
EFECTOS DEL CLORURO DE LITIO SOBRE LA LOCALIZACIÓN ...
78
Tesina para optar por el grado de Licenciado en Ciencias Biológicas Profundización en Biología Celular EFECTOS DEL CLORURO DE LITIO SOBRE LA LOCALIZACIÓN SUBCELULAR DE MARCKS DURANTE LA NEURULACIÓN EN EMBRIONES DE POLLO ANA MAITE FOLLE LOPEZ Docente Orientador: Dr. Flavio R. Zolessi Sección Biología Celular Tribunal: Dra. Cristina Arruti Dra. Nibia Berois Dr. Flavio R. Zolessi Montevideo, Uruguay Febrero 2012
Transcript of EFECTOS DEL CLORURO DE LITIO SOBRE LA LOCALIZACIÓN ...

Tesina para optar por el grado de Licenciado en Ciencias Biológicas
Profundización en Biología Celular
EFECTOS DEL CLORURO DE LITIO SOBRE LA LOCALIZACIÓN SUBCELULAR DE MARCKS DURANTE LA NEURULACIÓN EN
EMBRIONES DE POLLO
ANA MAITE FOLLE LOPEZ
Docente Orientador: Dr. Flavio R. Zolessi
Sección Biología Celular
Tribunal:
Dra. Cristina Arruti
Dra. Nibia Berois
Dr. Flavio R. Zolessi
Montevideo, Uruguay
Febrero 2012

A Josefina…

Agradecimientos __________________________________________________
A mi orientador, Flavio Zolessi, quién permitió que este trabajo fuera posible
A Cristina Arruti, por abrirme las puertas del laboratorio
A mi amigo y compañero de tesis, Gonzalo Aparicio, con el que compartimos
gran parte de este trabajo
A mis colegas, Andrea Toledo, Daniel Prieto y Lucía Ruiz por el buen humor y su
alegre compañía
A Luis, mi padre, mi madre y mi hermano por su apoyo incondicional
A avícola Prodhin, por proveernos de huevos embrionados

Indice __________________________________________________
I Resumen 1
II Introducción 2
i Origen del Sistema Nervioso: cerebro y médula espinal 2
ii Proceso de formación del tubo neural 2
iii Neurulación primaria 4
a - Formación de la placa neural 5
b - Modelado de la placa neural 5
c - Flexión de la placa neural 6
d - Cierre del tubo neural 6
iv Defectos en el cierre del tubo neural 9
v MARCKS 10
a - MARCKS en el desarrollo del sistema nervioso 11
b - MARCKS y defectos en el cierre del tubo neural 11
vi Efectos celulares del cloruro de litio 12
vii Nuestro estudio 13
III Objetivos 14
IV Resultados 15
i Neurulación en condiciones de cultivo 15
a - Distribución de MARCKS 18
ii Tratamientos farmacológicos de embriones en condiciones de
cultivo durante la neurulación 20
a - Tratamiento con 100 mM de cloruro de litio 21
b - Tratamiento con 60 mM de cloruro de litio 30
c - Tratamiento con 20 mM de cloruro de litio 33
iii Efectos del cloruro de litio sobre MARCKS 37
V Discusión 43
i Modelo de estudio y método de cultivo de embriones 43
ii Tratamientos farmacológicos de embriones en condiciones de
cultivo durante la neurulación 45
a - Efectos del cloruro de litio sobre la neurulación 47
b - Efectos del cloruro de litio sobre la muerte celular 48

c - Efectos del cloruro de litio sobre la somitogénesis 49
d - Efectos del cloruro de litio sobre MARCKS 49
e - Mecanismo de acción del cloruro de litio 50
VI Conclusiones 53
VII Materiales y métodos 54
i Obtención de embriones 54
ii Cultivo de embriones 54
iii Tratamiento farmacológico 57
iv Cortes a congelación 58
v Inmunodetección sobre cortes 58
vi Análisis de imágenes 60
VIII Apéndice 61
IX Bibliografía 63

1
I Resumen __________________________________________________
La construcción del sistema nervioso es uno de los eventos más fascinantes del
desarrollo embrionario. El proceso comienza con la formación, modelado y
flexión de la placa neural para dar lugar al cierre del tubo neural, a partir del cual
se formará el cerebro y la médula espinal. Las fallas durante este proceso se
conocen como NTDs y pueden ser influenciadas por factores genéticos y
ambientales. Dentro de los últimos se encuentran las drogas anti-depresivas y
para el tratamiento de trastornos bipolares, como el litio. Se estudió la acción del
cloruro de litio sobre el proceso de neurulación utilizando el embrión de pollo
como modelo experimental. Paralelamente, se intentó determinar una posible
correlación entre la localización subcelular de la proteína MARCKS y NTDs
producidos por exposición a este fármaco. Los tratamientos farmacológicos
incluyeron ensayos con 3 dosis de cloruro de litio (20, 60 y 100 mM) en
embriones tempranos de pollo en cultivo, utilizando el método EC y una nueva
forma de aplicación de la droga por disolución en el medio de cultivo. Los
resultados obtenidos muestran que el efecto del cloruro de litio es dosis-, tiempo-
y estadio-dependiente. El proceso de neurulación fue afectado por las dosis más
altas como también el proceso de gastrulación cuando los embriones fueron
tratados en etapas más tempranas del desarrollo. La dosis más alta provocó un
aumento significativo de muerte celular en el tubo neural. El cloruro de litio no
afectó la abundancia ni localización subcelular de MARCKS durante el cierre del
tubo neural con conservación de su apicalización en el neuroepitelio en los
tratamientos a corto plazo. Se observó un aumento en la inactivación por
fosforilación de la enzima GSK3β. Esta enzima es uno de los blancos propuestos
para el litio y mediaría su acción estabilizadora del ánimo. Actualmente los
mecanismos de acción del litio no se comprenden en su totalidad y es de suma
importancia profundizar su conocimiento debido al acotado rango que existe
entre dosis terapéuticas y teratogénicas.

2
II Introducción __________________________________________________
La vida de un animal comienza con una sola célula, el huevo fecundado. Durante
el desarrollo, las repetidas divisiones celulares producen células muy diferentes
entre sí hasta finalmente configurar un patrón de complejidad y precisión
espectaculares. En último término el genoma determina este patrón y el
rompecabezas de la biología del desarrollo consiste en entender cómo suceden
estos fenómenos (Alberts et al., 2003). Dentro de los temas más fascinantes en
la formación de un nuevo individuo se encuentra el nacimiento y la construcción
de lo que se convertirá en la red más compleja y completa que existe: el sistema
nervioso.
i Origen del Sistema Nervioso: cerebro y médula espinal
El sistema nervioso queda definido muy tempranamente en el desarrollo durante
un proceso conocido como neurulación. Tanto el cerebro como la médula espinal
derivan del tubo neural, una estructura que se forma durante la embriogénesis,
gracias a una secuencia muy coordinada de eventos morfogenéticos (Karfunkel,
1974; Schoenwolf, 1982, 1994; Gordon, 1985; Copp et al., 1990; Schoenwolf y
Smith, 1990; Jacobson, 1991). La formación del tubo neural es uno de los
primeros y más críticos eventos que suceden durante el desarrollo del sistema
nervioso central en los vertebrados (Sadler, 1998). Su precursor embriológico es
la placa neural, o neuroepitelio, una región más gruesa del ectodermo dorsal
ubicada en la superficie del embrión (Wallingford, 2005; Sanes et al., 2006).
ii Proceso de formación del tubo neural
Existen dos modos principales de convertir la placa neural en un tubo neural. La
neurulación primaria se produce en la porción anterior del embrión y permite la
formación del tubo neural que dará origen al cerebro y la mayor parte de la
médula espinal. En vertebrados este proceso involucra la formación y el plegado
del neuroepitelio dorsal. Las células que rodean a la placa neural estimulan a las
células de la misma a proliferar, invaginarse y separarse de la superficie por
fusión de los pliegues neurales, para formar un tubo hueco (Weil et al., 1997). De
esta manera se crean dos capas epiteliales: el ectodermo superficial y el tubo

3
neural (Copp et al., 1990; Colas y Schoenwolf, 2001; Copp et al., 2003b) (Figura
1).
Figura 1.- Imágenes de embriones de pollo in toto (derecha) y cortes transversales (izquierda) obtenidos con microscopía electrónica de barrido en diferentes etapas de la neurulación primaria. Los cortes transversales fueron realizados a nivel del futuro mesencéfalo; las imágenes B y C corresponden al mismo embrión pero a niveles distintos del eje céfalo-caudal. A. Etapa de modelado y flexión temprana de la placa neural. Se observa el punto bisagra medial (mhp). B. Estadio de surco neural. C. Estadio de tubo neural inicial, los pliegues neurales ya están en contacto pero no se han fusionado aún. D. Estadio de tubo neural definitivo; los pliegues neurales ya se han fusionado formando el techo del tubo neural, la cresta neural (flechas) y el ectodermo epidérmico dorsal medio. dlph, punto bisagra dorsolateral; e, endodermo: ee, ectodermo epidermal; fg, intestino anterior; hm, mesodermo de la cabeza; mhp, punto bisagra medial; n, notocorda; nf, pliegue neural; np, placa neural (Colas y Schoenwolf, 2001).
En la neurulación secundaria, que generalmente se produce en la porción
posterior del embrión, se forma el tubo neural que dará origen a la porción más
caudal de la médula espinal. En este caso el tubo neural se origina a partir de la
unión de células mesenquimáticas para formar un cordón sólido que
posteriormente se ahueca. Esta canalización del epitelio crea el tubo neural
secundario, cuyo lumen es continuo con el del tubo neural primario (Schoenwolf
y Franks, 1984; Copp y Brook, 1989) (Figura 2). De esta manera, el tubo neural
completo se forma por la unión de estos dos tubos formados separadamente
(Sanes et al., 2006; Gilbert, 2006).

4
Figura 2.- Micrografías electrónicas de un embrión de pollo in toto (A), corte parasagital (B), criofractura transversal (C, E) y sección transversal (D) ilustrando la neurulación secundaria. Las tres imágenes transversales muestran etapas progresivas en el desarrollo del tubo neural secundario. La línea en A indica la posición de los cortes parasagitales que se muestran en B. e, endodermo; ee, ectodermo epidérmico; ic, células internas del cordón medular que van a ser removidas durante la cavitación; mc, cordón medular sometido a cavitación; n, notocorda; oc, células externas del cordón medular que formarán el neuroepitelio secundario; p, lumen primario formado a partir del surco neural; pnt, terminación caudal del tubo neural primario; s, lumen secundario formado a partir de la cavitación del cordón medular; tb, botón de cola (Colas y Schoenwolf, 2001).
iii Neurulación primaria
A nivel tisular, la neurulación primaria consta de cuatro etapas: formación,
modelado y flexión de la placa neural sumado al cierre del tubo neural (Colas y
Schoenwolf 2001; Pulikkunnel y Thomas, 2005; Wallingford, 2005) (Figuras 1 y
3). La neurulación se inicia con la formación de la placa neural, gracias a un
proceso que se describe como inducción neural (Lemaire y Kodjabachian, 1996;
Tanabe y Jessell, 1996; Gould y Grainger, 1997; Hemmati-Brivanlou y Melton,
1997; Sasai y De Robertis, 1997; Weinstein y Hemmati-Brivanlou, 1999; Harland,
2000; Jessell y Sanes, 2000). Estudios recientes muestran que la inducción
neural involucra la supresión del destino epidérmico más que la inducción del
destino neural en sí. La señal de supresión es generada por el nodo de Hensen
(el equivalente en aves al organizador de Spemann) (Figura 3A) e involucra la
unión de moléculas inhibitorias a ligandos secretados, tales como BMPs o Wnts,
bloqueando la señal (Colas y Schoenwolf 2001; Stern, 2002).

5
a - Formación de la placa neural
En humanos, ratones, anfibios, aves y la mayoría de los vertebrados la
construcción de todo el sistema nervioso comienza con la formación de la placa
neural debido a un engrosamiento del ectodermo dorsal (Greene y Copp, 2009)
(Figura 3A). Esto sucede cuando el mesodermo dorsal subyacente (y el
endodermo faríngeo de la cabeza) señaliza a las células suprayacentes a fin de
elongar las células de la placa neural a columnares. Esta forma elongada
distingue a las células de la placa neural prospectiva de las células más planas
pre-epidérmicas que las rodean. Aproximadamente el 50% del ectodermo es
incluido en la placa neural (Wallingford, 2005) la cual se extiende a lo largo del
eje antero-posterior y se estrecha hacia el extremo caudal. Dicha placa se halla
constituida por células polarizadas, en las cuales la actina y la miosina se
acumulan en el borde apical del epitelio. Los movimientos coordinados de
invaginación, elevación y convergencia se deben a la acción de fuerzas
extrínsecas e intrínsecas, estas últimas dirigidas por el citoesqueleto de actina
(Schoenwolf, 1991; Smith y Schoenwolf, 1997).
b - Modelado de la placa neural
Durante el modelado, la placa neural continúa engrosándose a nivel ápico-basal.
Adicionalmente, se somete a movimientos de extensión convergente que
permiten que la placa se estreche a nivel medio-lateral y se alargue a nivel
céfalo-caudal (Figura 3B, C). Experimentos de aislamiento han demostrado que
el modelado está guiado por cambios en el comportamiento de las células del
neuroepitelio (Schoenwolf, 1988; Schoenwolf y Alvarez, 1989; Moury y
Schoenwolf, 1995). Aunque los procesos de neurulación y gastrulación pueden
ser experimentalmente desacoplados, la formación completa de la porción
rostro-caudal de la placa neural así como su extensión requieren movimientos de
gastrulación normales (especialmente la regresión de la línea primitiva) (Colas y
Schoenwolf, 2001).

6
c - Flexión de la placa neural
Para que el proceso de flexión se lleve a cabo debe ocurrir elevación y
convergencia de los pliegues neurales hacia la línea dorsal media (Figura 3B-D).
En un comienzo, las células de la línea media de la placa neural se anclan a la
notocorda por debajo de ellas, recibiendo el nombre de células del punto bisagra
medio. Su formación requiere una señal inductiva proveniente de la notocorda
subyacente (Smith y Schoenwolf, 1989; Nieuwkoop, 1999; Bachiller et al., 2000).
Esta señal, mediada por la proteína secretada Sonic hedgehog, también está
involucrada en la formación de la placa del piso del tubo neural, un importante
centro de señalización (Jessell y Sanes, 2000). Poco después, otras dos
regiones laterales denominadas puntos bisagra dorsolaterales forman surcos
cerca de la conexión de la placa neural con el resto del ectodermo. Las células
en estos tres puntos aumentan su altura y adquieren forma de cuña, permitiendo
que las bisagras actúen como un eje que dirige la rotación de las células
alrededor de éstas llevando a que la placa se flexione. La elevación en el punto
bisagra medio produce un espacio llamado surco neural, el cual se convertirá en
el lumen del tubo neural primitivo luego del cierre (Wallingford, 2005). Los
pliegues neurales también juegan un importante rol durante el plegado de la
placa neural. Cada pliegue es bilaminar y consiste en una capa de neuroepitelio
cubierta por una capa de ectodermo epidérmico (Colas y Schoenwolf, 2001).
Experimentos de aislación han demostrado que el plegamiento de la placa
neural no sólo es dirigido por cambios en el comportamiento en las células
neuroepiteliales (Schoenwolf, 1988; Moury y Schoenwolf, 1995). Las fuerzas que
dirigen el plegado son generadas, en gran medida, por el ectodermo epidérmico
y consisten en el aplanamiento, intercalación celular y orientación de las mitosis
(Schoenwolf y Alvarez, 1991; Sausedo et al., 1997) que permiten que el
ectodermo se expanda medialmente (Lawson et al., 2001).
d - Cierre del tubo neural
El tubo neural se cierra a medida que el par de pliegues neurales es llevado
hacia la línea dorsal media. Los pliegues se adhieren entre sí y las células a
partir de ambos pliegues se fusionan. El cierre no se produce simultáneamente a

7
lo largo de todo el ectodermo. Esto se observa mejor en aquellos vertebrados
(aves y mamíferos) cuyo eje corporal es alargado antes de la neurulación. En
amniotas, la inducción en la cabeza comienza antes que en el tronco. Por tanto,
en un embrión de pollo de 24 horas coexisten una avanzada neurulación en la
región cefálica con la gastrulación en la región caudal (Sadler, 2000). El tubo
neural finalmente forma un cilindro cerrado que se separa del ectodermo
superficial. Se piensa que esta separación está mediada por la expresión de
diferentes moléculas de adhesión (Gilbert, 2006). La fusión establece el techo
del tubo neural y lo separa del ectodermo epidérmico que lo cubre, el cual
contribuirá a formar la piel dorsal del embrión. Los dos extremos del tubo neural
que permanecen abiertos en esta etapa son denominados neuroporo anterior y
neuroporo posterior. Una tercera población de células ectodérmicas se forma a
medida que los pliegues neurales se elevan y convergen (en el ratón) o durante
la fusión (en el pollo). Estas son las células precursoras de la cresta neural que
se originan en la interfase entre la placa neural y la epidermis prospectiva y
provienen de ambos tejidos (Selleck y Bronner-Fraser, 1995). La cresta neural es
una importante población celular que contribuye a la formación del sistema
nervioso periférico y otras estructuras (células pigmentarias, esqueleto facial,
entre otras) (Anderson, 1999; García-Castro y Bronner-Fraser, 1999; Groves y
Bronner- Fraser, 1999; Hall, 1999; Le Douarin y Kalcheim, 1999).

8
Figura 3.- Embriones de pollo in toto durante la neurulación primaria, vistos desde la superficie dorsal del blastodermo. Los estadios que se muestran (HH4-11) fueron establecidos por Hamburger y Hamilton (1951) y representan 24 horas de desarrollo. A. La placa neural recién se ha formado. B. La placa neural durante el modelado. C. Comienzo de la flexión de la placa neural, simultánea al modelado, donde se establece el surco neural. D. Contacto entre los pliegues neurales a nivel de la región del futuro mesencéfalo del tubo neural (flecha). E. Se ha formado el tubo neural a nivel del futuro cerebro y mayor parte de la futura médula espinal. Se delinean los bordes aproximados de la placa neural para cada caso (A-C). hn, nodo de Hensen; n, notocorda; nf, pliegue neural; ng, surco neural; np, placa neural; nt/b, nivel del futuro cerebro en el tubo neural; nt/sc, nivel de la futura médula espinal en el tubo neural; ps, línea primitiva (Colas y Schoenwolf, 2001).
Figura 4.- Resumen del proceso de neurulación primaria en el pollo. A la izquierda se muestran cortes transversales y a la derecha se representa esquemáticamente la secuencia de eventos que suceden durante el cierre del tubo neural (Gilbert, 2006).

9
iv Defectos en el cierre del tubo neural
Cuando ocurren fallas en la neurulación se producen malformaciones a lo largo
del eje neural que pueden interrumpir la diferenciación del sistema nervioso
central y la inducción de los arcos vertebrales. Estas son colectivamente
conocidas como defectos en el cierre del tubo neural (NTDs) (Inagaki et al.,
2000; Pulikkunnel y Thomas, 2005). Con una incidencia aproximada de 1 en
1000 en humanos, es la segunda causa más común de defectos de nacimiento
luego de los defectos cardíacos congénitos (Afman et al., 2003; Detrait et al.,
2005). En humanos, los NTDs más frecuentes son anencefalia y
mielomeningocele (Wallingford, 2005). La anencefalia resulta de un defecto en el
cierre del extremo rostral del tubo neural y se caracteriza por una ausencia total
o parcial de la bóveda craneana y hemisferios cerebrales. El mielomeningocele
también es causado por el cierre defectuoso del tubo neural pero, en este caso,
a nivel de la columna vertebral. Los pacientes afectados pueden sufrir
consecuencias leves a severas, dependiendo del tamaño y localización del
defecto (Tolmie, 1996). La predisposición genética y los factores ambientales
juegan un rol importante en la etiología de los NTDs (Daly et al., 1997; Detrait et
al., 2005; Dhaulakhandi et al., 2010). Desórdenes genéticos asociados a NTDs
incluyen mutaciones en un único gen (ej: Síndrome de Meckel) y anomalías
cromosómicas (ej: trisomías 13 y 18). Sin embargo, los NTDs más comunes
tienen herencia multifactorial en la cual la predisposición genética es poligénica y
está influenciada por interacciones entre genes (Pulikkunnel y Thomas, 2005).
Como lo revelan varios estudios epidemiológicos, los factores ambientales
también pueden incidir y son numerosos los teratógenos y deficiencias
nutricionales que se han sugerido como posibles agentes causales (Afman et al.,
2003). Se ha descrito un aumento en el riesgo de NTDs en respuesta a
diferentes agresiones ambientales. Entre las más dramáticas, se encuentra el
uso de medicamentos anti-convulsivos o para tratamientos de desórdenes
bipolares (enfermedad maníaco-depresiva) por parte de la madre.

10
v MARCKS
MARCKS (Myristoylated Alanine-Rich C Kinase Substrate) es una proteína
perteneciente a una familia compuesta por dos miembros: MARCKS y MRP
(MARCKS Related Protein). Se trata de una proteína pequeña, intrínsecamente
desplegada, ácida, rica en alanina, glicina, prolina y ácido glutámico. Es un
sustrato mayor de la proteína quinasa C (PKC). Posee tres regiones altamente
conservadas, la región aminoterminal, la región MH2 de función desconocida, y
el dominio efector (DE), o dominio de fosforilación (Stumpo et al., 1989;
Blackshear et al., 1992). El dominio amino terminal está formado por los
primeros catorce aminoácidos que conforman la secuencia consenso para su
miristilación (modificación co-traduccional por la cual se adiciona un ácido
mirístico al grupo amino de la glicina en el extremo N-terminal mediante un
enlace amida) (Towler et al., 1988). El dominio MH2 posee el único sitio de
empalme del ARNm (Blackshear et al., 1992). Finalmente el DE es una
secuencia de 25 aminoácidos (Graff et al., 1989a; Graff et al., 1991), es
extremadamente básico a diferencia del resto de la proteína, y contiene las
serinas fosforilables por PKC. Esta proteína presenta un tamaño variable
dependiendo de la especie. En el pollo, la secuencia de MARCKS consta de 280
aminoácidos y un peso molecular de 27.8 kDa (Graff et al., 1989a; Zolessi et al.,
2004). Se trata de una proteína de membrana ubicua particularmente abundante
en el tejido nervioso (Albert et al., 1986). Su concentración también es alta en
bazo y pulmón, moderada en testículo, páncreas, glándula adrenal, riñón e
hígado, y muy baja en corazón y músculo esquelético (Albert et al., 1987).
Originalmente fue identificada como el principal sustrato de la proteína quinasa C
(PKC) (Albert et al., 1986) y desde entonces ha sido implicada en un gran
número de procesos celulares. Entre ellos, se destaca la regulación de la
dinámica del citoesqueleto cortical de actina en eventos como el desarrollo del
sistema nervioso, la coordinación de los eventos de señalización membrana-
citoesqueleto tales como la migración celular y la adhesión, así como en los
procesos de fagocitosis, endocitosis y exocitosis (revisión en Arbuzova et al.,
2002) y crecimiento de las neuritas en una amplia variedad de células (Aderem,
1992; Blackshear, 1993). MARCKS es sustrato de distintas quinasas de la familia
de la PKC y de quinasas dirigidas por prolina (MAPK y Cdks) e interacciona de

11
manera excluyente con varias moléculas tales como la calmodulina unida a
calcio (Ca-CaM), los fosfolípidos de membrana cargados negativamente y los
microfilamentos de actina. La proteína defosforilada entrecruza los filamentos de
actina, lo que puede ser inhibido por la unión de calmodulina o por la
fosforilación mediada por PKC. La fosforilación no impide la unión a la actina,
pero la torna más laxa lo cual impide el entrecruzamiento de los microfilamentos
(Hartwig et al., 1992) y provoca su translocación al citosol haciendo que
disminuya en la fracción particulada (Rosen et al., 1990).
a - MARCKS en el desarrollo del sistema nervioso
Análisis realizados en ratones knockout demostraron que se trata de una
proteína esencial en el desarrollo normal del sistema nervioso central así como
también en la formación de la retina (Stumpo et al., 1995; Blackshear, 1997). Si
bien está altamente expresada en el cerebro y en médula de animales adultos,
su expresión es aún mayor en cerebros de rata fetales (Blackshear et al., 1986;
Albert et al., 1986; Lobach et al., 1993). Se ha documentado que la disrupción
completa de la expresión de esta proteína (o de MRP) en el ratón, resulta en una
falla en el desarrollo del sistema nervioso central (NTDs) produciendo muerte
perinatal. Esto sugiere que la expresión de MARCKS durante la embriogénesis
en el ratón es necesaria para el desarrollo normal del sistema nervioso central
(Stumpo et al., 1995; Afman et al., 2003). Por otra parte, resultados previos
mostraron que MARCKS se acumula transitoriamente en la región apical de las
células neuroepiteliales durante el plegamiento del tubo neural (Zolessi y Arruti,
2001). Sin embargo, aunque se propone una posible regulación "upstream" por
MARCKS en algunas funciones del citoesqueleto cortical de actina, se
desconoce la función celular de la proteína en este proceso, así como los
mecanismos que llevan a su acumulación apical.
b - MARCKS y defectos en el cierre del tubo neural
Como mencionamos anteriormente, existe un incremento en el riesgo de NTDs
en respuesta a diferentes agresiones ambientales. Dentro de los numerosos
teratógenos estudiados se encuentran los medicamentos anti-convulsivos o
destinados al tratamiento de desórdenes bipolares. Se ha documentado que la

12
administración de estos medicamentos a madres embarazadas induce
malformaciones congénitas. Exposiciones al valproato (fármaco antiepiléptico)
durante el embarazo están asociadas a un aumento cercano al triple en la tasa
de anormalidades mayores, tales como espina bífida (llega a tener una
incidencia de hasta el 2%) y en casos raros anencefalia. El valproato funciona
como teratógeno en la mayoría de las especies estudiadas pero el embrión
humano parece ser el más susceptible (Ornoy, 2009; Watson et al., 1998). Se ha
demostrado que este teratógeno reduce la concentración de MARCKS en células
inmortales del hipocampo (Watson et al., 1998). Otro compuesto muy utilizado
para el tratamiento de desórdenes bipolares es el litio. Administraciones crónicas
de cloruro de litio (LiCl) en rata reducen marcadamente los niveles de MARCKS
de la porción soluble del hipocampo y este efecto persiste luego de terminado el
tratamiento. Sin embargo, no es claro aún si este cambio aparente en los niveles
de MARCKS se debe a una reducción de la concentración de la proteína o
simplemente a un cambio en la distribución subcelular de la misma (Lenox y
Watson, 1994). Aunque se desconoce el mecanismo exacto, algunas evidencias
indican que el efecto de estas drogas sobre MARCKS podría ser indirecto.
vi Efectos celulares del litio
A pesar de las importantes aplicaciones clínicas que posee el litio, todavía no ha
sido dilucidado el mecanismo molecular por el cual ejerce su efecto terapéutico
en humanos o modelos animales. A lo largo de los años se ha demostrado su
capacidad de inhibir varias enzimas, dentro de las cuales se encuentran
monofosfatasas de inositol (Berridge et al., 1989) y la glucógeno sintasa kinasa 3
(GSK3) (Stambolic et al., 1996; Klein y Melton, 1996). Sin embargo, debido a la
falta de modelos animales para trastornos bipolares, los efectos de la inhibición
enzimática en la acción terapéutica del litio no se conocen en profundidad
(Beaulieu y Caron, 2008).

13
vii Nuestro estudio…
En este trabajo nos enfocamos en el estudio de las fallas en la neurulación
primaria que resultan en defectos en el cierre del tubo neural (NTDs), en
particular al estudio del efecto del LiCl sobre este proceso en el embrión
temprano de pollo.
Se trata de un modelo experimental muy utilizado para el estudio de los procesos
tempranos del desarrollo (Darnell y Schoenwolf, 2000a), que se corresponde con
el primer mes del desarrollo embrionario de mamíferos (Dalgic et al., 2009). A su
vez, las zonas de neurulación primaria y secundaria se solapan en forma
bastante similar a los humanos (Schoenwolf y Delongo 1980; Kobus et al., 2009).
El desarrollo in ovo ocurre en ausencia de factores maternos, convirtiéndolo en
un modelo sencillo y especialmente adecuado para la investigación de fármacos
durante el desarrollo de los embriones. Los tejidos neurales en el embrión de
pollo muestran una variación en el grado de desarrollo a lo largo del eje céfalo-
caudal. Este hecho lo torna un muy buen modelo experimental para el estudio de
los efectos producidos por agentes químicos durante el proceso de cierre del
tubo neural (Dalgic et al., 2009). En este modelo se han logrado identificar
múltiples causantes de NTDs dentro de los cuales se encuentran la cotinina
(metabolito de la nicotina) (Dalgic et al., 2009), diazepam (Guney et al., 1999),
cadmio (Fernández et al., 2004), anestésicos (Lee y Nagele, 1985) altas dosis de
progesterona (Erdincler et al., 2009), entre otros.

14
III Objetivos __________________________________________________
El objetivo general de este proyecto es contribuir a la comprensión del proceso
de cierre del tubo neural en el embrión de pollo e intentar ahondar en las
funciones de la proteína MARCKS durante esta etapa.
Se intenta estudiar los posibles efectos del LiCl sobre la neurulación en
embriones de pollo. A su vez, veremos si es posible correlacionar la localización
subcelular de MARCKS con defectos en el cierre del tubo neural en embriones
tratados con este fármaco.

15
IV Resultados __________________________________________________
i Neurulación en condiciones de cultivo
En el comienzo de este trabajo pusimos a punto la técnica de cultivo de
embriones de pollo que utilizaríamos posteriormente en el laboratorio para
abordar los objetivos propuestos. Seleccionamos dos métodos de cultivo de la
bibliografía que podrían ser útiles: el cultivo de New y el cultivo EC. El primero
consiste en el crecimiento del embrión in vitro sobre la membrana vitelina
tensada en un anillo de vidrio y colocado sobre un sustrato de albúmina líquida
del propio huevo como medio nutritivo (New, 1955). El segundo utiliza un papel
de filtro para mantener el embrión bajo la tensión de la membrana vitelina, el cual
crece sobre un sustrato semisólido de agar-albúmina (Chapman et al., 2001).
Luego de realizar meticulosas pruebas con ambos métodos, decidimos utilizar el
método de cultivo EC (ver Materiales y Métodos para descripción de la técnica).
Este método permitió que los embriones de pollo continuasen su desarrollo
normal, hasta HH12 aproximadamente, una vez que fueron extraídos del huevo
(a partir de 12 horas de incubación). Observamos que la tasa de desarrollo de
los embriones en cultivo fue un poco más lenta que los embriones incubados in-
ovo. Las horas de incubación propuestas por Hamburger y Hamilton (1951) para
los estadios tempranos del desarrollo muchas veces no coincidieron con los
estadios alcanzados por los embriones en cultivo. También observamos
diferencias en la supervivencia de los embriones según el estadio de inicio del
cultivo. La supervivencia fue siempre mayor cuanto más avanzado el estadio.
Seguramente en los embriones más pequeños, al ser más frágiles y delicados,
aumenta la probabilidad de daño durante la manipulación que lleva al cultivo,
provocando mayor letalidad. A su vez, los cultivos realizados a partir de estadios
más avanzados resultaron técnicamente mucho más sencillos, probablemente
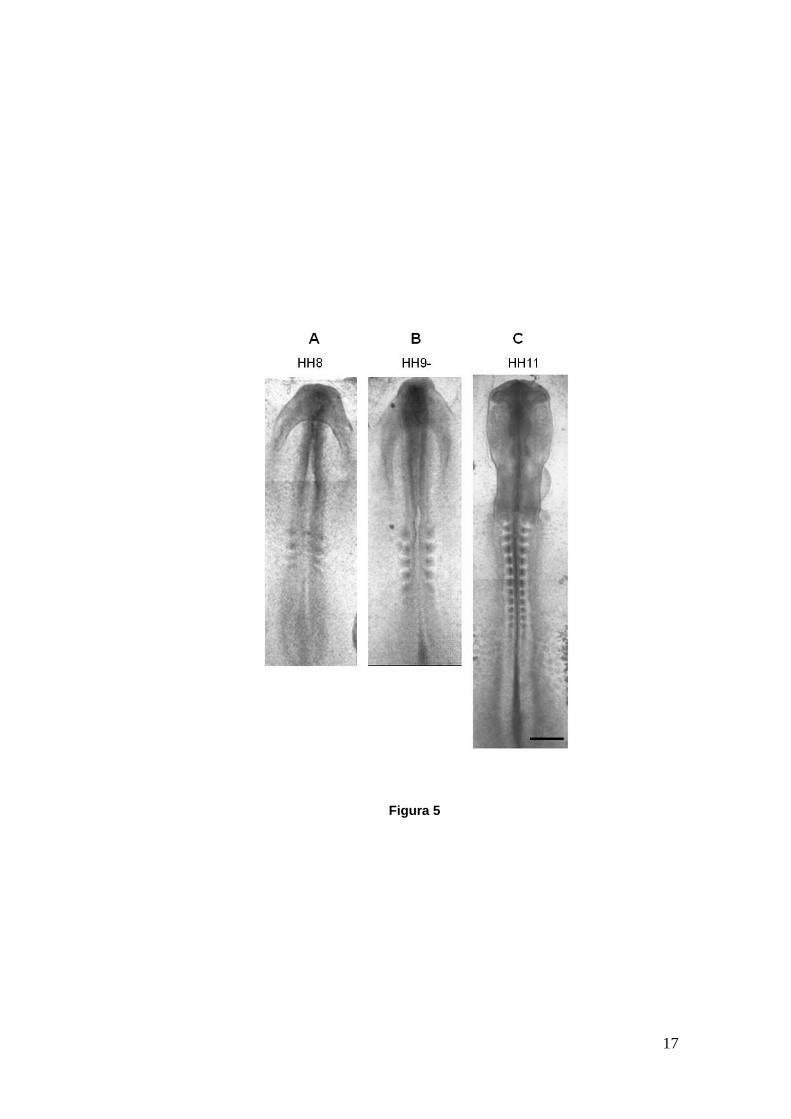
por el motivo mencionado anteriormente. En la Figura 5 mostramos una imagen
de embriones de pollo en cultivo utilizando el método EC. Como podemos
observar se trata de tres estadios distintos del desarrollo; los dos primeros
corresponden al proceso de neurulación (HH8 y HH9-) mientras que en el último
ya se ha formado completamente el tubo neural a lo largo de todo el eje céfalo-

16
caudal (HH11). En estos embriones podemos distinguir la región cefálica (arriba)
y la región caudal (abajo) así como también los somites que aumentan en
número a medida que el embrión avanza de estadio. En la Figura 5C el embrión
posee vesículas ópticas y cerebrales en formación y su corazón ya late aunque
recién comienza a tomar forma. Parte de la puesta a punto del método de cultivo
fue importante para observar y determinar los estadios implicados en el proceso
de neurulación en el pollo útiles para nuestra investigación. Utilizamos como
ayuda la tabla de estadios normales del desarrollo en el pollo de Hamburguer y
Hamilton (1951) para realizar las dataciones. El contacto entre los pliegues
neurales que lleva a la formación del tubo neural comienza en HH8, como
mostramos en la Figura 5A. Sin embargo, la formación de la placa neural junto
con el posterior modelado y flexión comienzan en etapas anteriores. Para HH9-
(Figura 5B) el cierre ha avanzado a lo largo de toda la región cefálica y parte del
tronco y para estadios posteriores (HH10 en adelante) ya se observa un cierre
casi total o total del tubo neural. Decidimos entonces realizar los tratamientos
farmacológicos con LiCl desde el inicio de la neurulación (HH6). También se
entendió pertinente realizar cultivos con el fármaco desde etapas anteriores
(HH3) para evaluar el efecto del tiempo en los tratamientos y una vez establecido
el contacto de los pliegues neurales (HH8). En todos los casos la incubación con
LiCl se extendió hasta aproximadamente HH9 o estadios más avanzados.
17
Figura 5

18
Figura 5.- Embriones de pollo in toto en cultivo EC. A. Embrión en HH8 con 4 pares de somites. El cierre del tubo neural está comenzado en la región cefálica mientras que en el resto del eje céfalo-caudal todavía se observa el surco neural. B. Embrión en HH9- con 6 pares de somites. El cierre del tubo neural ha avanzado hasta la región del tronco. Se observan los pliegues neurales muy próximos entre sí a la altura de los somites. C. Embrión en HH11 con 12 pares de somites. El cierre del tubo neural ya ha ocurrido a lo largo de todo el eje céfalo-caudal. Se observan las vesículas ópticas y cerebrales en formación y el codo de la arteria aorta hacia la derecha. Las imágenes fueron tomadas en microscopio compuesto (A) y lupa estereoscópica (B, C). Barra: 300 µm.
a - MARCKS en condiciones de cultivo
Con el objetivo de analizar la distribución subcelular de la proteína MARCKS
durante el proceso de neurulación en condiciones de cultivo, realizamos
inmunomarcaciones con el anticuerpo policlonal Polo52, dirigido contra la
proteína entera de pollo y marcadores para el neuroepitelio (N-CAM) y los
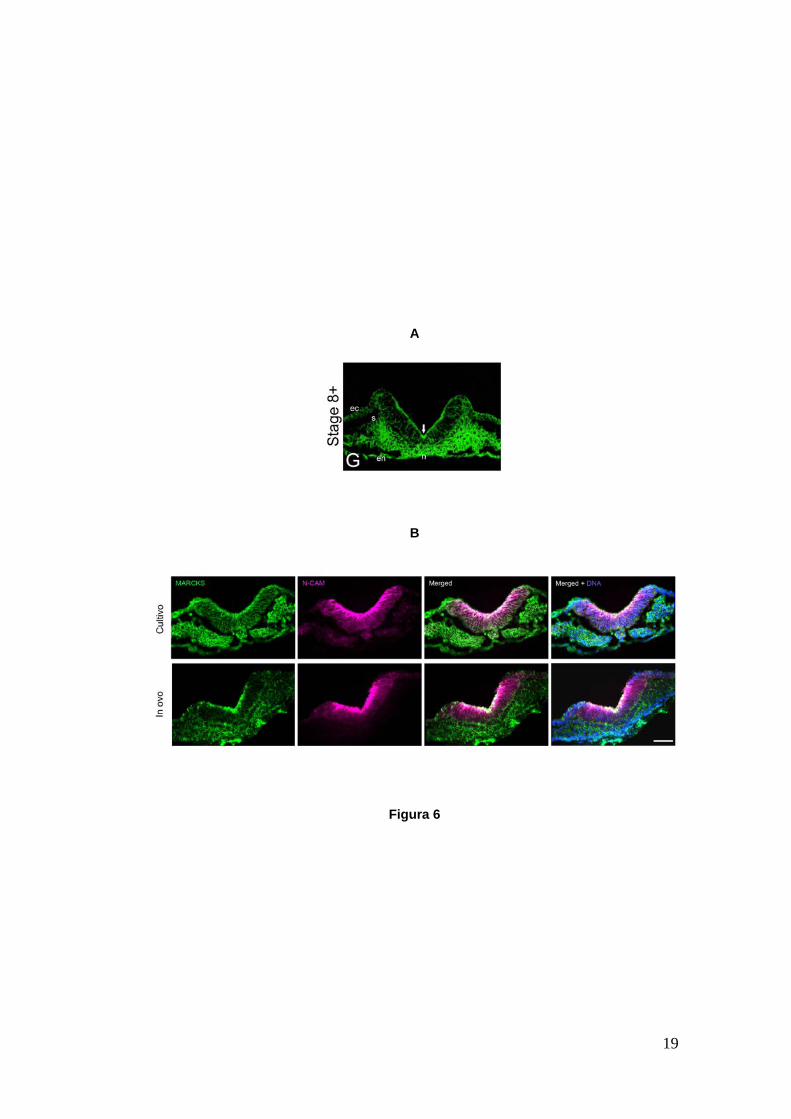
núcleos. En la Figura 6 se muestran los resultados obtenidos previamente en el
laboratorio (Zolessi y Arruti, 2001) y los logrados en nuestros ensayos. Como
podemos observar, la Figura 6A ilustra un corte transversal a nivel del tronco de
un embrión en HH8+. La neurulación se halla en proceso y se observa la flexión
de la placa neural, la cual ha adquirido forma de V. Los somites se evidencian a
ambos lados de la placa neural, entre el ectodermo y el endodermo. MARCKS se
observa apicalizada a lo largo del neuroepitelio durante esta etapa (Zolessi y
Arruti, 2001). En la Figura 6B se muestran cortes transversales de embriones en
HH8+ a nivel del tronco en condiciones de cultivo y extraídos directamente del
huevo. MARCKS se mantiene apicalizada entre los embriones cultivados en
comparación con los obtenidos directamente del huevo. En ambos casos se
observó una distribución apical de MARCKS durante HH8+ (neurulación) como
fuera descrito por Zolessi y Arruti (2001). Cabe mencionar que MARCKS
presenta una distribución homogénea en forma previa y posterior al cierre del
tubo neural. Los filamentos de actina se mantienen también en posición apical
durante y también luego del proceso de neurulación y muestran co-localización
con MARCKS en esa zona del neuroepitelio (Zolessi y Arruti, 2001).
19
A
B
Figura 6

20
Figura 6.- Cortes transversales de embriones de pollo en HH8+ a nivel del tronco. A. Se muestra la apicalización de la proteína MARCKS en el neuroepitelio que ya ha comenzado el proceso de modelado (Zolessi y Arruti, 2001). B. Embriones en condiciones de cultivo y extraído del huevo (in-ovo). Se observa la misma apicalización de MARCKS que en A. Se muestran la proteína MARCKS (verde), el marcador temprano de placa neural N-CAM (fucsia) y ambas imágenes superpuestas. En las últimas se agregan los núcleos celulares (azul). ec, ectodermo; en, endodermo; n, notocorda; s, somite. Las micrografías fueron tomadas en microscopio de fluorescencia. Barra: 50 µm.
ii Tratamiento farmacológico de embriones en condiciones de cultivo
durante la neurulación
Para abordar el objetivo planteado en este trabajo realizamos tratamientos con
LiCl en embriones en cultivo durante el período de neurulación. Como
mencionamos anteriormente, los cultivos fueron realizados principalmente a
partir de HH6, cuando el proceso de formación del tubo neural comienza, hasta
HH9, momento en el que el cierre ya se ha producido en la región cefálica y del
tronco. En algunos casos, los cultivos fueron iniciados desde estadios más
tempranos (gastrulación) y más tardíos (neurulación avanzada) para evaluar: i) la
existencia de un período crítico de acción del teratógeno y, ii) dependencia del
tiempo de exposición. Según Thomas y Collins (1987) se necesitan 3 dosis para
determinar la dosis teratogénica óptima de un fármaco: una dosis alta que sea
tóxica pero no letal, una dosis baja que se encuentre cercana al rango clínico y
que no cause efectos evidentes y una dosis intermedia. Utilizamos entonces 3
dosis distintas de la droga: 20, 60 y 100 mM, que de acuerdo a la bibliografía
causaban efectos desde escasos o nulos a severos, respectivamente (Giles y
Bannigan, 1999). El modo de aplicación de la droga fue por disolución en el
medio de cultivo (agar-albúmina). En este caso, la parte dorsal del embrión
permanece directamente en contacto con el sustrato y el LiCl actúa por difusión.
También se ensayó la aplicación directa de mínimos volúmenes de la droga
sobre el embrión. En este procedimiento el fármaco queda en contacto directo
con la parte ventral del embrión. A pesar de estas diferencias, ambos métodos
arrojaron los mismos resultados.

21
a - Tratamientos con 100 mM de cloruro de litio
Los cultivos de embriones tempranos de pollo a partir de HH3 tratados con una
concentración de 100 mM de LiCl hasta HH9, mostraron efectos muy severos del
fármaco. En este caso se aplicó la dosis más alta y el mayor tiempo de
exposición, por lo cual era de esperar dicho resultado. En HH3 el embrión inicia
la gastrulación, proceso que involucra movimientos celulares que darán origen a
las 3 capas embrionarias. Muchos de los embriones cultivados bajo estas
condiciones no sobrevivieron al tratamiento. Los embriones supervivientes
resultaron altamente afectados exhibiendo morfologías muy diferentes a los
individuos control, con mínimo desarrollo y ausencia de estructuras normales en
relación al tiempo de incubación. La Figura 7 muestra un corte transversal de un
embrión control y uno tratado bajo estas condiciones. En la primera se pueden
distinguir fácilmente las distintas estructuras del embrión. El tubo neural, formado
por el neuroepitelio plegado y la luz, se observa en la zona superior y se
encuentra casi cerrado. La apicalización de MARCKS ya ha desaparecido,
producto del cierre casi total del tubo neural. Se pueden distinguir el ectodermo
suprayacente, el mesodermo en la parte central y el endodermo por debajo. En
la imagen inferior se observan claramente los efectos del LiCl, el cual causó
importantes modificaciones en la morfología del embrión. No es posible distinguir
estructuras, ni capas embrionarias con certeza. El embrión presenta visibles
defectos en la neurulación con respecto al control. No cuenta con un
neuroepitelio definido, ni un tubo neural en proceso de formación (de hecho no
se logra visualizar ninguna estructura que se le asemeje). Tampoco es posible
diferenciar las regiones ventral y dorsal del embrión como consecuencia de los
drásticos cambios ocurridos. Además, no se observa apicalización de MARCKS
en ningún sector de la imagen, que podría constituir una guía para definir la
región dorsal. Los núcleos que se encuentran en la parte superior formando un
cordón (de izquierda a derecha de la imagen) podrían ser parte del neuroepitelio
que no llegó a plegarse. Una imagen del mismo corte del embrión con
inmunomarcación para actina filamentosa muestra una pequeña acumulación en
la parte superior, lo que sugiere que podría corresponder al lado dorsal, aunque
no podemos dar certeza de esto.

22
Figura 7

23
Figura 7.- Cortes transversales de embriones de pollo a nivel cefálico cultivados desde HH3 hasta HH9. La imagen superior corresponde a un embrión control. Ilustra el tubo neural en formación, con un neuroepitelio plegado bien definido. La imagen inferior es de un embrión tratado con una dosis de 100 mM de LiCl. Evidencia un drástico efecto del fármaco con pérdida de ejes corporales y de la morfología normal. Se muestra la distribución de la proteína MARCKS (verde) y los núcleos celulares (azul). ec, ectodermo; ne, neuroepitelio; n, notocorda. Las imágenes fueron tomadas en microscopio de fluorescencia. Barra: 50 µm.
Las dificultades en la identificación de los ejes corporales nos llevaron a utilizar
un marcador de placa neural en las inmunodetecciones para ubicar el
neuroepitelio. De esta manera es posible reconocer los lados dorsal y ventral del
embrión cuando los efectos de la droga modifican la morfología del embrión.
Esta estrategia es especialmente útil en los casos de afectación más leves
aunque de limitada aplicación en los tratamientos más severos (como el que
analizamos anteriormente). El anticuerpo que escogimos se denomina 5e y es
un marcador de las uniones intercelulares del neuroepitelio. Muestra la presencia
de la glicoproteína de unión celular N-CAM y se utiliza como marcador temprano
de placa neural (Sanes et al., 2006). Si bien se localiza preferentemente en el
neuroepitelio, observamos que también marca la notocorda y con menor
intensidad el mesodermo lateral. Antes de utilizarlo en nuestras
inmunomarcaciones realizamos ensayos con distintas diluciones del anticuerpo
para determinar la concentración más adecuada. Decidimos emplear una
dilución 1/400.
Cuando el tratamiento con 100 mM de LiCl se realizó en embriones de estadios
más avanzados (HH5 y 6 y hasta HH11) sometidos a la misma exposición, el
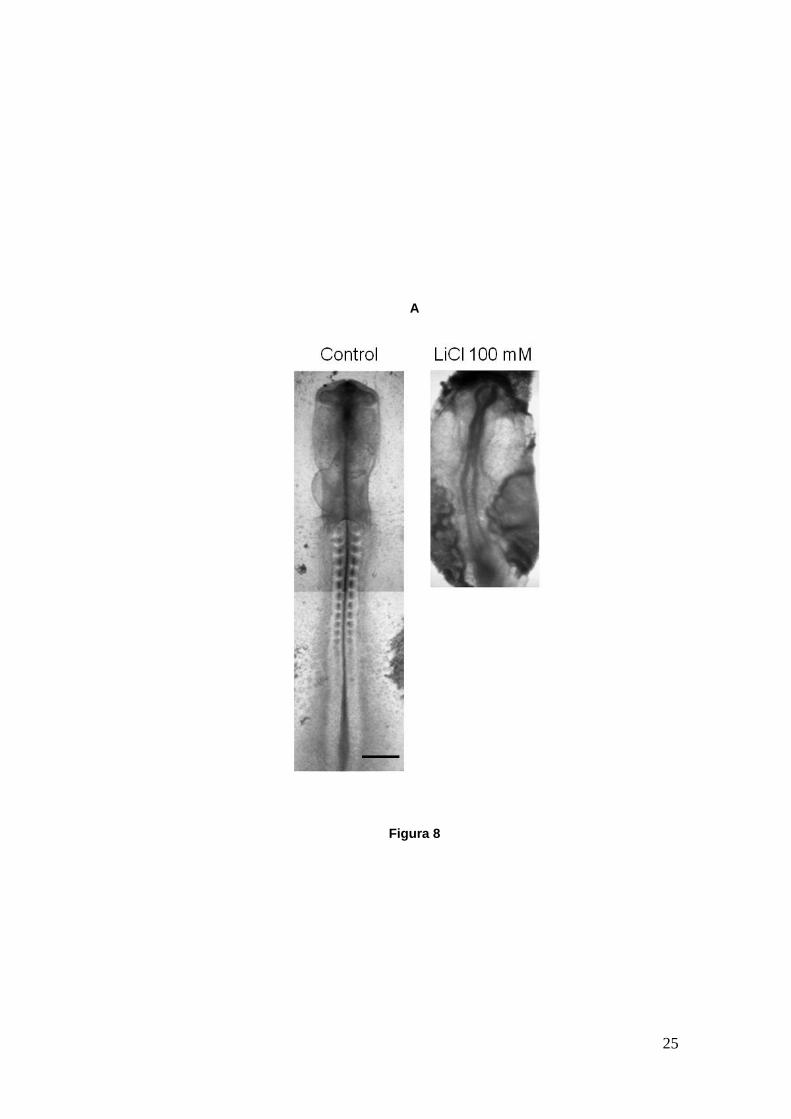
efecto de la droga fue menos severo. En la Figura 8A se muestra una imagen de
embriones in toto control y tratado. Como podemos observar, el embrión control
se ha desarrollado normalmente y ha llegado a HH11. Presenta una región
cefálica en formación, con las divisiones características del cerebro en esta
etapa (prosencéfalo, mesencéfalo y rombencéfalo), vesículas ópticas en
desarrollo, corazón en funcionamiento (podemos visualizar la aorta hacia el lado
derecho), 13 pares de somites y un tubo neural completamente cerrado desde el
extremo cefálico al caudal. Si se comparan el embrión control y el tratado se
evidencian claras diferencias. Este último presenta un tamaño mucho menor, la

24
distancia entre la región cefálica y caudal se ha reducido en forma notable y
muestra un importante retraso en el desarrollo. Si bien en este caso se
conservan los ejes corporales, el embrión carece de las estructuras indicadas
para el control. La región cefálica no presenta vesículas cerebrales ni ópticas, no
se observa el corazón y tampoco somites. Es posible observar un tubo neural en
proceso de formación con pocas zonas de contacto de los pliegues neurales. En
la mayor parte del eje antero-posterior el tubo neural permanece abierto. No se
ha producido un cierre normal del tubo neural que en el pollo transcurre desde la
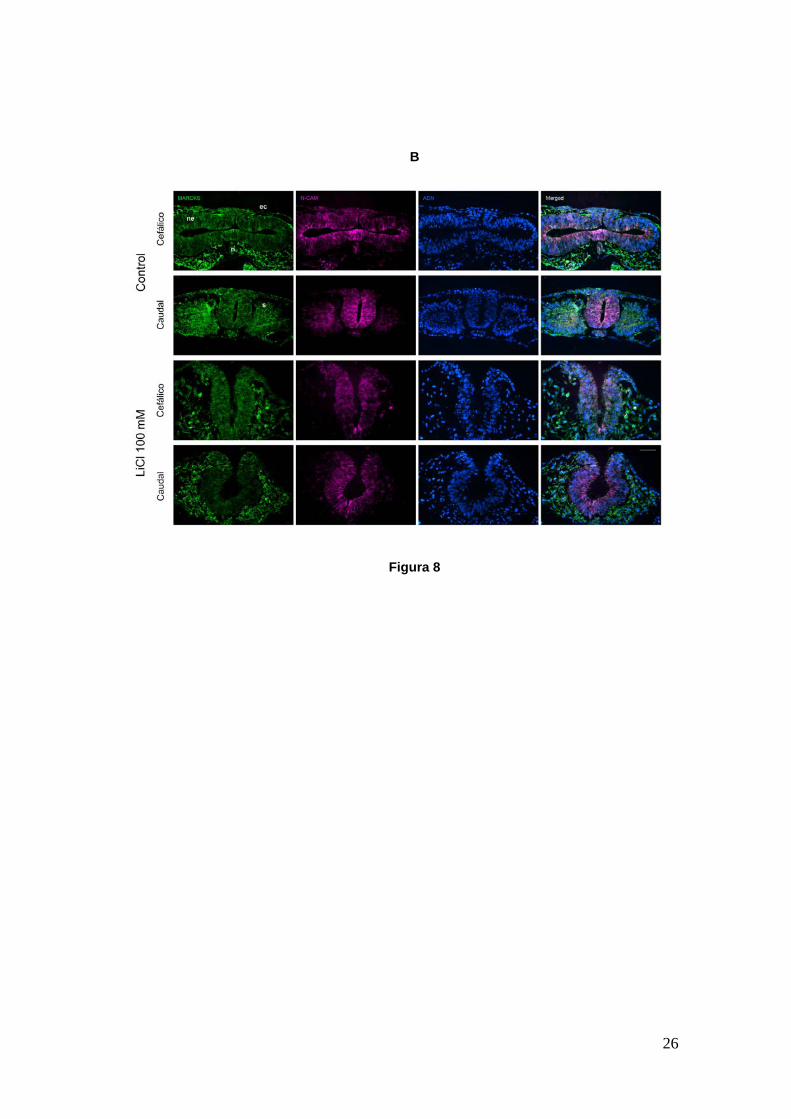
región cefálica hacia la caudal. En la Figura 8B, se muestran cortes
transversales de los embriones de la parte A en dos posiciones distintas del eje
antero-posterior, una cefálica y otra más caudal, donde se utilizaron distintos
marcadores. Se observan las diferencias que mencionamos entre ambos
embriones. En el embrión tratado no se ha producido el cierre del tubo neural,
aunque es evidente que los movimientos de flexión de la placa neural han tenido
lugar considerando la cercanía de los pliegues neurales. En el estadio alcanzado
por el embrión control (HH11), MARCKS ha modificado su localización apical en
el neuroepitelio y muestra nuevamente una distribución homogénea.
Interesantemente, en el embrión tratado ocurre lo mismo pese a que el cierre del
tubo neural no se ha producido.
25
A
Figura 8
26
B
Figura 8

27
Figura 8.- Embriones de pollo en cultivo, control y tratado con una dosis de 100 mM de LiCl desde HH6 hasta HH11. A. Embriones in toto. El control ha alcanzado un desarrollo normal para el tiempo de incubación mientras que el tratado muestra defectos importantes: retraso en el desarrollo, largo reducido del eje céfalo-caudal, no presenta vesículas cerebrales ni ópticas, corazón ni tampoco somites; presenta un tubo neural en formación pero que permanece mayoritariamente abierto. B. Cortes transversales a la altura de la región cefálica y más caudales. Se muestra la proteína MARCKS (verde), el marcador temprano de placa neural N-CAM (fucsia) y los núcleos celulares (azul). El embrión control posee un tubo neural cerrado en las dos regiones ilustradas mientras que en el tratado se encuentra aún abierto. ec, ectodermo; en, endodermo; n, notocorda, s, somite. Las imágenes fueron tomadas en lupa estereoscópica (A) y microscopio de fluorescencia (B). Barra A: 300 µm; Barra B: 50 µm.
Analizando más detalladamente cortes finos transversales marcados para ADN
de los embriones de la Figura 8, corroboramos que el tubo neural se encuentra
prácticamente cerrado en el embrión control, mientras que en el tratado
permanece abierto con los pliegues neurales separados. Logramos determinar
que el embrión tratado presenta un importante incremento en la muerte celular a
lo largo del neuroepitelio, hecho que se manifiesta por el aumento de los núcleos
picnóticos en comparación con el control (Figura 9).
Finalmente realizamos tratamientos con la misma dosis de LiCl utilizada
anteriormente, 100 mM, pero esta vez a partir de un estadio más tardío y un
período de tiempo menor. Los embriones fueron cultivados a partir de HH8 y
hasta aproximadamente HH9. Los resultados obtenidos muestran algunas
diferencias entre embriones tratados y control, que podemos visualizar en la
Figura 10. El proceso de cierre del tubo neural se encuentra retrasado en el
embrión tratado; el contacto de los pliegues neurales se ha producido
únicamente en la región anterior, mientras que en todo el tronco y región
posterior los pliegues permanecen distantes. Los somites muestran una mayor
separación en comparación con el respectivo control.

28
Figura 9
29
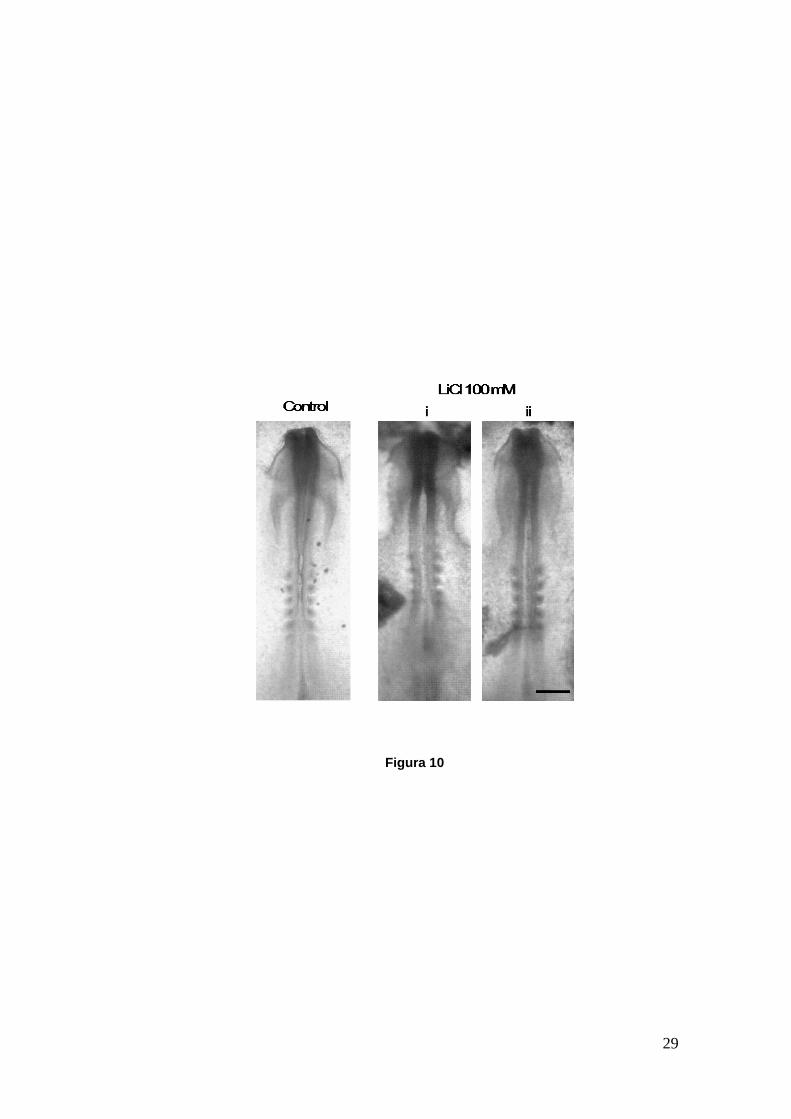
Figura 10

30
Figura 9.- Cortes transversales de embriones de pollo a nivel cefálico cultivados desde HH6 hasta HH11. La imagen superior corresponde a un embrión control. Muestra el neuroepitelio plegado. Los pliegues neurales han hecho contacto y la luz del tubo neural se observa expandida hacia los lados, ya que pertenece a la región cefálica. La imagen inferior es de un embrión tratado con una dosis de 100 mM de LiCl. Se observa el neuroepitelio en forma de U. El tubo neural no se ha cerrado, los pliegues neurales permanecen distantes. Se observa un aumento en el número de núcleos picnóticos correspondientes a muerte celular. Se muestran los núcleos celulares (azul). ec, ectodermo; ne, neuroepitelio; n, notocorda. Las imágenes fueron tomadas en microscopio de fluorescencia. Barra: 50 µm.
Figura 10.- Embriones de pollo in toto en cultivo; control y tratados con una dosis de 100 mM de LiCl desde HH8 hasta HH8+. En el embrión control el cierre del tubo neural ha avanzado hasta el inicio de los somites, donde comienza el contacto entre los pliegues neurales. Los embriones tratados (i, ii) muestran un retraso evidente en el cierre del tubo neural y una mayor separación entre los pliegues neurales en gran parte del eje céfalo-caudal. Las imágenes fueron tomadas en lupa estereoscópica. Barra: 300 µm.
b - Tratamientos con 60 mM de cloruro de litio
Los embriones tratados con una dosis de 60 mM de LiCl (HH6 hasta
aproximadamente HH9) mostraron algunas diferencias en su desarrollo con
respecto a los controles. En la Figura 11A podemos observar un embrión control
en HH9- con 6 pares de somites en el cual el cierre del tubo neural abarca la
mayor parte del eje céfalo-caudal. Encontramos dos tipos de morfologías
características en los embriones expuestos a tratamientos con 60 mM de LiCl. El
embrión i muestra una longitud céfalo-caudal similar al control. Si bien el cierre
del tubo neural es similar al control en el sector anterior, se observa una
separación mayor de los pliegues neurales a nivel de los somites. El número de
somites se corresponde con el estadio alcanzado pero los mismos muestran un
aspecto menos definido que en el control. El embrión ii muestra una mayor
longitud del eje antero-posterior. Los pliegues neurales no han hecho contacto
aún a lo largo de este eje. Podemos observar un número menor de somites (3
pares) que no concuerda con el estadio del desarrollo en relación al tiempo de
incubación. Este embrión muestra una morfología poco común en la parte
posterior o caudal, donde se ha producido un plegamiento incorrecto de la placa
neural que se muestra expandida como lo evidencia el marcador N-CAM. La

31
placa contiene un surco mayor en el medio y dos surcos ectópicos más
pequeños a los costados que también son positivos para N-CAM (Figura 11B).
También realizamos tratamientos con una dosis de 60 mM de LiCl por un lapso
menor (HH8 hasta aproximadamente HH9). En la Figura 12 mostramos los
resultados macroscópicos obtenidos. Al momento de la fijación, el embrión
control y el tratado i habían alcanzado un HH8+ mientras que el tratado ii se
hallaba apenas más avanzado en su desarrollo (HH9). Si bien el cierre del tubo
neural avanza desde cefálico a caudal en el embrión tratado, el proceso parece
algo retrasado. En el embrión tratado ii el cierre ha llegado hasta la altura del
tronco, como en el control, aunque fue necesario un mayor tiempo de incubación
para lograrlo. Otra diferencia comprende al neuroporo anterior, que se encuentra
más abierto en los embriones que fueron expuestos al fármaco.
32
A
B
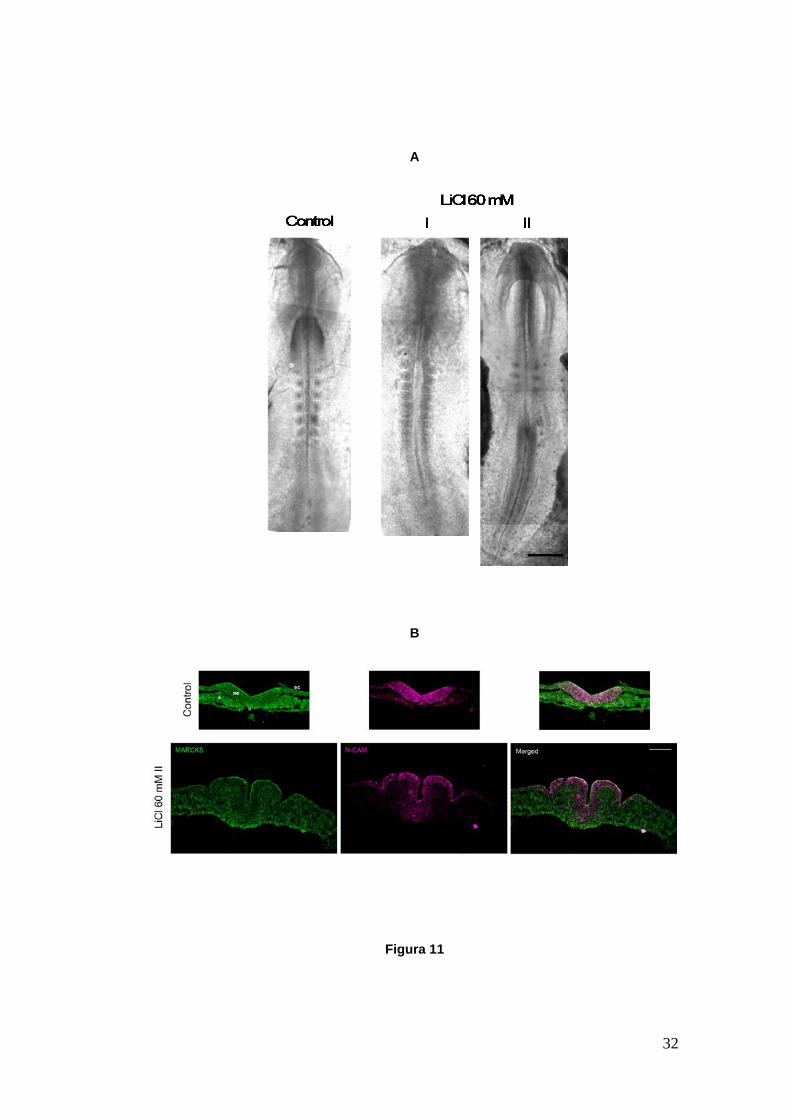
Figura 11

33
Figura 11.- Embriones de pollo en cultivo, control y tratados con una dosis de 60 mM de LiCl (desde HH6 hasta aproximadamente HH9). A. Embriones in toto control y tratados. Para el caso de los embriones tratados se muestran las dos morfologías obtenidas: i. El largo del eje céfalo-caudal es similar al control pero el cierre del tubo neural se encuentra retrasado en el tronco y los somites aparecen menos definidos; ii. El largo del eje céfalo-caudal se ha modificado, mostrando un aumento debido a la prolongación de los pliegues neurales posteriores y únicamente 3 pares de somites. B. Cortes transversales a nivel caudal de los embriones control y tratado II. Este último posee una placa neural expandida con dos surcos neurales ectópicos a los lados del surco principal. Se muestra la proteína MARCKS (verde), el marcador temprano de placa neural N-CAM (fucsia) y la imagen superpuesta. ec, ectodermo; ne, neuroepitelio; n, notocorda; s, somite. Las imágenes fueron tomadas en microscopio compuesto (A) y en microscopio de fluorescencia (B). Barra A: 300 µm. Barra B: 50 µm.
c - Tratamientos con 20 mM de cloruro de litio
Los tratamientos más leves fueron realizados con una concentración de 20 mM
de LiCl, dosis propuesta como aquella cercana al rango clínico y que no causa
efectos evidentes (Thomas y Collins, 1987; Giles y Bannigan, 1999). Los
embriones tratados con esta dosis fueron cultivados desde HH3 o HH6 hasta
HH9 y HH11. En este caso no fueron observados efectos a nivel macro- o
microscópico en comparación con el control. En la Figura 12A se ilustran 2
embriones obtenidos por cultivo desde HH6 hasta HH11, uno tratado con 20 mM
de LiCl y otro control. El grado de desarrollo alcanzado por el embrión tratado es
normal para el tiempo de incubación, así como también el número de somites y
la distancia céfalo-caudal. El cierre del tubo neural se ha completado en casi
toda la longitud del embrión, salvo en la región caudal como también ocurre en el
control. Las inmunomarcaciones sobre criocortes transversales caudales de
estos embriones (Figura 13B) confirman los resultados observados in toto. En
ninguno de los embriones expuestos a esta dosis detectamos efectos del LiCl
sobre las estructuras en formación, así como tampoco sobre el cierre del tubo
neural.
34
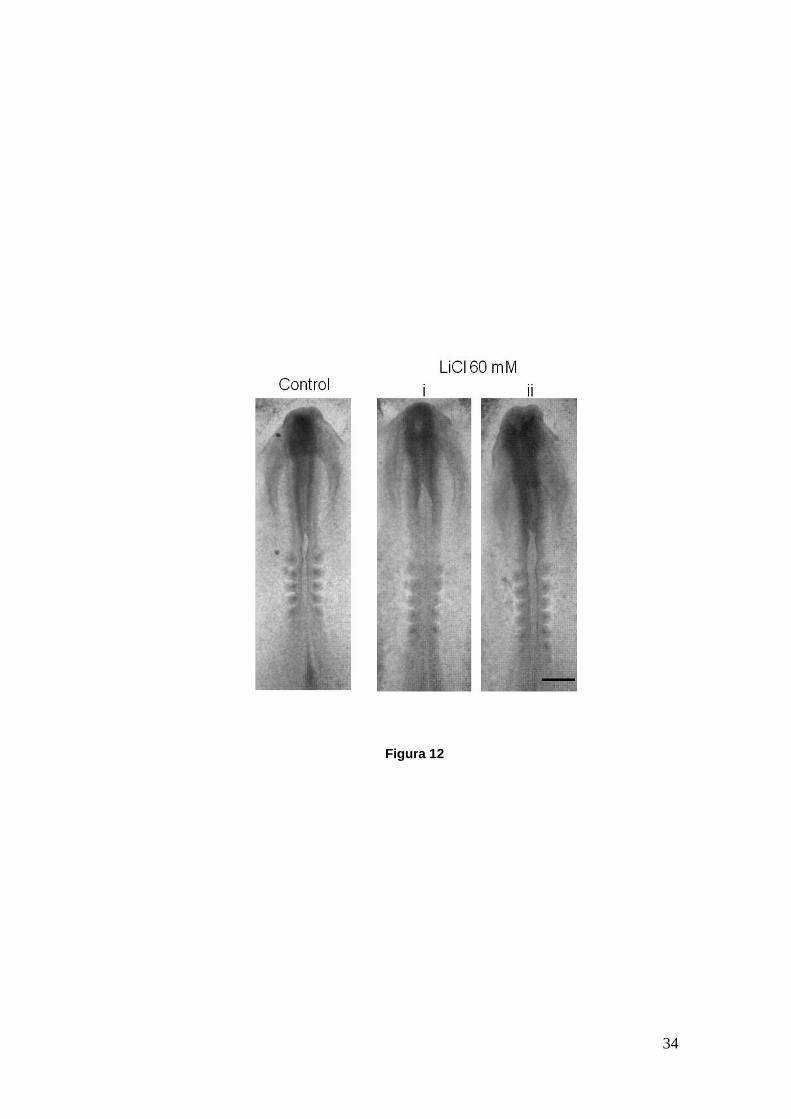
Figura 12

35
Figura 12. Embriones de pollo in toto en cultivo, control y tratados con una dosis de 60 mM de LiCl desde HH8 hasta aproximadamente HH9. En el embrión control (HH8+) se ha alcanzado el cierre del tubo neural hasta el inicio de los somites donde comienzan a establecer contacto los pliegues neurales. Para los embriones tratados se muestran un HH8+ (i) y HH9 (ii). Se observa un retraso en el cierre del tubo neural y una apertura mayor del neuroporo anterior en comparación con el embrión control. Las imágenes fueron tomadas en lupa estereoscópica. Barra: 300 µm.
36
A
B
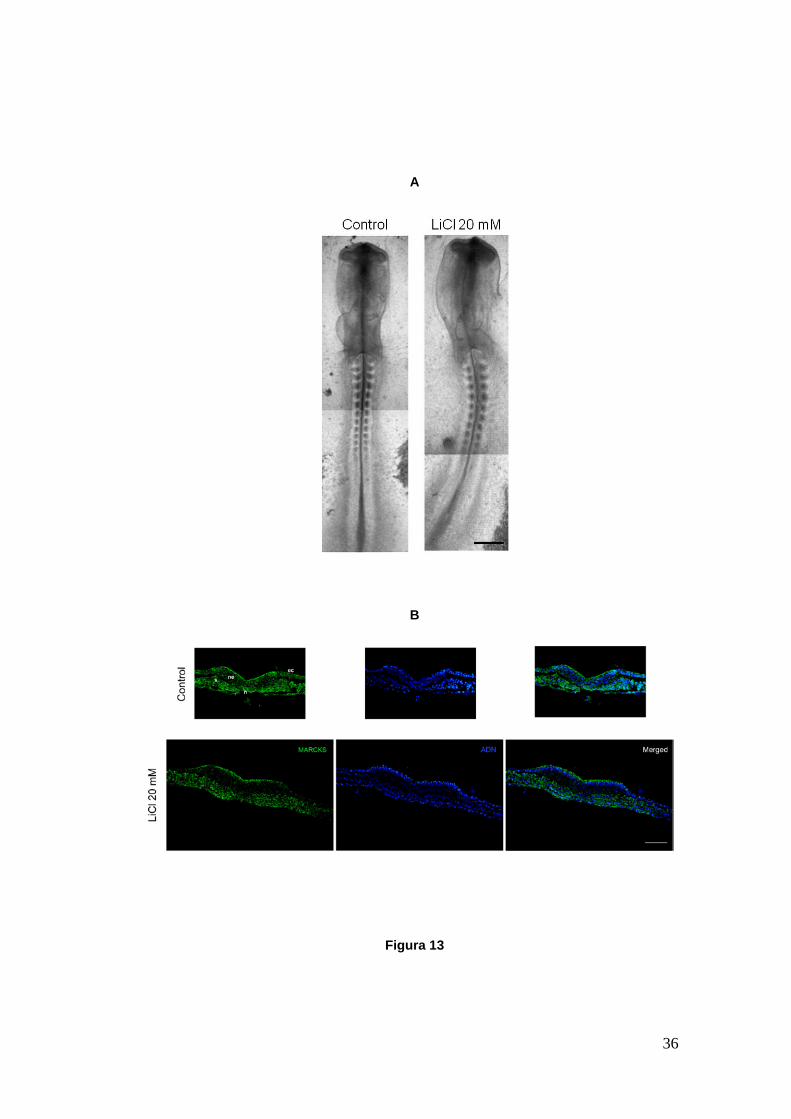
Figura 13

37
Figura 13.- Embriones de pollo en cultivo, control y tratado con una dosis de 20 mM de LiCl. A. Embriones in toto cultivados desde HH6 hasta HH11. El embrión tratado no exhibe diferencias con respecto al control. Ambos muestran una morfología normal para el estadio del desarrollo alcanzado. El cierre del tubo neural está llegando a su fin. Los embriones presentan vesículas ópticas en formación y 13 pares de somites. B. Cortes transversales de embriones cultivados desde HH6 a HH9 a la altura de la región caudal. Se muestra la proteína MARCKS (verde), los núcleos celulares (azul) y la superposición de ambas imágenes. Tanto el embrión control como el tratado muestran una placa neural en etapa de modelado, donde MARCKS comienza a observarse en la región apical del neuroepitelio. ec, ectodermo; ne, neuroepitelio; n, notocorda; s, somite. Las imágenes fueron tomadas en lupa estereoscópica (A) y microscopio de fluorescencia (B). Barra A: 300 µm; Barra B: 50 µm.
iii Efectos del cloruro de litio sobre la localización subcelular de MARCKS
Luego de analizar los efectos generales que causó el tratamiento con LiCl sobre
embriones de pollo en cultivo, nos enfocamos en estudiar su acción sobre la
proteína MARCKS. Como mencionamos anteriormente, era de nuestro particular
interés observar si la proteína sufría algún cambio en su abundancia o
localización subcelular durante el proceso de cierre del tubo neural en
comparación con los respectivos controles. En la Figura 14 mostramos algunas
imágenes obtenidas al inmunomarcar MARCKS en embriones en cultivo control
y tratados con dos de la dosis utilizadas en nuestro trabajo: 20 y 60 mM. De
acuerdo a lo reportado por Zolessi y Arruti (2001) esta proteína posee un
particular comportamiento a medida que transcurre la neurulación en el pollo.
Durante el cierre del tubo neural adopta una posición apical en las células que
componen el neuroepitelio y una vez finalizado el proceso vuelve a adquirir una
distribución homogénea. Cuando comparamos la distribución de MARCKS
durante la neurulación en las distintas situaciones analizadas pudimos observar
que, al igual que mostramos en la Figura 6, el embrión control presenta una
apicalización de la proteína a nivel del neuroepitelio. Se evidencia una marcación
más intensa (verde) en la zona apical de las células que lo componen. Al
comparar la imagen control con las correspondientes a las distintas dosis de LiCl
se comprobó que ocurre el mismo fenómeno respecto a la localización
subcelular de MARCKS. La proteína se mantiene apicalizada durante el proceso
y no muestra una disminución en el sector apical ni aumento en el citoplasma
aparentes, incluso cuando el embrión ha experimentado modificaciones en su

38
morfología producto de las acción del litio (embrión tratado con LiCl 60 mM). No
llegamos a obtener imágenes de embriones tratados con 100 mM y cultivados
desde HH6 a HH9, lo cual nos impide realizar una comparación con las
imágenes de la Figura 14. Los cortes obtenidos para esta dosis pertenecían a
embriones más avanzados en el desarrollo (Figura 6) donde MARCKS ya no se
observa apicalizada y regresa a su distribución original. Si bien obtuvimos
embriones que fueron cultivados desde HH3 a HH9 con 100 mM con
inmunomarcación para este proteína (Figura 7), la severidad del efecto del LiCl
impidió la comparación con los otros experimentos.
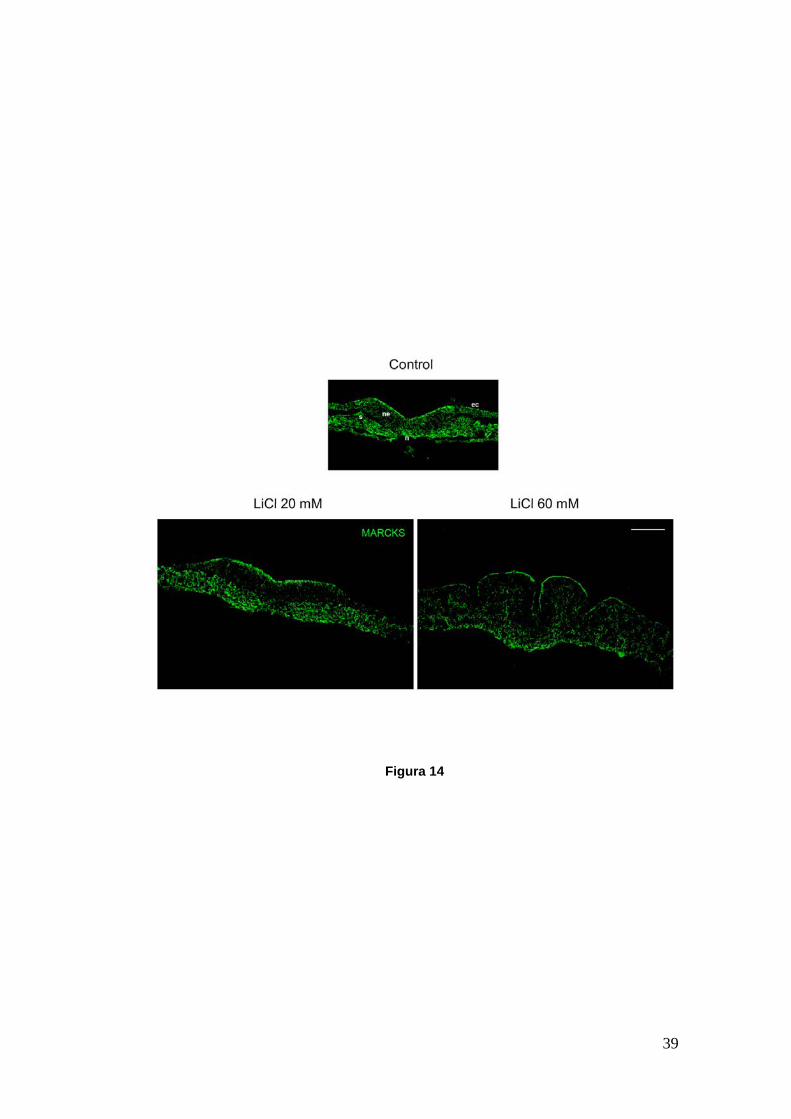
39
Figura 14

40
Figura 14.- Cortes transversales de embriones de pollo a nivel caudal cultivados desde HH6 hasta HH9. Se muestran embriones control y tratados con 20 y 60 mM de LiCl marcados para la proteína MARCKS (verde). En todos los casos MARCKS se mantiene apicalizada en el neuroepitelio durante el modelado y flexión de la placa neural. El LiCl no afecta la abundancia ni localización subcelular de MARCKS. ec, ectodermo; ne, neuroepitelio; n, notocorda; s, somite. Las micrografías corresponden a las mostradas previamente en las Figuras 11B y 13B. Barra: 50 µm.
Debido a que no encontramos cambios en la abundancia y localización
subcelular de MARCKS durante la neurulación en embriones tratados, decidimos
verificar la acción del LiCl sobre los embriones en cultivo. Para corroborarlo
utilizamos cortes transversales de embriones control y tratado con una dosis de
20 mM de la droga sobre los cuales realizamos inmunodetecciones para la
enzima GSK3β. Stambolic y colaboradores y Klein y Melton demostraron en
1996 la capacidad del litio de inhibir esta enzima a través de su fosforilación. En
la Figura 15A se exponen los resultados obtenidos al inmunomarcar la forma
fosforilada de la enzima (pGSK3β) en cortes transversales cefálicos. También se
adjunta en la misma el promedio de la cuantificación de intensidad de
fluorescencia (Figura 15B). Se puede observar un aumento en la intensidad de
fluorescencia de la enzima pGSK3β en el embrión tratado con respecto al
control. No sólo se incrementa la intensidad en cada célula, sino también el
número de células marcadas. Si observamos las medidas promedio de
intensidad de fluorescencia obtenidas vemos que el aumento llega a ser casi el
doble en los embriones tratados. Se logra una importante inactivación de la
enzima por la exposición a la dosis más pequeña (20 mM) para la cual no
encontramos efectos del LiCl en el desarrollo embrionario.
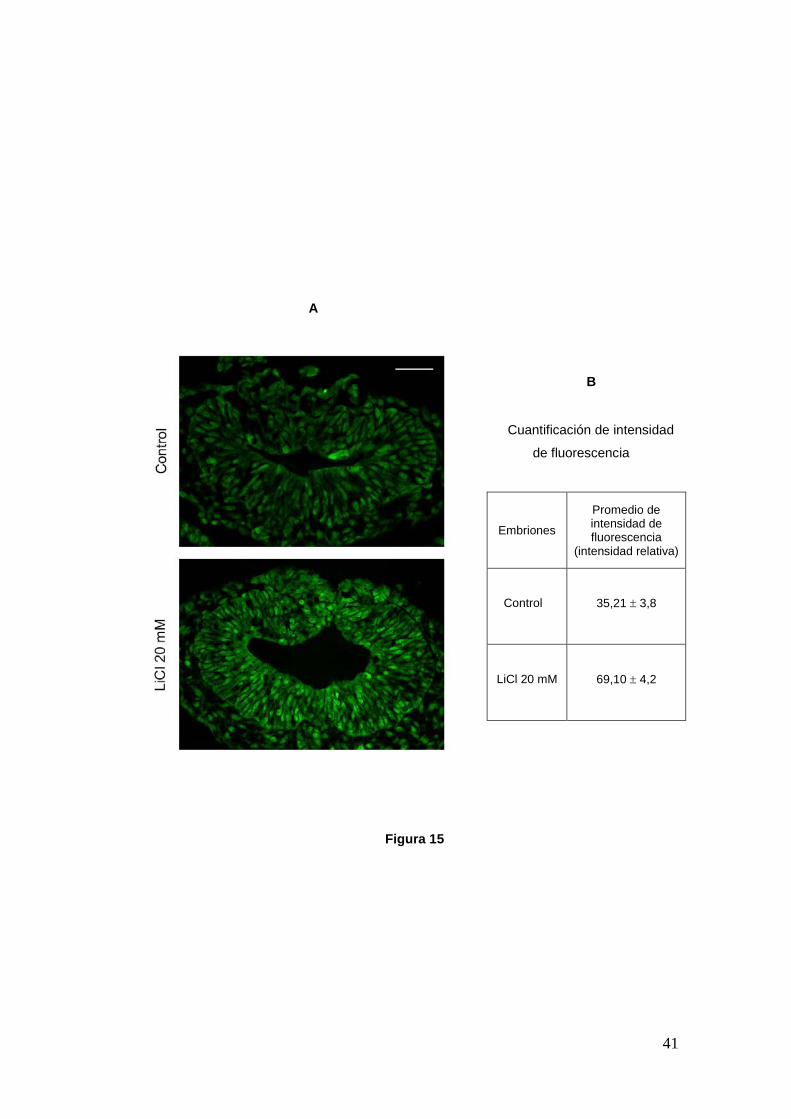
41
A
B
Cuantificación de intensidad
de fluorescencia
Figura 15
Embriones
Promedio de intensidad de fluorescencia
(intensidad relativa)
Control 35,21 ± 3,8
LiCl 20 mM 69,10 ± 4,2

42
Figura 15.- A. Cortes transversales de embriones de pollo a nivel cefálico cultivados desde HH6 a HH9. Se muestran embriones control y tratado con una dosis de 20 mM de LiCl marcados para la forma inactiva de la enzima GSK3β (pGSK3β; verde). Las imágenes fueron tomadas en microscopio de fluorescencia. Barra: 50 µm. B. Cuantificación de la intensidad relativa de fluorescencia de pGSK3β para imágenes cefálicas, a nivel del tronco y caudales de un total de 4 embriones: 2 controles y 2 tratados con 20 mM de LiCl. El litio aumenta la proporción de la enzima fosforilada en el embrión tratado, aún a concentraciones bajas donde no se aprecian efectos del fármaco.

43
V Discusión __________________________________________________
i Modelo de estudio y método de cultivo de embriones
Como primera conclusión podemos afirmar que el embrión de pollo resultó un
excelente modelo para nuestra investigación. Durante muchos años numerosos
investigadores han optado por utilizarlo en diversos estudios gracias a las
ventajas que ofrece y su relativa sencillez de manipulación (Giles y Banning,
1999; Stern, 2002; Dalgic et al., 2009). Si bien las aves están alejadas
filogenéticamente de los mamíferos, los procesos que suceden durante el
desarrollo temprano son extrapolables de forma tal que pueden guiarnos hacia
un mejor entendimiento de lo que ocurre en estas fascinantes etapas de la vida.
A su vez, el método de cultivo elegido funcionó correctamente y resultó ser
adecuado para lograr los objetivos propuestos. El método EC, propuesto por
Chapman y colaboradores en 2001, es una simple pero poderosa herramienta
para el estudio del desarrollo embrionario temprano. A pesar de ser un sistema
in vitro, que puede no reproducir perfectamente la realidad, cuenta con
importantes ventajas. Cuando realizamos el procesamiento de embriones en
cultivo y los comparamos con las imágenes correspondientes a embriones
extraídos directamente del huevo en los mismos estadios, pudimos verificar que
se correlacionan. Estos primeros resultados mostraron que los cultivos
reproducen el desarrollo normal del pollo en su etapa temprana. Dicho método
mostró además ser práctico, efectivo y útil para el estudio del proceso de
neurulación, permitiendo la observación y el seguimiento continuo de los
embriones bajo condiciones controladas de temperatura y humedad. Esto último
posibilita realizar su fijación en el momento deseado para su posterior
procesamiento. Una ventaja adicional es que permite la realización de varios
cultivos simultáneos con diferentes situaciones problema y con un número
significativo de individuos para cada caso. Esta metodología ha demostrado
también ser apropiada para desarrollar ensayos con el propósito de evaluar el
efecto de sustancias nocivas para el desarrollo o teratógenos. Hemos
comprobado que resulta un excelente modelo para estudiar NTDs. La técnica
que ideamos de disolución de la droga en el medio semisólido de agar-albúmina
permite la implementación de estudios dosis-efecto y elimina a la vez la

44
necesidad de agregar volúmenes adicionales al cultivo del embrión. En el
método EC se realiza el cultivo en ausencia de líquido sobre el lado ventral del
embrión (sólo necesita el contacto con el medio por su lado dorsal) y agregados
externos, aunque pequeños, podrían influir en el desarrollo normal. Los retrasos
en el desarrollo observados en embriones en cultivo con respecto a los
incubados in ovo probablemente se deban a la manipulación para iniciar los
cultivos. Los embriones son sacados fuera de las condiciones óptimas para su
crecimiento (37°C y humedad constante), extraídos del huevo, lavados y
colocados sobre un sistema muy distinto al de origen. Todos estos cambios
afectan el crecimiento del embrión hasta que es colocado nuevamente en la
incubadora, bajo condiciones óptimas, donde puede continuar su desarrollo.
Como se describe en la Introducción, el proceso de neurulación primaria en el
pollo es ampliamente conocido en la actualidad y se ha descrito en forma
detallada la secuencia de eventos que llevan a la formación del tubo neural
(Colas y Schoenwolf, 2001). Las observaciones realizadas en el presente estudio
permitieron identificar con precisión esta secuencia y determinar con exactitud
los estadios correspondientes a cada fenómeno. En el embrión de pollo, los
eventos preparatorios de la neurulación primaria comienzan en HH4 cuando las
células de la placa neural se hacen distinguibles en la región del ectodermo
dorsal, adquiriendo una forma alargada. El plegamiento comienza alrededor de
HH6 a medida que las células del punto bisagra neural medial se anclan a la
notocorda y cambian su forma, mientras que las células epidérmicas presuntivas
se mueven hacia la línea media dorsal. Los pliegues neurales son elevados a
medida que la epidermis presuntiva continúa moviéndose hacia esta línea. Se
produce la convergencia de los pliegues neurales cuando nos acercamos a HH8
a medida que las células del punto bisagra dorsolateral adoptan forma de cuña y
las células epidérmicas empujan hacia el centro. Finalmente desde HH8 en
adelante los pliegues neurales se contactan y las células de la cresta neural, que
en un principio unen el tubo neural con la epidermis, se dispersan (Gilbert, 2006).

45
ii Tratamientos farmacológicos de embriones en condiciones de cultivo
durante la neurulación
La neurulación es uno de los procesos más relevantes del desarrollo. Una serie
de etapas consecutivas deben sucederse con precisión para que el sistema
nervioso pueda formarse y llegar a comandar las innumerables funciones de
todo un organismo (Wallingford, 2005). Pero como todo proceso biológico, puede
involucrar fallas que interfieran con su curso normal. Cuando falla el proceso de
cierre del tubo neural aparecen defectos conocidos como NTDs (Inagaki et al.,
2000; Pulikkunnel y Thomas, 2005). Los NTDs pueden originarse por factores
genéticos o ambientales (Daly et al., 1997; Detrait et al., 2005; Dhaulakhandi et
al., 2010). La droga que utilizamos durante este estudio, el LiCl, se administra
comúnmente para el tratamiento de trastornos bipolares y maníaco-depresivos.
A pesar de su conocida eficiencia clínica (Price y Henninger, 1994), su uso debe
ser controlado con precaución ya que existe un pequeño margen entre la dosis
terapéutica y la tóxica. La intoxicación por litio genera una toxicidad sistémica y
se ha visto que las manifestaciones clínicas características están directamente
correlacionadas con su concentración sérica (Kansagra et al., 2011). Los
tratamientos llevados a cabo con LiCl en este trabajo sobre embriones de pollo
en cultivo mostraron algunos efectos interesantes sobre los procesos tempranos
del desarrollo. Trabajos pioneros en este tema ya demostraban que en
vertebrados los efectos de la administración de litio dependen de la especie que
se emplea como modelo experimental y la dosis y duración del tratamiento. En
ensayos a corto plazo se pudo determinar que los niveles de LiCl que resultan
teratogénicos en el ratón son 6 veces superiores a los niveles terapéuticos de
litio en suero utilizados en humanos (Smithberg y Dixit, 1982). Incluso se han
documentado diferencias en la sensibilidad a la droga entre rata y ratón (Hansen
et al., 1990). Esta variación que parece existir en las dosis teratogénicas entre
diferentes especies nos impide generalizar resultados entre los distintos modelos
utilizados. Las tres dosis utilizadas en los cultivos de embriones de pollo
(seleccionadas según Thomas y Collins, 1987), si bien fueron algo superiores a
las alcanzadas en el suero humano bajo tratamiento terapéutico, mostraron
fenotipos correspondientes a los mencionados por estos autores para determinar

46
la dosis teratogénica óptima de un fármaco. El tratamiento con 100 mM de LiCl
resultó una dosis alta y tóxica pero no letal en la mayoría de los casos; la
administración de 20 mM de LiCl una dosis baja cercana al rango clínico sin
efectos evidentes mientras que la dosis de 60 mM de LiCl mostró efectos
intermedios.
A partir de los resultados obtenidos pudimos concluir que el efecto del LiCl
aumenta a medida que administramos cantidades crecientes de la droga.
Concentraciones altas (100 mM) de LiCl causaron importantes defectos en los
procesos tempranos del desarrollo de los embriones en cultivo y en algunos
casos la muerte. La dosis intermedia (60 mM) provocó efectos sobre la
neurulación sin afectar demasiado la morfología normal de los embriones ni los
ejes corporales. Por último, la concentración más baja (20 mM) no causó efectos
evidentes sobre los embriones expuestos comparados con los controles. Este
estudio demuestra entonces que el litio tiene un efecto teratogénico dependiente
de la concentración (dosis dependiente) como reportaron Giles y Bannigan
(1999) para el desarrollo vascular temprano en embriones de pollo en cultivo.
Además, se comprobó que los efectos provocados por el LiCl aumentan con el
tiempo de exposición en las diferentes etapas del desarrollo analizadas y son
dependientes del momento de administración. La primera conclusión se deduce
a partir de los resultados obtenidos para los tratamientos que difieren en el
tiempo de exposición al fármaco: los tratamientos más prolongados causaron
efectos mayores que los breves. Estos últimos, realizados desde HH8 hasta HH9
fueron los más leves para las dosis de 60 y 100 mM. Según Nokhbatolfoghahai y
Parivar (2008) uno de los factores críticos es el estadio del desarrollo en el cual
los embriones son expuestos a agentes teratógenos. En nuestro caso, pudimos
corroborar esta afirmación cuando la droga se empleó a iguales concentraciones
(100 mM) y tiempo de exposición pero comenzando su administración en
distintos momentos (estadios) del desarrollo. Concluimos que los efectos del LiCl
difieren según el momento de administración. Para los tratamientos realizados
con 100 mM iniciados desde los estadios más tempranos (HH3) los efectos
fueron drásticos. Los embriones mostraron fenotipos muy difíciles de interpretar
cuando sobrevivían al tratamiento. Observamos una pérdida casi completa de
los ejes corporales, no solo a nivel macroscópico sino también en

47
inmunomarcaciones. La aplicación de litio desde etapas muy tempranas parece
tener efectos sobre los procesos del desarrollo anteriores a la neurulación que
ocurren en ese momento. Este hecho puede observarse claramente en la Figura
8, donde los cortes transversales muestran que no se visualizan las tres capas
embrionarias resultantes del proceso de gastrulación. Un trabajo reciente,
llevado a cabo por Martin y colaboradores (2011) describe las malformaciones
causadas en embriones expuestos al litio desde el inicio de la gastrulación. Los
análisis histológicos muestran que ocurre una importante desorganización de
todos los tejidos internos a lo largo del eje antero-posterior. Estos resultados
concuerdan con los obtenidos en nuestro estudio. Pueden ser explicados por
cambios que se producen a nivel de la expresión proteica luego de la exposición
al litio durante la gastrulación y neurulación, como se ha reportado para
embriones de anfibio (Lazou y Beis, 1993). Por el contrario, cuando los
tratamientos fueron iniciados desde etapas más avanzadas (HH6; igual tiempo
de exposición), los embriones mostraron morfologías similares a los controles
aunque con defectos fácilmente detectables.
a - Efectos del cloruro de litio sobre la neurulación
Durante varias décadas, estudios clínicos y experimentales han demostrado que
la administración a largo plazo de sales de litio causa múltiples efectos
teratogénicos en embriones animales y humanos (Lewicki et al., 2006; Garcia et
al., 2007; Fujii et al., 2007). También se ha observado que el litio resulta
teratogénico y tóxico incluso en adultos (Sharma e Iqbal, 2005; Zarnescu y
Zamfirescu, 2006; Allagui et al., 2006; Tsaltas et al., 2007). Desde el inicio de
estos estudios se ha comprobado que su administración puede causar defectos
cardíacos y cardiovasculares durante el embarazo aún a niveles terapéuticos en
suero (Warkany, 1988; Cohen et al., 1994; Moore et al., 1995). Además, se ha
observado que el litio produce muerte fetal y defectos del esqueleto (Marathe y
Thomas, 1986), anomalías en ojos y oídos (Wright et al., 1971), paladar hendido
(Szabo, 1970) y NTDs (Jurand, 1988; Hansen et al., 1990) durante el desarrollo
de varios animales de laboratorio. En este trabajo pudimos verificar que el litio
causa defectos en la neurulación en el embrión de pollo. A partir de los
tratamientos con las concentraciones más altas utilizadas (60 y 100 mM)

48
logramos observar un desarrollo embriológico retardado con respecto al proceso
de cierre del tubo neural (Jurand, 1988). También ocurrieron malformaciones del
tubo neural presentando expansiones de la placa neural y surcos neurales
ectópicos. En algunos casos embriones cuyos pliegues neurales no lograban
contacto aún en estadios ya avanzados que debían mostrar tubos neurales ya
cerrados. Algunos embriones expuestos también mostraron el neuroporo anterior
más abierto que los embriones control (Grover y Gupta, 2005). Estos resultados
indican que el litio interfiere de alguna forma con el proceso de neurulación
normal (Hansen et al., 1990; Giles y Bannigan, 1997; Grover y Gupta, 2005).
b - Efectos del cloruro de litio sobre la muerte celular
Otro proceso que vimos afectado durante el tratamiento con LiCl fue la muerte
celular. Los embriones expuestos a concentraciones de 100 mM desde HH6 a
HH11 mostraron un aumento en el número de núcleos picnóticos en el
neuroepitelio. Si bien es sabido que el proceso de muerte celular es un requisito
esencial para que el desarrollo embrionario pueda ocurrir con normalidad,
nuestros resultados parecen indicar niveles anormales de este fenómeno que
afectan la neurulación (Greenblatt et al., 2010). Esta observación también es
apoyada por otros autores que afirman que la muerte celular en el neuroepitelio
inducida por litio puede causar NTDs en el ratón (Giles y Bannigan, 1997). Estos
autores observaron que la muerte celular se vuelve evidente en el tubo neural
incluso poco después de iniciado el tratamiento. Estudios similares muestran que
el valproato, otro fármaco utilizado como anti-convulsivo o para tratamiento de
desórdenes bipolares, también puede inducir mecanismos apoptóticos (Umur et
al., 2012). Por el contrario, numerosas investigaciones apoyan la acción
antiapoptótica y neuroprotectora de estos fármacos (Jorda et al., 2000; Hennion
et al., 2002) y trabajos recientes a nivel molecular indican que ambos
compuestos son inhibidores de los factores de stress que inducen apoptosis (Xu
et al., 2003; Zhong et al., 2006; Kappes et al., 2007). Los efectos antagónicos
pro y antiapoptóticos inducidos por este tipo de fármacos dependen, en último
término, de la dosis y del modelo experimental utilizados (Opiela y Ksiazkiewicz,
2006; Song et al., 2002; Zhang et al., 2005) Según estas dos variables el litio
puede ocasionar efectos muy distintos que incluso pueden llegar a ser

49
antagónicos. Por tal motivo es necesario conocer con exactitud los rangos
terapéuticos y tóxicos para cada modelo experimental al que se administren
estas drogas en función de los efectos que se intenten inducir.
c - Efectos del cloruro de litio sobre la somitogénesis
Algunos embriones tratados mostraron somites anómalos y/o en menor número
con respecto a los controles. Esta alteración fue previamente detectada por
Linask y colaboradores (1998) comprobando que la molécula de adhesión N-
cadherina y las cateninas intracelulares son importantes en la segmentación, la
formación de los somites y del compartimento del miótomo. Los embriones de
pollo y ratón que fueron expuestos a LiCl en dicho trabajo también desarrollaron
somites anómalos. En base a esto ellos sugirieron que la señalización mediada
por Wnt podría estar implicada en la formación de grandes clusters de N-
cadherina/catenina, un factor común en las vías de diferenciación del esqueleto
y el músculo cardíaco, donde interfiere el litio.
d - Efectos del cloruro de litio sobre la proteína MARCKS
Desde 1995 ya se había observado que la disrupción completa de la proteína
MARCKS, uno de los más destacados sustratos de PKC, impide el desarrollo
normal del sistema nervioso central en el ratón (Stumpo et al., 1995). A su vez,
se ha demostrado que exposiciones crónicas al litio producen una reducción
significativa de MARCKS en el cerebro (Lenox et al., 1992; Watson and Lenox,
1996). Esta alteración de la expresión de la proteína persiste aún cuando se ha
detenido la administración del mismo y no se observa luego del tratamiento
agudo con litio. Zolessi y Arruti (2001) demostraron que esta proteína posee un
comportamiento particular durante la flexión de la placa neural ya que adopta
una posición apical en las células del neuroepitelio durante este proceso. Si bien
nuestros resultados no muestran efectos del LiCl sobre la localización o
abundancia subcelular de MARCKS, algunos investigadores han documentado
su disminución en el hipocampo de la rata y en la línea celular inmortal de
hipocampo HN33 bajo tratamiento crónico. La disminución máxima se observó
recién a los 7 días de exposición al litio (Lenox et al., 1996). La exposición de
esta línea celular al litio produce una disminución tiempo y dosis dependiente de

50
MARCKS que a su vez depende de la concentración de inositol presente en el
medio (Watson and Lenox, 1996). Los tratamientos realizados en nuestro trabajo
muestran que la proteína se mantiene apicalizada durante la flexión de la placa
neural aún bajo tratamiento con LiCl. Sin embargo, considerando que la
exposición fue a corto plazo (el fármaco estuvo en contacto con los embriones
sólo durante algunas horas) es posible que el tratamiento no haya sido suficiente
para reproducir los efectos descritos por estos autores. Son numerosos los
estudios que han establecido las diferencias entre los tratamientos a corto y
largo plazo con litio. En 1991, Lenox y colaboradores detectaron una reducción
significativa de la fosforilación de MARCKS por parte de PKC en el hipocampo
luego de la exposición crónica al litio. Este fenómeno persistía incluso después
de finalizado el tratamiento pero no ocurría en las exposiciones a corto plazo. La
alteración en la fosforilación de MARCKS se refleja en una reducción en la
fracción soluble del hipocampo. Se plantea una inhibición transcripcional de los
genes Macs como posible explicación de estas observaciones (Wang et al.,
2001).
e - Mecanismo de acción del litio
Los resultados obtenidos en nuestro laboratorio muestran aumento en la
fosforilación de la enzima GSK3β en respuesta a la exposición con la menor
dosis de LiCl utilizada (20 mM). Si bien no llegamos a obtener mediciones de la
enzima para los tratamientos con dosis más altas (donde podría ser aún más
notorio este aumento), existen varios trabajos que reportan la presencia de esta
inhibición. Aunque hasta ahora no se ha dilucidado por completo el mecanismo
de acción del litio, se ha reunido una considerable evidencia acerca de la acción
de este fármaco y sus moléculas blanco. A dosis por encima del rango
terapéutico, los efectos teratogénicos del litio han sido una herramienta útil para
comprender las vías moleculares que regulan la embriogénesis (Martin et al.,
2011). Cabe destacar que la reducción de la proteína MARCKS producida por la
exposición al litio puede revertirse con la adición de inositol al medio (Lenox et
al., 1996). Por más de una década se sostuvo que el mecanismo de acción del
litio consistía en la inhibición de monofosfatasas de inositol y enzimas
relacionadas. Teóricamente, esta inhibición previene el reciclaje de inositol en el

51
cerebro, ya que secuestra el inositol libre requerido para cascadas de
señalización intracelular mediadas por fosfo-inositol (Gould y Manji, 2005). El
litio, al inhibir la enzima inositol-monofosfatasa provoca la depleción del inositol
libre en un sistema en el cual el inositol exógeno no está disponible fácilmente
(Berridge et al., 1989), situación que puede existir en el embrión temprano
debido a su alta tasa de proliferación y metabolismo. Las células que se
enfrentan a esta depleción no son capaces de responder a señales
extracelulares dependientes del ciclo fosfatidilinositol ya que no logran realizar la
transducción de estas señales (Giles y Bannigan,1999). En 1996 se descubrió
que el litio era capaz de inhibir a la enzima GSK3β (Klein y Melton, 1996;
Stambolic et al., 1996) y más tarde se identificó que el efecto ocurría a través de
la competencia por magnesio (Gurvich y Klein, 2002; Ryves y Harwood, 2001).
Este hallazgo llevó a pensar que la inhibición de esta enzima podría jugar un
papel importante en el tratamiento de trastornos bipolares. Desde hace algunos
años se sostiene la hipótesis que la inhibición de GSK3β constituye un blanco
terapéutico relevante en las propiedades estabilizadoras del ánimo del litio. Una
de sus cualidades más notables es la gran cantidad de vías de señalización que
convergen en esta enzima y, más llamativo aún, el número de blancos biológicos
que posee (Frame y Cohen, 2001; Grimes y Jope, 2001b). Las evidencias
reunidas hasta el momento sugieren que el litio reduce el inositol cerebral, inhibe
la actividad de la enzima GSK3β e interfiere en la formación del complejo
Akt·βArr2·PP2A en neuronas cerebrales dopaminérgicas (debido a la inhibición
de GSK3β) a través de la competencia por la unión al magnesio (Beaulieu y
Caron, 2008; O'Brien et al., 2011). Es posible que también afecte las funciones
de las proteínas G de una manera dependiente de magnesio (Srinivasan et al.,
2004). Se ha demostrado que la reducción de MARCKS debida a niveles
terapéuticos crónicos de litio, depende no solo de la concentración relativa de
inositol sino también del estado de activación del receptor, confiriendo la acción
selectiva del litio en el cerebro cuando el inositol es limitado (Watson y Lenox,
1996). Estudios preliminares de Watson y Lenox (1997) han aportado evidencias
que indican una reducción paralela de la proteína MARCKS inducida por litio y
los niveles de ARNm de MARCKS. Estos resultados, junto con otros que apoyan
lo antes mencionado, constituyen una sólida evidencia del rol que PKC juega a

52
largo plazo en la exposición crónica al litio y también sobre la desestabilización
del ARNm de MARCKS (Brooks et al., 1991, 1992; Lindner et al., 1992; Manji y
Lenox, 1994).

53
VI Conclusiones __________________________________________________
Finalmente, y a modo de conclusión, podemos afirmar que hemos puesto a
punto un método sencillo pero muy útil para el estudio de fármacos durante el
desarrollo temprano en el embrión de pollo. Particularmente establecimos una
nueva forma de aplicación de la droga por disolución en el medio de cultivo que
permitió evaluar los efectos del litio sobre el proceso de neurulación. Hallamos
que la acción de este fármaco es dosis, tiempo y estadio dependiente.
Detectamos diversos defectos en el cierre del tubo neural a concentraciones de
60 y 100 mM. También observamos que aparecen defectos en la gastrulación
cuando los embriones son tratados a altas dosis en etapas tempranas.
Asimismo, se detectó un aumento del porcentaje de muerte celular en el
neuroepitelio en ensayos realizados durante la neurulación (100 mM). Si bien ha
sido documentado que el litio reduce los niveles de MARCKS, no hallamos
diferencias en la localización ni abundancia de la proteína en los embriones
tratados con respecto a los controles. Este hecho podría explicarse por la
brevedad de los tratamientos aplicados de LiCl. A lo largo de los años se ha
reunido evidencia sobre los mecanismos que utiliza este fármaco para su acción.
Hoy en día sabemos que reduce el inositol cerebral, inhibe la actividad de la
enzima GSK3β e interfiere en la formación del complejo Akt·βArr2·PP2A a través
de la competencia por unión al magnesio. A partir de aquí debemos profundizar
el conocimento sobre los mecanismos de acción del litio y otras drogas de uso
médico a fin de dilucidar cómo contribuyen al tratamiento de trastornos
convulsivos o bipolares y logran afectar el desarrollo temprano aún a dosis
terapéuticas.

54
VII Materiales y métodos __________________________________________________
i Obtención de embriones
Los huevos de pollo embrionados, gentilmente donados por la avícola Prodhin,
fueron incubados a 37° C con humedad constante hasta los estadios deseados.
Las dataciones de los embriones fueron realizadas de acuerdo a la tabla de
estadios normales de desarrollo del pollo de Hamburger y Hamilton (1951).
ii Cultivo de embriones
Los embriones fueron cultivados, desde estadios tempranos (HH3) a más tardíos
(HH8) y luego del cultivo se volvieron a incubar bajo las mismas condiciones
iniciales hasta el cierre del tubo neural (HH9 a 11). El método de cultivo utilizado
se denomina “EC” (Chapman et al., 2001). Para llevarlo a cabo se retiran los
huevos de la incubadora y se dejan enfriar por 15-30 minutos (min). Luego se
sostiene el huevo con su eje mayor orientado perpendicularmente sobre el borde
de una placa de petri (100 mm) y se realiza una muesca dando golpecitos en la
parte inferior de la cáscara. Se vacía su contenido rompiendo la cáscara a partir
de la muesca. El blastodermo debe quedar posicionado hacia arriba y lo más
centrado posible en la yema; la membrana vitelina debe permanecer intacta. Se
remueve la albúmina gruesa que cubre el blastodermo con papel tisú plegado
apoyándolo sobre el borde del blastodermo y retirando suavemente hacia afuera.
Es crucial remover la albúmina gruesa ya que su presencia impide que la
membrana vitelina se adhiera correctamente al papel de filtro que este método
utiliza para mantener el embrión bajo tensión. La tensión del blastodermo
asegura un correcto desarrollo in vitro. Una vez retirada la albúmina gruesa se
coloca el papel de filtro con una abertura central (cuadrados de 2 x 2 cm con 4
orificios de perforadora parcialmente superpuestos y centrados; Figura 16) sobre
la membrana vitelina, de manera tal que el embrión quede centrado en el orificio.
Con la ayuda de una pinza se hace presión sobre el papel de filtro para lograr
que la membrana vitelina quede firmemente adherida al mismo. Utilizando una
tijera, se procede a cortar esta membrana alrededor del papel, teniendo cuidado
de cortar completamente por todo el perímetro. Se retira cuidadosamente el

55
papel de filtro de la yema en dirección oblicua con una pinza y se limpia la yema
y restos de albúmina que puedan haber quedado unidas. Para finalizar la
limpieza se sumerge y se retira el papel de filtro con el embrión adherido en una
placa de petri que contiene solución salina. Se repite cuidadosamente el
procedimiento las veces que sea necesario para lograr una limpieza óptima. El
paso siguiente consiste en secar el excedente de solución apoyando el papel de
filtro por sus bordes sobre un papel absorbente. Luego se coloca el mismo en
una placa de cultivo sobre un sustrato semi-sólido de agar-albúmina (ver
Apéndice) de forma tal que la parte ventral del blastodermo mire hacia arriba. Se
debe verificar que no haya burbujas entre el embrión y el medio. Las placas de
cultivo se colocan en una cámara húmeda y se llevan a la incubadora a 37° C
con humedad constante hasta que los embriones alcancen los estadios
deseados (Figura 16).

56
Figura 16

57
Figura 16.- Secuencia de eventos en la preparación de embriones para el cultivo EC. A. Se incuban los huevos hasta que alcancen al estadio deseado. Luego se llevan a temperatura ambiente por 15-30 min y se rompen en una placa de Petri. B. Se utiliza papel tisú para sacar cuidadosamente la albúmina gruesa que rodea la yema, dejando el blastodermo lo más limpio posible. C. Se centra el papel de filtro sobre el blastodermo, intentando alinearlo con el eje céfalo-caudal. D. Se corta la membrana vitelina alrededor del papel de filtro. E. Si la albúmina ha sido removida adecuadamente, el papel de filtro quedará pegado a la membrana vitelina luego del corte. F. Se levanta el papel de filtro fuera de la yema en dirección oblicua. G. Se elimina el exceso de yema del papel de filtro con pinzas desde adentro hacia afuera. H. Para remover la mayor cantidad de yema posible puede sumergirse el papel de filtro en solución salina. I. Se coloca el papel de filtro sobre el sustrato semi-sólido de agar albúmina y se cubre con una tapa (K). Se incuban las placas en un recipiente cerrado con humedad constante. J. Como método alternativo para limpiar la membrana vitelina se puede colocar el papel de filtro directamente sobre la placa con el medio de cultivo y lavar la yema con solución salina utilizando una pipeta Pasteur. Se debe eliminar todo el exceso de líquido al finalizar. L. Embrión cultivado 44 horas bajo el método EC, desde el estadio 4 (Chapman et al., 2001).
iii Tratamientos farmacológicos
Los tratamientos fueron realizados sobre embriones de pollo en cultivo utilizando
LiCl. Como control empleamos concentraciones equimolares de cloruro de sodio
en el vehículo correspondiente para esta droga, el agua. Los embriones fueron
fotografiados en microscopio compuesto o lupa estereoscópica una vez
alcanzados los estadios planificados.
Realizamos ensayos preliminares para ajustar el protocolo de tratamiento
farmacológico, particularmente en cuanto a dos parámetros: modo de aplicación
y dosis. Con respecto al modo de aplicación probamos realizarlo directamente
sobre el embrión, en el centro del blastodisco, con mínimos volúmenes (10-20
µL) así como mediante disolución en el sustrato semi-sólido de agar-albúmina
que el método EC utiliza. También ensayamos distintas dosis de LiCl en
embriones en cultivo y determinamos un rango para el cual se obtenían efectos
nulos o leves a severos. Finalmente decidimos realizar los tratamientos con 3
concentraciones distintas dentro de este rango: 20, 60 y 100 mM.

58
iv Cortes a congelación
Cuando los embriones en cultivo, tratados y control, llegaron a los estadios
planeados fueron procesados para la realización de criocortes. Primeramente
fueron fijados por inmersión con PFA al 4% en PBS para inmovilizar y preservar
la estructura interna del embrión. Luego se realizaron 3 lavados (1 h c/u) con el
fin de quitar por completo el fijador y se comenzó con la crioprotección del
material biológico, utilizando soluciones de sacarosa al 4% primero y más tarde
al 20% manteniendo allí los ejemplares hasta que se sumergieron por completo.
Logrado esto, se incluyeron las muestras en un medio de soporte apto para la
realización de criocortes. En esta ocasión se utilizó gelatina al 7.5% (Stern y
Holland, 1993) que luego se colocó en placas de petri de 35 mm junto con los
embriones (pese a ser un método más laborioso es mejor utilizar gelatina como
medio de inclusión para embriones pequeños ya que permite mejor resolución en
las técnicas de inmunomarcación). Los embriones fueron posicionados
correctamente a medida que la gelatina solidificaba y mantenidos a 4°C hasta su
utilización (máximo 6 días). Para los criocortes se tallaron pequeños bloques de
gelatina con una hoja de bisturí y se colocaron sobre la platina del crióstato
utilizando como pegamento el medio de inclusión por congelación OCT Jung.
Los bloques fueron finalmente congelados con nitrógeno líquido de forma
indirecta. El bloque y la platina fueron introducidos en el crióstato previo a la
realización de los cortes para nivelar la temperatura (-24º C). Se realizaron
cortes a congelación transversales de 5-7 μm de espesor en un crióstato
(Cryocut E, Reichert-Jung, Alemania) a -24º C. Los bloques fueron tallados para
que todas las superficies fueran paralelas, facilitando la obtención de cortes que
fueron levantados en portaobjetos previamente lavados y recubiertos con
gelatina crómica (ver Apéndice). Luego de secados, los cortes se almacenaron
en vasos Coplin a 4ºC hasta su posterior utilización para inmunofluorescencia
(máximo 6 días).
v Inmunodetección sobre cortes
Realizamos inmunodetecciones sobre criocortes con distintos marcadores para
observar el neuroepitelio y sus características.

59
Para comenzar la inmunofluorescencia se debe retirar el excedente de gelatina
sobre los portaobjetos que contienen los cortes. Para esto se lava el medio de
inclusión sumergiendo los portaobjetos en un Coplin con PBS durante 20-40 min
(la duración del lavado depende del tiempo de almacenamiento) en un baño a
37° C. A continuación se retiran los portaobjetos, se secan por sus bordes (sin
dejar secar los cortes) y se realizan separaciones con parafina para delimitar los
distintos grupos sobre los cuales se va a realizar la técnica. Luego de 3 lavados
con PBS (2 min c/u) se realiza una post-fijación, para adherir los cortes al
portaobjetos, colocando paraformaldehído al 4% en PBS por 10 min. Siguen 5
lavados con PBS (2 min c/u). Se bloquean los sitios inespecíficos con albúmina
sérica bovina (BSA) al 1% en PBS por 40 min en cámara húmeda a 37° C.
Posteriormente los cortes fueron incubados en presencia del anticuerpo primario
bajo las mismas condiciones. Los anticuerpos primarios utilizados y sus
respectivas diluciones (en BSA al 1% en PBS) se detallan en la Tabla 1. Para la
incubación se utilizaron entre 20 y 40 μL de las diluciones por grupo de cortes,
por encima de los cuales se colocó una lámina de Parafilm para lograr una
distribución homogénea del anticuerpo. En todos los casos se realizaron
controles negativos por omisión del anticuerpo primario. Los anticuerpos que
utilizamos fueron tanto mono- como policlonales. Cada uno cuenta con sus
ventajas y sus desventajas. La mayor ventaja de los primeros es el
reconocimiento monoespecífico, es decir que el anticuerpo reconoce un epítope
único, lo que permite analizar la presencia y la distribución de un antígeno con
gran precisión; los segundos, por el contrario, reconocen una gran multiplicidad
de epítopes. Las aplicaciones posibles también difieren a la hora de su
utilización. La gran desventaja que presentan los anticuerpos monoclonales del
mismo origen es que no permiten realizar dobles marcados en un mismo
preparado. En consecuencia, para analizar cortes con varios anticuerpos
monoclonales debemos realizar cortes seriados, y analizar con un anticuerpo
cada corte, para luego comparar los cortes adyacentes (Blottière et al., 1995;
Nelson et al., 2000).
60
Tabla1.- Anticuerpos primarios. Nombres, diluciones y estructuras que identifican.
Anticuerpos Dilución Estructura que identifican
Polo52 * 1:5000 MARCKS total
5e 1:400 Glicoproteína de unión celular N-CAM
GSK3β 1:10 Enzima GSK3β activa
pGSK3β 1:10 Enzima GSK3β inactiva (fosforilada) * El anticuerpo policlonal de conejo Polo52 fue producido previamente en el laboratorio en colaboración con el Polo Tecnológico de Pando, Facultad de Química, UdelaR y el Institut Pasteur de Montevideo, a partir de proteína MARCKS recombinante de pollo.
Pasado este tiempo se eliminó el excedente de anticuerpo primario realizando 5
lavados con PBS (2 min c/u) y se procedió a la incubación con el anticuerpo
secundario (diluido en BSA al 1% en PBS) en cámara húmeda a 37° C durante
40 min. Se utilizaron 2 anticuerpos secundarios (cabra): anti IgG de conejo
conjugado a Alexa 488 (1:2000) y anti IgG de ratón conjugado a Alexa 546
(1:2000). Luego se lavó el anticuerpo secundario realizando 5 lavados con PBS
(2 min c/u). La contratinción de los núcleos celulares (ADN) se realizó con
Hoechst 33342 (1:8000) en PBS por 10 min y en algunos casos con un marcador
fluorescente de actina F (Faloidina-Texas Red, o Faloidina-TRITC; 1:5000) en
PBS por 20 min. Por último se realizaron 3 lavados finales (2 min c/u) con PBS.
El montaje del cubreobjetos sobre los portaobjetos se realizó con una pequeña
cantidad de glicerol al 50% en PBS y esmalte de uñas incoloro para sellar. Las
preparaciones se almacenaron a -20° C. La observación de los preparados se
efectuó en un microscopio de fluorescencia Nikon Microphot FXA con los juegos
de filtros correspondientes y los mismos fueron fotografiados con una cámara
digital Nikon D70S.
vi Análisis de imágenes
Las imágenes obtenidas en el microscopio de fluorescencia a partir de los
preparados fueron procesadas en el programa Photoshop CS3. La cuantificación
de intensidad de fluorescencia para pGSK3β también fue realizada con este
programa. Las medidas fueron tomadas a partir de un total de 4 embriones (2
controles y 2 tratados con 20 mM de LiCl), utilizando 3 imágenes por embrión

61
(cefálica, a nivel del tronco y caudal). Sobre cada imagen se realizaron 10
medidas al azar para un área cuadrada de 50 µm de lado. Se promediaron los
10 valores de intensidad para cada imagen y luego se obtuvo un valor promedio
por embrión. Los valores de intensidad de fluorescencia relativa mostrados en la
Figura 15 corresponden al promedio obtenido a partir de los 2 embriones
tratados con 20 mM de LiCl y los 2 embriones control. Se calculó el desvío
estándar para los dos grupos de embriones tomando todas las medidas
realizadas para cada uno.

62
VIII Apéndice __________________________________________________
i Preparación de placas de cultivo EC control y con LiCl
Recolectar albúmina líquida en un vaso de bohemia (romper el huevo por su extremo cónico y eliminar la albúmina gruesa ayudados de una pinza). Preparar solución salina fisiológica para control (0,3 osmolar) y solución para tratamiento farmacológico (LiCl con concentraciones equimolares de sal), llevar a ebullición y colocar el agar agitando hasta que se disuelva. Llevar la albúmina líquida y las soluciones con agar a un baño a 49° C y dejar equilibrar esta temperatura. Mezclar iguales cantidades da ambas preparaciones, revolver y colocar 2 ml de la mezcla en placas de petri de 3,5 cm de diámetro. Dejar reposar a temperatura ambiente hasta que el agar solidifique. Guardar a 4° C hasta su uso (Para más detalles ver Darnell y Schoenwolf, 2000b). ii Soluciones PBS (Eisen): Cloruro de sodio 8 g
Cloruro de potasio 0,2 g Fosfato disódico anhidro 0,912 g Fosfato monopotásico 0,2 g Agua destilada csp 1 L
Paraformaldehído al 3,7 %: Paraformaldehído 7,4 g PBS 200 mL
Disolver el paraformaldehído en PBS y calentando entre 60 y 80°C (no debe
superar este valor) agitando hasta que se diluya por completo y quede transparente la solución. Dejar enfriar y enrasar. Alicuotar y mantener a -20° C hasta su utilización.
Gelatina crómica: Gelatina 0,5 g - Alumbre de cromo 50 mg - Agua destilada csp 100 mL
Diluir el alumbre de cromo en agua calentando sin superar los 60°C. Agregar
suavemente la gelatina con agitación y dejar disolver por completo. Filtrar y guardar a 4° C.
Glicerol al 50% en PBS: Glicerol 500 µL - PBS 5X 200 µL - Agua bidestilada 300 µL

63
XI Bibliografía __________________________________________________ Aderem A (1992) Signal transduction and the actin cytoskeleton: the roles of MARCKS and profilin. Trends Biochem Sci 17:438-43. Afman LA, Blom HJ, Van der Put NMJ, Van Straaten HWM (2003) Homocysteine Interference in Neurulation: A Chick Embryo Model. Birth Defects Research (Part A) 67:421-428. Albert KA, Walaas SI, Wang J, Greenard P (1986) Widespread occurrence of 87 kDa, a major specific substrate for protein kinase. Procl Natl Acad Sci 83: 2822-2826. Albert KA, Nairn AC, Greengard P (1987) The 87-kDa protein, a major specific substrate for protein kinase C: purification from bovine brain and characterization. Proc Natl Acad Sci 84: 7046-7050. Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2004) Biología Molecular de la Célula. 4a edición, España. Ediciones Omega. Allagui MS, Hfaiedh N, Vincent C, Guermazi F, Murat JC, Croute F, El Feki A (2006) Changes in growth rate and thyroid- and sex-hormones blood levels in rats under sub-chronic lithium treatment. Hum Exp Toxicol May; 25(5): 243-250. Anderson DJ (1999) Lineages and transcription factors in the specification of vertebrate primary sensory neurons. Current opinion in neurobiology 9: 517-524. Arbuzova A, Schmitz AAP, Vergères G (2002) Cross-talk unfolded: MARCKS proteins. Biochem J 362: 1-12. Bachiller D, Klingensmith J, Kemp C, Belo JA, Anderson RM, May SR, McMohan JA, McMohan AP, Harland RM, Rossant J, De Robertis EM (2000) The organizer factors Chordin and Noggin are required for mouse forebrain development. Nature 403: 658-661. Beaulieu JM, Caron MG (2008) Looking at lithium: Molecular Moods and Complex Behaviour. Mol Interv Oct; 8(5): 230-41. Review. Berridge MJ, Downes CP, Hanley MR (1989) Neural and developmental actions of lithium: a unifying hypothesis. Cell 59: 411-419. Blackshear PJ (1993) The MARCKS family of protein kinase C substrates. J Biol Chem 268: 1501-04. Blackshear PJ, Wen L, Glynn BP, Witters LA (1986) Protein kinase C-stimulated phosphorylation in vitro of a Mr 80,000 protein phosphorylated in response to phorbol esters and growth factors in intact fibroblasts. Distinction from protein kinase C and prominence in brain. J Biol Chem 261: 1459-69.

64
Blackshear PJ, Tuttle JS, Oakey RJ, Seldin MF, Chery M, Philippe C, Stumpo DJ (1992) Chromosomal mapping of the human (MACS) and mouse (Macs) genes encoding the MARCKS protein. Genomics 14: 168-174. Blackshear PJ, Silver J, Nairn AC, Sulik KK, Squier MV, Stumpo DJ, Tuttle JS (1997) Widespread neuronal ectopia associated with secondary defects in cerebrocortical chondroitin sulfate proteglycans and basal lamina in MARCKS-deficient mice. Exp Neurol 145; 46-61. Blottière HM, Daculsi G, Anegon I (1995) Utilization of activated U937 monocytic cells as a model to evaluate biocompatibility and biodegradation of synthetic calcium phosphate. Biomaterials 16: 497-503. Brooks SF, Herget T, Erusalimsky JD and Rozengurt E (1991) Protein kinase C activation potently down-regulates the expression of its major substrate, 80K, inSwiss 3T3 cells. EMBO J 10: 2497-2505. Brooks SF, Herget T, Broad S, Rozengurt E (1992) The expression of 80K/ MARCKS, a major substrate of protein kinase C (PKC), is down-regulated through PKC-dependent and -independent pathways. J Biol Chem 267: 14212-14218. Chapman SC, Collignon J, Schoenwolf GC, Lumsden A (2001) Improved Method for Chick Whole-Embryo Culture Using a Filter Paper Carrier. Dev Dyn 220: 284-289. Cohen LS, Friedman JM, Jefferson JW, Marshalljohnson E, Weiner ML (1994) A reevaluation of risk of in utero exposure to lithium. Journal of the American Medical Association 271, 146±150. Colas JF, Schoenwolf GC (2001) Towards a cellular and molecular understanding of neurulation. Dev Dyn 221: 117-145. Copp AJ, Brook FA (1989) Does lumbosacral spina bifida arise by failure of neural folding or by defective canalisation? J Med Genet 26: 160-166. Copp AJ, Brook FA, Estibeiro JP, Shum ASW, Cockroft DL (1990) The embryonic development of mammalian neural tube defects. Prog Neurobiol 35: 363-403. Copp AJ, Greene NDE, Murdoch JN (2003b) The genetic basis of mammalian neurulation. Nat Rev Genet 4: 784-793. Dalgic A, Armagan E, Helvacioglu F, Okay O, Daglioglu E, Take G, Unlu A, Belen D (2009) High Dose Cotinine may Induce Neural Tube Defects in a Chick Embryo Model. Turkish Neurosurgery Vol: 19, No. 3: 224-229. Daly S, Millis JL, Molloy AM, Conley M, Lee YJ, Kirke PN, Weir DG, Scott JM (1997) Minimum effective dose of folic acid for food fortifcation to prevent neural-tube defects. Lancet 350: 1666-1669.

65
Darnell DK, Schoenwolf GC (2000a) The chick embryo as a model system for analyzing mechanisms of development. Methods Mol Biol 135: 25-9. Darnell DK, Schoenwolf GC (2000b) Culture of avian embryos. Methods Mol Biol 135: 31-8. Detrait ER, George TM, Etchevers HC, Gilbert JR, Vekemans M, Speer MC (2005) Human neural tube defcts: developmental biology, epidemiology and genetics. Neurotoxicol Teratol 27(3): 515-24. Review. Dhaulakhandi DB, Rohilla S, Rattan KN (2010) Neural Tube Defects: Review of Experimental Evidence on Stem Cell Therapy and Newer Treatment Options. Fetal Diagn Ther 28: 72-78. Erdincler P, Dalgic A, Cepni I, Sanli E, Kafadar A, Tasyurekli M (2009) The effects of high dose progesterone on neural tube development in early chick embryos. Neurol India 54: 178-181. Fernandez EL, Svenson C, Dencker L, Gustafson AL (2004) Disturbing endoderm signaling to anterior neural plate of vertebrates by the teratogen cadmium. Reprod Toxicol 18: 653-660. Frame S, Cohen P (2001) GSK3 takes centre stage more than 20 years after its discovery. Biochem J 359: 1-16. Fujii S, Oku H, Takahashi R, Kanbara Y, Sugasawa J, Ikeda T (2007) Optic nerve dysfunction secondary to long-term use of lithium carbonate. Jpn J Ophthalmol 51: 79-81. Garcia-Castro M, Bronner-Fraser M (1999) Induction and differentiation of the neural crest. Curr Opin Cell Biol 11: 695-698. Garcia C, Mayaudon H, Dupuy O, Leberre JP, Bordier L, Bauduceau B (2007) Silent thyroiditis in a patient under lithium therapy. Rev Med Interne 28: 46-47. Gilbert SF (2006) Biología del desarrollo. 7a edición, México. Panamericana. Giles JJ, Bannigan JG (1997) The effects of lithium on neurulation stage mouse embryos. Arch Toxicol 71(8): 519-28. Giles JJ, Bannigan JG (1999) The effects of lithium on vascular development in the chick area vasculosa. J Anat 194; 197±205. Gordon R (1985) A review of the theories of vertebrate neurulation and their relationship to the mechanics of neural tube birth defect. Rev J Embryol Exp Morphol 89 (Suppl.): 229-255.

66
Gould SE, Grainger RM (1997) Neural induction and antero-posterior patterning in the amphibian embryo: past, present and future. Cell Mol Life Sci 53(4): 319-38. Gould TD, Manji HK (2005) Glycogen Synthase Kinase-3: a Putative Molecular Target for Lithium Mimetic Drugs. Neuropsychopharmacology 30: 1223-1237. Graff JM, Stumpo DJ, Blackshear PJ (1989a) Molecular cloning, sequence and expression of a cDNA encoding the chicken Myristoylated Alanine-Rich C Kinase Substrate (MARCKS). Mol Endo 3: 1903-1906. Graff JM, Rajan RR, Randall RR, Nairn AC, Blackshear PJ (1991) Protein kinase C substrate and inhibitor characteristics of peptides derived from the Myristoylated Alanine-rich C Kinase Substrate (MARCKS) protein phosphorylation site domain. J Biol Chem 266: 14390-14398. Greene NDE, Copp AJ (2009) Development of the vertebrate central nervous system: formation of the neural tube. Prenat Diagn 29: 303-311. Greenblatt DY, Ndiaye M, Chen H, Kunnimalaiyaan M (2010) Lithium inhibits carcinoid cell growth in vitro. Am J Transl Res May; 10; 2(3): 248-53. Grimes CA, Jope RS (2001b) The multifaceted roles of glycogen synthase kinase 3beta in cellular signaling. Prog Neurobiol 65: 391-426. Grover S, Gupta N (2005) Lithium-associated anencephaly. Can J Psychiatry Mar; 50(3): 185-6. Groves AK, Bronner-Fraser M (1999) Neural crest diversification. Curr Top Dev Biol 43: 221-258. Guney O, Selcuki M, Unlu A, Bagdatoglu C (1999) The effect of diazepam on the development of neural tube defects in early chıck embryos. Turk Neurosurg 9: 44-7. Gurvich N, Klein PS (2002) Lithium and valproic acid: parallels and contrasts in diverse signaling contexts. Pharmacol Ther 96: 45-66. Hall BK (1999) The Neural Crest in Development and Evolution. New York: Springer-Verlag, Inc. Hamburger V, Hamilton HL (1951) A series of normal stages in the development of the chick embryo. J Morph 88: 49-92. Hansen DK, Walker RC, Grafton TF (1990) Effect of lithium carbonate on mouse and rat embryo in vitro. Teratology 41: 155-160. Harland R (2000) Neural induction. Curr Opin Genet Dev 10(4): 357-62.

67
Hartwig JH, Thelen M, Rosen A, Janmey PA, Nairn AC, Aderem A (1992) MARCKS is an actin filament crosslinking protein regulated by protein kinase C and calcium-calmodulin. Nature 356: 618-22. Hemmati-Brivanlou A, Melton D (1997) Vertebrate neural induction. Annu Rev Neurosci 20: 43-60. Hennion JP, el-Masri MA, Huff MO (2002) Evaluation of neuroprotection by lithium and valproic acid against oubain-induced cell demage. Bipolar Disord 3; 201-206. Inagaki T, Smith JL, Walker ML, Schoenwolf GC (2000) Neural tube defects. Methods Mol Biol 136: 161-166. Jacobson AG (1991) Experimental analyses of the shaping of the neural plate and tube. Am Zool 31: 628-643. Jessell TM, Sanes JR (2000) Development. The decade of the developing brain. Curr Opin Neurobiol Oct;10(5): 599-611. Jorda EG, Verdaguer E, Canudas AM, Jimenez A, Garcia de Arriba S, Allgaier C, Pallas M, Camins A (2000) Implications of cyclin-dependent kinase in the neuroprotective properties of lithium. Neuroscience 134; 1001-1111. Jurand A (1988) Teratogenic activity of lithium carbonate: an experimental update. Teratology 38; 101±111. Kansagra AJ, Yang E, Nambiar S, Patel PS, Karetzky MS (2011) A Rare Case of Acute Respiratory Distress Syndrome Secondary to Acute Lithium Intoxication. Am J Ther [Epub ahead of print]. Kao KR, Elinson RP (1989) Dorsalization of mesoderm induction by lithium. Dev Biol 132: 81-90. Kappes A, Vaccaro A, Kunnimalaiyaan M, Chen M (2007) Lithium ions: a novel treatment for pheochromocytomas and paragangliomas. Surgery 141; 161-165. Karfunkel, P (1974) The mechanism of neural tube formation. Int Rev Cytol 38: 245-271. Review. Klein PS, Melton DA (1996) A molecular mechanism for the effect of lithium on development. Proc Natl Acad Sci USA 93: 8455-8459. Kobus K, Nazari EM, Rauh Müller YM (2009) Effects of folic acid and homocysteine on spinal cord morphology of the chicken embryo. Histochem Cell Biol 132: 525-532. Lazou A, Beis A (1993) Lithium induces changes in the plasma membrane protein pattern of early amphibian embryos. Biol Cell 77(3): 265-8.

68
Lawson A, Anderson H, Schoenwolf GC (2001) Cellular mechanisms of neural fold formation and morphogenesis in the chick embryo. Anat Rec Feb; 1, 262(2): 153-68. Le Douarin N, Kalcheim C (1999) The neural crest. 2a edición, Cambridge. Cambridge University Press. Lee H, Nagele RG (1985) Neural tube defects caused by local anaesthetics in early chick embryos. Teratology 31: 119-127. Lemaire P, Kodjabachian L (1996) The vertebrate organizer: structure and molecules. Trends Genet Dec; 12(12): 525-31. Lenox RH, Watson DG, Ellis J (1991) Muscarinic receptor regulation and protein kinase C: sites for the action of chronic lithium in the hippocampus. Psychopharmacol Bull 1; 27(3): 191-7. Lenox RH, Watson DG, Patel J, and Ellis J (1992) Chronic lithium administration alters a prominent PKC substrate in rat hippocampus. Brain Res 570, 333-340. Lenox RH, Watson DG (1994) Lithium and the brain: a psychopharmacological strategy to a molecular basis for manic depressive illness. Clin Chem 40(2): 309-14. Review. Lenox RH, McNamara RK, Watterson JM, Watson DG (1996) Myristoylated alanine-rich C kinase substrate (MARCKS): a molecular target for the therapeutic action of mood stabilizers in the brain? J Clin Psychiatry 57 (Suppl) 13: 23-31; discussion 32-3. Lewicki M, Paez H, Mandalunis PM (2006) Effect of lithium carbonate on subchondral bone in sexually mature Wistar rats. Exp Toxicol Pathol 58: 197-201. Lindner D, Gschwendt M, Marks F (1992) Phorbol ester-induced downregulation of the 80-kDa myristoylated alanine-rich C kinase substrate-related protein in Swiss 3T3 fibroblasts. J Biol Chem 267: 24–26. Linask KK, Ludwig C, Han MD, Liu X, Radice GL, Knudsen KA (1998) N-cadherin/catenin-mediated morphoregulation of somite formation. Dev Biol Oct; 1, 202(1): 85-102. Lobach DF, Rochelle JM, Watson ML, Seldin MF, Blackshear PJ (1993) Nucleotide sequence, expression, and chromosomal mapping of Mrp and mapping of five related sequences. Genomics 17(1): 194-204. Manji HK, Lenox RH (1994) Long-term action of lithium: A role for transcriptional and posttranscriptional factors regulated by protein kinase C. Synapse 16: 11-28. Marathe MR, Thomas GP (1986) Embryotoxicity and teratogenicity of lithium carbonate in Wistar rat. Toxicology Letters 34: 115±120.

69
Martin LK, Bratoeva M, Mezentseva NV, Bernanke JM, Remond MC, Ramsdell AF, Eisenberg CA, Eisenberg LM (2011) Inhibition of heart formation by lithium is an indirect result of the disruption of tissue organization within the embryo. Dev Growth Differ doi: 10.1111/j.1440-169X.2011.01313.x. Moore JA, Iehr Scientific Committee (1995) An assessment of lithium using the IEHR evaluative process for assessing human developmental and reproductive toxicity of agents. Reproductive Toxicology 9: 175±210. Moury JD, Schoenwolf GC (1995) Cooperative model of epithelial shaping and bending during avian neurulation: autonomous movements of the neural plate, autonomous movements of the epidermis, and interactions in the neural plate/epidermis transition zone. Dev Dyn 204: 323-337. Nelson PN, Reynolds GM, Waldron EE, Ward E, Giannopoulos KP, Murray G (2000) Monoclonal antibodies. Mol Pathol 53: 111-117. Nieuwkoop PD (1999) The neural induction process: Its morphogenetic aspects. Int J Dev Biol 43: 615-623. Nokhbatolfoghahai M, Parivar K (2008) Teratogenic Effect of Lithium Carbonate in Early Development of Balb/C Mouse. The anatomical record 291: 1088-1096. O'Brien WT, Huang J, Buccafusca R, Garskof J, Valvezan AJ, Berry GT, Klein PS (2011) Glycogen synthase kinase-3 is essential for β-arrestin-2 complex formation and lithium-sensitive behaviors in mice. J Clin Invest Sep; 1, 121(9): 3756-62. Opiela J, Ksiazkiewicz KL (2006) Role of Bcl-2 protein family in a control of apoptosis in ovaries (Polish). Biotechnologia 1: 290-296. Ornoy A (2009) Valproic acid in pregnancy: how much are we endangering the embryo and fetus? Reprod Toxicol Jul; 28(1): 1-10. Price LH, Heninger GR (1994) Lithium in the treatment of mood disorders. New England Journal of Medicine 331: 591±599. Pulikkunnel ST, Thomas SV (2005) Neural Tube Defects: Pathogenesis and Folate Metabolism. JAPI Vol. 53. Rosen A, Keenan KF, Thelen M, Nairn AC, Aderem A (1990) Activation of protein kinase C results in the displacement of its Myristoylated, Alanine-rich Substrate from punctate structures in macrophage filopodia. J Exp Med 172: 1211-1215. Ryves WJ, Harwood AJ (2001) Lithium inhibits glycogen synthase kinase-3 by competition for magnesium. Biochem Biophys Res Commun 280: 720-725. Sadler TW (1998) Mechanisms of neural tube closure and defects. Mental retardation and developmental disabilities research reviews 4: 247-253.

70
Sadler TW (2000) Langman’s medical embryology. 8a edición. Baltimore: Williams and Wilkins. Sanes DH, Reh TA, Harris WA (2006) Development of the Nervous System. 2a edición, USA . Elsevier Inc. Sasai Y, De Robertis EM (1997) Ectodermal patterning in vertebrate embryos. Dev Biol Feb; 1, 182(1): 5-20. Sausedo RA, Smith JL, Schoenwolf GC (1997) Role of nonrandomly oriented cell division in shaping and bending of the neural plate. J Comp Neurol May; 19, 381(4): 473-88. Schoenwolf GC (1982) On the morphogenesis of the early rudiments of the developing central nervous system. Scan Elec Micmsc I: 289-308. Schoenwolf, GC (1988) Microsurgical analyses of avian neurulation: separation of medial and lateral tissues. J Comp Neurol Oct; 22, 276(4): 498-507. Schoenwolf GC (1991) Cell movements driving neurulation in avian embryos. Development 2: 157-68. Schoenwolf GC (1994) Formation and patterning of the avian neuraxis: One dozen hypotheses. En: “Neural Tube Defects”, Bock G, Marsh J (eds). CIBA Foundation Symposium 181. Chichester:John Wiley and Sons, pp. 25-50. Schoenwolf GC, Delongo J (1980) Ultrastructure of secondary neurulation in the chick embryo. Am J Anat May; 158(1): 43-63. Schoenwolf GC, Franks MV (1984) Quantitative analyses of changes in cell shapes during bending of the avian neural plate. Dev Biol 105: 257-272. Schoenwolf GC, Alvarez IS (1989) Roles of neuroepithelial cell rearrangement and division in shaping of the avian neural plate. Development 106: 427-439. Schoenwolf GC, Smith JL (1990) Mechanisms of neurulation- traditional viewpoint and recent advances. Development 109: 243-270. Schoenwolf GC, Alvarez IS (1991) Specification of neurepithelium and surface epithelium in avian transplantation chimeras. Development Jul; 112(3): 713-22. Selleck MA, Bronner-Fraser M (1995) Origins of the avian neural crest: the role of neural-plate epidermal interactions. Development 121: 525-538. Sharma SD, Iqbal M (2005) Lithium induced toxicity in rats: a hematological, biochemical and histopathological study. Biol Pharm Bull May; 28(5): 834-7. Smith JL, Schoenwolf GC (1989) Notochordal induction of cell wedging in the chick neural plate and its role in neural tube formation. J Exp Zool Apr; 250(1): 49-62.

71
Smith JL, Schoenwolf GC (1997) Neurulation: coming to closure. Trends Neurosci 20: 510-517. Smithberg M, Dixit PK (1982) Teratogenic effects of lithium in mice. Teratology 26: 239-240. Smithberg M, Dixit PK, Singer L (1984) Uptake and transfer of lithium in pregnancy and lactation in the mouse. Proc Soc.Exp Biol Med 175: 164-168. Song L, De Sarno P, Jope R (2002) Central role of glycogen synthase kinase-3 in endoplasmic reticulum stress induced caspase-3 activation. J Biol Chem 277: 44701-4708. Srinivasan C, Toon J, Amari L, Abukhdeir AM, Hamm H, Geraldes CF, Ho YK, Mota de Freitas D (2004) Competition between lithium and magnesium ions for the G-protein transducin in the guanosine 5′-diphosphate bound conformation. J Inorg Biochem 98: 691-701. Stambolic V, Ruel L, Woodgett, JR (1996) Lithium inhibits glycogen synthase kinase-3 activity and mimics wingless signalling in intact cells. Curr Biol 6: 1664-1668. Stern CD (2002) Induction and initial patterning of the nervous system - the chick embryo enters the scene. Curr Opin Genet Dev 12: 447-451. Stern CD, Holland PWH (1993) Essential developmental biology a practical approach. Oxford University Press Chapter 21: 192-212. Stumpo DJ, Graff JM, Albert KA, Greengard P, Blackshear PJ (1989) Molecular cloning, characterization, and expression of a cDNA encoding the "80- to 87-kDa" myristoylated alanine-rich C kinase substrate: a major cellular substrate for protein kinase C. Proc Natl Acad Sci 86: 4012-6. Stumpo DJ, Bock CB, Tuttle JS, Blackshear PJ (1995) MARCKS deficiency in mice leads to abnormal brain development and perinatal death. Proc Natl Acad Sci 92: 944-8. Szabo KT (1970) Teratogenic effect of lithium carbonate in the foetal mouse. Nature 225: 7-75. Tanabe Y, Jessell TM (1996) Diversity and pattern in the developing spinal cord. Science Nov; 15, 274(5290): 1115-23. Tolmie J (1996) Neural tube defects and other congenital malformations of the central nervous system. En: Principles and Practice of Medical Genetics. Emery AE, Rimoins DL (eds.). Churchill Livingston, New York. Towler DA, Gordo JI (1988) The biology and enzymology of eukaryotic protein acylation. Ann Rev Biochem 57: 69-99.

72
Tsaltas E, Kontis D, Boulougouris V, Papakosta VM, Giannou H, Poulopoulou C, Soldatos C (2007) Enhancing effects of chronic lithium on memory in the rat. Behav Brain Res Feb; 12, 177(1): 51-60. Umur AS, Selcuki M, Bursali A, Umur N, Kara B, Vatansever HS, Duransoy YK (2012) Simultaneous folate intake may prevent advers effect of valproic acid on neurulating nervous system. Childs Nerv Syst Jan 14 [Epub ahead of print]. Wallingford JB (2005) Neural tube closure and neural tube defects: studies in animal models reveal known knowns and known unknowns. Am J Med Genet C Semin Med Genet 135: 59-68. Wang L, Liu X, Lenox RH (2001) Transcriptional down-regulation of MARCKS gene expression in immortalized hippocampal cells by lithium. J Neurochem Nov; 79(4): 816-25. Warkany J (1988) Teratogen update: lithium. Teratology 38: 593±596. Watson DG, Lenox RH (1996) Chronic lithium-induced down-regulation of MARCKS in immortalized hippocampal cells: Potentiation by muscarinic receptor activation. J Neurochem 67: 767-777. Watson DG, Lenox RH (1997) Chronic lithium exposure leads to the downregulation of both MARCKS mRNA and protein in immortalized hippocampal cells. Soc Neurosci Abstr 23: 1111. Watson DG, Watterson JM, Lenox HL (1998) Sodium Valproate Down-regulates the Myristoylated Alanine- Rich C Kinase Substrate (MARCKS) in Immortalized Hippocampal Cells: A Property of Protein Kinase C-Mediated Mood Stabilizers. JPET 285: 307-316. Weil M, Jacobson MD, Raff MC (1997) Is programmed cell death required for neural tube closure? Current Biology 7: 281-284. Weinstein DC, Hemmati-Brivanlou A (1999) Neural induction. Annu Rev Cell Dev Biol 15: 411-33. Wright TL, Hoffman LH, Davies J (1971) Teratogenic effects of lithium in rats. Teratology 4: 151±156. Xu J, Culman J, Blume A, Brecht S, Gohlke P (2003) Chronic treatment with a low dose of lithium protects the brain against ischemic injury by reducing apoptotic death. Stroke 34: 1287-1292. Zarnescu O, Zamfirescu G (2006) Effects of lithium carbonate on rat seminiferous tubules: an ultrastructural study. Int J Androl Dec; 29(6): 576-82. Zhang WV, Jullig M, Connolly AR, Stott NS (2005) Early gene response in lithium chloride induced apoptosis. Apoptosis 10: 75-90.

73
Zhong J, Lee W, Yao W (2006) Lithium protects ethanol induced neuronal apoptosis. Biochem Biophys Res Commun 350: 905-910. Zolessi FR, Arruti C (2001) Apical accumulation of MARCKS in neural plate cells during neurulation in the chick embryo. BMC Developmental Biology 1: 7. Zolessi FR, Durán R, Engström U, Cerveñansky C, Hellman U, Arruti C (2004) Identification of the chicken MARCKS phosphorylation site specific for differentiating neurons as ser 25 using a monoclonal antibody and mass spectrometry. J Proteom Res 1: 84-90.


















