COMPLEJO DE GOLGI Becker,W.M. y col. 2000 COMPLEJO DE GOLGI.
Upload
sesi-sp-servico-social-da-industriaCategory
view
4.624download
0
COMPONENTES
CELULARES
Do Glicocálix ao Complexo de Golgi

Os limites da célula viva
Uma célula viva é um compartimento microscópico, isolado
do ambiente por pelo menos uma barreira: a membrana
plasmática.
Está é uma película extremamente fina e delicada, que
exerce severa “fiscalização” sobre todas as substâncias e
partículas que entram e saem da célula.

Os limites da célula viva
Dada a relativa fragilidade da membrana plasmática, a
maioria das células apresenta algum tipo de envoltório
que dá proteção e suporte físico à membrana.
Entre esses envoltórios destacam-se o glicocálix, presente
na maioria das células animais, e a parede celulósica,
presente em células de plantas e de algumas algas.

Glicocálix
Se isolássemos uma célula de nosso corpo, notaríamos que ela esta envolta por uma espécie de malha feita de moléculas de glicídios (carboidratos) frouxamente entrelaçadas.
Esta malha protege a célula como uma vestimenta: trata-se do glicocálix (do grego glykys, doce, açúcar, e do latim calyx, casca envoltório).
Diversas funções têm sido sugeridas para o glicocálix. Acredita-se que, além de ser uma proteção contra agressões físicas e químicas do ambiente externo, ele funcione como uma malha de retenção de nutrientes e enzimas, mantendo um microambiente adequado ao redor de cada célula.
Confere às células a capacidade de se reconhecerem, uma vez que células diferentes têm glicocálix formado por glicídios diferentes e células iguais têm glicocálix formado por glicídios iguais.


Parede celulósica
As células vegetais possuem um envoltório externo, espesso
e relativamente rígido: a parede celulósica, também
chamada membrana esquelética celulósica;
Células vegetais jovens apresentam uma parede celulósica
fina e flexível, denominada parede primária.

Parede celulósica
A parede primária é elástica, de modo a permitir o crescimento celular. Depois que a célula cresceu e atingiu o tamanho e a forma definitivos, forma-se a parede secundária, mais espessa e rígida.
A celulose que constitui a parede secundária é secretada através da membrana plasmática, e se deposita entre esta e a superfície interna da parede primária, na qual adere fortemente.

Constituição da parede celulósica
A parede das células vegetais é constituída por longas e resistentes microfibrilas do polissacarídeo celulose.
As microfibrilas celulósicas se mantém unidas por meio de uma matriz formada por glicoproteínas (proteínas ligadas a açucares), hemicelulose e pectina (polissacarídeos).

Constituição da parede celulósica
A estrutura molecular da parede celulósica aplica o
mesmo princípio do concreto armado, no qual
longas e resistentes varetas de ferro estão
mergulhadas em uma argamassa de cimento e
pedras.
Na parede celular, as microfibrilas de celulose
correspondem às varetas de ferro do concreto,
enquanto as glicoproteínas e os polissacarídeos da
matriz correspondem à argamassa.

Membrana plasmática Toda a célula, seja procarionte ou eucarionte, apresenta
uma membrana que isola do meio exterior: a membrana plasmática.
A membrana plasmática é tão fina (entre 6 a 9 nm) que os mais aperfeiçoados microscópios ópticos não conseguiram torná-la visível.
Foi somente após o desenvolvimento da microscopia eletrônica que a membrana plasmática pode ser observada.
Nas grandes ampliações obtidas pelo microscópio eletrônico, cortes transversais da membrana aparecem como uma linha mais clara entre duas mais escuras, delimitando o contorno de cada célula.

Constituição química da membrana
plasmática
Estudos com membranas plasmáticas isoladas
revelam que seus componentes mais abundantes
são fosfolipídios, colesterol e proteínas.
É por isso que se costumam dizer que as
membranas plasmáticas têm constituição
lipoprotéica.

Uma vez identificados os fosfolipídios e as proteínas como os principais
componentes moleculares da membrana, os cientistas passaram a
investigar como estas substâncias estavam organizadas.

O modelo do mosaico fluido
A disposição das moléculas na membrana plasmática foi
elucidada recentemente, sendo que os lipídios formam
uma camada dupla e contínua, no meio da qual se
encaixam moléculas de proteína.
A dupla camada de fosfolipídios é fluida, de consistência
oleosa, e as proteínas mudam de posição continuamente,
como se fossem peças de um mosaico.
Esse modelo foi sugerido por dois pesquisadores, Singer e
Nicholson, e recebeu o nome de Modelo Mosaico Fluido.

O modelo do mosaico fluido
Os fosfolipídios têm a função de manter a estrutura da
membrana e as proteínas têm diversas funções.
As membranas plasmáticas de um eucariócitos contêm
quantidades particularmente grande de colesterol.
As moléculas de colesterol aumentam as propriedades da
barreira da bicamada lipídica e devido a seus rígidos
anéis planos de esteróides diminuem a mobilidade e torna
a bicamada lipídica menos fluida.


Funções das proteínas nas membranas
plasmáticas
As proteínas da membrana plasmática exercem grandes
variedades de funções: atuam preferencialmente nos
mecanismos de transporte, organizando verdadeiros túneis
que permitem a passagem de substâncias para dentro e
para fora da célula, funcionam como receptores de
membrana, encarregadas de receber sinais de substâncias
que levam alguma mensagem para a célula, favorecem a
adesão de células adjacentes em um tecido, servem como
ponto de ancoragem para o citoesqueleto.

Proteínas de adesão: em células adjacentes, as proteínas da membrana podem aderir
umas às outras.
Proteínas que facilitam o transporte de substâncias entre células.
Proteínas de reconhecimento: determinadas glicoproteínas atuam na membrana como
um verdadeiro “selo marcador”, sendo identificadas especificamente por outras células.
Proteínas receptoras de membrana.
Proteínas de transporte: podem desempenhar papel na difusão facilitada, formando
um canal por onde passam algumas substâncias, ou no transporte ativo, em que há gasto
de energia fornecida pela substância ATP. O ATP (adenosina trifosfato) é uma molécula
derivada de nucleotídeo que armazena a energia liberada nos processos
bioenergéticos que ocorrem nas células (respiração aeróbia, por exemplo). Toda vez
que é necessária energia para a realização de uma atividade celular (transporte ativo,
por exemplo) ela é fornecida por moléculas de ATP.
Proteínas de ação enzimática: uma ou mais proteínas podem atuar isoladamente como
enzima na membrana ou em conjunto, como se fossem parte de uma “linha de
montagem” de uma determinada via metabólica.
Proteínas com função de ancoragem para o citoesqueleto.
Funções das proteínas nas membranas plasmáticas

Transporte pela membrana plasmática
A capacidade de uma membrana de ser atravessada por algumas substâncias e não por outras define sua permeabilidade. Em uma solução, encontram-se o solvente (meio líquido dispersante) e o soluto (partícula dissolvida). Classificam-se as membranas, de acordo com a permeabilidade, em 4 tipos:
Permeável: permite a passagem do solvente e do soluto;
Impermeável: não permite a passagem do solvente nem do soluto;
Semipermeável: permite a passagem do solvente, mas não do soluto;
Seletivamente permeável: permite a passagem do solvente e alguns tipos de soluto

Transporte pela membrana plasmática
Nessa última classificação se enquadra a membrana
plasmática.
A passagem aleatória de partículas sempre ocorre de um
local de maior concentração para outro de concentração
menor (a favor do gradiente de concentração).
Isso se dá até que a distribuição das partículas seja
uniforme. A partir do momento em que o equilíbrio for
atingido, as trocas de substâncias entre dois meios tornam-
se proporcionais.

Transporte pela membrana plasmática
A passagem de substâncias através das membranas celulares envolve vários mecanismos, entre os quais podemos citar:
Transporte passivo
Osmose
Difusão simples
Difusão facilitada
Transporte ativo
Bomba de sódio e potássio
Endocitose e exocitose
Fagocitose
Pinocitose

Citoplasma
Os primeiros citologistas acreditavam que o interior da
célula viva era preenchido por um fluído homogêneo e
viscoso, no qual estava mergulhado o núcleo. Esse
fluido recebeu o nome de citoplasma (do grego kytos,
célula, e plasma, aquilo que dá forma, que modela).
Hoje se sabe que o espaço situado entre a membrana
plasmática e o núcleo é bem diferente do que
imaginaram aqueles citologistas pioneiros. Além da
parte fluida, o citoplasma contém bolsas e canais
membranosos e organelas ou orgânulos
citoplasmáticos, que desempenham funções específicas
no metabolismo da célula eucarionte.


Citoplasma O fluido citoplasmático é constituído principalmente por água,
proteínas, sais minerais e açucares.
No citosol ocorre a maioria das reações químicas vitais, entre elas a fabricação das moléculas que irão constituir as estruturas celulares.
É também no citosol que muitas substâncias de reserva das células animais, como as gorduras e o glicogênio, ficam armazenadas.
Na periferia do citoplasma, o citosol é mais viscoso, tendo consistência de gelatina mole.
Essa região é chamada de ectoplasma (do grego, ectos, fora).
Na parte mais central da célula situa-se o endoplasma (do grego, endos, dentro), de consistência mais fluida.

Célula
Animal
Célula
vegetal
Célula
animal

Ciclose
O citosol encontra-se em contínuo movimento, impulsionado pela contração rítmica de certos fios de proteínas presentes no citoplasma, em um processo semelhante ao que faz nossos músculos se movimentarem.
Os fluxos de citosol constituem o que os biólogos denominam ciclose. Em algumas células, a ciclose é tão intensa que há verdadeiras correntes circulatórias internas.
Sua velocidade aumenta com elevação da temperatura e diminui em temperaturas baixas, assim como na falta de oxigênio.

Movimento amebóide
Alguns tipos de células têm a capacidade de alterar rapidamente a consistência de seu citosol, gerando fluxos internos que permitem à célula mudar de forma e se movimentar.
Esse tipo de movimento celular, presente em muitos protozoários e em alguns tipos de células de animais multicelulares, é chamado movimento amebóide.

Como são as organelas?
Algumas das organelas do citoplasma são membranosas,
isto é, são revestidas por uma membrana lipoprotéica
semelhante à membrana plasmática.
Estamos nos referindo a retículo
endoplasmático, mitocôndrias, complexo de golgi,
lisossomos, peroxissomos, glioxissomos, cloroplastos e vacúo
los.
As organelas não membranosas são os ribossomos e os
centríolos.

Retículo endoplasmático
O citoplasma das células eucariontes contém inúmeras
bolsas e tubos cujas paredes têm uma organização
semelhante à da membrana plasmática.
Essas estruturas membranosas formam uma complexa rede
de canais interligados, conhecida pelo nome de retículo
endoplasmático.
Pode-se distinguir dois tipos de retículo: rugoso (ou
granular) e liso (ou agranular).

Retículo endoplasmático
rugoso (RER) e liso (REL)
O retículo endoplasmático rugoso (RER), também
chamado de ergastoplasma, é formado por sacos
achatados, cujas membranas têm aspecto verrugoso
devido à presença de grânulos – os ribossomos –
aderidos à sua superfície externa (voltada para o
citosol).
Já o retículo endoplasmático liso (REL) é formado
por estruturas membranosas tubulares, sem
ribossomos aderidos, e, portanto, de superfície lisa.

Os dois tipos de retículo estão interligados e a transição entre eles é gradual.
Se observarmos o retículo endoplasmático partindo do retículo rugoso em direção ao liso, vemos
as bolsas se tornarem menores e a quantidade de ribossomos aderidos diminuir progressivamente,
até deixar de existir.

Funções do retículo endoplasmático
O retículo endoplasmático atua como uma rede de
distribuição de substâncias no interior da célula.
No líquido existente dentro de suas bolsas e tubos,
diversos tipos de substâncias se deslocam sem se
misturar com o citosol.

Produção de lipídiosUma importante função de retículo endoplasmático liso é a produção de lipídios. A lecitina e o colesterol, por exemplo, os principais componentes lipídicos de todas as membranas celulares são produzidos no REL. Outros tipos de lipídios produzidos no retículo liso são os hormônios esteróides, entre os quais estão a testosterona e os estrógeno, hormônios sexuais produzidos nas células das gônadas de animais vertebrados.
DesintoxicaçãoO retículo endoplasmático liso também participa dos processos de desintoxicação do organismo. Nas células do fígado, o REL, absorve substâncias tóxicas, modificando-as ou destruindo-as, de modo a não causarem danos ao organismo. É a atuação do retículo das células hepáticas que permite eliminar parte do álcool, medicamentos e outras substâncias potencialmente nocivas que ingerimos.
Armazenamento de substânciasDentro das bolsas do retículo liso também pode haver armazenamento de substâncias. Os vacúolos das células vegetais, por exemplo, são bolsas membranosas derivadas do retículo que crescem pelo acúmulo de soluções aquosas ali armazenadas.
Produção de proteínasO retículo endoplasmático rugoso, graças à presença dos ribossomos, é responsável por boa parte da produção de proteínas da célula. As proteínas fabricadas nos ribossomos do RER penetram nas bolsas e se deslocam em direção ao aparelho de Golgi, passando pelos estreitos e tortuosos canais co retículo endoplasmático liso.

Vacúolos
Os vacúolos das células vegetais são interpretados com regiões expandidas do retículo endoplasmático.
Em células vegetais jovens observam-se algumas dessas regiões, formando pequenos vacúolos isolados um do outro.
Mas, à medida que a célula atinge a fase adulta, esses pequenos vacúolos se fundem, formando-se um único, grande e central, com ramificações que lembram sua origem reticular.
A expansão do vacúolo leva o restante do citoplasma a ficar comprimido e restrito à porção periférica da célula. Além disso, a função do vacúolo é regular as trocas de água que ocorrem na osmose.

Vacúolos
Em protozoários de água doce existem vacúolos
pulsáteis (também chamados contráteis), que
exercem o papel de reguladores osmóticos.
O ingresso constante de água, do meio para o
interior da célula, coloca em risco a integridade
celular.
A remoção contínua dessa água mantém constante a
concentração dos líquidos celulares e evita riscos de
rompimento da célula. É um trabalho que consome
energia.


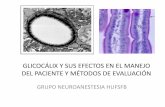
Aparelho de Golgi
A denominação aparelho ou complexo de Golgi é uma homenagem ao citologista italiano Camilo Golgi, que, em 1898, descobriu essa estrutura citoplasmática.
Ao verificar que certas regiões com citoplasma celular se coravam por sais de ósmio de prata, Golgi imaginou que ali deveria existir algum tipo de estrutura, posteriormente confirmada pela microscopia eletrônica.
O aparelho de Golgi está presente em praticamente todas as células eucariontes, e consiste de bolsas membranosas achatadas, empilhadas como pratos. Cada uma dessas pilhas recebe o nome dedictiossomo.
Nas células animais, os dictiossomos geralmente se encontram reunidos em um único local, próximo ao núcleo. Nas células vegetais, geralmente há vários dictiossomos espalhados pelo citoplasma.


Funções do aparelho de Golgi
O aparelho de Golgi atua como centro de armazenamento, transformação, empacotamento e remessa de substâncias na célula.
Muitas das substâncias que passam pelo aparelho de Golgi serão eliminadas da célula, indo atuar em diferentes partes do organismo. É o que ocorre, por exemplo, com as enzimas digestivas produzidas e eliminadas pelas células de diversos órgãos (estômago, intestino, pâncreas, etc).
Outras substâncias, tais como o muco que lubrifica as superfícies internas do nosso corpo, também são processadas e eliminadas pelo aparelho de Golgi.
Assim, o principal papel dessa estrutura citoplasmática é a eliminação de substâncias que atuam fora da célula, processo genericamente denominado secreção celular.

Secreção de enzimas digestivas
As enzimas digestivas do pâncreas, por exemplo, são produzidas no RER e levadas até as bolsas do aparelho de Golgi, onde são empacotadas em pequenas bolsas, que se desprendem dos dictiossomos e se acumulam em um dos pólos da célula pancreática.
Quando chega o sinal de que há alimento para ser digerido, as bolsas cheias de enzimas se deslocam até a membrana plasmática, fundem-se com ela e eliminam seu conteúdo para o meio exterior.
A produção de enzimas digestivas pelo pâncreas é apenas um entre muitos exemplos do papel do aparelho de Golgi nos processos de secreção celular. Praticamente todas as células do corpo sintetizam e secretam uma grande variedade de proteínas que atuam fora delas.


Acrossomo do espermatozóide
O aparelho de Golgi desempenha um papel importante na formação dos espermatozóides.
Estes contêm bolsas repletas de enzimas digestivas, que irão perfurar as membranas do óvulo e permitir a fecundação.
A bolsa de enzimas do espermatozóide maduro, originada no aparelho de Golgi, é o acrossomo (do grego acros, alto, topo, e somatos, corpo), termo que significa “corpo localizado no topo do espermatozóide”.


Formação da lamela média em
vegetais
Nas células vegetais o complexo de Golgi participa ativamente da formação da lamela média, a primeira membrana que separa duas células recém-originadas na divisão celular.
Os dictiossomos acumulam o polissacarídeo pectina, que é eliminado entre as células irmãs recém formadas, constituindo a primeira separação entre elas e, mais tarde, a lâmina que as mantém unidas.