en los
organismos aerobios.
Energética del Ciclo de Krebs
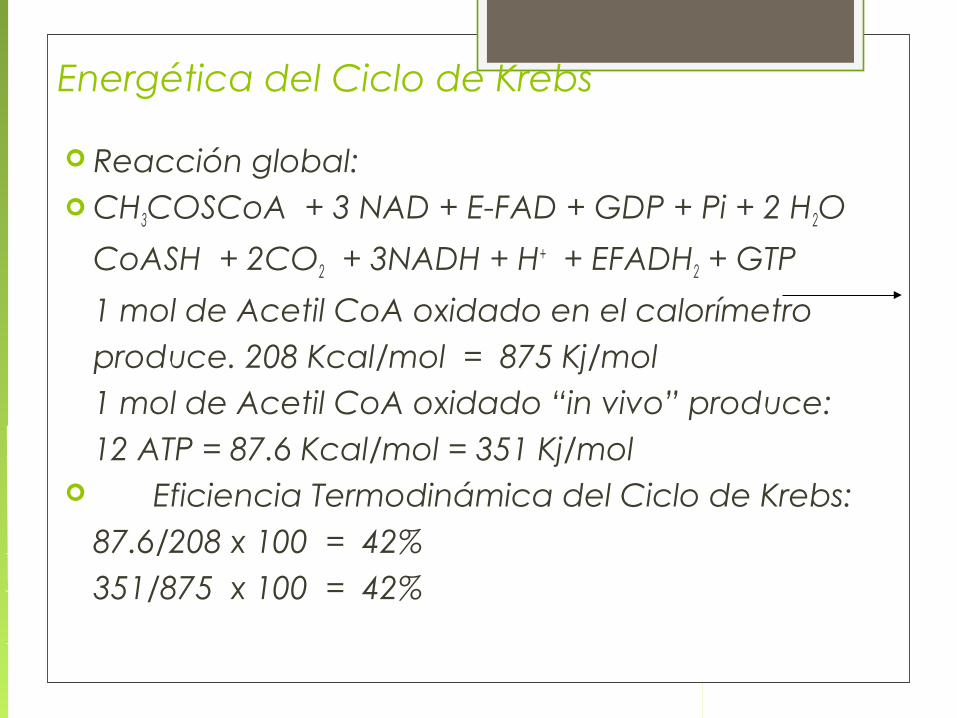
Reacción global: CH3COSCoA + 3 NAD + E-FAD + GDP + Pi + 2 H2O
CoASH + 2CO2 + 3NADH + H+ + EFADH2 + GTP
1 mol de Acetil CoA oxidado en el calorímetro produce. 208 Kcal/mol = 875 Kj/mol1 mol de Acetil CoA oxidado “in vivo” produce:12 ATP = 87.6 Kcal/mol = 351 Kj/mol
Eficiencia Termodinámica del Ciclo de Krebs:87.6/208 x 100 = 42%351/875 x 100 = 42%
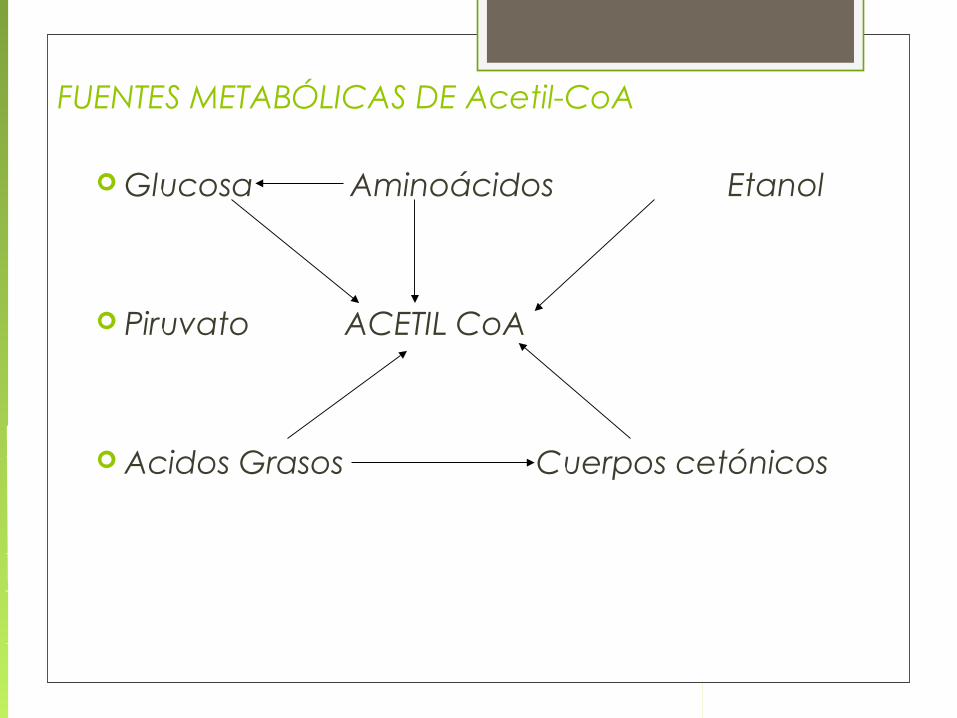
FUENTES METABÓLICAS DE Acetil-CoA
Glucosa Aminoácidos Etanol
Piruvato ACETIL CoA
Acidos Grasos Cuerpos cetónicos
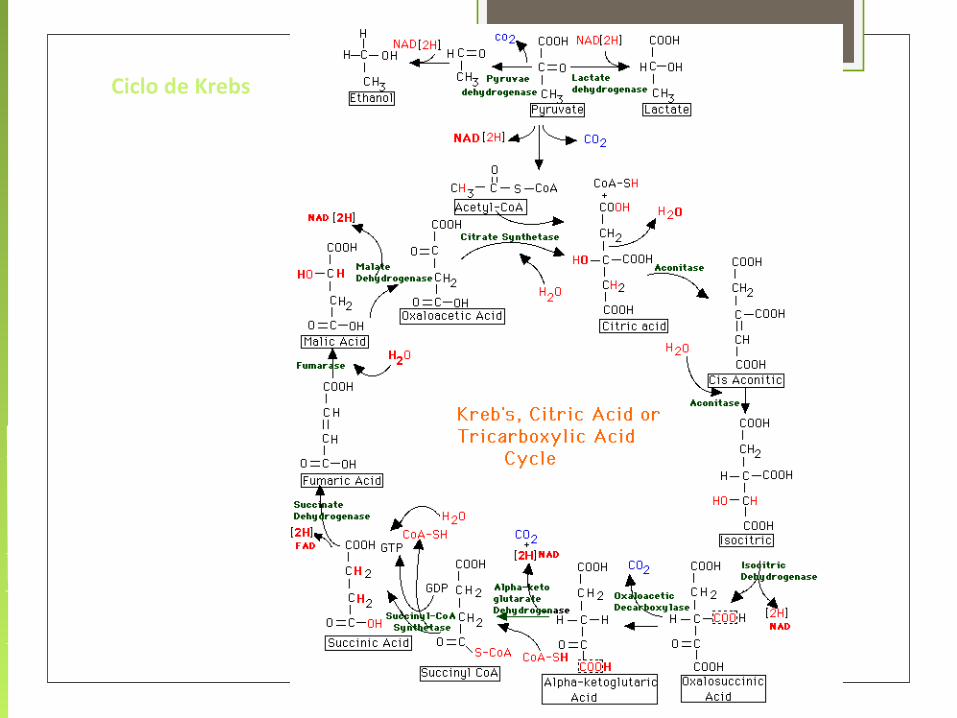
Ciclo de Krebs
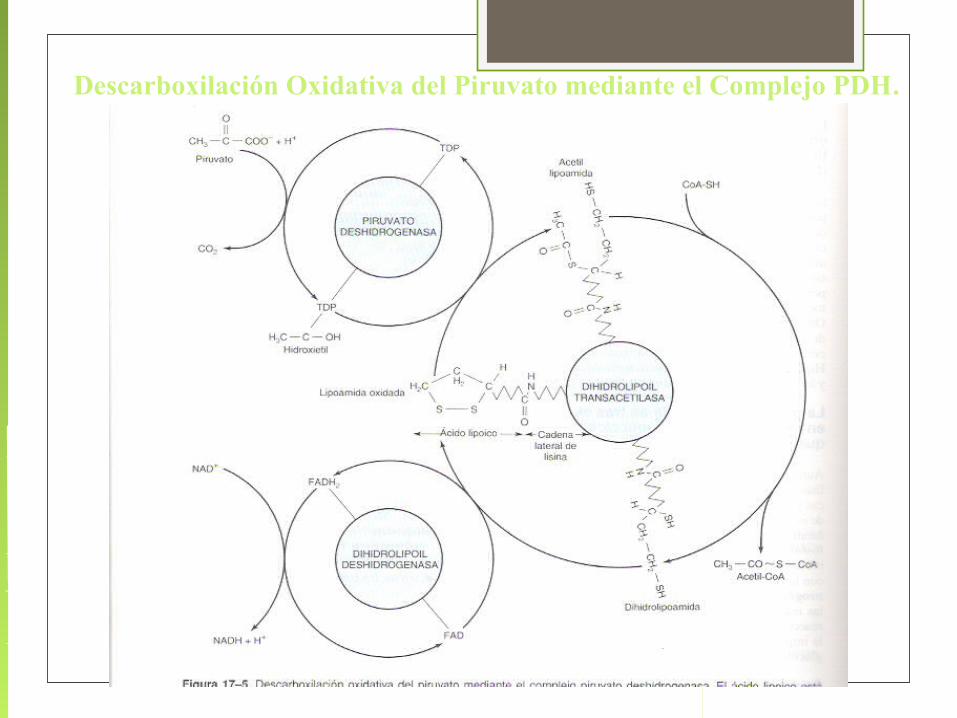
Descarboxilación Oxidativa del Piruvato mediante el Complejo PDH.
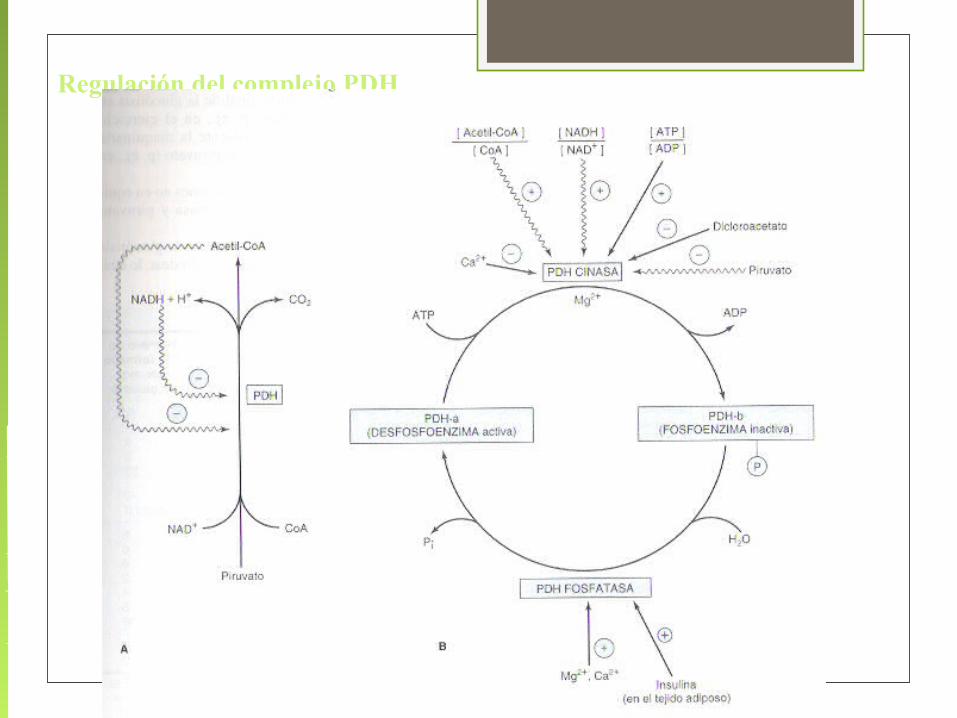
Regulación del complejo PDH

Reacciones del Ciclo del Ácido Cítrico
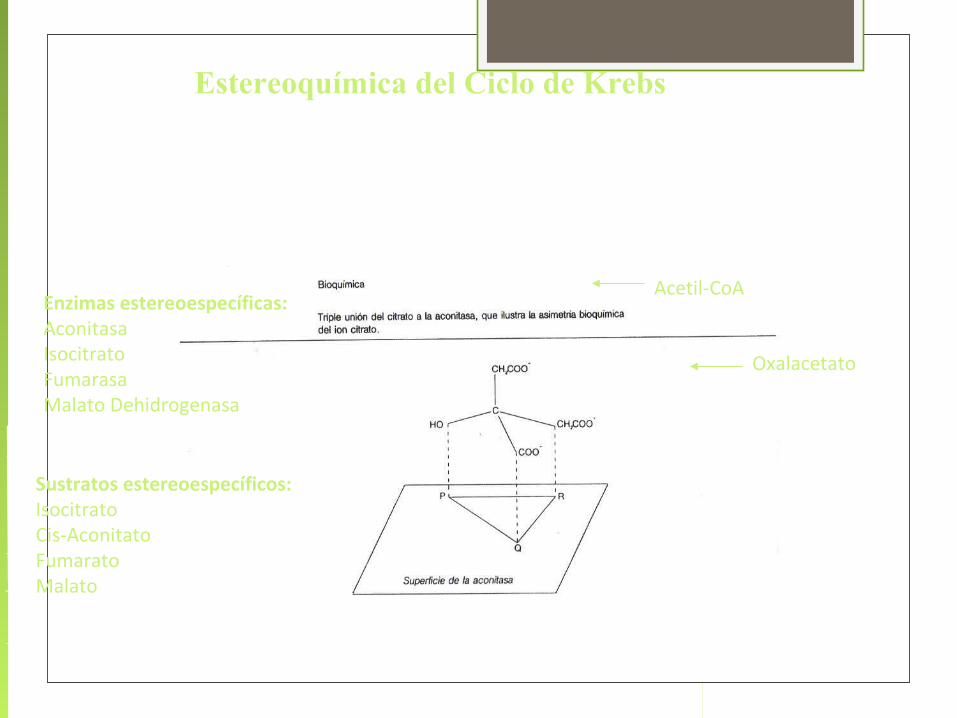
Estereoquímica del Ciclo de Krebs
Enzimas estereoespecíficas:AconitasaIsocitratoFumarasaMalato Dehidrogenasa
Sustratos estereoespecíficos:IsocitratoCis-AconitatoFumaratoMalato
Acetil-CoA
Oxalacetato
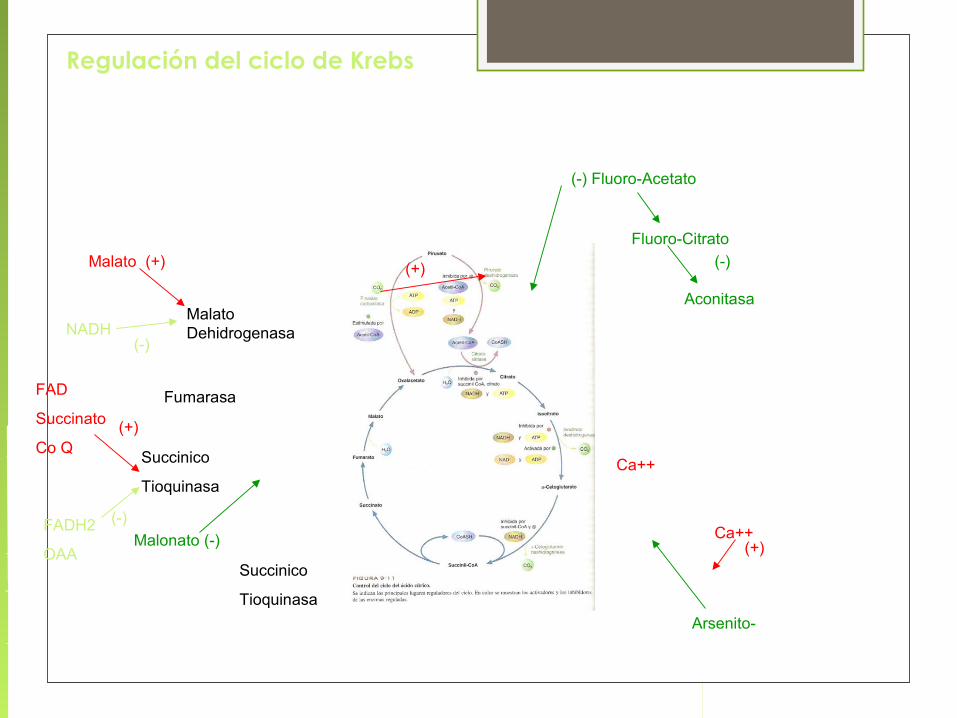
Regulación del ciclo de Krebs
(-) Fluoro-Acetato
Fluoro-Citrato
Aconitasa
(-)
Ca++
Ca++(+)
Arsenito-
Succinico
Tioquinasa
Succinico
Tioquinasa
Malonato (-)FADH2
OAA
(-)
FAD
Succinato
Co Q(+)
Fumarasa
Malato Dehidrogenasa
Malato (+)
NADH(-)
(+)
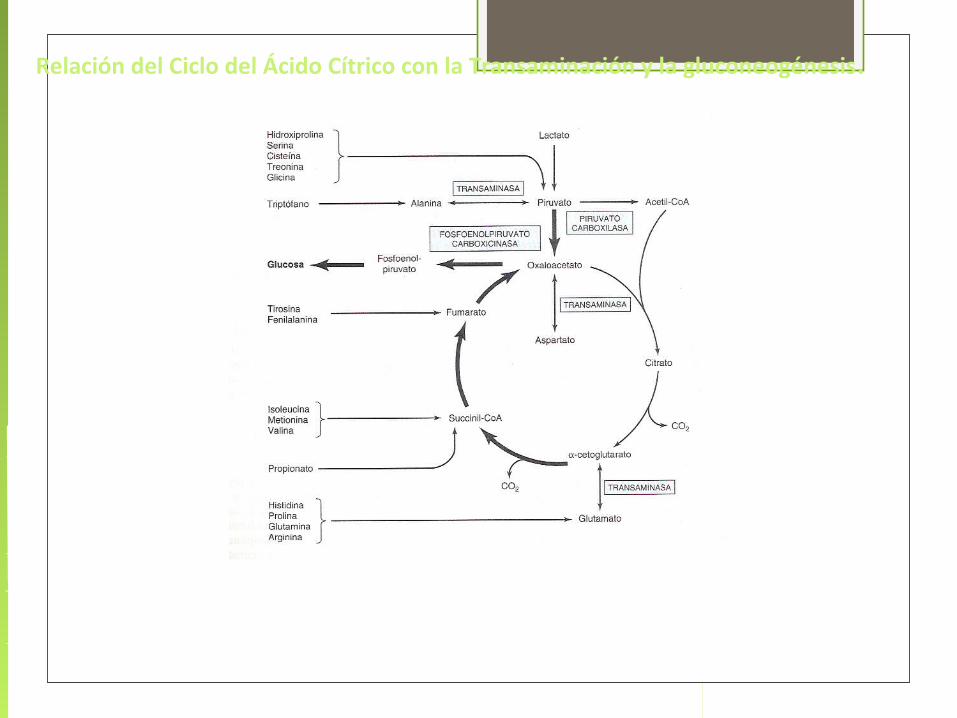
Relación del Ciclo del Ácido Cítrico con la Transaminación y la gluconeogénesis.
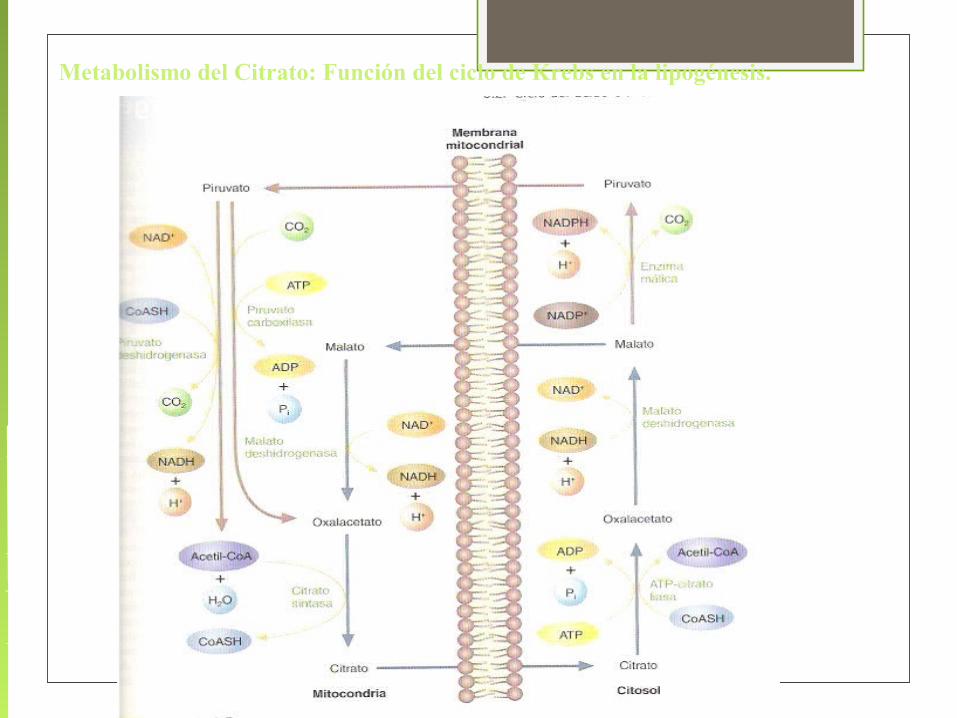
Metabolismo del Citrato: Función del ciclo de Krebs en la lipogénesis.
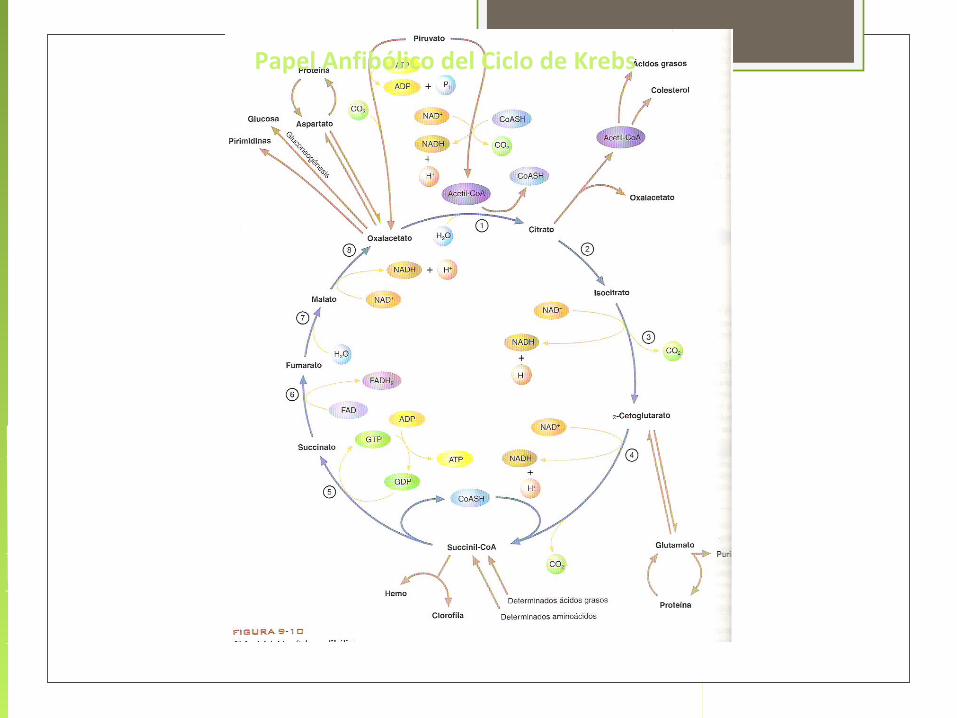
Papel Anfibólico del Ciclo de Krebs

Reposición de intermediarios del TCA especialmente OAA:
Piruvato Carboxilasa
Piruvato OAA
Enzima Málica
Piruvato Malato OAA
ATP
Biotina
CO2
NADP
NADPH+H
Reacciones Anapleróticas
Mitocondrial
Mitocondrial y en citosol
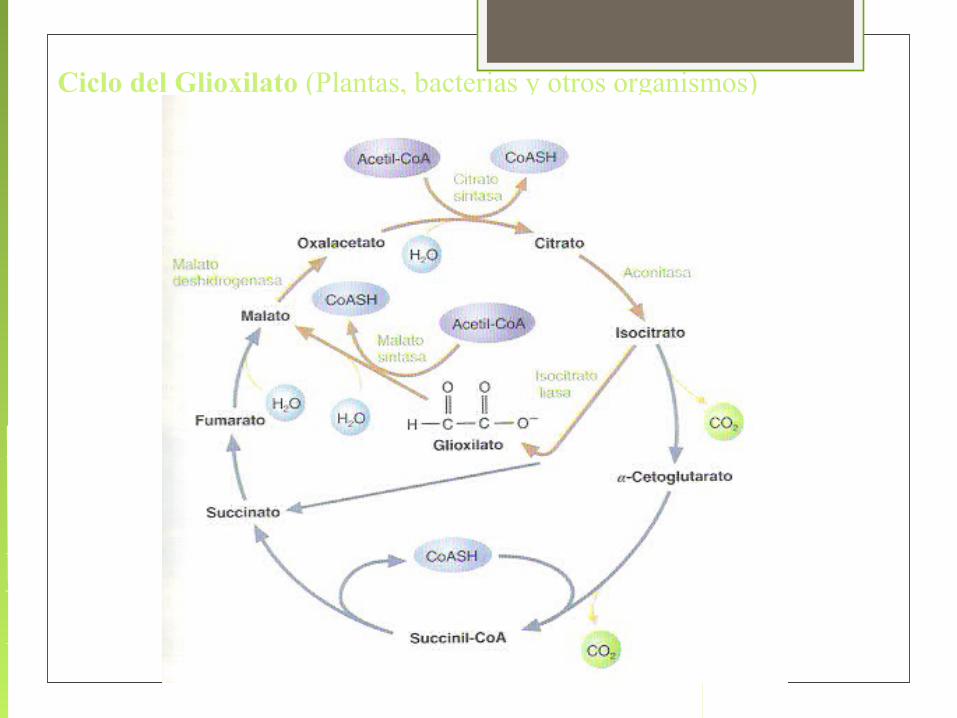
Ciclo del Glioxilato (Plantas, bacterias y otros organismos)
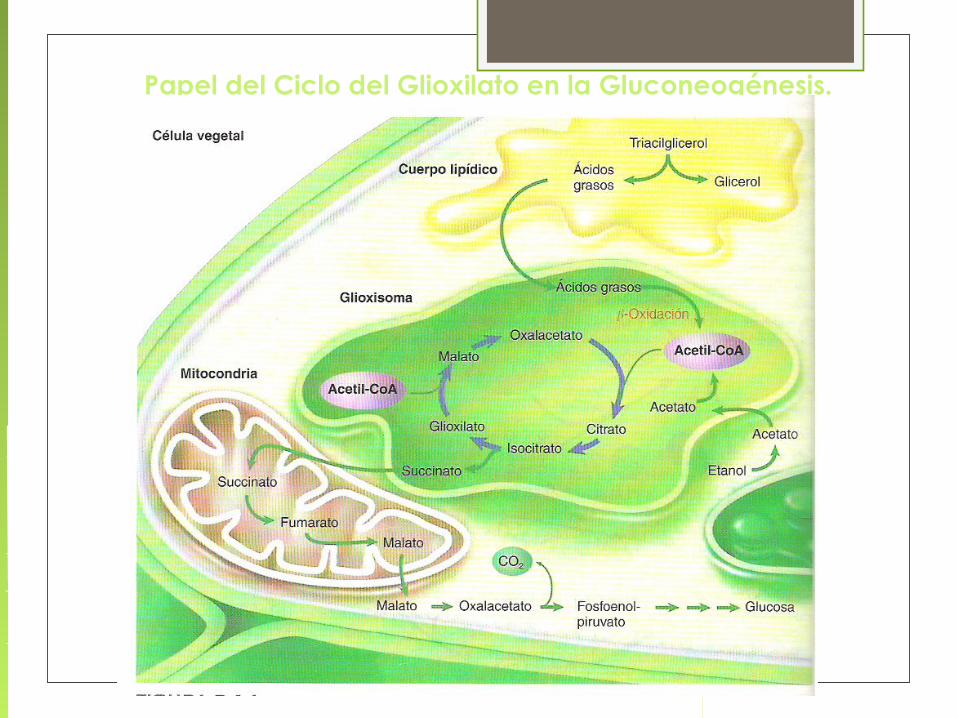
Papel del Ciclo del Glioxilato en la Gluconeogénesis.


















