Estudio de la Interacción entre los Grupos Fosfato del ...
192
Estudio de la Interacción entre los Grupos Fosfato del Ácido Desoxirribonucleico y Fármacos modelo que poseen Grupos Básicos Q.F. Liliana Paola Alarcón Ramírez 2017
Transcript of Estudio de la Interacción entre los Grupos Fosfato del ...

Estudio de la Interacción entre los
Grupos Fosfato del Ácido
Desoxirribonucleico y Fármacos modelo
que poseen Grupos Básicos
Q.F. Liliana Paola Alarcón Ramírez
2017


Estudio de la Interacción entre los
Grupos Fosfato del Ácido
Desoxirribonucleico y Fármacos modelo
que poseen Grupos Básicos
Trabajo de tesis para acceder al grado de
Doctor en Ciencias Químicas de
Liliana Paola Alarcón Ramírez
Director
Prof. Dr. Rubén Hilario Manzo
Comisión Asesora
Prof. Dra. Graciela Panzetta Prof. Dra. Marcela Longhi Prof. Dr. Gustavo Pino
Evaluador externo
Prof. Dra. Ana Lea Cukierman


Dedicado al Único y Soberano
Dios, Jesucristo con todo mi amor


El manuscrito aquí presente representa el trayecto del camino de mi vida
que más ha impactado y trascendido en mí ser, permitiendo la resignificación del
ayer, consolidando el presente a cada paso y afirmando la esperanza de un
futuro mejor; transformando para siempre la visión del mundo, de las sociedades,
de la amistad, de la familia, del amor, de Dios y visualizando el papel de la
Ciencia y de la Investigación en la construcción de un país no solo en cuanto a los
avances productivos, sino también en la formación de nuevas generaciones
apasionadas por el conocimiento y el deseo de ser auténticamente mejores. Es
por eso que deseo expresar mi más profundo agradecimiento:
A mi director de tesis, Prof. Dr. Rubén H. Manzo, a quien admiro y quiero,
gracias por decidir formarme, un maestro;
A los profesores que integran la Comisión Asesora: Prof. Dra. Graciela
Panzetta, Prof. Dra. Marcela Longhi, Prof. Dr. Gustavo Pino, por sus valiosos
aportes y acompañamiento;
A la Prof. Dra. Ana Lea Cukierman por haber aceptado evaluar nuestro
trabajo;
A las instituciones que financiaron la investigación, Conicet, FONCyT,
SECyT;
A la Prof. Dra. María Eugenia Olivera, por su apoyo desinteresado;
Al Prof. Dr. Álvaro Jimenéz k, Prof. Dra. Fabiana Alovero;
A mis compañeros de laboratorio;
Al Prof. Dr. Daniel Allemandi, Prof. Dr. Santiago Palma y mis compañeros de
docencia;
Al personal docente y no docente de los Departamentos de Farmacia,
Orgánica y Fisico-química;
A las chicas y profesoras de Farmacognosia
A Argentina, A Colombia
A los hermanos de la IPUC e IPNJ
A mi familia amada y añorada, a mis amigos del alma, y a quien está
dedicado este trabajo, al Señor Jesús, sin ellos no hubiese sido posible, les debo
todo, no sería lo que soy sin ellos, todo mí amor.



Resumen
El trabajo desarrollado en esta tesis doctoral aborda la caracterización in vitro de la interacción iónica entre los grupos fosfato del Ácido Desoxirribonucleico (ADN), tomado como modelo de ácido nucleico (AN), con las especies protonadas de un grupo seleccionado de fármacos (F) con grupos básicos, a través de la determinación de constantes de afinidad, la reversibilidad de la interacción y el efecto sobre la estructura secundaria de la macromolécula.
Comprendió la caracterización en dispersión acuosa de la sal sódica del ADN de esperma de salmón (comercial (ADN-Na) y de referencia (ADN-NaR)), la forma ácida (HADN) y de los complejos ADN-F con Atenolol (At), lidocaína (Li), timolol (Ti), bencidamina (Be) y propranolol (Pr). F con diferentes propiedades básicas, lipofílicas-hidrofílicas y solubilidad.
El estudio de las constantes de afinidad (Kpi), a través de la determinación de especies en el equilibrio demostró que, una alta proporción de F se encuentra acomplejado con el ADN por condensación iónica [R-FH+], exhibiendo Kpi del orden de 106. A su vez, el acomplejamiento produce una progresiva disminución del potencial electrocinético negativo con el aumento en la proporción del F acomplejante, lo que es consistente con la formación de los [R-FH+].
Los F fueron liberados lentamente desde los sistemas ADN-F en celdas bicompartimentales, mediante una cinética difusional análoga a la informada con otros polielectrolitos (PE) ácidos, demostrando que los complejos se comportan como reservorios de los F, de manera consistente con el alto grado de condensación iónica.
Utilizando dicroísmo circular se observó un comportamiento diferenciado del efecto de la interacción sobre la estructura secundaria del ADN. Esta no se modificó por el acomplejamiento con At, Li y Ti, pero fue significativamente alterada por F más lipofílicos como Be y Pr, revelando modificaciones en la estructura secundaria del ADN de carácter reversible, excepto con el Pr.
La caracterización in vitro de las interacciones ADN-F desarrollada en esta tesis, proporciona una base físico-química con proyecciones en varios campos de interés farmacoterapéutico, tales como la identificación in vivo del efecto de las interacciones descriptas sobre la funcionalidad de los AN, la utilización de ADN como portador de F y la transfección intracelular de AN.


INDICE GENERAL
LISTADO DE SIGLAS Y SÍMBOLOS
LISTADO DE FIGURAS
CAPÍTULO 1: INTRODUCCIÓN GENERAL Y OJETIVOS 1.1. Marco en el que se desarrolla la tesis……………………………………………… 3
1.2. Objetivo General ……………………………………………………………………….. 3
1.3. Importancia del proyecto …………………………………………………………….. 4
1.4. Objetivos Específicos ………………………………………………………………….. 4
1.5. Fuente de ADN a utilizar en el estudio……………………………………………. 5
1.6. Antecedentes……………………………………………………………………………….. 6
1.7. Ácidos Nucleicos (AN)…………………………………………………………………… 7
1.8. Estructura de los AN……………………………………………………………………… 8
1.8.1. Bases Nitrogenadas…………………………………………………………. 8
1.8.2. Pentosas………………………………………………………………………… 9
1.8.3. Grupo fosfato………………………………………………………………….. 10
1.8.4. Estructura primaria………………………………………………………….. 11
1.8.5. Estructura secundaria………………………………………………………. 12
1.8.6. Otro tipo de organizaciones……………………………………………… 14
1.9. Ubicación de los AN en la estructura intracelular……………………………… 15
1.10. Interacciones ADN y Fármacos (F) que afectan su funcionalidad (unión
a surcos e intercalantes)………………………………………………………………… 16
1.10.1 Tipos de unión………………………………………………………………. 17
1.10.1.1. Unión a los surcos…………………………………………. 17
1.10.1.2. Intercalantes………………………………………………… 18
1.11. Interacción Polielectrolito (PE)-F……………………………………………………. 19

1.11.1. Propiedades de las dispersiones PE-F……………………………… 20
1.11.2. Caracterización de la condensación iónica……………………….. 23
1.11.3. Caracterización de los complejos en estado sólido……………. 24
CAPÍTULO 2: METODOLOGÍA GENERAL
2.1. Materiales…………………………………………………………………………………… 29
2.1.1. ADN……………………………………………………………………………. 29
2.1.2. Obtención del HADN a partir de ADN-Na………………………… 29
2.1.3. Fármacos modelo………………………………………………………….. 30
2.1.4. Reactivos……………………………………………………………………… 30
2.2. Metodología…………………………………………………………………………………. 31
2.2.1. Determinación Espectrofotométrica de ADN…………………….. 31
2.2.2. Determinación de pH…………………………………………………….. 31
2.2.3. Potenciometría Diferencial de Barrido (PDB)…………………….. 32
2.2.4. Determinación de Sodio…………………………………………………. 32
2.2.5. Determinación de conductividad……………………………………… 33
2.2.6. Espectroscopia de correlación fotónica (dynamic light
scanning DLS)……………………………………………………………….
33
2.2.7. Electroforesis en gel………………………………………………………. 33
2.2.8. Dicroísmo circular………………………………………………………….. 34
2.2.9. Preparación de complejos………………………………………………. 35
2.2.10. Determinación de Especies en el Equilibrio y de la constante
de afinidad Kpi………………………………………………………………. 36
2.2.11. Evaluación de la reversibilidad de la interacción Efecto del
agregado de sales neutras sobre los equilibrios de formación
de pares iónicos……………………………………………………………… 36
2.2.12. Estudios de Liberación en celdas bicompartimentales de
difusión………………………………………………………………………… 37

2.2.13. Reversibilidad de los cambios en la estructura del ADN
determinado por DC……………………………………………………… 38
2.2.14. Difracción de rayos X…………………………………………………….. 38
CAPÍTULO 3: MATERIALES SELECCIONADOS Y CARACTERIZACIÓN DE ADN-Na Y HDNA EN DISPERSIÓN ACUOSA
PARTE I………………………………………………………………………………. 41
3.1. Introducción………………………………………………………………………………… 41
3.2. Sumario………………………………………………………………………………………. 42
3.3. Materiales y metodologías…………………………………………………………….. 44
3.4. Resultados y Discusión…………………………………………………………………. 44
3.4.1. Caracterización por Espectroscopia UV……………………………. 44
3.4.2. Caracterización por potenciometría diferencial de barrido
(PDB)…………………………………………………………………………… 46
3.4.3. Determinación potenciométrica de Sodio…………………………. 48
3.4.4. Caracterización de la estructura secundaria por Dicroísmo
Circular (DC)………………………………………………………………… 49
3.4.5. Caracterización de tamaño molecular por electroforesis……. 52
3.4.6. Ensayo de permeación de los polímeros………………………….. 53
3.5. Conclusiones……………………………………………………………………………….. 54
PARTE II…………………………………………………………………………….. 55
3.1. Introducción………………………………………………………………………………… 55
3.2. Set de F Modelo…………………………………………………………………………… 55
3.3. Calidad y origen de los materiales utilizados…………………………………… 58

CAPÍTULO 4 CARACTERIZACIÓN DE LOS SISTEMAS ADN-F
EN DISPERSIÓN
4.1. Introducción…………………………………………………………………………………. 61
4.2. Materiales y Metodologías……………………………………………………………… 62
4.3. Resultados y Discusión………………………………………………………………….. 63
4.3.1. pH de las dispersiones ADN-F…………………………………………. 65
4.3.2. Mediciones de conductividad………………………………………….. 66
4.3.3. Mediciones de potencial electrocinético……………………………. 67
4.3.4. Mediciones de electroforesis…………………………………………… 68
4.3.5. Distribución de especies en las dispersiones ADN-F………….. 69
4.3.5.1. Procesamiento de datos para la determinación de
Especies en el Equilibrio y de la constante de
afinidad (Kpi)………………………………………………… 70
4.3.5.2. Determinaciones analíticas en condiciones de
equilibrio………………………………………………………… 72
4.3.6. Reversibilidad del Sistema: Efecto de la relación de
volúmenes compartimento donor ( d ) y receptor ( r )……….. 76
4.3.7. Reversibilidad del Sistema: Efecto de la adición de otras
especies sobre los equilibrios…………………………………………… 78
4.3.8. Difracción de rayos X de los complejos ADN-F…………………… 80
4.4. Conclusiones………………………………………………………………………………... 82
CAPÍTULO 5 CINÉTICA Y MECANISMO DE LIBERACIÓN DE
LOS F DESDE LOS COMPLEJOS ADN-F
5.1. Introducción…………………………………………………………………………………. 87
5.2. Materiales y Metodologías……………………………………………………………… 91
5.3. Resultados y Discusión………………………………………………………………….. 92
5.3.1. Liberación en agua………………………………………………………….

5.4. Conclusiones…………………………………………………………………………………. 100
CAPÍTULO 6 EFECTO DE LA INTERACCIÓN PE-F SOBRE LA
ESTRUCTURA SECUNDARIA DEL ADN
6.1. Introducción…………………………………………………………………………………. 103
6.1.1 Aplicaciones del DC en el análisis estructural del ADN……….. 103
6.2. Materiales y Metodologías……………………………………………………………… 105
6.3. Resultados y Discusión…………………………………………………………………… 106
6.3.1. Efecto de la lipofilicidad de los F sobre la estructura
secundaria…………………………………………………………………….. 106
6.3.2. Reversibilidad de los cambios en la estructura del ADN………. 111
6.3.3. Efecto de la temperatura sobre la estructura secundaria de
los complejos ADN-F………………………………………………………. 113
6.4. Conclusiones………………………………………………………………………………… 114
CONCLUSIONES GENERALES Y PROYECCIONES………………….… 119
BIBLIOGRAFÍA…………………………………………………………………………………. 125
PUBLICACIONES y PRESENTACIONES……………………………………… 145
ANEXO I MANEJO DEL ELECTRODO DE SODIO…………………………………… 149
ANEXO II BASES TEÓRICAS DE DICROÍSMO CIRCULAR…………………….. 155


LISTADO DE SIGLAS Y SÍMBOLOS
AN: Ácidos nucleicos, este término hace referencia tanto al ADN como al ARN.
ADN: Ácido desoxirribonucleico, denominación general que incluye las diferentes
muestras de: ADN-Na, ADN-NaR, HADN y HADNR
ADN-Na : Sal sódica del ácido desoxirribonucleico marca Saporitti
ADN-NaR: Sal sódica del ácido desoxirribonucleico marca Sigma, referencia
ADN-Fx : Complejo iónico Ácido desoxirribonucleico - Fármaco, dónde x representa
el % de neutralización del polímero con el fármaco.
ADN-FH+ : Par iónico
[ADN-FH+ ]: Concentración estequiométrica del par iónico
ARN: Ácido ribonucleico
At: Atenolol
Be: Bencidamina
CD: Compartimento donor
CR: Compartimento receptor
DC: Dicroísmo Circular
ds: doble hélice
F: Fármaco
F : Especie neutra del fármaco básico
[F ]: Concentración estequiométrica de la especie neutra del fármaco
F - : Grupo ionizado del fármaco ácido
FDA US: Administración de Drogas y Alimentos de Estados Unidos

FH : Fármaco ácido
FH+: Grupo protonado del fármaco básico
[FH+]: Concentración estequiométrica de la especie protonada del fármaco
HADN: Forma ácida del ácido desoxirribonucleico obtenido a partir de ADN-Na
HADNR: Forma ácida del ácido desoxirribonucleico marca Sigma, referencia
I+D: Investigación y Desarrollo
Kpi: Constante de formación de par iónico
Li: Lidocaína
Log P: Lipofilicidad
M: Molaridad
mV: mili Voltios
N: normalidad
nm: nanómetros
pb: pares de pases
PDB: Potenciometría diferencial de barrido
PE: Polielectrolito
PE-F: Complejo polielectrolíto-Fármaco
–PO4H: Grupo fosfato ácido
–PO4- : Grupo fosfato ionizado
pKa: Constante de disociación ácida
PM: Peso molecular
Pr: Propranolol
QbD: Quality by desing
r.p.m: revoluciones por minute

Rx: Difractometría de Rayos X de polvos
SPF: Sistema portador de F
Ti: Timolol
USP: Farmacopea de Estados Unidos
UV: Ultravioleta
Kesp: Conductividad específica
Kl: Constante de liberación del fármaco
Kpi: Constante de afinidad
Λ: Conductancia Estequiométrica (µS.cm2/M)
Θ: Ángulo de incidencia de rayos X sobre la muestra
Δθ: Ángulo de elepticidad
λ: longitud de onda de máxima absorción
µS.cm-1:Unidad de conductancia, micro Siemmens por centímetro
ζ : Potencial electrocinético
% p/v: Porcentaje peso en volumen


LISTADO DE FIGURAS
Figura Titulo pp
1.1 Estructura química y pKa de las bases nitrogenadas del ADN y ARN … 8
1.2 Formación del Nucleósido Adenosina ……………………………………………. 10
1.3 Formación del Nucleótido Adenosin 5'-monofosfato ……………………….. 10
1.4 Estructura primaria del ADN ………………………………………………………… 12
1.5 Estructuras secundarias del ADN …………………………………………………. 12
1.6 Estructura terciaria y cuaternaria del ADN…………………..…………………. 14
1.7 Estructura química de netropsina (a) y de cisplatino (b)…………………. 18
1.8 Estructura química de daunomicina …………………………………………….. 19
1.9 Potenciales electrocinéticos (ζ) exhibidos por dispersiones acuosas de
PE cargados con F………………………………………………………………………. 21
1.10 Distribución de especies luego de la partición de los hidrogeles (PE-
F50) con un solvente orgánico………………………………………………………. 23
3.1 (a) Espectro de Absorción y (b) relación absorbancia vs
concentración de ADN-Na en agua MQ pH 7,2……….…………………….. 45
3.2 Perfiles de titulación potenciométrica de ADN-NaR (a), HADN (c),
HADNR (e)…………………………………………………………………………………. 47
3.3 Curvas de calibración del NaCl y ADN-Na para la determinación de
Sodio………………………………………………………………………………………….. 48
3.4 Perfil de DC de ADN-NaR, DNA-Na y HADNR en agua Milli-Q, pH
≈7,0. La concentración del PE en las dispersiones acuosas es de 0,05
mg/mL ……………………………………………………………………………….…….. 50
3.5 Perfil de DC de DNA-Na en agua Milli-Q, pH ≈7,0. La concentración
del PE en las dispersiones acuosas varía en el intervalo de 0,005 a
0,049 mg/mL………………………………………………………………………………. 50

3.6 Efecto de la temperatura sobre el perfil DC de ADN-NaR en agua
Milli-Q, pH ≈7,0. La concentración del PE en las dispersiones acuosas
es de 0,05 mg/mL ……………………………………………………………………… 51
3.7 Electroforesis en gel de agarosa de ADN-Na y HADN, corridas en gel
de agarosa al 0,75% y teñidas con SYBR Green I. ……………………….. 52
3.8 Porcentaje de permeación de polímero a través de membrana de
celulosa (12,000 Da), usando como medio receptor agua………………… 53
3.7 Fármacos seleccionados para estudiar los sistemas (PE-Fx)……………… 57
4.1 Variación de la conductividad específica y del pH con la proporción de
At en la dispersión………………………………………………………………………. 66
4.2 Variación del potencial electrocinético en función del agregado de moles % de F para la dispersión ADN-Fx…………………………………………
67
4.3 Electroforesis de gel en agarosa de complejos ADNR -F100 y 150 en gel
de agarosa 0,75%, revelados con Syber Green I……………………………. 68
4.4 Determinación del tiempo de diálisis necesario para alcanzar el
equilibrio en los sistemas ADN-F, utilizando como modelo el complejo
ADN-Pr43 a pH≈ 7,0. Relación de volumen entre el compartimento d
y r : a) 1/40 b) 1/10…………………………………………………………………… 73
4.5 Efecto de la proporcionalidad de F modelo que neutraliza ADN-Na
sobre la Kpi. Comparación del comportamiento entre las dos fuentes
de ADN-Na ensayadas. a) ADN-Na 0,60 % p/v b) ADN-NaR 0,12 %
p/v ……………………………………………………………………………………………. 75
4.6
Distribución de especies en el equilibrio de los sistemas ADN-At37-92 a
pH≈ 7,0 obtenidos por diálisis………………………………………………………. 76
4.7 Efecto del cambio del volumen del compartimiento receptor en el
equilibrio de los sistemas ADN-F introducidos en el compartimento d,
utilizando como modelo el complejo ADN-At37-92 a pH≈ 7,0……………. 77
4.8 Efecto de la adición de concentraciones crecientes de contraiones
inorgánicos (NaCl), sobre la distribución de especies de los sistemas
ADN-F, utilizando como modelo el complejo ADN-At85. Los valores de

pH corresponden al compartimento d en el equilibrio……………………… 79
4.9 a) Difracción de rayos X de ADN-Na, de los complejos iónicos ADN-Li
y Li como F modelo seleccionado, en el intervalo 2θ/θ. (b) Difracción
de rayos X de ADN-Na, de los complejos iónicos ADN-Pr y Pr como F
modelo seleccionado, en el intervalo 2θ/θ………………………………….. 80
5.1 Perfiles de liberación acumulativa de atenolol en agua. ADN-F50, y F
de referencia en células de Franz en función del (a) tiempo y (b) del
tiempo1/2. ADN-Na se incluyó como blanco………………………… 94
5.2 Perfiles de liberación acumulativa de lidocaína en agua. ADN-F50, y F
de referencia en células de Franz en función del (a) tiempo y (b) del
tiempo1/2. ADN-Na se incluyó como blanco………………………… 95
5.3 Perfiles de liberación acumulativa de propranolol en agua. ADN-F50,
como de F de referencia en células de Franz en función del (a)
tiempo y (b) del tiempo1/2. ADN-Na se incluyó como blanco…………… 96
5.4 Liberación acumulativa de At desde el complejo ADN-Atx (x: 42%,
64 % y 85% y At de referencia equivalente a 100%......................... 98
5.5 Liberación acumulativa de Li desde el complejo ADN-Lix (x: 25%,
42 % y 85% y Li de referencia equivalente a 100%......................... 99
5.6 Liberación acumulativa de Pr desde el complejo ADN-Prx (x: 25%,
42 % y 85% y Pr de referencia equivalente a 100%......................... 99
6.1 Espectro de DC para ARN-A dextrógiro y ADN-A, ADN-B dextrógiro, y
ADN-Z levógiro……………………………………………………………………………. 104
6.2 Perfil de DC de ADN-NaR y ADN-Na en agua Milli Q, pH 7,0…………… 107
6.3 Efecto de la temperatura sobre el perfil de DC de una dispersión
acuosa de ADN-NaR (0,04 mg / ml) a pH 7,0………………………………….. 107
6.4 Perfil de DC de ADN-NaR y ADN-At50 ………………………………..………….. 108
6.5 Perfil de DC de ADN-NaR y ADN-Li50 …………………………..……………….. 109
6.6 Perfil de DC de ADN-NaR y ADN-Ti50 …………………………..……………….. 109
6.7 Perfil de DC de ADN-NaR y ADN-Bz50 …………………………..………………. 110

6.8 Perfil de DC de ADN-NaR y ADN-Pr50 ……………………………………………. 111
6.9 Perfil de DC de ADN-NaR y ADN-Bz50 después de realizar la diálisis
exhaustiva…………………………………………………………………………………. 112
6.10 Perfil de DC de ADN-NaR y ADN-Pr50 después de realizar la diálisis
exhaustiva…………………………………………………………………………………. 112
611 Efecto de la temperatura sobre el perfil de DC de una dispersión
acuosa de ADN-Ti50 …………………………………………………………………… 113
612 Efecto de la temperatura sobre el perfil de DC de una dispersión
acuosa de ADN-Pr50 …………………………………………………………………… 114


INTRODUCCIÓN
GENERAL
Capítulo 1
En este capítulo se describen los objetivos del trabajo doctoral y el contexto
en el cual se desarrollan. También la estructura y conformaciones de los AN,
sus propiedades fisicoquímicas e interacciones con F actualmente conocidas.
Finalmente, se presentan las propiedades de relevancia de las dispersiones
PE-F.


Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
3
1.1. Marco en el que se desarrolla la tesis
El estudio de la interacción entre polielectrolitos (PE) ácidos y básicos y
fármacos (F) ionizables con grupos básicos y ácidos respectivamente, y sus
aplicaciones farmacéuticas se viene desarrollando desde hace más de una
década en el grupo de investigación. En particular su aplicación como sistemas
portadores de fármacos (SPF) basados en la alta afinidad PE-F para generar
complejos iónicos.
Entre los PE ácidos que han sido utilizados previamente en proyectos de
investigación y desarrollo (I + D) se destacan: PE sintéticos (poli-metacrilatos),
semisintéticos (carboximeticelulosa) y PE naturales (ácido algínico y ácido
hialurónico).
La disponibilidad de una plataforma tecnológica y una razonable
infraestructura, constituyeron las bases para proponer el estudio de la
interacción entre los grupos fosfato de los ácidos nucleicos (AN) y F modelo con
grupos básicos protonables.
En este marco, se propusieron para el desarrollo de la tesis los objetivos
que se detallan a continuación.
1.2. Objetivo General
Se propuso entonces:
Realizar un estudio amplio de la interacción reversible de los grupos
fosfato del ácido desoxirribonucleico (ADN), como modelo de AN,
con un grupo seleccionado de F con grupos básicos protonables,
desde un enfoque propio de la química supramolecular.

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
4
1.3. Importancia del proyecto
La disponibilidad de información sistemática sobre la interacción de F
protonables de utilidad farmacoterapéutica con los grupos ionizados de los AN,
que contemple la afinidad y reversibilidad de la interacción, así como los efectos
sobre la estructura primaria y secundaria de la macromolécula es de
importancia en varios campos del conocimiento sobre las propiedades y
aplicaciones de estas importantes macromoléculas.
Entre ellos podemos reconocer:
- Utilización de ADN comercial como portador de principios activos.
- Situaciones farmacoterapéuticas en las que esta clase de F pueden
interaccionar con los AN.
- Generación de propiedades en los AN que favorezcan su inserción intracelular
mediante el apropiado acomplejamiento.
Estos aspectos serán analizados detalladamente a lo largo del desarrollo
del texto de la tesis.
1.4. Objetivos específicos
Se propusieron los siguientes objetivos:
1. Establecer una batería de ensayos que permita una detallada
caracterización fisicoquímica tanto del ADN, como de los complejos ADN-
F obtenidos en dispersión utilizando un grupo seleccionado de F modelo.
2. Determinar cuantitativamente la afinidad ADN-F, la reversibilidad de la
interacción y la cinética y mecanismo de la liberación del F desde la
macromolécula.

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
5
3. Evaluar los efectos de la interacción ADN-F sobre propiedades físicas de
relevancia, como el potencial electrocinético, estructura secundaría,
movilidad electroforética, entre otras.
1.5. Fuente de ADN a utilizar en el estudio
Es importante destacar que el uso de ADN se encuentra aprobado en
Europa como materia prima para productos cosméticos según la decisión
2006/257/CE de la comisión de la Comunidad Europea (Comisión Europea,
2006).
La fuente de ADN utilizada en este estudio, es la sal sódica extraída y
purificada de esperma de salmón, de la especie Oncerhynchus keta, conocido
también como ADN de testículo de salmón. En la actualidad esta sustancia es
comercializada con diferentes propósitos, como suplemento nutricional y
materia prima para productos cosméticos.
En esos campos se proclaman como beneficios funcionales: anti-
envejecimiento para cuerpo y cerebro, aumento del rendimiento deportivo y
recuperación y mejoramiento de la salud hepática y de la piel (Martínez O,
1997; Navarro J, 1999; Holen E, 2005); también se le atribuye la capacidad de
absorber rayos UV (Sasaki Y, 2010). Sin embargo, no existe suficiente evidencia
reportada para los usos indicados.
En el campo farmacéutico se han propuesto films de ADN / Colágeno
para el tratamiento de heridas, los que han sido considerados como promisorios
biomateriales para vendajes (Van Den Beucken J, 2006; Shen X, 2008).
Asociaciones ADN / lípidos han presentado interesante actividad antibacteriana
(Tanaka K, 1996; Inoue Y, 2003; Yamada M, 2005), también se reportan
aplicaciones odontológicas para la cicatrización de heridas alrededor de
implantes orales (Fukushima T, 2001; Getts R, 2016).

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
6
1.6. Antecedentes
Cómo se detallará más adelante en este capítulo, en las unidades del
ADN se encuentran grupos fosfato orientados al exterior de la molécula, estos
poseen un hidrógeno ácido que en solución acuosa se disocia comportándose
como un PE.
La disposición de los grupos fosfato ha sido muy utilizada para generar
interacciones electrostáticas con PE y lípidos catiónicos (formando los
denominados lipoplejos) a los efectos de adecuar, por acomplejamiento, las
propiedades de plásmidos y ADN en relación a la denominada terapia génica
(Rolland A, 2003) que involucra la captación de estas entidades por células pro
y eucariotas. En la actualidad se observa una importante dinámica en la
investigación de esta problemática (Tiera M, 2011; Shimanovich U, 2011). Por
otra parte, la unión electrostática de los grupos fosfato ha sido considerada la
principal interacción con moléculas de amplia distribución en el organismo, tales
como las poliaminas: espermina (Feuerstein B,1990; Utsuno K, 2010) y
espermidina (McGregor T, 2002; Hackl E, 2005).
Respecto a la interacción de los AN con moléculas orgánicas endógenas y
exógenas de pesos moleculares por debajo de 1000 g/mol (Um S, 2006;
Wahyuni E, 2010), un intervalo en el que se encuentra una alta proporción de
principios activos usados actualmente en farmacoterapia, muchas
investigaciones se han focalizado en las moléculas que intercalan el ADN, dada
su importancia en la interrupción de las propiedades biológicas del AN (Cheng
X, 2009; Hajian R, 2009; Shah A, 2009; Mizuno Y, 2010; Stepankova J, 2011;
Getts R, 2016).
Sin embargo, se ha prestado menos atención al estudio de F que son
capaces de interaccionar iónicamente con los AN, a pesar que:

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
7
Una importante fracción de los F utilizados en terapéutica tienen grupos
básicos que se encuentran protonados a pHs fisiológicos, por lo que son
capaces de interactuar como contraiones de los grupos fosfato ionizados
de los AN.
Durante la administración de muchas formas farmacéuticas, tales como
inyecciones intramusculares, gotas oftálmicas y parches transdérmicos,
una concentración apreciable del F está en contacto con las células de
los tejidos involucrados (Banker G, 1996).
1.7. Ácidos Nucleicos (AN)
Se describirán aquí las estructuras y conformaciones de los AN, los sitios
de ionización y las distintas clases de interacciones entre el ADN y F que han
sido informadas en la actualidad.
Los AN son las biomoléculas portadoras de la información genética, con
funciones celulares en la conservación, replicación, y transmisión de esta
información.
Desde el punto de vista químico, los AN son macromoléculas formadas
por polímeros lineales de unidades monoméricas llamadas nucleótidos, unidas
por enlaces éster de fosfato, la secuencia de las unidades no tiene periodicidad
aparente; presentan un alto grado de polimerización, siendo las moléculas más
grandes que se conocen.
Se clasifican en dos tipos de AN, ácidos desoxirribonucleicos (ADN) y
ácidos ribonucleicos (ARN). Se diferencian básicamente en tres aspectos:

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
8
Citosina (C) Timina (T) Uracilo (U)
1) El azúcar (pentosa) que forma parte de los nucleótidos: desoxirribosa
en el ADN y ribosa en el ARN. 2) Las bases nitrogenadas que contienen:
Adenina, Guanina, Citosina y Timina en el ADN; y Adenina, Guanina, Citosina y
Uracilo en el ARN. 3) La estructura de las cadenas: en el ADN se encuentra
organizada en una cadena doble y en el ARN una cadena sencilla.
Históricamente es de relevancia mencionar el descubrimiento de los AN
en 1869 por Meischer, en su trabajo con leucocitos y espermatozoides de
salmón, que la denominó nucleína por encontrarse en el núcleo. Así como, el
descubrimiento de la estructura tridimensional compleja del ADN en 1953 por
Watson y Crick (Watson J, 1953; Crick F, 1954; Manning G, 1978)
1.8 Estructura de los AN
1.8.1. Bases Nitrogenadas
Purinas
Pirimidínicas
Figura 1.1 Estructura química y pKa de las bases nitrogenadas del
ADN y ARN (Acharya S, 2009).
Adenina (A) Guanina (G)

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
9
Las bases nitrogenadas contienen la información genética, son
estructuras planas, aromáticas. Establecen interacciones por formación de
puentes de hidrógeno entre bases complementarias de la cadena doble (Crick
F, 1954; Taylor P, 2015), e interacciones hidrofóbicas que ayudan a estructurar
las cadenas mediante el acomodamiento entre bases vecinas de la misma
cadena (Burkard M, 1999; Ohmichi T, 2002).
Los AN tienen varios centros de ionización presentes en las bases (Figura
1.1 (Acharya S, 2009). Cada una de las bases tiene un sitio de protonación o de
desprotonación a pHs ácido o alcalino, respectivamente. Sin embargo, dado que
se comportan como ácidos o bases débiles, tales ionizaciones no son de
relevancia a pHs cercanos a 7,0.
1.8.2. Pentosas
La unión de una base nitrogenada a una pentosa da lugar a los
compuestos llamados nucleósidos (Figura 1.2). Se efectúa a través de un
enlace glicosídico (Blackburn G, 1996; Acharya S, 2009), con configuración beta
(β) entre el carbono 1 de la ribosa o desoxirribosa, y un nitrógeno de la base, el
1 en las pirimidinas, y el 9 en las purinas, con la pérdida de una molécula de
agua.
Para evitar confusiones en la nomenclatura de nucleósidos y nucleótidos,
los átomos de la pentosa se designan con números seguidos de un apóstrofe
(1',2', 3', 4' y 5'), para distinguirlos de los de las bases, por lo que los enlaces
de los nucleósidos se designan como β (1'-1) en las pirimidinas y β (1'-9) en las
purinas (Blackburn G, 1996; Acharya S, 2009).
UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
10
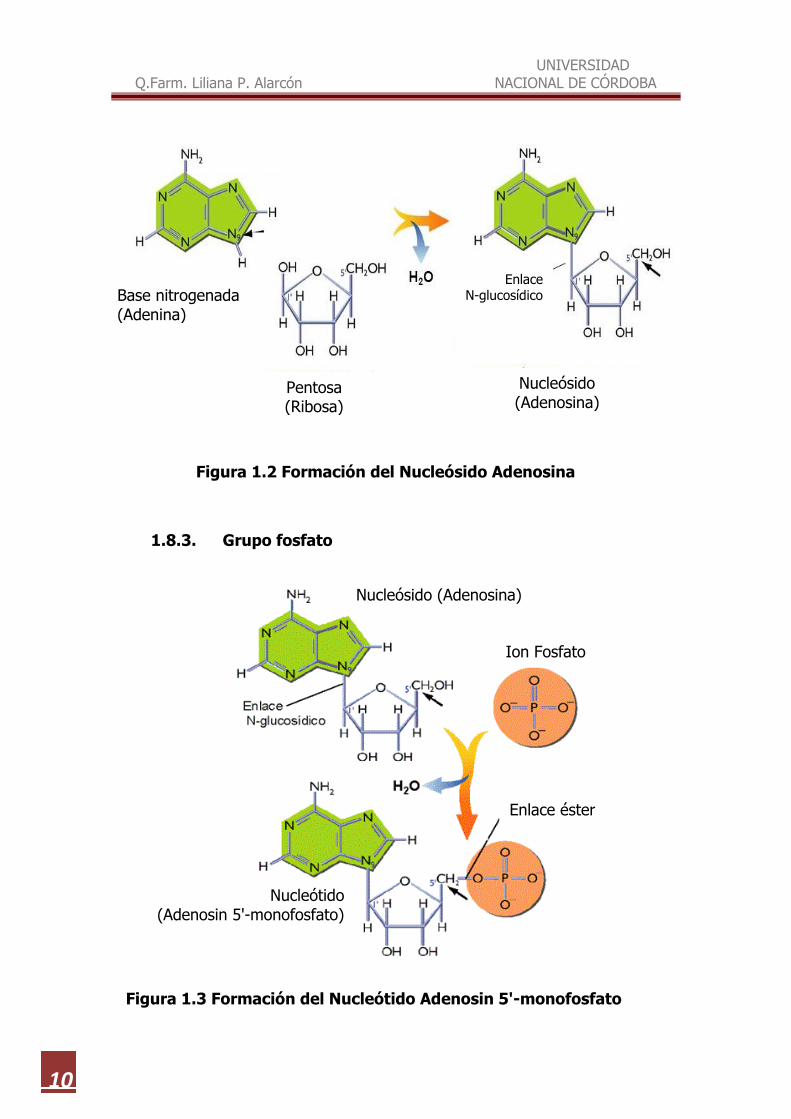
Figura 1.2 Formación del Nucleósido Adenosina
1.8.3. Grupo fosfato
Figura 1.3 Formación del Nucleótido Adenosin 5'-monofosfato
Nucleósido (Adenosina)
Nucleótido (Adenosin 5'-monofosfato)
Ion Fosfato
Base nitrogenada (Adenina)
Pentosa (Ribosa)
Nucleósido (Adenosina)
Enlace éster
Enlace N-glucosídico

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
11
Los nucleótidos (Figura 1.3) son los ésteres fosfóricos de los nucleósidos.
Están formados por la unión de un grupo fosfato al carbono 5' de una pentosa.
A su vez la pentosa lleva unida al carbono 1' una base nitrogenada
Se forman cuando se une un ácido fosfórico a un nucleósido en forma de
ion fosfato (-O-HOPO-O-) mediante un enlace éster con el grupo alcohol -OH
del carbono 5' de la pentosa. Aunque la ribosa tiene tres posiciones en las que
se puede unir el fosfato (2', 3' y 5'), y en la desoxirribosa dos (3' y 5'), los
nucleótidos naturales más abundantes son los que tienen fosfato en la posición
5’. Nucleótidos con fosfato en 3' aparecen en la degradación de los ácidos
nucleicos (Robert H, 2008).
Los grupos de fosfato internucleotídico tienen una pKa de ionización de
1,5; mientras que los fosfatos terminales tienen valores de pKa de 1,5 y 6,5
(Chamberlin S, 2002; Acharya S, 2009). Esto significa que a pH fisiológico los
fosfatos internucleótidos están completamente ionizados.
1.8.4. Estructura primaria
Se trata de la secuencia de desoxirribonucleótidos de una de las hebras.
Estos últimos se unen entre sí mediante el grupo fosfato del segundo
nucleótido, que sirve de puente de unión entre el carbono 5' del primer
nucleótido y el carbono 3' del siguiente nucleótido (Figura 1.4). Es decir, la
secuencia de las unidades de monómeros se ordena 5' 3' (Blackburn G, 1996;
Acharya S, 2009).

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
12
Figura 1.4 Estructura primaria del ADN
1.8.5. Estructura secundaría
Watson y Crick (Watson J, 1953; Crick F, 1954; Manning G, 1978)
postularon el modelo que representa dos hebras de ADN dispuestas juntas en
una doble hélice de diámetro constante. Las hebras están enrolladas en torno a
un eje imaginario, que puede girar hacia la derecha (dextrógira) o izquierda
(levógira), según el tipo de ADN, uniéndose a través de puentes de hidrógeno
de manera complementaria y antiparalela (Figura 1.5).
Figura 1.5 Estructuras secundarias del ADN

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
13
En esta estructura, las bases presentes en una hebra se acoplan con las
bases de la hebra opuesta de manera complementaria y antiparalela a través
de la formación de puentes de hidrógeno (Taylor P, 2015). Esta interacción
determina el acomodamiento en forma de doble hélice (Ohmichi T, 2002). Por
lo tanto, la interacción entre las bases opuestas son las fuerzas predominantes
que estabilizan las estructuras secundarias del ADN. Otro factor que aporta a la
estabilización de la estructura es la orientación del grupo fosfato hacia el
exterior de la hélice que minimizan las fuerzas de repulsión.
El proceso de auto-ensamblaje entre las bases, ocurre así: Adenina (A) y
tiamina (T) forman dos puentes de hidrógeno; y entre guanina (G) y citosina
(C) se forman tres puentes de hidrógeno.
El esqueleto de la estructura espiral del ADN describe dos surcos - mayor
y menor. Los surcos difieren no sólo en tamaño, sino también en polaridad y
características químicas (Nielsen J, 2001). Las bases presentes en cada surco y
la secuencia de unión son específicas. En el surco mayor A-T tiene una
secuencia aceptor-donor-aceptor y G-C tiene una secuencia de aceptor-aceptor-
donor.
Dependiendo del nivel de hidratación del biopolímero y de la naturaleza
de los contraiones presentes, la doble hélice puede encontrarse en diferentes
configuraciones helicoidales: formas A, B o Z (Figura 1.5) (Robert H, 2008).
La forma A se produce en condiciones anhidras (<75% de humedad
relativa) y se encuentra en las esporas bacterianas. Tiene 11 pares de bases
(pb) por giro helicoidal, una elevación helicoidal por pb de 0,26 nm, y un
diámetro de 2,6 nm, las bases nitrogenadas forman un ángulo de 20° con el eje
azúcar – fosfato.
La forma B es la forma más común encontrada en condiciones naturales
y de altos porcentajes de humedad. Tiene 10 pb por giro helicoidal, una

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
14
elevación helicoidal por pb de 0,34 nm, y un diámetro de 2,0 nm, las bases
nitrogenadas forman un ángulo de 90° con el eje azúcar-fosfato. Ambas formas
A y B son hélices formadas hacía la derecha.
La forma Z es una hélice formada hacía la izquierda y se cree que sirve
como un interruptor genético, ya que se encuentra acompañado de la unión de
grupos metilo (-CH3). Tiene 12 pb por vuelta helicoidal, una elevación helicoidal
por pb de 0,37 nm y un diámetro de 1,8 nm.
1.8.6. Otro tipo de organizaciones
Figura 1.6 Estructuras terciaria y cuaternaria del ADN

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
15
La estructura terciaria del ADN se establece cuando la doble hélice es
capaz de enrollarse sobre sí misma, formando una especie de súper-hélice. Esta
disposición se conoce como ADN superenrrollado, y se debe a la acción de
enzimas Topoisomerasas-II, esta estructura da estabilidad y reduce la longitud
de la molécula (Robert H, 2008). En células procariotas está asociado a una
pequeña cantidad de proteínas; en eucariotas el empaquetamiento es más
complejo y compacto comprendiendo desde nucleosoma, collar de perlas y fibra
cromatínica, acompañado por un mayor número de proteínas.
La estructura cuaternaria se conoce como cromosomas, que es la forma
aún más compacta del ADN.
El ADN se encuentra generalmente como doble hélice, pero también
existe en cadena sencilla, en forma circular (Ban C, 1994; Uytterhoeven K,
2002), horquillas (Kopka M, 1985; Feigon J, 1999), tripletes (Bordelon J, 2002)
y cuadruplete (Robert H, 2008).
El ARN funciona generalmente en la forma de cadena sencilla y se
organiza (se pliega a sí mismo) en estructuras secundarias y terciarias (Braña
M, 2001), dependiendo de su tamaño, de la estructura y de las funciones,
existen diferentes tipos, algunos son: ARN mensajero, ARN transferencia, ARN
ribosomal, ARN de interferencia, Micro ARN, ARN interferente pequeño, entre
otros.
1.9. Ubicación de los AN en la estructura intracelular
El Ácido desoxirribonucleico (ADN), es el material genético de todos los
organismos vivos y de la mayoría de los virus. Lleva la información necesaria
para la replicación y para dirigir la síntesis de ARNs y proteínas. Es de
relevancia recordar que, tanto las células eucariotas como procariotas poseen

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
16
una membrana plasmática que las separa del medio y poseen ribosomas dónde
se encuentra ubicado el ARN. Sin embargo, en las células eucariotas el ADN es
lineal y se encuentra ubicado en el núcleo, organizado en forma de
cromosomas (el núcleo está rodeado por una membrana). Mientras que las
células procariotas al carecer de dicha membrana contienen el ADN, que es
circular, compactado en una estructura llamada nuceloide en el citoplasma.
1.10. Interacciones ADN y F que afectan su funcionalidad (unión a surcos e intercalantes)
Existen F que pueden actuar directamente sobre los AN interrumpiendo
la replicación y transcripción (Hasanzadeh M, 2016). Es por esto que el blanco
más utilizado en las farmacoterapias contra el cáncer es el ADN, donde se
busca inhibir la rápida replicación de las células cancerosas. También en
muchas enfermedades como la diabetes, el lupus, la enfermedad de Alzheimer,
entre otras, que pueden atribuirse a la sobre o subproducción de proteínas o la
producción de proteínas mutadas, se busca entonces, modular el proceso de
traducción de proteínas (Sisto M, 2014; Bougas A, 2017).
Se han descripto diferentes mecanismos de interacción entre el ADN y
los F, siendo los más relevantes la unión a surcos y la intercalación. Sin
embargo, el resultado en todos los casos es la modificación de la actividad del
biopolímero (Hasanzadeh M, 2016). El efecto del F sobre el ADN está a menudo
relacionado con la geometría de unión. De este modo, la actividad potencial de
un F podría ser evaluada identificando su tipo de unión al ADN.

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
17
1.10.1 Tipos de unión
Para la mayoría de los F, el tipo de unión que se da con el ADN no
implica formación de enlaces covalentes, por lo tanto, es un proceso de
equilibrio. La constante de unión se puede determinar midiendo la proporción
de las formas libres y unidas del F. Las interacciones están impulsadas por
atracciones electrostáticas, interacciones hidrófobas, enlaces de hidrógeno y
fuerzas de van der Waals (Shaikh S, 2004).
Se han descripto dos tipos de interacciones no covalentes entre el F y el
ADN: la unión a los surcos y la intercalación. También pueden ocurrir
combinaciones de estos modos y la unión no especıfica.
1.10.1.1. Unión a los Surcos
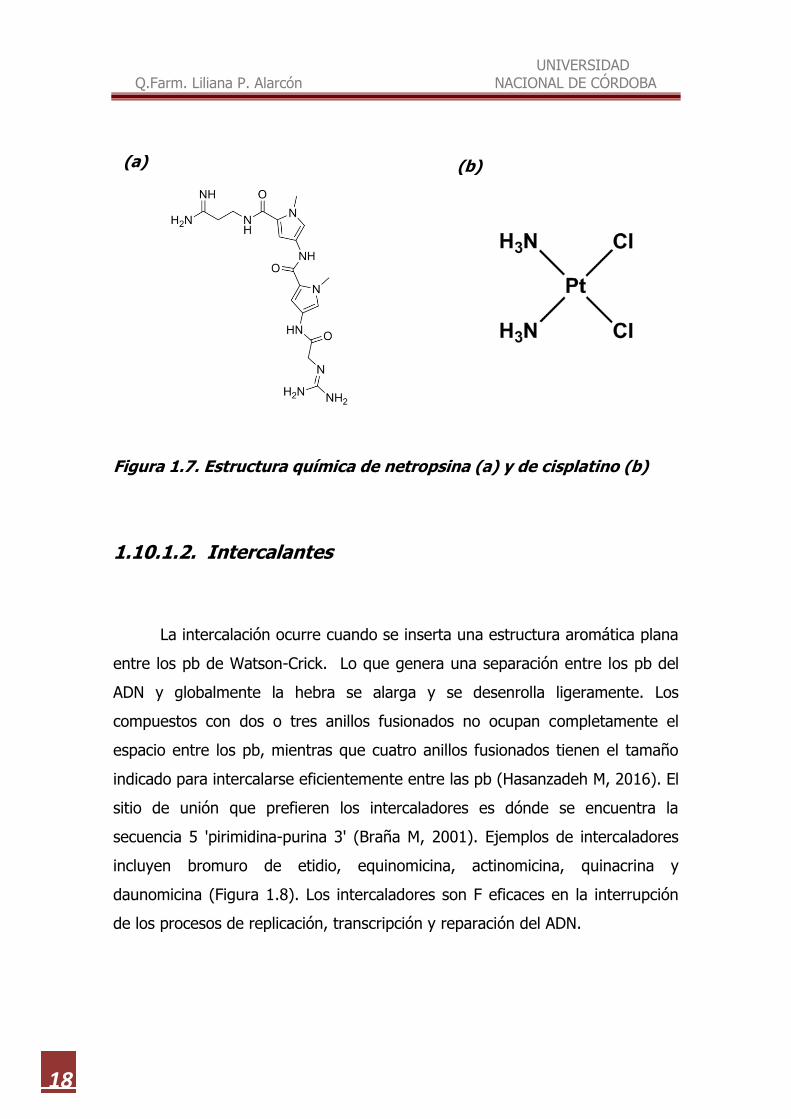
Los F de unión al surco menor son generalmente delgados y en forma de
media luna, sin grupos laterales que producen impedimento estérico. Algunos
ejemplos de ligandos de unión al surco menor incluyen distamicina,
cromomicina, y netropsina (Figura 1.7, a). Estos F no producen cambios
significativos a la estructura del ADN.
Existen menos F de unión al surco mayor, aunque éste es un modo de
unión muy común para que las proteínas regulen la expresión génica. Ejemplos
de los principales F de unión al surco mayor incluyen alquilantes, mostazas y
cisplatino (Figura 1.7, b).
UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
18
Figura 1.7. Estructura química de netropsina (a) y de cisplatino (b)
1.10.1.2. Intercalantes
La intercalación ocurre cuando se inserta una estructura aromática plana
entre los pb de Watson-Crick. Lo que genera una separación entre los pb del
ADN y globalmente la hebra se alarga y se desenrolla ligeramente. Los
compuestos con dos o tres anillos fusionados no ocupan completamente el
espacio entre los pb, mientras que cuatro anillos fusionados tienen el tamaño
indicado para intercalarse eficientemente entre las pb (Hasanzadeh M, 2016). El
sitio de unión que prefieren los intercaladores es dónde se encuentra la
secuencia 5 'pirimidina-purina 3' (Braña M, 2001). Ejemplos de intercaladores
incluyen bromuro de etidio, equinomicina, actinomicina, quinacrina y
daunomicina (Figura 1.8). Los intercaladores son F eficaces en la interrupción
de los procesos de replicación, transcripción y reparación del ADN.
(b) (a)

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
19
Figura 1.8 Estructura química de daunomicina
1.11. Interacción PE-F
Considerando que el principal objetivo planteado consiste en determinar
en qué grado los AN se comportan como polielectrolitos (PE) ácidos, en esta
sección se describen propiedades relevantes de las interacciones PE-F.
Los PE ácidos o básicos en dispersión acuosa reaccionan con moléculas
que posean grupos básicos o ácidos, respectivamente. La reacción ácido base
genera en dispersión acuosa una alta proporción de pares iónicos. Los
equilibrios se representan en las siguientes ecuaciones:
RH + F R- + FH+ [R-FH+] Ec.1
Dónde RH y R- representan el grupo ácido del PE aniónico (los grupos
funcionales pueden ser R-COOH, R-SO3H o -O-HOPO-O-, etc.), que reacciona
con un F básico. F y FH+ representan las especies del F. El resultado de los
equilibrios es la generación del producto de condensación contraiónica
denominado par iónico [R-FH+].

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
20
Del mismo modo, un PE básico interacciona con un fármaco ácido (FH )
RR1R2N + FH RR1R2NH+ + F - [ RR1R2NH+F - ] Ec.2
Donde RR1R2N representa el grupo básico del PE catiónico, en el que R1
y R2 pueden ser hidrógenos o grupos alquílicos, FH y F - representan las
especies neutra e ionizada del FH y [RR1R2NH+F - ] es el producto de la
condensación.
1.11.1. Propiedades de las dispersiones PE-F
De manera general, cuando se neutralizan los grupos ácidos (RH) de un
PE ácido con F en un medio acuoso se generan los siguientes equilibrios:
RH R- + H+ Ec.3
F + H+ FH+ Ec.4
R- + FH+ [R-FH+ ] Ec.5
Como producto de la neutralización del PE se genera una alta proporción
de pares iónicos [R-FH+]. Esta condensación contraiónica, se adjudica a un
balance entre la atracción electrostática de un contraion por la cadena del PE y
la pérdida de entropía traslacional del contraion, debido a su localización en la
vecindad de la cadena de la macromolécula (Dobrynin A, 2005).

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
21
Los PE en dispersión generan cargas positivas o negativas según su
naturaleza básica o ácida. Sus sales inorgánicas exhiben altos potenciales
electrocinéticos (ζ). Del mismo modo, los complejos con contraiones orgánicos
(ej. F modelo) aun cuando mantienen un alto grado de condensación
contraiónica, mantienen esta propiedad.
En nuestro grupo de investigación se han obtenido resultados con
diversos complejos PE-F que han exhibido valores de ζ altos (ver figura 1.9).
Figura 1.9 Potenciales electrocinéticos (ζ) exhibidos por dispersiones
acuosas de PE cargados con F. C: Carbomer; EE: Eudragit E; EL:
Eudragit L. Los subíndices indican el % de F y contraión inorgánico
expresado en moles % que neutraliza los grupos ionizables de cada
PE (Battistini F, 2015).

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
22
Como se señaló, las dispersiones obtenidas con un PE ácido producen un
ζ negativo, así como las obtenidas con PE básicos producen un ζ positivo. El
comportamiento observado puede interpretarse con los modelos teóricos de
doble capa eléctrica, en otras palabras, un modelo constituido por dos fases
interpenetradas: fase macromolecular y fase fluida (Martin A, 1993; Jiménez-
Kairuz A, 2004).
El modelo propone que, en condiciones de equilibrio, un PE ácido que
interacciona con un contraión FH+, conforma un microentorno macromolecular
donde la concentración de especies catiónicas (H+ y FH+) es mayor que en el
seno de la solución. Esto ocurre como consecuencia de la atracción
electrostática de las cargas negativas remanentes del PE que están siendo
neutralizadas con FH+ por condensación iónica, en equilibrio con una proporción
de contraiones con mayor grado de libertad, algunos de los cuales poseen
suficiente energía cinética para migrar al seno de la solución. De esta manera,
el microentorno macromolecular tendrá permanentemente cargas iónicas con
capacidad de interactuar con un campo eléctrico.
Como ocurre con contraiones inorgánicos, la interacción ácido-base entre
los PE ácidos y moléculas de F protonable mantienen un alto ζ que contribuye a
la estabilidad física de las dispersiones.
La concentración de OH-, en cambio, es mayor en el seno de la solución
debido a fuerzas de repulsión entre especies cargadas del mismo signo y a los
requerimientos de electroneutralidad en dicho seno: [FH+] + [H+] = [OH-].
Como se mencionó anteriormente, el modelo supone que la concentración de
especies neutras es igual en ambas fases, tanto en el microentorno
macromolecular como en el seno de la solución.
De acuerdo a este modelo, el microentorno se caracteriza por un valor
de pH menor al del seno de la solución y por una alta proporción de FH+ bajo la
forma de pares iónicos.
Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
23
1.11.2. Caracterización de la condensación iónica
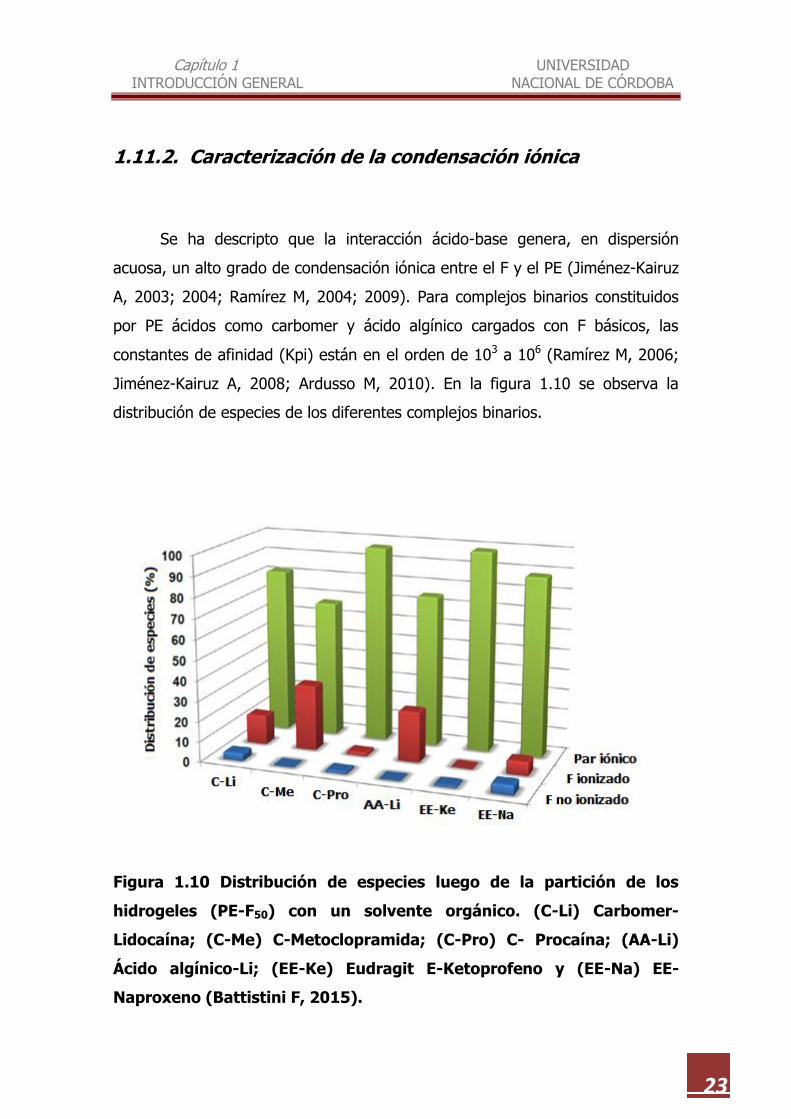
Se ha descripto que la interacción ácido-base genera, en dispersión
acuosa, un alto grado de condensación iónica entre el F y el PE (Jiménez-Kairuz
A, 2003; 2004; Ramírez M, 2004; 2009). Para complejos binarios constituidos
por PE ácidos como carbomer y ácido algínico cargados con F básicos, las
constantes de afinidad (Kpi) están en el orden de 103 a 106 (Ramírez M, 2006;
Jiménez-Kairuz A, 2008; Ardusso M, 2010). En la figura 1.10 se observa la
distribución de especies de los diferentes complejos binarios.
Figura 1.10 Distribución de especies luego de la partición de los
hidrogeles (PE-F50) con un solvente orgánico. (C-Li) Carbomer-
Lidocaína; (C-Me) C-Metoclopramida; (C-Pro) C- Procaína; (AA-Li)
Ácido algínico-Li; (EE-Ke) Eudragit E-Ketoprofeno y (EE-Na) EE-
Naproxeno (Battistini F, 2015).

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
24
Recientemente se han observado los mismos resultados para el
biopolímero ácido hialurónico (Battistini F, 2013). Para los PE básicos como
Eudragit E100 con F ácidos, se observó un grado de condensación iónica mayor
al 88%, algunos de los F ácidos investigados tienen grupos fosfato como la
dexametasona (Quinteros D, 2012; 2011; Guzmán M, 2012).
La reversibilidad de la condensación iónica se ha estudiado a través del
efecto sobre los equilibrios de la incorporación de concentraciones crecientes de
sales inorgánicas, la extracción de los F por diálisis o partición con solventes y
mediante estudios cinéticos de liberación de los F.
La incorporación de sales inorgánicas produce un desplazamiento parcial
del equilibrio promoviendo una mayor liberación de la molécula del F desde el
entorno del PE. Esta reversibilidad confiere a las dispersiones PE-F propiedades
particulares como portador de F de dimensiones nanométricas. Estas
propiedades son de relevancia en aplicaciones farmacotécnicas y
biofarmacéuticas tales como: aumento de la solubilidad aparente de F poco
solubles (Jiménez-Kairuz A, 2002; 2003; Quinteros D, 2008 ), estabilidad
química (Jiménez-Kairuz A, 2004; Esteban S, 2009), resistencia al ataque
enzimático (Quinteros D, 2011), modulación de la liberación del F desde el PE
en presencia de fluidos biológicos (Jiménez-Kairuz A, 2002; 2003; Quinteros D,
2011), interacción con membranas biológicas (bioadhesividad y promoción de la
permeabilidad de F) (Romero V, 2010).
1.11.3. Caracterización de los complejos en estado sólido
La caracterización de las propiedades farmacotécnicas en estado sólido,
conduce a la obtención de información fundamental en el desarrollo de
formulaciones, debido a la mayor estabilidad de los compuestos en estado
sólido que en dispersión. Por esto, es usual encontrar formas farmacéuticas

Capítulo 1 UNIVERSIDAD
INTRODUCCIÓN GENERAL NACIONAL DE CÓRDOBA
25
como polvos para reconstituir antes de su administración (Ansel H, 2011). La
interacción iónica entre el PE y F en estado sólido se ha evidenciado
principalmente mediante espectroscopia FT-IR, análisis térmico y difracción de
rayos X de polvos (Guzmán M, 2012; Ramírez M, 2009). Por difracción de rayos
X se analiza el estado cristalino o amorfo de los complejos PE-F, estos últimos
se obtienen como sólidos amorfos y no muestran las propiedades cristalinas
originales de los F precursores.
Los complejos PE-F en estado sólido pueden obtenerse mediante
diferentes técnicas, como se observa en el esquema 1.1, tales como:
evaporación del solvente, liofilización, coacervación o secado por atomización.
(Jiménez-Kairuz A, 2008; Quinteros D, 2008).
Esquema 1.1 Procedimientos utilizados para la preparación y
obtención de los complejos sólidos PE-F.
PE (Dispersión acuosa o
hidroalcohólica)
F contraión (Solución o dispersión)
Condensación del complejo PE-F
evaporación del solvente
liofilización secado por spray
Producto sólido

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
26

METODOLOGÍA
GENERAL
Capítulo 2
En este capítulo se describen los materiales y metodologías utilizadas tanto en la
caracterización del polímero como de los complejos.


Capítulo 2 UNIVERSIDAD
METOGOLOGÍA GENERAL NACIONAL DE CÓRDOBA
29
2.1. Materiales
2.1.1. ADN
Como fuente de ADN de referencia (ADN-NaR) se utilizó la sal sódica de
ADN de esperma de salmón, conocido también como ADN de testículo de
salmón. El ADN-NaR es una macromolécula de doble hélice, extraída y
purificada de la especie Oncerhynchus keta. Los siguientes valores son
reportados en la ficha técnica del producto: peso molecular aproximadamente
de 2000 pb (pares de bases), contenido de G-C es de 41,2%, temperatura de
fusión (Tm) 87,5°C. (Tm es la temperatura a la cual el 50% de la molécula se
encuentra como doble hebra). El HADNR es la forma ácida del ADN. Ambas
referencias fueron provistas por Sigma Chemical Co., St Louis, USA.
El ADN de esperma de salmón provisto por Parafarm®, Bs. As. Argentina
(ADN-Na), fue usado para la elaboración de lotes de dispersión de mayor
volumen. Según el informe técnico, el ADN es extraído y purificado de la
especie Oncerhynchus keta. El certificado de análisis indica que el contenido de
fósforo es del 87% y el control higiénico de bacterias aerobias y coliformes
conforme.
2.1.2. Obtención del HADN a partir de ADN-Na
El HADN es la forma ácida obtenida de la muestra provista por
Parafarm®, utilizando una resina de intercambio iónico. Se preparó una
dispersión inicial de una cantidad pesada de aproximadamente 500 mg de ADN-
Na en la mínima cantidad de agua Milli-Q aproximadamente 20 mL, que
presentó un pH inicial de 7,0; se eluyó a través de una columna de vidrio

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
30
conteniendo una resina sulfónica de intercambio catiónico (Amberlite IR 120 en
su ciclo de protones (Sigma-Aldrich), R-Ar-SO3H). Luego de la recolección de la
muestra se añadieron sucesivas alícuota de agua Milli-Q a la columna con el fin
de obtener la mayor recuperación del HADN. La dispersión recolectada presentó
un pH de 1,9; posteriormente fue liofilizada (Liofilizador Labbconco®) bajo
vacío de 1 x 10-3 mbar previo congelamiento en freezer.
2.1.3. FÁRMACOS MODELO
Los F seleccionados para obtener los productos ADN-F fueron de grado
farmacéutico. Atenolol (At) y los clorhidratos de lidocaína (Li), propranolol (Pr) y
bencidamina (Bz) y timolol maleato (Ti), fueron provistos por Parafarm®, Bs.
As. Argentina.
2.1.4. REACTIVOS
Los reactivos y solventes utilizados fueron de calidad pro-análisis. Se
preparó una solución al 0,9% P/V de cloruro de sodio (Cicarelli Laboratorios).
Para las electroforesis se utilizó SYBR Green I (SIAL), agarosa,
marcadores de ADN de alto (1kb) y bajo (100 pb) peso molecular provistos por
Sigma Chemical Co., St Louis, USA. Se preparó un buffer de fosfato 10mM
(PBS, 10mM, pH 6,8) acorde a la USP 34-NF 29, 2011. Se utilizó agua Milli-Q
para la realización de todas las determinaciones.

Capítulo 2 UNIVERSIDAD
METOGOLOGÍA GENERAL NACIONAL DE CÓRDOBA
31
2.2. Metodología
2.2.1. Determinación Espectrofotométrica de ADN
Los espectros de absorción fueron determinados en un
Espectrofotómetro Thermo Electron Corporation (Evolution 300), con un
programa Visión proTM Software, en celdas de cuarzo de 1 cm, el barrido fue
realizado en un intervalo de longitud de onda entre 200-400 nm partiendo de
una solución acuosa stock del polímero de concentración de 1,2 mg/mL, se
realizaron diluciones para obtener valores de absorbancia entre 0,2 a 1,0. La
solución stock se prepara sin agitación dejando hidratar el polímero con el 50%
del Agua Milli-Q necesaria para la dispersión. Se determinó el grado de
contaminación proteica a través del índice obtenido de la relación de
absorbancias A260/A280.
2.2.2. Determinación de pH
Las mediciones de pH de las dispersiones acuosas de ADN-Na, HADN y
de la serie de complejos ADN-F, fueron realizadas a temperatura ambiente
utilizando un pH-metro Mettler Toledo Seven Multi equipado con electrodo de
vidrio combinado Ag/AgCl (Mettler Toledo DG 115-SC).

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
32
2.2.3. Potenciometría Diferencial de Barrido (PDB)
Las mediciones se realizaron en un titulador manual y en un titulador
automático (Titrando 905 de METROHM) con un pH-metro Mettler Toledo,
Seven Multi con electrodo de vidrio Ag/AgCl DG 115-SC a 25°C. Se utilizaron
soluciones estandarizadas de NaOH 0,0486 N y HCl 0,0500 N como referencia
(R) y titulante, respectivamente.
La curva de referencia (R) se realizó utilizando como muestra 7 mL de
NaOH 0,0486 N. Para la curva problema (P) se elaboró una muestra con 7 mL
de NaOH 0,0486 N y una cantidad pesada de aproximadamente 100 mg de
polímero, se agita suavemente hasta completa dispersión. El volumen de cada
muestra se completó a 50 mL con agua libre de dióxido de carbono. El registro
de pH se tomó en el intervalo de 0 a 20 mL de reactivo titulante, el intervalo de
dosificación fue de 0,2 y 0,1 mL.
El procesamiento de los datos se realizó utilizando el programa Excel.
2.2.4. Determinación de Sodio
Los equivalentes -O-Na+OPO-O- por gramo de polímero fueron
determinados mediante la cuantificación del catión sodio. Para ello se utilizó un
electrodo selectivo de Sodio marca METTLER TOLEDO perfectIONTM comb Na+
(Mettler Toledo, Germany).
Para el uso de este sensor se elaboró un instructivo de uso (POE004 manejo del
electrodo de Sodio), dónde se consignaron las condiciones de medición
estandarizadas. Para el detalle de la técnica ver Anexo 1.

Capítulo 2 UNIVERSIDAD
METOGOLOGÍA GENERAL NACIONAL DE CÓRDOBA
33
2.2.5. Determinación de conductividad
La conductividad de las dispersiones de ADN y de los complejos ADN-Fx
fue medida utilizando un conductímetro Mettler Toledo Seven Multi equipado
con una celda de conductividad (InLab® 741). Los valores de conductividad
medidos (Siemens –S-), fueron multiplicados por la constante de la celda del
electrodo correspondiente (0,101078 cm-1) para obtener la conductividad
específica (Kesp, S/cm). Las muestras fueron termostatizadas a 25°C.
2.2.6. Espectroscopía de dispersión dinámica dela luz (dynamic light
scattering DLS)
Los coeficientes de difusión y potenciales electrocinéticos (ξ) fueron
determinados usando el equipo Delsa Nano C (Beckman Coulter, Osaka, Japón)
equipado con un láser de diodo de 658 nm a un ángulo de dispersión fijado en
165° y un controlador de temperatura. Los valores de ξ se obtuvieron de la
función de autocorrelación provista por el software Delsa Nano 2.20 (Beckman
Coulter. Osaka. Japan) que emplea la ecuación de Smoluchowsky (Lyklema
J,2003). El dato informado corresponde al promedio de tres mediciones de la
muestra. Las mediciones fueron tomadas por duplicado a 25°C a dispersiones
de PE en concentración entre 0,1 a 1,2% P/V solo y acomplejado con
proporciones crecientes de F (ADN-Fx).
2.2.7. Electroforesis en Gel
El peso molecular de las muestras de ADN fue determinado por
electroforesis siguiendo la metodología descripta en Sambrook et al,1989.

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
34
utilizando un equipo Mini-Sub® Cell GT Cell (Bio Rad Life Science China) con
una fuente POWER PAC BASIC 100-120V/220-240V (Bio Rad Life Science
Singapur). Las muestras se sembraron por duplicado en un gel de agarosa al
0,75% de 10cm de longitud. Previamente el gel fue teñido con 2µL SYBR
Green. Se sembró un volumen final de 12 µL de muestra previamente
incorporada a un buffer de carga que está compuesto por azul de bromofenol
(0,25%), xileno ciamol (0,25%), glicerina (80,0%) y agua Milli-Q (19,5%). El
gel fue sumergido en un aparato horizontal con buffer TBE 1x (Tris base 90
mM, ácido bórico 90 mM y EDTA 2mM) y la corrida se desarrolló durante 1,5
horas a 100 Voltios.
2.2.8. Dicroísmo Circular (DC)
Los espectros fueron medidos en un espectropolarímetro Jasco 81, se
mantuvo el compartimento de la celda con un flujo continuo de nitrógeno de
alta pureza. Los parámetros establecidos para el equipo fueron: velocidad de
corrida: 50 nm/ min, número de acumulaciones: 3, tiempo de respuesta: 4 s,
ancho de banda: 2 nm y longitud de onda: 190 - 350 nm. Se utilizaron celdas
de cuarzo de 0,02 cm (Sigma Chemical Co., St Louis, USA), en las cuáles se
colocaron 0,5 mL de sistema a evaluar, siendo inicialmente agua Milli-Q, luego
el blanco constituido por F y agua Milli-Q y finalmente la muestra. Se realizaron
diluciones, tanto de la muestra original como de los blancos, de manera que la
concentración final fuese de 0,3 mg/mL, utilizando como medio de dilución
agua Milli-Q desgasificada y filtrada. El software permite realizar una corrección
del perfil de la muestra sustrayendo el perfil del blanco correspondiente. Cada
espectro representa el promedio de 3 determinaciones por muestra. Las
determinaciones fueron tomadas a temperatura ambiente.
Antes de proceder con las determinaciones de DC, a las muestras se les
determinó el espectro UV verificando un valor de absorbancia entre 0,8 a 1,0 a

Capítulo 2 UNIVERSIDAD
METOGOLOGÍA GENERAL NACIONAL DE CÓRDOBA
35
la longitud de onda de máxima absorción del ADN (260 nm). El procesamiento
de los datos se realizó utilizando el programa Sigma Plot 11 y fueron
representados como las variaciones en el ángulo de elipticidad (Δε).
Se realizaron determinaciones luego que las muestras fueran sometidas
a un ciclo de temperatura (calentamiento - enfriamiento) controlado por un
baño termostatizado adaptado al equipo que recubre el soporte de la celda, el
control de la temperatura dentro de la muestra se efectuó con la ayuda de un
termómetro digital. El ascenso paulatino de temperatura se llevó hasta un
máximo de 82°C, se sostuvo por 4 minutos, y se dejó enfriar a temperatura
ambiente.
2.2.9. Preparación de Complejos
Se obtuvieron las dispersiones acuosas de los complejos binarios ADN-Fx
y HADN-Fx, mezclando soluciones acuosas de las sales de los F con dispersiones
del polímero previamente hidratado sin agitación durante 24 horas y en
relaciones tales que el polímero, en el sistema, estuviera a una concentración
final de 0,12% P/V para ADN-NaR y 0,6% P/V en el caso de ADN-Na y HADN.
En todos los casos el subíndice “x” corresponde al porcentaje de los grupos
fosfato del PE neutralizados con el F contraión, siendo x = % dentro del
intervalo de 25% a 150%. Las dispersiones se agitaron suavemente durante 1
hora y se almacenaron durante 15 horas a 4-8°C. Antes de utilizarlas para las
determinaciones se dejan temperar al ambiente y se verifica el pH del
complejo, ajustando de ser necesario a valores de pH cercanos a la neutralidad,
adicionando HCl 1N.
Los complejos binarios en estado sólido, se obtuvieron por liofilización de
las dispersiones PE-F bajo vacío de 1 x 10-3 mbar, previo congelamiento en
freezer de -80°C.

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
36
2.2.10. Determinación de Especies en el Equilibrio y de la constante de
afinidad Kpi
Las proporciones de las especies [F ], [FH+] y [R - FH+] de la serie de
complejos ADN-Fx se determinaron mediante el equilibrio de diálisis de las
dispersiones usando tubos de membrana de acetato de celulosa previamente
hidratado en agua Milli-Q durante 30 min (12,000 Da; Sigma, St Louis, MA,
USA). En el tubo de diálisis [compartimento donor ( d )] se incorporan
exactamente 10 mL de las dispersiones acuosas PE-F con una concentración de
PE de 0,12% o 0,6% P/V y distintos porcentajes de neutralización del polímero
con el F. El tubo sellado con clips fue introducido en un vaso de precipitado
sellado con Parafilm® [compartimento receptor ( r )] que contiene 100mL
(1/10) o 400 mL (1/40) de agua Milli-Q desgasificada.
Las fases estuvieron en contacto durante 24 horas hasta que alcanzaran
el equilibrio. Durante este tiempo, los sistemas se mantuvieron en el equipo de
agitación termostatizado a 25°C y constante agitación a 32 rpm.
Finalmente, se registró el pH de los compartimentos d y r, y se midió la
concentración del F en el compartimento r mediante espectrofotometría UV a
la longitud de onda de máxima absorción de cada F.
2.2.11. Evaluación de la reversibilidad de la interacción: Efecto del
agregado de sales neutras sobre los equilibrios de formación de pares
iónicos
El efecto de la adición de NaCl se evaluó mediante diálisis. Se prepararon
sistemas de la siguiente composición ADN-At85-NaCly, el subíndice “y”
corresponde al porcentaje de los grupos fosfato del PE neutralizados con el Na+

Capítulo 2 UNIVERSIDAD
METOGOLOGÍA GENERAL NACIONAL DE CÓRDOBA
37
contraión, siendo y= 25%, 50% y 100%. La diálisis se efectuó tal como se
describe en el numeral 2.10 para una relación de volumen 1/40 ( d/r ). Se
seleccionó At como F modelo para este ensayo, debido a que, en estudios
previos, presentó los valores de constantes de afinidad más altos; por lo tanto,
los resultados pueden ser extrapolados a sistemas con menores valores de
afinidad.
2.2.12. Estudios de Liberación en celdas bicompartimentales de
difusión
Los estudios de liberación in vitro de los F desde las dispersiones de los
complejos ADN-Fx, se llevaron a cabo utilizando celdas de difusión
bicompartimentales del tipo Franz (ver figura 5.1), separadas por una
membrana semipermeable de acetato de celulosa (12,000 Da; Sigma. St
Louis.MA. USA) previamente hidratada en agua Milli-Q durante 30 min. El área
efectiva de difusión fue de 1,25 cm2. En el compartimento receptor se colocan
16 mL de medio receptor (agua Milli-Q o solución de NaCl 0,9% P/V)
desgasificado y atemperado a 37°C. En el compartimento donor se introduce
exactamente 1mL de dispersión ADN-Fx o una solución acuosa de F en una
concentración equivalente a la utilizada en el complejo. La concentración se
establece a fines de garantizar las condiciones de sumidero, es decir, que
prevalezca la difusión del F en dirección donor-receptor.
El sistema se mantiene termostatizado a (37,0 ± 0,1) °C y el
compartimiento receptor es sometido a una agitación magnética suave y
constante. Se estableció una frecuencia de muestreo durante 3 horas de la
siguiente manera: 5, 10, 15, 30, 60, 90, 120 y 180 min, extrayendo 0,9 mL del
medio receptor con reposición de medio fresco y atemperado.

UNIVERSIDAD
Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
38
La concentración de F liberado se cuantifica por espectrofotometría UV a
la longitud de onda de máxima absorción de cada F. Los resultados informados
son producto del promedio de tres mediciones independientes entre sí.
2.2.13. Reversibilidad de los cambios en la estructura del ADN
determinado por DC
Los complejos ADN-Pr50 y ADN-Bz50 fueron sometidos a una diálisis
exhaustiva para recuperar el ADN sin F. Para llevar esto a cabo, se colocaron 10
mL del complejo en dispersión en el tubo de diálisis, tal como se describe en el
numeral 2.10 para una relación de volumen 1/40 ( d/r ) por 24 horas, usando
como medio receptor buffer fosfato sódico 10 mM a pH 7,0; el cuál fue
renovado en su totalidad cada 2 horas durante las primeras 8 horas. La
cuantificación de F, que migra al compartimento receptor, se determinó por
espectroscopia UV y las modificaciones estructurales del ADN en el medio
donor, por DC.
2.2.14. Difracción de rayos X
Los patrones de difracción de Rx fueron obtenidos usando un
difractómetro (PANalyical XPert Pro, Países Bajos) provisto con un software
específico para el procesamiento de datos y utilizando una lámpara de Cu (Kα)
(λ=1,5418 Å). Las muestras sólidas se colocaron en un porta-muestra de
aluminio y se estableció un intervalo de barrido entre 5 y 7 en 2θ/θ y una
velocidad de barrido de 0,04 2θ/s. Previo al ensayo las muestras fueron
pulverizadas a polvo fino.

Materiales
seleccionados y
Caracterización de
ADN-Na y HDNA en
dispersión acuosa
Capítulo 3
En este capítulo se describen las materias primas: PARTE I del ADN generalidades,
selección de las metodologías para caracterizar su funcionalidad como PE, resultados
y discusión de los hallazgos. PARTE II descripción de los F seleccionados.


Capítulo 3 MATERIALES GENERALIDADES UNIVERSIDAD CARACTERIZACIÓN ADN Y HADN NACIONAL DE CÓRDOBA
41
PARTE I
3.1. Introducción
Como se señaló previamente en el capítulo 1, el ADN se encuentra
aprobado como materia prima para uso en productos cosméticos según la
decisión 2006/257/CE de la comisión de la Comunidad Europea (Comisión
Europea, 2006).
Para evaluar la pureza y principales características estructurales de los
ADN a utilizar, se desarrolló una serie de ensayos utilizando cuatro muestras: la
sal sódica ADN-NaR y la forma ácida HADNR, obtenidas de Sigma-Aldrich y
denominadas como referencias (R), la sal sódica ADN-Na provista por
Parafarm®, Bs. As. Argentina y la forma ácida HADN obtenida a partir del ADN-
Na.
El objetivo de este capítulo es describir y analizar la caracterización
fisicoquímica de los ADN descriptos anteriormente, para la cual se seleccionaron
técnicas desarrolladas y aplicadas en anteriores estudios con otros PE, así como
la exploración de nuevas técnicas que permitieran reunir la mayor información
posible sobre la identidad, pureza y propiedades específicas de las sustancias
en estudio como se describe en el esquema 3.1.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
42
Esquema 3.1 Batería de ensayos establecida para la caracterización de las muestras de ADN. Donde UV: caracterización espectroscópica, PDB: Potenciometría diferencial de barrido, DC: Dicroísmo circular
3.2. Sumario
Mediante la titulación por PDB de las dispersiones acuosas de ADN-Na a
0,60 % P/V y ADN-NaR a 0,12 % P/V se determinó que en ambos casos los
grupos fosfato están salificados. La determinación potenciométrica de Na+
reveló que ADN-Na contiene 3,71 meq/g y ADN-NaR contiene 3,51 meq/g de
grupos fosfato salificados. Ambas muestras exhiben al UV, un máximo de
absorción a 260 nm y un valor del cociente de absorbancias de A260/ A280
cercano a 1,8; indicando un aceptable grado de pureza.
ADN-Na
HADN
UV
Identificación y pureza.
ds o ss
PDB
Grado de salificación
Peso Equivalente
Determinación de Na
Número de equivalentes
DC
Tipo de estructura
conformacional
Electroforesis
Tamaño medido en pb.
Potencial Electrocinético
Carga, estabilidad

Capítulo 3 MATERIALES GENERALIDADES UNIVERSIDAD CARACTERIZACIÓN ADN Y HADN NACIONAL DE CÓRDOBA
43
Por DC se reveló que ambas muestras presentan una estructura
secundaria tipo-B, la cual es característica de la estructura nativa de doble
hélice del ADN. La electroforesis en gel indicó que el ADN-NaR exhibe una banda
correspondiente a una cadena que contiene entre 2000 a 8000 pares de bases
(pb), mientras que el ADN-Na corresponde a un tamaño menor a 100 pb. Las
dispersiones exhibieron potenciales electrocinéticos de -28,0 ± 1,0 para ADN-Na
al 0,60% P/V y -35,9 ± 1,6 para ADN-NaR al 0,12% P/V.
Por último, la permeación a través de una membrana semipermeable de
acetato de celulosa (12,000 Da; Sigma. St Louis.MA. USA) permitió evidenciar la
pureza de las sales sódicas del ADN en cuanto a la ausencia de fragmentos
correspondientes a oligonucleótidos, mientras que las formas ácidas se
encuentran contaminadas con dichos fragmentos que migraron a través de la
membrana.
A continuación, en la tabla 3.1 se resumen los resultados de los
parámetros medidos durante los ensayos de caracterización.
Tabla 3.1 Sumario de información obtenida en la caracterización de
ADN-Na implementando la batería de ensayos definida.
Parámetro Teórico ADN-Na ADN-NaR
λ max 260 260 260
A (50µg/mL) 1,00 1,00 1,08
a (µg/mL)-1 cm-1 0,0200 0,0209 0,0218
Relación de A 260:280
1,80 1,72 1,88
pH --- 7,20 7,25
PM pb --- < 100 2000-8000
ζ (mV)2 --- -28,0 ± 1,0 -35,9 ± 1,6
Equivalentes/g --- 3,71 x10-3 3,51 x10-3
DC Doble hélice, tipo-B

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
44
3.3. Materiales y metodologías
Las descripciones detalladas de las metodologías utilizadas se informaron
en el capítulo 2. Metodologías generales.
3.4. Resultados y Discusión
3.4.1. Caracterización por Espectroscopia UV
Esta técnica nos permitió obtener, a través de la longitud de onda de
máxima absorción, información sobre la identidad de la muestra. Algunos
autores (Lehninger A, 2008; García JP 2014) han definido el coeficiente de
extinción y la absorbancia que una solución 50µg/mL debe presentar, en caso
de ser un ADN de hélice sencilla o doble. A través del cálculo del cociente de las
absorbancias en 260/280 se obtiene información sobre las condiciones de
pureza, específicamente contaminación con proteínas y otros compuestos
orgánicos.
En la figura 3.1 se observa el perfil del espectro de absorción del ADN-
Na, donde se evidencia un máximo de absorción a una longitud de onda de 260
nm. En la tabla 3.2 se expresan los resultados de los parámetros mencionados
para todas las muestras analizadas, donde se constata que todos los polímeros
exhiben un máximo de absorción a 260 nm, un coeficiente de extinción cercano
a 0,0200 y una absorbancia de 1,00 para una solución de 50µg/mL de
concentración, resultados que corresponden a un ADN de doble hélice. El valor
del cociente de absorbancias de A260/ A280 cercano a 1,80; indica un grado de
pureza aceptable para la muestra de proveedor nacional y un alto grado de
pureza para la referencia certificada.

Capítulo 3 MATERIALES GENERALIDADES UNIVERSIDAD CARACTERIZACIÓN ADN Y HADN NACIONAL DE CÓRDOBA
45
Figura 3.1 (a) Espectro de Absorción y (b) relación absorbancia vs
concentración de ADN-Na en agua Milli-Q pH 7,2.
Tabla 3.2 Parámetros de identificación y pureza en agua Milli-Q a pH
7,2
1. La desviación estándar fue calculada para las mediciones de cada muestra de manera
independiente, por la similaridad entre los valores de los errores se reporta solo uno.
Al realizar el monitoreo de control de estos parámetros del ADN durante
el almacenamiento, se observó que la muestra sufre algún proceso de
degradación, ocasionado probablemente por la continua apertura del envase y
exposición del material al ambiente. Razón por la cual se implementa una
frecuencia de análisis cada 3 meses y se fortalecen las medidas sanitarias en la
manipulación.
Parámetro Teórico ADN-Na HADN ADN-NaR HADNR
σ1
Lote 1 Lote 2 Lote 1 Lote 2 Lote 1
λ máx. 260 260 260 260 260 260 -----
A (50µg/mL)
1,00 1,04 1,00 1,22 1,08 1,11 ± 0,01
a (µg/mL)-1 cm-1
0,0200 0,0208 0,0209 0,0235 0,0218 0,0219 ± 0,0005
Relación de A 260:280
1,80 1,62 1,72 1,49 1,76 1,88 ± 0,03
0
0,2
0,4
0,6
0,8
1
1,2
200 220 240 260 280 300 320
A
λnm
y= 20,8 x - 0,0056 R² = 0,998
0
0,2
0,4
0,6
0,8
1
1,2
1,4
0 0,02 0,04 0,06
A
mg/mL
a b

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
46
Los parámetros del HADN obtenido por resina, revelan que su paso por
esta degrada la molécula hasta oligonucleótidos, por lo tanto, se alejan del
requerimiento de doble hélice y del grado de pureza aceptable.
3.4.2. Caracterización por potenciometría diferencial de
barrido (PDB)
La PDB es útil para caracterizar las propiedades ácido-base de sistemas
que estén constituidos por una molécula ionizable en solución o por una mezcla
de las mismas. Adicionalmente, permite identificar y cuantificar los grupos
ácidos y/o básicos presentes a través de las áreas que describen los perfiles.
Permite dar conclusiones acerca de la pureza iónica de los compuestos.
Las figuras 3.2 a, c y e muestran los perfiles de titulación de las
muestras de ADN-NaR, HADN y HADNR respectivamente, mientras que las
figuras b, d y f muestran cualitativamente el área delimitada por el diferencial
de pH entre la solución de referencia y la muestra de PE en función del
volumen del titulante. El área positiva del perfil de ADN-NaR de los grupos
fosfato (-O-Na+OPO-O) presentes en el PE están en su forma ionizada (-O- -
OPO-O-), es decir que los fosfatos se encuentran salificados, mientras que el
área negativa para los HADN y HADNR indican que los grupos ionizables del PE
se encuentran en su forma ácida (-O-HOPO-O). El resultado de la muestra
HADN confirma la eficiencia de las resinas de intercambio iónico para obtener el
HADN a partir del ADN-Na.
Se debe considerar el aporte a las respectivas áreas del perfil, por parte
de los grupos hidroxilo presentes en las bases guanidina y tiamina, que se
comportan como ácidos débiles (pKa= 9,9 y 9,4, respectivamente), y de los
nitrógenos protonables presentes en adenina y citosina, centros básicos muy
débiles (pKa= 4,2 y 3,51) (Acharya S, 2006).
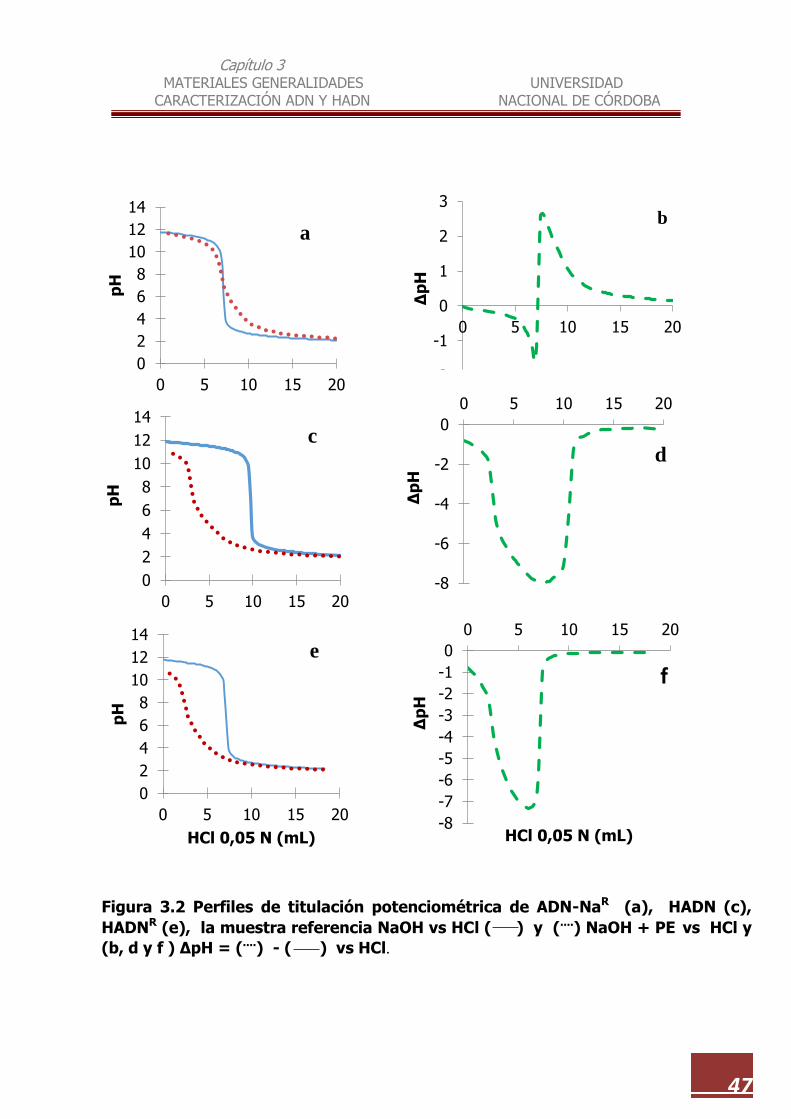
Capítulo 3 MATERIALES GENERALIDADES UNIVERSIDAD CARACTERIZACIÓN ADN Y HADN NACIONAL DE CÓRDOBA
47
-8
-7
-6
-5
-4
-3
-2
-1
0
0 5 10 15 20
Δp
H
HCl 0,05 N (mL)
f
-2
-1
0
1
2
3
0 5 10 15 20
Δp
H
b
-8
-6
-4
-2
0
0 5 10 15 20Δ
pH
0
2
4
6
8
10
12
14
0 5 10 15 20
pH
a
0
2
4
6
8
10
12
14
0 5 10 15 20
pH
0
2
4
6
8
10
12
14
0 5 10 15 20
pH
HCl 0,05 N (mL)
e
c
d
Figura 3.2 Perfiles de titulación potenciométrica de ADN-NaR (a), HADN (c),
HADNR (e), la muestra referencia NaOH vs HCl ( ) y (....) NaOH + PE vs HCl y
(b, d y f ) ΔpH = (....) - ( ) vs HCl.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
48
3.4.3. Determinación potenciométrica de Sodio
Considerando que todos los grupos fosfato se encuentran como sal
sódica (-O-Na+OPO-O), los equivalentes por gramo de polímero fueron
determinados mediante la cuantificación potenciométrica de sodio. El ADN-Na
presenta 3,71 ± 0,01 meq y el ADN-NaR contiene 3,51 ± 0,01 meq (-O-
Na+OPO-O) por gramo de material sólido (tabla 3.3). Los datos obtenidos
están cercanos al límite inferior del intervalo de peso molecular informado para
una unidad de nucleótido (288,2 – 328,2 para las bases de menor y mayor peso
molecular).
Tabla 3.3 Determinación de los mequivalentes de grupos fosfato por
gramo de material, a través de la determinación de sodio.
Ecuación de la
recta
Error Valor
X´ [M] mg/mL PE Eq.g
NaCl Y= 54,9 X+101,2 ±1,5
ADN-Na Y= 53,0 X+ 89,7 ±1,4 -2,9 1,3 x 10-3 0,35 269,2 (3,71 ± 0,01) x 10-3
ADN-NaR Y= 56,3 X+96,5 ±1,6 -2,8 1,4 x 10-3 0,41 284,9 (3,51 ± 0,01) x 10-3
Figura 3.3 Curvas de calibración del NaCl y ADN-Na para la
determinación de Sodio.
y = 54,9 x + 101,2
R² = 0,998
y = 53,0 x + 89,7 R² = 0,998
-140
-120
-100
-80
-60
-40
-20
0
20
40
60
-5 -4 -3 -2 -1 0
mV
Lg[M]
NaCl
DNA.Na C

Capítulo 3 MATERIALES GENERALIDADES UNIVERSIDAD CARACTERIZACIÓN ADN Y HADN NACIONAL DE CÓRDOBA
49
3.4.4. Caracterización de la estructura secundaria por
Dicroísmo Circular (DC)
Todas las dispersiones acuosas en agua Milli-Q de la figura 3.4, fueron
tomadas bajo las mismas condiciones de concentración y pH ≈ 7,0 Es
importante destacar que los resultados obtenidos con esta técnica sobre la
estructura secundaria son de tipo cualitativo.
Las dispersiones de los polímeros presentaron un perfil de DC
correspondiente a una estructura secundaria de doble hélice tipo-B.
Comportamiento característico de la estructura nativa, dónde las bases se van
uniendo complementariamente girando hacia la derecha, en entornos de alta
humedad relativa.
En la figura 3.4 se observan tres señales corrientemente adjudicadas a la
configuración B, un efecto Cotton positivo en 275 nm, un efecto Cotton
negativo en 240 nm y un punto de cruce con el eje en 258 nm, que coincide
con la longitud de onda de máxima absorción del ADN. Finalmente, en la región
de longitud de onda corta entre 180 y 200 nm aparece una señal positiva.
En la figura 3.5 se puede evidenciar el efecto de la concentración del PE
sobre la intensidad de las señales, estas determinaciones fueron necesarias a
fines de seleccionar la concentración óptima del PE, que permitiera definir con
mayor claridad los efectos Cotton positivo y negativo descriptos en el párrafo
anterior.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
50
Figura 3.4. Perfil de DC de ADN-NaR, DNA-Na y HADNR en agua Milli-Q,
pH ≈7,0. La concentración del PE en las dispersiones acuosas es de
0,05
mg/mL.
Figura 3.5. Perfil de DC de DNA-Na en agua Milli-Q, pH ≈7,0. La
concentración del PE en las dispersiones acuosas varía en el intervalo
de 0,005 a 0,049 mg/mL.
nm
180 200 220 240 260 280 300 320 340 360
-2
-1
0
1
2
3
ADN-Na
ADN-NaR
HADNR
nm
180 200 220 240 260 280 300 320 340 360
-2
-1
0
1
2
3 0.005 mg/mL
0.018 mg/mL
0.024 mg/mL
0.037 mg/mL
0.049 mg/mL
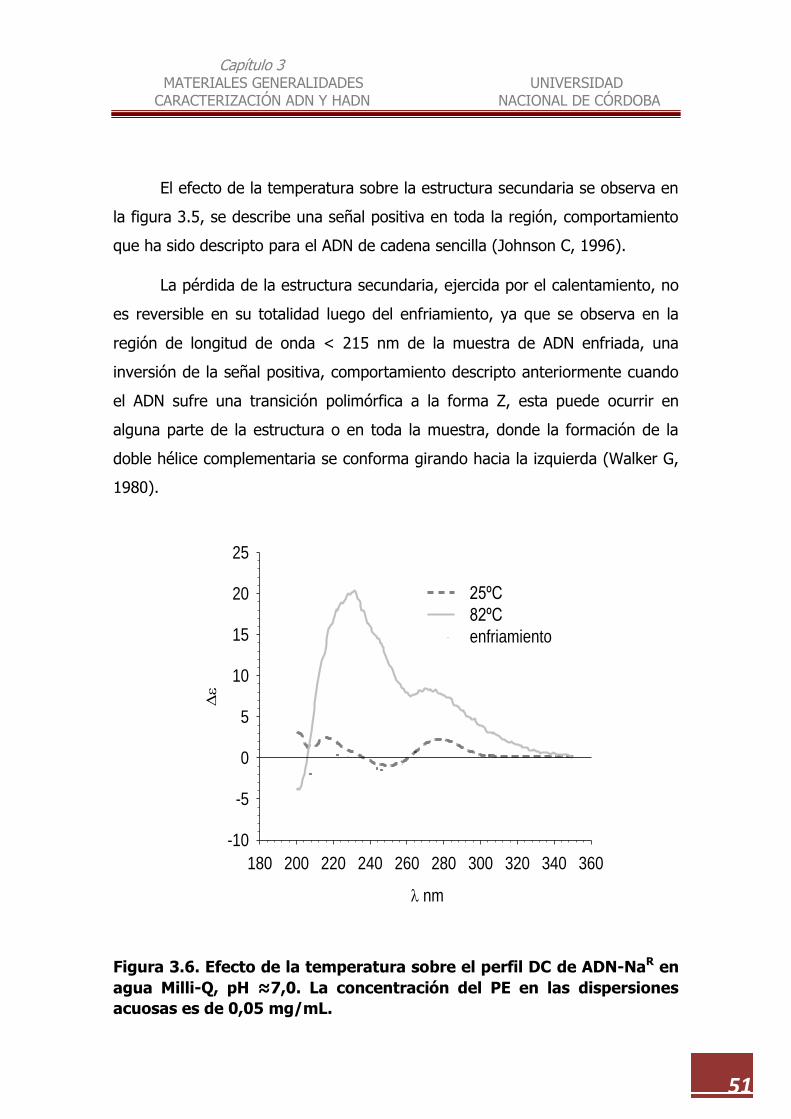
Capítulo 3 MATERIALES GENERALIDADES UNIVERSIDAD CARACTERIZACIÓN ADN Y HADN NACIONAL DE CÓRDOBA
51
El efecto de la temperatura sobre la estructura secundaria se observa en
la figura 3.5, se describe una señal positiva en toda la región, comportamiento
que ha sido descripto para el ADN de cadena sencilla (Johnson C, 1996).
La pérdida de la estructura secundaria, ejercida por el calentamiento, no
es reversible en su totalidad luego del enfriamiento, ya que se observa en la
región de longitud de onda < 215 nm de la muestra de ADN enfriada, una
inversión de la señal positiva, comportamiento descripto anteriormente cuando
el ADN sufre una transición polimórfica a la forma Z, esta puede ocurrir en
alguna parte de la estructura o en toda la muestra, donde la formación de la
doble hélice complementaria se conforma girando hacia la izquierda (Walker G,
1980).
Figura 3.6. Efecto de la temperatura sobre el perfil DC de ADN-NaR en
agua Milli-Q, pH ≈7,0. La concentración del PE en las dispersiones
acuosas es de 0,05 mg/mL.
nm
180 200 220 240 260 280 300 320 340 360
-10
-5
0
5
10
15
20
25
25ºC
82ºC
enfriamiento

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
52
3.4.5. Caracterización de tamaño molecular por
electroforesis
La figura 3.7 muestra el resultado de la electroforesis en gel de agar de
los PE. En la parte superior de la fotografía se ubica el polímero de mayor peso
molecular, en este caso la muestra 4 correspondiente al ADN-NAR exhibe una
banda entre 2000 a 8000 pares de bases (pb). En las zonas inferiores se ubican
los polímeros de menor peso molecular, el ADN-Na, el HADN y el HADNR
exhibieron un tamaño menor a 100 pb.
Figura 3.7 Electroforesis en gel de agarosa de ADN-Na y HADN, corridas en gel de agarosa al 0,75% y teñidas con SYBR Green I.
Línea 1. HADN; Línea 2. ADN-Na; Línea 3. HADNR; Línea 4. ADN-Na
R
1 2 1 2 3 4

Capítulo 3 MATERIALES GENERALIDADES UNIVERSIDAD CARACTERIZACIÓN ADN Y HADN NACIONAL DE CÓRDOBA
53
3.4.6. Ensayo de permeación de los polímeros
La figura 3.8 muestra el resultado de la permeación de los polímeros a
través de la membrana, que será utilizada posteriormente en los ensayos de
determinación de constantes de afinidad y cinética de liberación. Para la
implementación de esas técnicas es requerimiento que el polímero no atraviese
la membrana. Como se observa en la figura, la forma ácida de ADN, tanto de la
referencia como el obtenido por resina, presentan migración al compartimento
receptor de componentes de muy bajo peso molecular. Aunque el peso de
estos polímeros, determinado por electroforesis, es similar al ADN-Na, es
posible que la forma ácida tenga un porcentaje de pequeños fragmentos no
caracterizados. El comportamiento observado sobre la pureza de estas
muestras, determinó que no se utilizarán en la preparación de complejos con
los F seleccionados.
Figura 3.8 Porcentaje de permeación de polímero a través de membrana de celulosa (12,000 Da), usando como medio receptor
agua.
0
10
20
30
40
50
60
70
80
90
0 30 60 90 120 150
% p
erm
ea
ció
n d
e p
oli
me
ro
Tiempo (min)
ADN-NaHADNHADNR

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
54
3.5. Conclusiones
Se obtuvo información y conocimiento acerca de la pureza y de las
características fisicoquímicas de mayor relevancia de las dispersiones ADN-Na y
ADN-NaR y sus respectivas formas ácidas.
La batería de ensayos propuesta logró la caracterización necesaria para
avanzar en el estudio de la interacción ADN-F.
Se determinaron los grupos ionizables fosfato por gramo del ADN
Se determinó la estructura secundaría del ADN y el efecto parcialmente
irreversible de la temperatura.
Se verificó que el ADN obtenido de proveedor nacional cumple con los
criterios de calidad y pureza, al ser comparado con ADN de referencia de
calidad certificada.
La diferencia entre el peso molecular de ADN-Na y ADN-NaR, produce
dispersiones de diferente aspecto y viscosidad, lo que lleva a utilizarlas
en diferente porcentaje P/V en los complejos.
Se evidenció que la forma ácida del ADN no es conveniente para la
elaboración de complejos con F.
La frecuencia de reanálisis establecida permitió detectar detrimento de la
calidad de la materia prima y tomar los correctivos pertinentes.

Capítulo 3 MATERIALES GENERALIDADES UNIVERSIDAD CARACTERIZACIÓN ADN Y HADN NACIONAL DE CÓRDOBA
55
PARTE II
3.1. Introducción
El criterio utilizado para seleccionar un grupo de F modelo consideró, por
una parte, utilizar F que hayan sido estudiados previamente como contraiones
de PE ácidos. Por otra parte, se consideró incorporar F con diferente basicidad y
que exhibieran diferentes propiedades hidrofílicas-lipofílicas.
Adicionalmente, se estimó que ambas especies, F y FH+, tuvieran el
mismo espectro de absorción.
3.2. Set de F Modelo
Todos los F seleccionados como modelo tienen grupos básicos en su
estructura constituidos por aminas alifáticas (Figura 3.7). La tabla 3.1 contiene
las principales propiedades físicas y químicas del conjunto de F.
Estos F, como se mencionó antes, han sido utilizados con otros PE como
el Carbomer, el Ácido Algínico, la Carboximetilcelulosa, el Ácido Hialurónico, el
Eudragit®S y Eudragit®L, lo que permite comparar los resultados.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
56
Atenolol Lidocaína
Timolol Propranolol
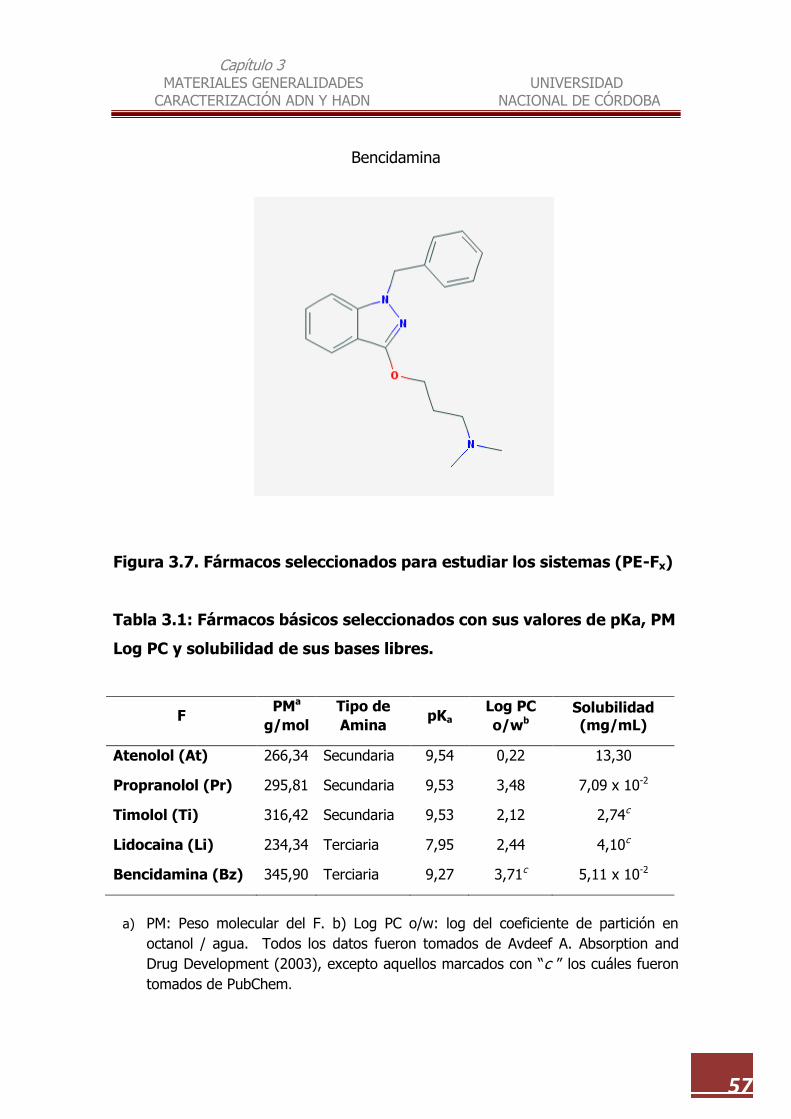
Capítulo 3 MATERIALES GENERALIDADES UNIVERSIDAD CARACTERIZACIÓN ADN Y HADN NACIONAL DE CÓRDOBA
57
Bencidamina
Figura 3.7. Fármacos seleccionados para estudiar los sistemas (PE-Fx)
Tabla 3.1: Fármacos básicos seleccionados con sus valores de pKa, PM
Log PC y solubilidad de sus bases libres.
F PMa
g/mol Tipo de
Amina pKa
Log PC
o/wb Solubilidad (mg/mL)
Atenolol (At) 266,34 Secundaria 9,54 0,22 13,30
Propranolol (Pr) 295,81 Secundaria 9,53 3,48 7,09 x 10-2
Timolol (Ti) 316,42 Secundaria 9,53 2,12 2,74c
Lidocaina (Li) 234,34 Terciaria 7,95 2,44 4,10c
Bencidamina (Bz) 345,90 Terciaria 9,27 3,71c 5,11 x 10-2
a) PM: Peso molecular del F. b) Log PC o/w: log del coeficiente de partición en
octanol / agua. Todos los datos fueron tomados de Avdeef A. Absorption and
Drug Development (2003), excepto aquellos marcados con “c ” los cuáles fueron
tomados de PubChem.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
58
3.3. Calidad y origen de los materiales utilizados
Los siguientes materiales fueron utilizados en el desarrollo de esta tesis
doctoral:
ADN de esperma de salmón (DNA-NaR) de Sigma Chemical Co., St Louis,
EE.UU. y ADN de esperma de salmón (ADN-Na) de Parafarm®, Bs. As,
Argentina. Atenolol y los clorhidratos de lidocaína, propranolol y bencidamina y
la sal de maleato de timolol, todos de calidad farmacéutica, Parafarm®, Bs. As,
Argentina. Se utilizaron Sigma SYBR Green I (SIAL), agarosa BioReagent, para
biología molecular, 1KB DNA Ladder y PCR 100 bp Low Ladder. Se preparó una
solución salina tamponada con fosfato 10 mM (pH 6,8 PBS), de acuerdo con la
USP 34-NF 29, 2011. Todos los demás reactivos químicos fueron de calidad
analítica y se utilizó agua Milli-Q en los experimentos.

En este capítulo se describen la caracterización de las propiedades
físicas de las dispersiones ADN-F. Así como un estudio preliminar de la
obtención de estos complejos en estado sólido y su posterior
redispersión.
Caracterización de
los Sistemas ADN-F
en dispersión
Capítulo 4


UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
61
4.1. Introducción
El objetivo de este capítulo es describir los resultados obtenidos en la
preparación de los diferentes complejos y la posterior caracterización de
algunas propiedades físicas de las dispersiones ADN-F, vinculadas a la
estabilidad de estos sistemas.
Con este propósito se realizaron diversas determinaciones dirigidas a
caracterizar tales propiedades, entre ellas determinaciones de pH,
conductividad, potencial electrocinético y comportamiento electroforético.
Además, se caracterizó la capacidad del ADN para formar pares iónicos
[R- FH+] con un set de F modelo. El grado de afinidad se determinó midiendo la
proporción de F unido al PE y las proporciones de especies libres bajo la forma
F y FH+ en situaciones de equilibrio utilizando diálisis.
De manera complementaria, dada la complejidad del ADN, se realizó un
estudio preliminar y exploratorio de la obtención de complejos en estado sólido
por liofilización, y su caracterización en cuanto a la pérdida de cristalinidad de
los F al interaccionar con el PE, utilizando la difracción de rayos x.
Este estudio, que aporta al entendimiento a escala molecular del sistema
PE-F, está vinculado con los resultados que se discutirán en capítulos
posteriores, sobre la cinética de liberación de los F y el efecto de la interacción
sobre la estructura secundaria del AN.

Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
62
En lo que respecta al ADN, los grupos fosfato presentes en los
nucleótidos, poseen un hidrógeno ácido (pKa 1,5 Acharya S, 2006) que se
disocia en un amplio intervalo de valores de pH de interés fisiológico y le
confieren las propiedades de los PE aniónicos. Considerando entonces, que los
equilibrios iónicos constituirían la principal interacción entre los grupos fosfato
del ADN y las especies protonadas FH+, la ecuación 1 puede ser reemplazada
por:
R-Na +FH+A- R- + FH++ Na+ + A- [R- FH+] + Na+ + A-
Ec.6
Donde R- representa un grupo fosfato ionizado de la sal sódica del ADN
y A- representa el anión de las sales de los F.
4.2. Materiales y Metodologías
Para desarrollar estas determinaciones se utilizaron las dispersiones de
ADN-Na y ADN-NaR y soluciones de F descriptos en el capítulo 3. Se utilizó
agua Milli-Q para la realización de los ensayos.
La metodología para la preparación de complejos ADN-Fx (X= 25, 42 y
85 moles %), tanto de la dispersión al 0,60 % P/V del ADN-Na, como al 0,12%
P/V del ADN-NaR y el ajuste de pH de los sistemas, y su posterior obtención en
estado sólido, se describe en el numeral 2.2.9 del capítulo 2. Se preparó el
complejo ADN-At(85)-NaCl(z), conforme al numeral 2.2.11.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
63
Los estudios de pH, conductividad, potencial ζ, electroforesis y
finalmente, los patrones de difracción de Rx fueron obtenidos tal y como se
describió en los numerales 2.2.2., 2.2.5. - 2.2.7. y 2.2.14. del capítulo 2.
Finalmente, los estudios de afinidad se llevaron a cabo utilizando bolsas
de diálisis de la manera como se describió en el numeral 2.2.10. del capítulo 2.
4.3. Resultados y Discusión
En general, la reacción de las dispersiones acuosas de ADN-Na al 0,60 %
P/V con proporciones crecientes de los F básicos modelo [atenolol (At),
lidocaína (Li), propranolol (Pr), timolol (Ti) y bencidamina (Bz)] generaron
dispersiones homogéneas ligeramente viscosas, transparentes sin la presencia
de sedimentos, excepto cuando reaccionaron con una alta proporción de F de
mayor lipofilicidad como el Pr y Bz, como se detalla en la tabla 4.1. Los
sistemas con presencia de precipitado no fueron utilizados para las posteriores
determinaciones.
Resultados similares fueron obtenidos con ADN-NaR al 0,12% P/V.
Aunque en este caso se usó una concentración más baja de PE debido a la
viscosidad producida por el alto peso molecular de la macromolécula. Sin
embargo, en ambos casos, un incremento de la concentración de los complejos,
también produce un incremento en la viscosidad de las dispersiones generando
hidrogeles traslucidos.
Por inspección visual se determinó que los sistemas son físicamente
estables durante el almacenamiento a 2-8°C durante 20 días.
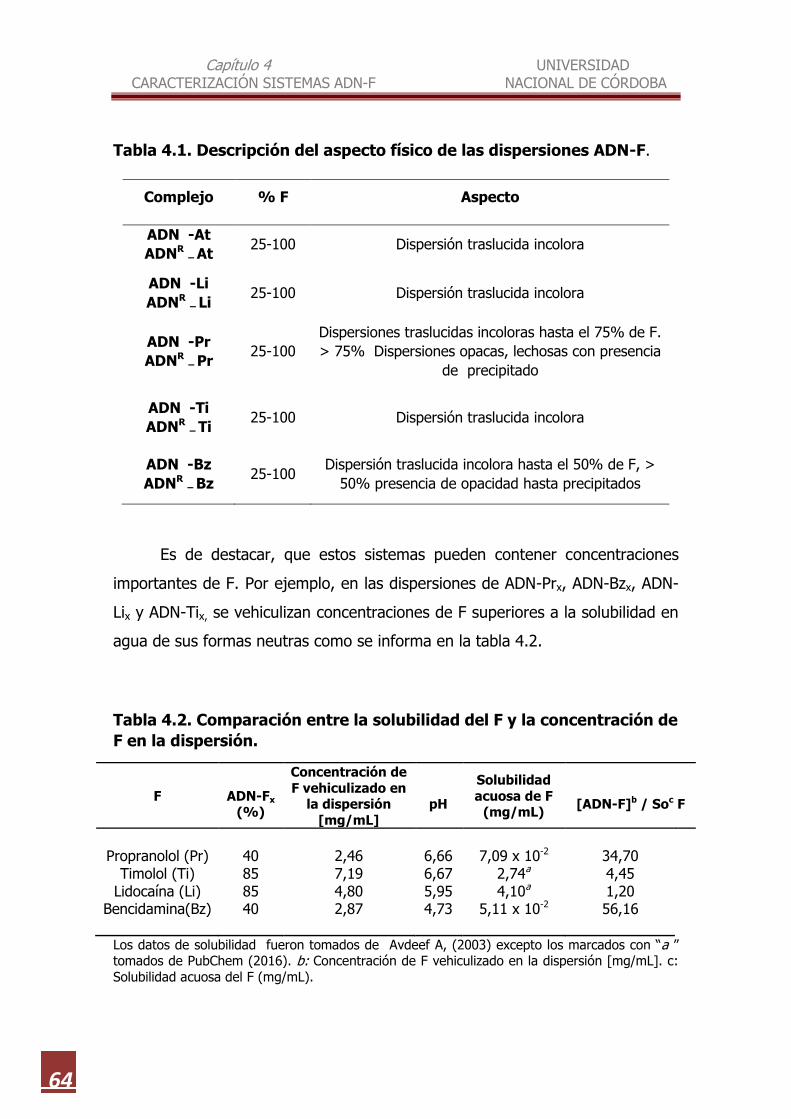
Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
64
Tabla 4.1. Descripción del aspecto físico de las dispersiones ADN-F.
Complejo % F Aspecto
ADN -At
ADNR – At 25-100 Dispersión traslucida incolora
ADN -Li
ADNR – Li 25-100 Dispersión traslucida incolora
ADN -Pr
ADNR – Pr 25-100
Dispersiones traslucidas incoloras hasta el 75% de F.
> 75% Dispersiones opacas, lechosas con presencia
de precipitado
ADN -Ti
ADNR – Ti 25-100 Dispersión traslucida incolora
ADN -Bz
ADNR – Bz 25-100
Dispersión traslucida incolora hasta el 50% de F, >
50% presencia de opacidad hasta precipitados
Es de destacar, que estos sistemas pueden contener concentraciones
importantes de F. Por ejemplo, en las dispersiones de ADN-Prx, ADN-Bzx, ADN-
Lix y ADN-Tix, se vehiculizan concentraciones de F superiores a la solubilidad en
agua de sus formas neutras como se informa en la tabla 4.2.
Tabla 4.2. Comparación entre la solubilidad del F y la concentración de
F en la dispersión.
F
ADN-Fx
(%)
Concentración de
F vehiculizado en la dispersión
[mg/mL]
pH
Solubilidad
acuosa de F (mg/mL)
[ADN-F]b / Soc F
Propranolol (Pr) 40 2,46 6,66 7,09 x 10-2 34,70
Timolol (Ti) 85 7,19 6,67 2,74a 4,45 Lidocaína (Li) 85 4,80 5,95 4,10a 1,20
Bencidamina(Bz) 40 2,87 4,73 5,11 x 10-2 56,16
Los datos de solubilidad fueron tomados de Avdeef A, (2003) excepto los marcados con “a ” tomados de PubChem (2016). b: Concentración de F vehiculizado en la dispersión [mg/mL]. c:
Solubilidad acuosa del F (mg/mL).

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
65
4.3.1. pH de las dispersiones ADN-F
El pH de las dispersiones al 0,60% P/V del ADN-Na y al 0,12% P/V del
ADN-NaR es de 7,2. La progresiva incorporación de los F en forma de sales
(clorhidrato y maleato) produce una disminución del pH como se informa en la
tabla 4.3 (a) y (b). El At que se incorpora como base libre genera valores de pH
entre 10,0 y 11,0 por lo que el sistema se lleva a neutralidad mediante el
agregado de HCl 1M.
Tabla 4.3. Valores de pH ± 0,05 de dispersiones ADN-F (a), ADNR-F
(b). F en proporciones crecientes expresadas como moles %
a) Proporción de F
%
ADN-F
Pr Me Li
25 7,17 7,06 7,01
50 7,03 6,47 6,75
75 6,97 6,24 6,62
100 6,89 6,07 6,58
b) Proporción de F
%
ADNR-F
Pr Be Li Ti
50 6,66 6,67 6,59 5,41
100 6,33 6,21 6,19 4,93
Los valores de pH de las dispersiones son directamente dependientes de
la basicidad (pKa) y porcentaje de F incorporado y de su contraión, así como
también de la concentración de polímero disperso en cada caso. El Ti está
salificado con el ácido dicarboxílico maleico cuya solución acuosa exhibe un
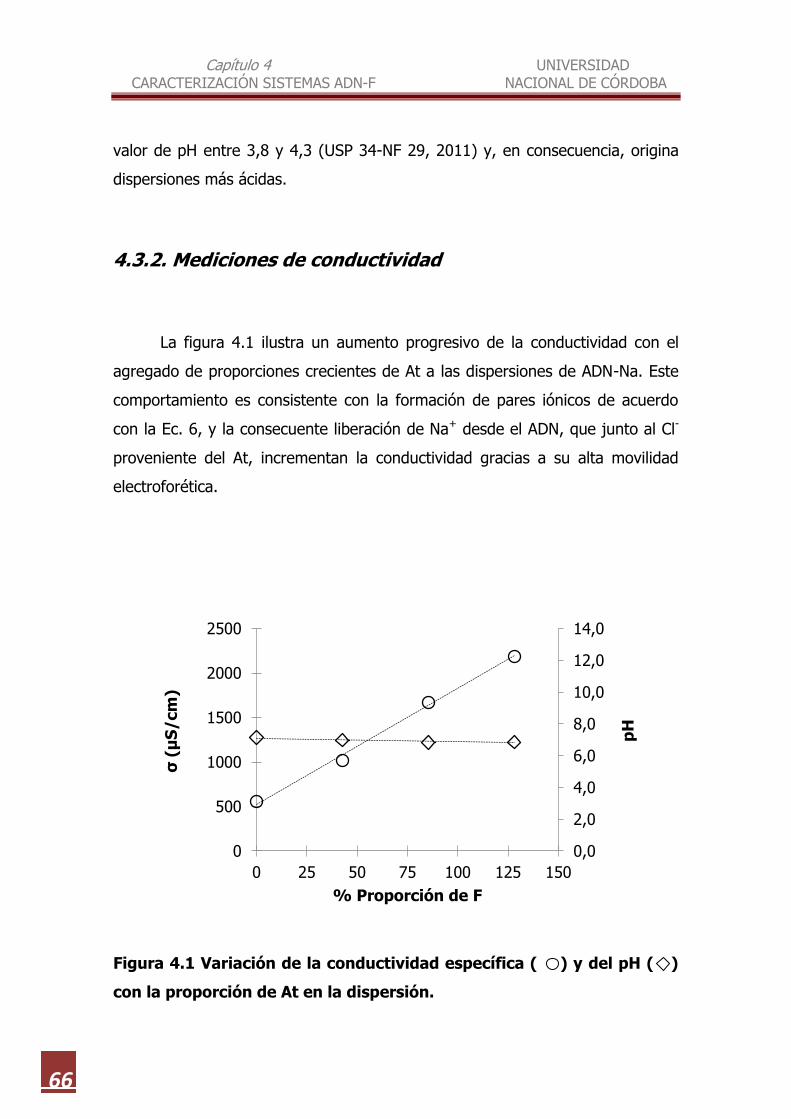
Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
66
valor de pH entre 3,8 y 4,3 (USP 34-NF 29, 2011) y, en consecuencia, origina
dispersiones más ácidas.
4.3.2. Mediciones de conductividad
La figura 4.1 ilustra un aumento progresivo de la conductividad con el
agregado de proporciones crecientes de At a las dispersiones de ADN-Na. Este
comportamiento es consistente con la formación de pares iónicos de acuerdo
con la Ec. 6, y la consecuente liberación de Na+ desde el ADN, que junto al Cl-
proveniente del At, incrementan la conductividad gracias a su alta movilidad
electroforética.
Figura 4.1 Variación de la conductividad específica ( ) y del pH ( )
con la proporción de At en la dispersión.
0,0
2,0
4,0
6,0
8,0
10,0
12,0
14,0
0
500
1000
1500
2000
2500
0 25 50 75 100 125 150
σ (
µS
/cm
)
% Proporción de F
pH
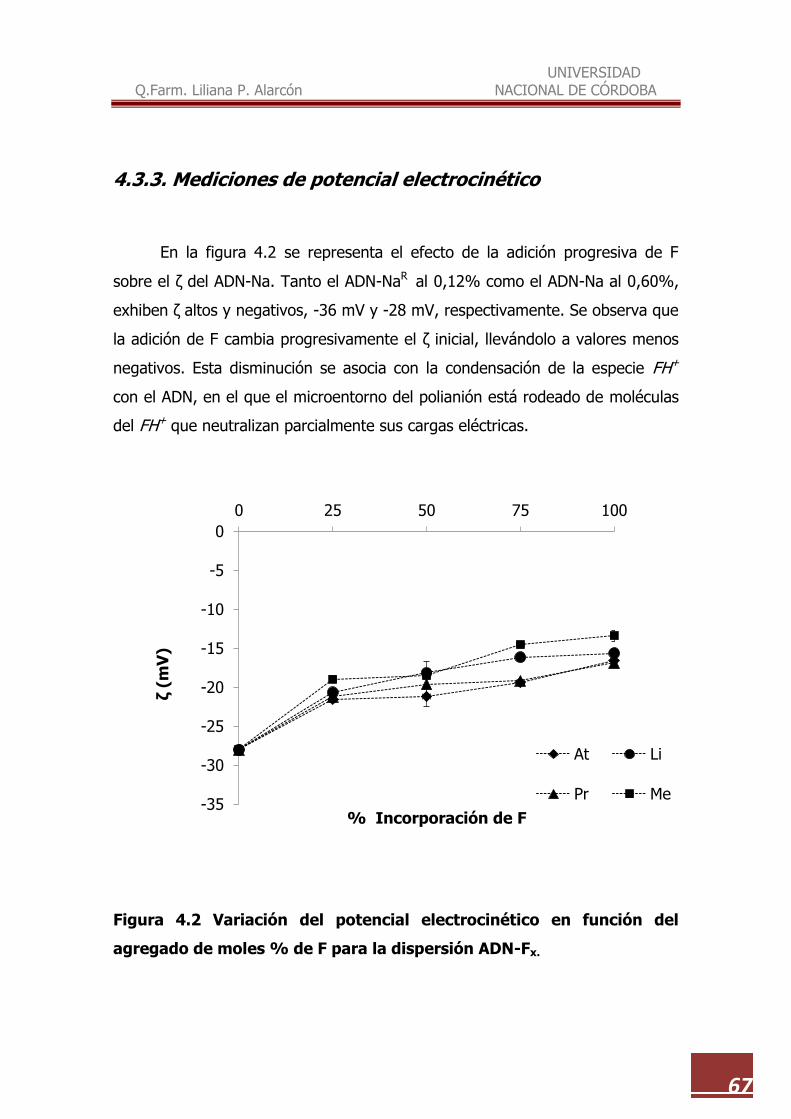
UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
67
4.3.3. Mediciones de potencial electrocinético
En la figura 4.2 se representa el efecto de la adición progresiva de F
sobre el ζ del ADN-Na. Tanto el ADN-NaR al 0,12% como el ADN-Na al 0,60%,
exhiben ζ altos y negativos, -36 mV y -28 mV, respectivamente. Se observa que
la adición de F cambia progresivamente el ζ inicial, llevándolo a valores menos
negativos. Esta disminución se asocia con la condensación de la especie FH+
con el ADN, en el que el microentorno del polianión está rodeado de moléculas
del FH+ que neutralizan parcialmente sus cargas eléctricas.
Figura 4.2 Variación del potencial electrocinético en función del
agregado de moles % de F para la dispersión ADN-Fx.
-35
-30
-25
-20
-15
-10
-5
0
0 25 50 75 100
ζ (m
V)
% Incorporación de F
At Li
Pr Me

Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
68
4.3.4. Mediciones de electroforesis
Para este ensayo se adicionaron al ADN-NaR proporciones de F
equivalentes al 100% y 150% de los grupos fosfato a fin de mantener una alta
proporción de condensación contraiónica.
Se observa en la Figura 4.3 la electroforesis en gel de agarosa de ADN-
NaR y los complejos de ADNR–At(100 y 150). Las líneas 2 y 3, representan el ADN
cuando interacciona con el At, la movilidad del PE se ve disminuida y la
intensidad de la banda no se modifica al incrementar el agregado de F.
Figura 4.3 Electroforesis de
gel en agarosa de complejos
ADNR-F 100 y 150 en gel de
agarosa 0,75%, revelados
con Syber Green I. Línea 1,
ADNR; Línea 2 ADNR-At (100);
Línea 3 ADNR-At (150).
1 2 1 2 3 6 7 8 911

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
69
4.3.5. Distribución de especies en las dispersiones ADN-F
Con el propósito de caracterizar los equilibrios involucrados en la
interacción ADN-F y el grado de condensación iónica de los F con los PE, se
determinó la distribución de las especies en el equilibrio (F, FH+ y [R- FH+]),
utilizando la técnica de diálisis. La metodología utilizada se puso a punto
previamente (Battistini F, 2014), en respuesta a las limitantes presentadas por
la técnica de partición con solventes orgánicos de la especie neutra del F que
interacciona con el ADN. Entre ellas la imposibilidad de analizar F de baja
lipofilicidad.
Esquema 4.1 Representación de las especies y equilibrios establecidos en los sistemas ADN-F en condiciones de diálisis.
La técnica de determinación de especies en el equilibrio mediante diálisis de las
dispersiones PE-F es un sistema agua/agua, en este caso el efecto de
separación se consigue con el uso de una membrana semipermeable, que
impide la difusión del ADN desde el compartimiento donor por su tamaño

Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
70
molecular, superior al tamaño del poro de la membrana como se muestra en la
figura 3.7. Los equilibrios que se establecen en este sistema están
representados en el esquema 4.1
4.3.5.1. Procesamiento de datos para la determinación de
Especies en el Equilibrio y de la constante de afinidad (Kpi)
Una vez alcanzado el equilibrio se determinaron las especies que
migraron al compartimento r, mediante:
a) La determinación molar del F por espectrofotometría UV. Dado que el
grupo básico es alifático en todos los F analizados, ambas especies F
y FH+ son analíticamente indistinguibles por lo que se determina FT =
FH+r + Fr
b) La determinación de [H+] en ambos compartimentos
c) Dado que F es la única especie capaz de difundir libremente entre los
compartimentos, se considera que su concentración en el equilibrio es
la misma en el compartimento d y r.
Es importante destacar que las mediciones espectrofotométricas se
realizan solamente en el compartimento r, en el que todas las especies están en
solución y no hay macromoléculas que puedan dispersar la radiación UV.
Sobre estas bases, conociendo la [F ] y la [H+] en el compartimento d ,
es posible determinar [FH+] a través de las ecuaciones descriptas a
continuación, y por defecto la fracción [FH+ ] que está formando parte del par
iónico. De acuerdo como lo describe la ecuación 6, en el equilibrio se cuenta
con la presencia de iones Na+ y A-, provenientes de la sal del PE y del F,
respectivamente. Estos iones por su tamaño molecular difunden libremente por
los compartimentos d y r.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
71
Dado que, la masa de Fr en mg fue restada de la masa inicial F0 para
obtener la masa del compartimento donor Fd, (F0 – Fr = Fd ), de la relación
([ F ]r /[ F ]d ) dada por la diálisis, se puede calcular la distribución de especies
de acuerdo a las siguientes ecuaciones:
[F]𝑟
[F]𝑑=
[𝐹𝐻+]𝑟 + [𝐹]𝑟
[𝐹]𝑑 + [𝐹𝐻+]𝑑 + [R− 𝐹𝐻+]𝑑
Ec.7
𝐾𝑎 =[𝐹][𝐻+]
[𝐹𝐻+]
Ec.8
La constante de afinidad de la condensación iónica (Kpi) fue calculada
siguiendo las siguientes ecuaciones (Ardusso M, 2010):
𝐾𝑝𝑖 =[R− 𝐹𝐻+] [𝐻+]
[RH] 𝐾𝑎 [𝐹𝐻+]
Ec.9
El término [RH] fue calculado de la ecuación 10.
[RH] = [RH]𝑡 - [R− 𝐹𝐻+] - [R− ]
= [RH]𝑡 - [R− 𝐹𝐻+] - [ 𝐹𝐻+] Ec.10

Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
72
Dadas las condiciones experimentales dónde la sumatoria de las especies
negativas [R− ] más [𝑂𝐻−] y [𝐶𝑙−] es igual a la sumatoria de las especies
positivas [𝐹𝐻+] más [𝐻+] y [𝑁𝑎+], y considerando que [ 𝐶𝑙−] =
[𝑁𝑎+], [R− ] ≫ [𝑂𝐻−] y [𝐹𝐻+] ≫ [𝐻+], entonces [R− ] se aproxima a [𝐹𝐻+].
De esta manera, la técnica conserva los principios fisicoquímicos de la
extracción por solventes, adicional de superar las limitantes anteriormente
mencionadas, abriendo el abanico para el análisis de una gran variedad de F
con diferentes propiedades fisicoquímicas, cuenta con la principal ventaja de
prescindir del uso de solventes orgánicos. Se obtienen resultados reproducibles
y confiables
4.3.5.2. Determinaciones analíticas en condiciones de equilibrio
La evaluación de la concentración de F en el compartimento r en función
del tiempo, permitió definir el tiempo en que el sistema alcanza el equilibrio. El
cual se define como la obtención de los mismos valores de concentración de F
en dos lecturas consecutivas, en un período no menor a 2 horas. De este modo
se estableció que en 24 horas el sistema alcanza el equilibrio.
El tiempo definido para alcanzar el equilibrio fue determinado por el
seguimiento de la diálisis durante 48 horas a 25°C del sistema ADN-Pr50. Como
puede observarse en la figura 4.4 a), para la relación de volúmenes 1/40 entre
los compartimentos d y r. En la figura 4.4 b) para la relación de volúmenes 1/10
entre compartimento d y r, en el intervalo de tiempo trascurrido entre las 24 y
36 horas se mantiene constante la concentración de F en el compartimento r ,
al igual que la distribución de especies. Conforme a los resultados, se define 24
horas como el tiempo en el cuál se alcanza el equilibrio para los sistemas ADN-
Fx.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
73
Figura 4.4 Determinación del tiempo de diálisis necesario para alcanzar el
equilibrio en los sistemas ADN-F, utilizando como modelo el complejo ADN-
Pr43 a pH≈ 7,0. Relación de volumen entre el compartimento d y r : a) 1/40
b) 1/10
En la tabla 4.4 se presenta la proporción inicial de F en el sistema ADN-Fx
(antes del equilibrio), la fracción molar del [F] en el equilibrio expresada como
porcentaje de grupos fosfato neutralizados, la distribución de las especies en el
equilibrio y la constante de afinidad (Kpi) para una serie de experimentos, en el
que los datos obtenidos se procesaron de acuerdo con las ecuaciones, que se
describieron previamente.
a
b
0
20
40
60
80
100
12 24 36
Dis
trib
uci
ón d
e E
speci
es
(% )
tiempo Horas
[ ADN- FH+]
[ F ]
[ FH+]
0
20
40
60
80
100
12 24
Dis
trib
uci
ón d
e E
speci
es
(%)
tiempo Horas
[ FH+]
[ F ]
[ ADN- FH+]

Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
74
Tabla 4.4. Distribución de especies en el equilibrio y log Kpi obtenido por diálisis de complejos ADN-Fx a 25 °C.
1: Relación de volumen entre el compartimento donor (d ) y receptor ( r ). 2: F cargado al ADN en las dispersiones (PE-Fx) colocadas en d antes de la extracción por
diálisis, expresada en mg/mL. La concentración en P/V del ADN-Na y ADN-NaR es 0,60% y
0,12%, respectivamente. 3: Porcentajes (moles %) de F acomplejado con el PE remanente en d luego de la diálisis 4: Valores de pH en r y d después de la diálisis 5: Distribución de especies expresadas como % total de F remanente en d después de la diálisis 6: Logaritmo de la constante de afinidad (Kpi) de la formación del par iónico
Vd/Vr
1. Complejo
F2 Concentración
[mg/mL]
Fx3
[mol
%]
pH4 Distribución de Especies5 (%) Log
Kpi6 r d [R-FH+] FH+ F
1/40
ADN-AtX
2,20 9,38 6,37 6,21 83,98 16,01 1,0 x 10-2 5,86
3,33 18,34 6,77 6,31 83,28 16,70 1,0 x 10-2 5,79
4,49 29,30 6,27 6,44 96,70 3,30 2,8 x 10-3 6,49
6,60 28,99 6,17 6,19 91,49 8,51 4,0 x 10-3 6,30
ADN-PrX
1,25 11,25 6,75 5,66 76,06 23,94 3,5 x 10-3 6,06
2,45 20,71 6,71 5,73 74,47 25,53 4,3 x 10-3 6,00
3,72 24,51 6,64 5,72 69,78 30,21 5,1 x 10-3 5,92
4,90 26,19 6,49 5,94 82,65 17,35 4,8 x 10-3 6,03
ADN-LiX
2,24 9,42 6,17 5,91 85,01 14,75 0,230 4,51
3,39 12,65 6,06 5,91 86,44 13,34 0,225 4,58
4,48 20,59 6,14 5,81 84,94 14,87 0,191 4,66
ADN -At85 4,48 28,79 6,27 6,44 96,70 3,30 2,8 x 10-3 6,49
ADN -At85-NaCl25 4,48 22,67 6,26 6,18 91,19 8,81 4,1 x 10-3 6,26
ADN -At85-NaCl50 4,48 14,08 6,19 6,29 89,98 10,01 6,3 x 10-3 6,04
ADN -At85-NaCl75 4,48 5,42 6,19 6,05 78,07 21,92 6,8 x 10-3 5,84
ADN -At85-NaCl100 4,48 13,32 6,20 5,99 49,35 50,63 1,8 x 10-2 5,37
1/10
ADN -AtX
2,20 36,53 6,36 6,43 98,67 1,33 1,1 x 10-3 6,95
3,33 59,44 6,42 6,41 99,27 0,73 6,0 x 10-4 7,42
4,49 75,37 6,37 6,44 98,80 1,20 9,6 x 10-4 7,40
5,52 92,46 6,27 7,00 97,22 2,77 8,7 x 10-3 6,98
ADNR-AtX 1,29 40,77 6,24 6,71 93,19 6,80 1,1 x 10-2 6,66
1,57 41,95 6,34 6,63 86,60 13,39 1,8 x 10-2 6,43
ADNR- PrX
0,71 30,77 6,25 6,28 88,45 11,54 6,8 x 10-3 6,63
1,45 43,60 6,55 6,25 63,24 36,74 2,1 x 10-2 6,09
2,10 55,13 6,38 6,26 68,25 31,74 1,8 x 10-2 6,28
ADNR- LiX 1,44 30,74 6,30 6,13 53,04 45,73 1,211 4,40
2,13 50,93 6,12 6,07 70,29 29,03 0,699 4,93

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
75
Como se puede observar en esta tabla, una alta fracción de F que
permanece en el compartimiento d cuando se alcanza el equilibrio, está
involucrada en la condensación iónica con el ADN.
Los F modelo exhiben altas afinidades con el ADN, expresadas como Kpi,
que son del orden de 106.
Los experimentos en una relación de volumen de 1/40 evidencian que At
y Pr exhiben Kpi similares, y a su vez mayores que la presentada por Li, que es
un F de menor basicidad (con un pKa inferior). Este comportamiento también
se observó anteriormente con otros PE ácidos (Ardusso M, 2010).
Adicionalmente, para los experimentos con una relación de volumen de
1/10, cuando el sistema ADN-Atx se carga con una mayor proporción de At, la
Kpi aumenta de manera proporcional con la carga. Finalmente, tanto los
resultados de la tabla 4.4 y la figura 4.5 muestran que el ADN-NaR de alto peso
molecular exhibió un comportamiento similar al observado con el ADN-Na.
2,0
3,0
4,0
5,0
6,0
7,0
8,0
5 10 15 20 25 30
Log K
pi
Proporción de F (%) en el equilibrio
Prop
Lid
2,0
3,0
4,0
5,0
6,0
7,0
8,0
25 35 45 55 65
Log K
pi
Proporción de F (%) en el equilibrio
Prop
Lid
b a
Figura 4.5 Efecto de la proporcionalidad de F modelo que neutraliza ADN-
Na sobre la Kpi. Comparación del comportamiento entre las dos fuentes
de ADN-Na ensayadas. a) ADN-Na 0,60 % P/V b) ADN-NaR 0,12 % P/V
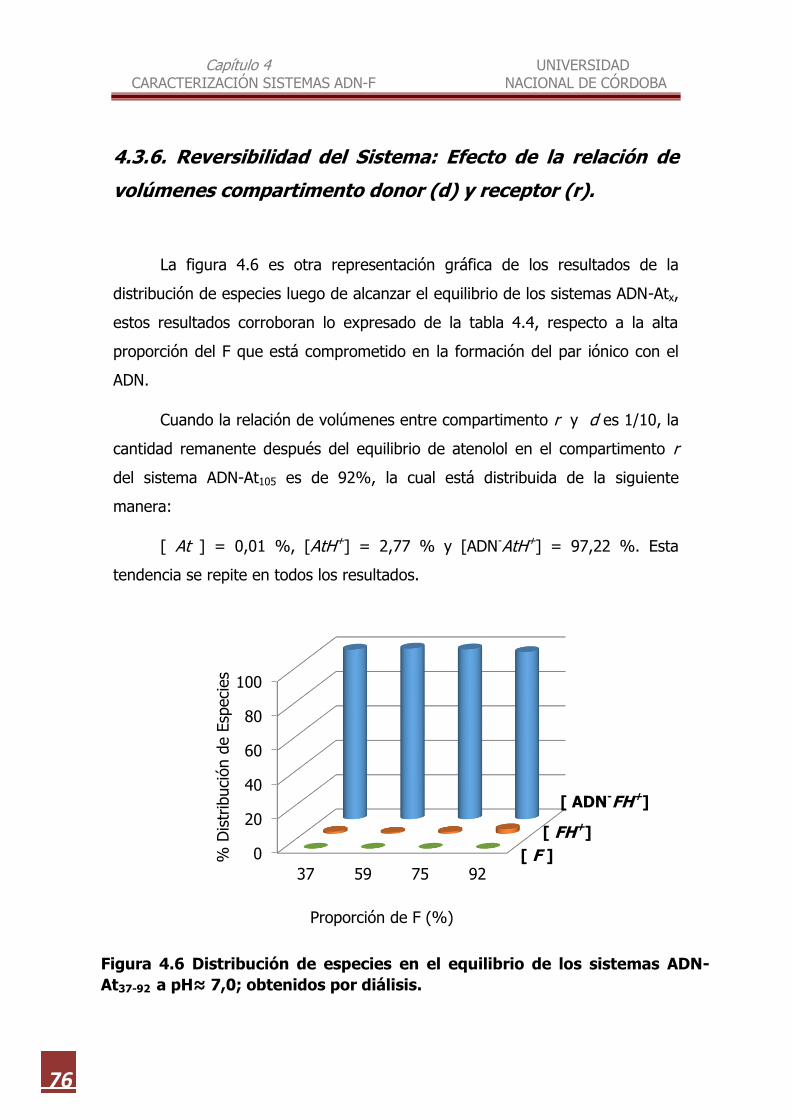
Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
76
4.3.6. Reversibilidad del Sistema: Efecto de la relación de
volúmenes compartimento donor (d) y receptor (r).
La figura 4.6 es otra representación gráfica de los resultados de la
distribución de especies luego de alcanzar el equilibrio de los sistemas ADN-Atx,
estos resultados corroboran lo expresado de la tabla 4.4, respecto a la alta
proporción del F que está comprometido en la formación del par iónico con el
ADN.
Cuando la relación de volúmenes entre compartimento r y d es 1/10, la
cantidad remanente después del equilibrio de atenolol en el compartimento r
del sistema ADN-At105 es de 92%, la cual está distribuida de la siguiente
manera:
[ At ] = 0,01 %, [AtH+] = 2,77 % y [ADN-AtH+] = 97,22 %. Esta
tendencia se repite en todos los resultados.
0
20
40
60
80
100
37 59 75 92
% D
istr
ibuci
ón d
e E
speci
es
Proporción de F (%)
Figura 4.6 Distribución de especies en el equilibrio de los sistemas ADN-
At37-92 a pH≈ 7,0; obtenidos por diálisis.
[ ADN-FH+]
[ FH+]
[ F ]
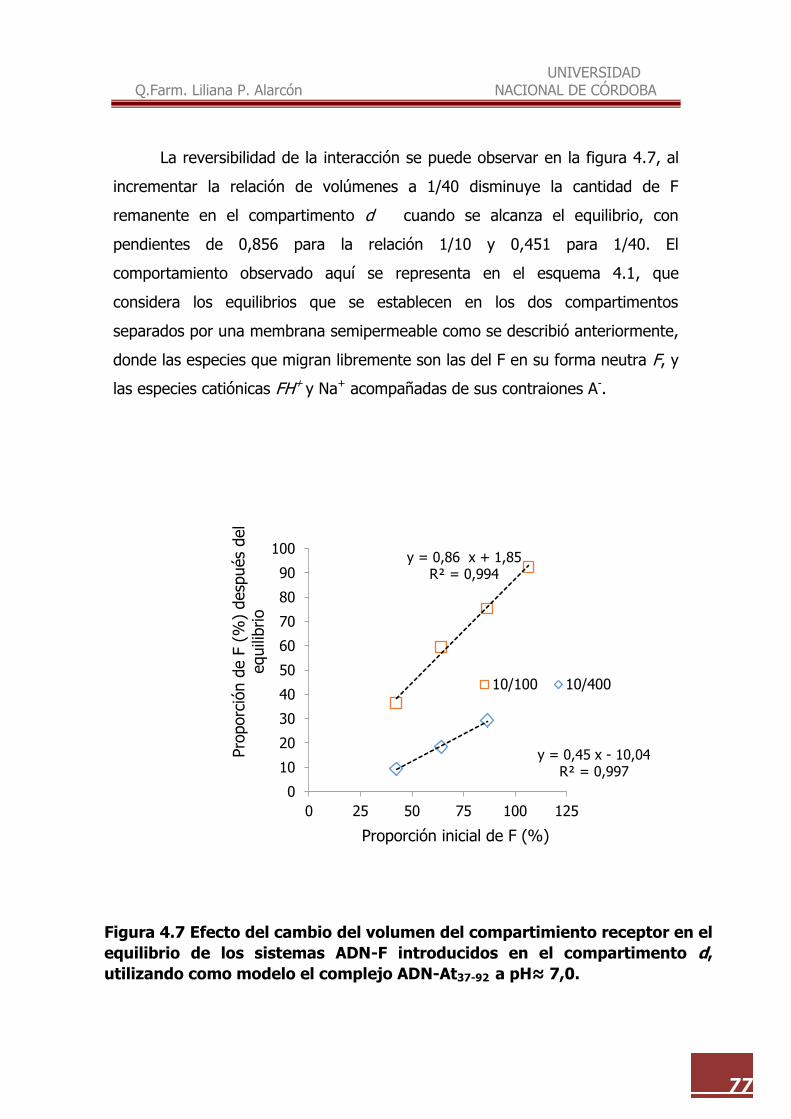
UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
77
La reversibilidad de la interacción se puede observar en la figura 4.7, al
incrementar la relación de volúmenes a 1/40 disminuye la cantidad de F
remanente en el compartimento d cuando se alcanza el equilibrio, con
pendientes de 0,856 para la relación 1/10 y 0,451 para 1/40. El
comportamiento observado aquí se representa en el esquema 4.1, que
considera los equilibrios que se establecen en los dos compartimentos
separados por una membrana semipermeable como se describió anteriormente,
donde las especies que migran libremente son las del F en su forma neutra F, y
las especies catiónicas FH+ y Na+ acompañadas de sus contraiones A-.
b
Figura 4.7 Efecto del cambio del volumen del compartimiento receptor en el
equilibrio de los sistemas ADN-F introducidos en el compartimento d,
utilizando como modelo el complejo ADN-At37-92 a pH≈ 7,0.
y = 0,86 x + 1,85 R² = 0,994
y = 0,45 x - 10,04 R² = 0,997
0
10
20
30
40
50
60
70
80
90
100
0 25 50 75 100 125
Pro
porc
ión d
e F
(%
) desp
ués
del
equili
brio
Proporción inicial de F (%)
10/100 10/400

Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
78
A medida que F migra al compartimento r, FH+ se disocia en agua y
como consecuencia de esta disminución de [FH+], esta especie se regenera a
partir de ADN-AtH+ actuando este par iónico como un reservorio de F, hasta
que, una vez saturado el compartimento r se alcanza el equilibrio químico.
Explicando el mecanismo de mayor migración de F al aumentar el volumen del
compartimento r.
4.3.7. Reversibilidad del Sistema: Efecto de la adición de
otras especies sobre los equilibrios.
El modelo de equilibrios propuesto, permite suponer que el agregado de
iones al sistema debería generar desplazamientos de especies como
consecuencia de intercambios iónicos y reagrupamiento de cargas. No obstante,
como se describió anteriormente en la ecuación 6, el sistema originalmente
cuenta con la presencia del ion Na+ proveniente del ADN-Na y del ion Cl-
proveniente de las sales de los F modelo, especies que también alcanzan un
equilibrio entre el compartimento d y r. bajo estas condiciones se determinaron
constantes de afinidad del orden de 106, evidenciando la fuerte atracción
electrostática entre el ADN y F.
R-Na +FH+A- R- + FH++ Na+ + A- [R- FH+] + Na+ + A-
Ec.6
Para un sistema sin interferencia original de iones Na+ y Cl- , el reagrupamiento
de cargas se describe en la ecuación 11 y 12.
[R- FH+] + NaCl- R- Na+ + FH++ Cl- Ec.11
FH+ F + H+ Ec.12

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
79
De este modo, para evaluar la reversibilidad de la interacción PE-F, se
usó como estrategia la adición de proporciones crecientes de NaCl al sistema
ADN-F que ya contiene Na+ y Cl-. Al aumentar la concentración del ion Na+ se
genera un desplazamiento de una cierta proporción de FH+. Sin embargo, como
puede observarse en la figura 4.8 y en la tabla 4.4, el efecto sobre la Kpi es
limitado, lo que evidencia la mayor afinidad de FH+ sobre Na+.
Figura 4.8 Efecto de la adición de concentraciones crecientes de
contraiones inorgánicos (NaCl), sobre la distribución de especies de los
sistemas ADN-F, utilizando como modelo el complejo ADN-At85. Los valores
de pH corresponden al compartimento d en el equilibrio.
5,8
5,9
6,0
6,1
6,2
6,3
6,4
6,5
3,0
4,0
5,0
6,0
7,0
8,0
9,0
0 25 50 75 100
Lo
g K
pi
Proporción de NaCl adicionado (%)
NaCl
pH
pH

Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
80
Patron de difracción de Rayos X
2
0 20 40 60 80 100
ADN-Na
ADN-Li50
Li
4.3.8. Difracción de rayos X de los complejos ADN-F
Se obtuvieron por liofilización complejos sólidos de color blanco, que se
almacenaron en envases de vidrio ámbar y protegidos de la humedad.
El ADN, como la mayoría de los polímeros, presenta un patrón de
difracción nulo, correspondiente a un sólido amorfo, que se caracteriza por
presentar una banda dispersa de baja intensidad a lo largo de todo el espectro
de 2θ / θ. En cambio, los F cristalinos producen un espectro de rayos X
característico, presentando picos intensos a determinados valores de 2θ / θ.
(a)
Figura 4.9 (a) Difracción de rayos X de ADN-Na, de los complejos iónicos ADN-Li y Li como F modelo seleccionado, en el intervalo 2θ/θ.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
81
Patron de difracción de Rayos X
2
0 20 40 60 80 100
ADN-Na
ADN-Pr50
Pr
(b)
Figura 4.9 (b) Difracción de rayos X de ADN-Na, de los complejos iónicos ADN-Pr y Pr como F modelo seleccionado, en el intervalo 2θ/θ.
En las figuras 4.9 (a) y (b) se muestran los patrones de rayos X
característicos para los F seleccionados como modelo de molécula contraión (Li
y Pr respectivamente), del polímero y de los complejos iónicos. Como puede
observarse, el ADN-Na, presenta un diagrama de difracción nulo, una banda
dispersa de baja intensidad semejante a otros PE estudiados. En contraste los F
presentan un difractograma de sólido cristalino.
En los difractogramas de los complejos obtenidos por liofilización,
desaparecen las señales del F cristalino completamente al 40% de
neutralización, asemejándose al de sólido amorfo.

Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
82
Respecto al comportamiento de los complejos PE-F, obtenidos por
interacción iónica entre un F cristalino con un PE. Estos dan origen a un
material amorfo, con ausencia de F libre, que se adjudica a la alta condensación
iónica del F con el PE. (Jiménez-Kairuz A, 2004; Ramírez M, 2006; Quinteros D,
2008; Esteban S, 2009).
Finalmente, se procedió a la redispersión de los complejos sólidos en la
misma proporción de agua Milli-Q inicial, observándose las mismas
características visuales de las dispersiones originales, sin presencia de
precipitados o turbidez. La determinación de la recuperación de las demás
propiedades fisicoquímicas se procederá en etapas posteriores a ser evaluadas.
4.4. Conclusiones
La reacción entre dispersiones acuosas de ADN-Na o ADN-NaR y
diferentes proporciones de las sales de los F utilizados, generan
complejos estables que permiten vehiculizar concentraciones de F
superiores a la solubilidad de sus formas no protonadas.
Los resultados que se reportan en este capítulo son consistentes con la
interpretación basada en la ecuación 6.
La incorporación al ADN de proporciones crecientes de los F genera una
progresiva disminución del pH, lo que es consistente con la alta acidez
de los grupos fosfato (pKa≈1) y la basicidad de las aminas alifáticas (pKa
= 7,95 - 9,53), genera también un aumento de conductividad debido a la
liberación de los contraiones Na+ y Cl- de alta movilidad electroforética.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
83
En todos los casos produce una progresiva disminución del alto potencial
electrocinético negativo de ADN-Na, que es consistente con la formación
de pares iónicos [R- FH+]. A su vez, disminuye la movilidad
electroforética de las macromoléculas, lo que puede interpretarse por la
disminución de las cargas netas y aumento del peso molecular.
La técnica definida para la determinación cuantitativa de la afinidad ADN-
F resultó ser idónea para el sistema estudiado, debido a que como se
informó mediante la figura 3.7, la membrana de 12,000 D es efectiva
para evitar el pasaje de la sal sódica del ADN. Además, la técnica
permite conservar la integridad del biopolímero, como no sucedería con
el uso de solventes orgánicos.
El análisis de la determinación cuantitativa de la afinidad, evidenció que
una alta proporción de los F ensayados se encuentra unida a los grupos
fosfato de los ADN en forma reversible exhibiendo constantes de afinidad
(Kpi) en el intervalo 104 – 107.
Las Kpi para los F con mayor basicidad (At, Pr) exhiben valores más altos
que los F con menor basicidad (Li).
El sistema es sensible a la adición de sales neutras que disminuyen
progresivamente el valor de las Kpi, pero pone en evidencia que la
afinidad de las especies protonadas FH+ es mucho mayor que la de los
cationes Na+.

Capítulo 4 UNIVERSIDAD
CARACTERIZACIÓN SISTEMAS ADN-F NACIONAL DE CÓRDOBA
84
Los resultados pueden interpretarse mediante la ecuación 6, lo que pone
de manifiesto que el ADN se comporta en forma análoga a otros PE
ácidos estudiados previamente. La tabla 4.5 presenta datos comparativos
de las Kpi de F con mayor (At, Pr, Me) y menor basicidad (Li), como
acomplejantes de PE lineales con grupos ácidos de diferente fuerza (ADN
pKa≈1.0, HA pKa≈2 y EL, ES pKa≈6)
Del análisis comparativo con otros PE, se observa que los valores de Kpi
para el ADN son similares al HA, estos son mayores que los obtenidos
con los Eudragit® ( EL, ES), y a su vez, EL y ES tienen Kpi similares.
Tabla 4.5. Comparativo de la distribución de especies en el equilibrio y log Kpi de complejos PE-Fx
Donde EL: Eudragit L; ES: Eudragit S; AH: Ácido Hialurónico; Me:
Metoclopramida
Complejo
Fx
[mol %] Distribución de Especies (%)
Log Kpi [R-FH+] FH+ F
EL-Li 25 86,80 12,40 0,80 3,45
50 76,24 33,31 1,45 3,23
ES-Li 25 88,60 8,60 2,80 3,02
50 82,86 13,64 3,49 3,02
EL-Me 50 49,74 50,22 0,04 4,55
ES-Me 50 68,50 31,41 0,09 4,59
AH-Li 25 56,07 42,99 0,94 4,25
50 62,23 36,99 0,77 4,36
AH-Pr 25 60,63 39,64 0,02 5,89
100 78,56 21,42 0,02 6,14
ADNR-AtX 29,30 96,70 3,30 0,003 6,49
40,77 93,19 6,80 0,011 6,66
ADNR- PrX 30,77 88,45 11,54 0,01 6,63
55,13 68,25 31,74 0,02 6,28
ADNR- LiX 30,74 53,04 45,73 1,21 4,40
50,93 70,29 29,03 0,69 4,93

En este capítulo se estudia la cinética y el mecanismo de liberación in vitro de F
modelos desde los complejos iónicos ADN-F, y los factores que pueden
contribuir a modificar la liberación, entendiendo el proceso molecular y
fisicoquímico que se da lugar, y a partir de esta base predecir el comportamiento
que los sistemas tendrán in vivo.
Cinética y
mecanismo de
liberación de los F
desde los complejos
ADN-F
Capítulo 5


UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
87
5.1. Introducción
Los estudios de la liberación de los F desde los complejos PE-F
generalmente se desarrollan en celdas bicompartimentales de tipo Franz. Esta
metodología fue introducida en la década de los 70´s (Franz T, 1978) y
posteriormente optimizada (Shah V, 1999) y fue denominada método de celdas
de difusión vertical por la Food and Drug Administration (FDA), que lo describe
como un método simple, confiable y reproducible para el monitoreo de la
liberación de principios activos desde cremas, ungüentos e hidrogeles (USP 34-
NF 29, 2011; FDA, 1997). La Federación Farmacéutica Internacional publicó
una guía de recomendaciones para la realización de ensayos in vitro para
nuevas formas farmacéuticas donde también las incluye (Brown C, 2011). En el
campo de la investigación, es el método más común referenciado en
publicaciones científicas para la evaluación de la liberación de F desde sistemas
nanoparticulados (Budhian A, 2008; Nerella A, 2014) e hidrogeles (Jiménez-
Kairuz A, 2004).
El dispositivo se representa en el esquema 5.2, compuesto por dos
compartimentos, separados por una membrana sintética semipermeable, el
compartimento donor es el receptáculo dónde se coloca el sistema a ser
analizado, mientras que el compartimento receptor contiene el medio en el que
se desea analizar el sistema. Dentro de las consideraciones para aplicar el
método se destacan, que el tamaño de poro de la membrana debe impedir el
paso de las macromoléculas como los PE y el aseguramiento de las condiciones
sink, es decir, condiciones alejadas de la saturación del F en el medio de
difusión durante todo el ensayo (USP 34-NF 29, 2011; FDA, 1997).
Para realizar una aproximación a los mecanismos involucrados en el
proceso de liberación del F desde el sistema portador se requiere analizar los
perfiles de cinética de liberación a través de modelos matemáticos.

Capítulo 5 CINÉTICA Y MECANISMO DE LIBERACIÓN UNIVERSIDAD DE LOS F DESDE LOS COMPLEJOS ADN-F NACIONAL DE CÓRDOBA
88
En la actualidad, no existe un modelo que considere todos los procesos
físicos y químicos que puedan ocurrir en los diferentes sistemas, pero se logran
interesantes correlaciones con la experimentación in vivo (Franz T, 1978; Batool
H, 2013).
El modelo más sencillo fue propuesto por T. Higuchi (Higuchi T, 1963;
Hasan E, 2003), en este modelo la fracción de F liberado es proporcional a la
raíz cuadrada del tiempo, como se describe en la ecuación 1.
Q = k √t Ec 1
Donde Q es la cantidad (o proporción) de F liberado a tiempo t, k es la
constante de velocidad de liberación. Los valores de k son expresados en
unidades mg.h-1/2 u h-1/2.
Se interpreta que, si se satisface esta correlación, el mecanismo que
gobierna la liberación del F es la difusión, como se ha descripto para numerosos
sistemas matriciales, transdermales, entre otros (Jiménez-Kairuz A, 2003; 2004)
Otro modelo, denominado Korsmeyer-Pepas (Korsmeyer R, 1983; Ritger
P, 1987), fue introducido más recientemente para el análisis de perfiles de
liberación de sistemas matriciales, utilizando la siguiente ecuación:
Mt /M∞ = k t n Ec 2
Donde Mt es la cantidad de F en el compartimento receptor en el tiempo
t; M∞, la cantidad inicial de F en el compartimento donor; Mt /M∞ es la fracción
(0,1 – 0,7) de F liberado a cada tiempo t; k es la constante de velocidad de
liberación y n es el exponente difusional cuyo valor se relaciona al mecanismo
de liberación. El modelo determina que el análisis de los datos se efectúa entre
el 10 y 70% de liberación del F.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
89
Un valor de n aproximadamente 0,5 en los sistemas matriciales se
atribuye a un mecanismo de liberación controlado por la difusión del F,
mientras que un valor de n próximo a 1, es atribuido a la erosión o relajación
de las cadenas del polímero. Los valores intermedios sugieren que ambos
procesos (difusión y erosión) regulan la velocidad de liberación (Ranga K, 1998;
Talukdar M, 1995; Lotfipour F, 2004).
Originariamente los modelos cinéticos fueron propuestos para el análisis
de sistemas matriciales, en la actualidad se ha aplicado exitosamente a
sistemas dispersos (Jiménez-Kairuz A, 2003; 2004; Ramírez M, 2006; Ardusso
M, 2010).
En el caso de sistemas compuestos por complejos PE-F, el entendimiento
del proceso cinético requiere un análisis molecular semejante al realizado en el
capítulo 4 en la determinación de las constantes de afinidad.
Se ha propuesto un modelo que contempla la difusión del F desde el
entorno del PE como se esquematiza en el esquema 5.1 para un sistema
constituido exclusivamente por un PE ácido neutralizado con el F básico
(Jiménez-Kairuz A, 2003; 2004; Battistini F, 2013):
De acuerdo con este modelo, cuando el medio receptor es agua, la
difusión del F desde el complejo la desarrolla esencialmente la especie
neutra F que puede difundir libremente al seno de la solución.
Se observó que la velocidad de difusión del F depende de la proporción
de pares iónicos presentes en el sistema. De manera que el equilibrio de
disociación de los pares iónicos en el entorno del complejo PE-F gobierna
la cinética.
Cuando se utiliza una solución salina neutra en el medio receptor (NaCl),
la difusión de los iones Cl- y Na+ al compartimento donor proporciona los
contraiones para que difunda significativamente la especie FH+
acompañando a F con el consecuente aumento de la velocidad de
liberación.

Capítulo 5 CINÉTICA Y MECANISMO DE LIBERACIÓN UNIVERSIDAD DE LOS F DESDE LOS COMPLEJOS ADN-F NACIONAL DE CÓRDOBA
90
En ambas condiciones, la disociación del par iónico es la etapa limitante en
la cinética de liberación, de esta manera, el sistema PE-F se comporta como un
reservorio de F capaz de liberarlo de una manera lenta al medio (Vilches A,
2002; Jiménez-Kairuz A, 2003; 2004; Ramírez M, 2006; Ardusso M, 2010).
Esquema 5.1 Modelo que representa la distribución de todas las
especies en equilibrio en sistema PE ácido- F básico (Jiménez-Kairuz
A, 2004)

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
91
El objetivo de este capítulo es estudiar la cinética y el mecanismo de
liberación in vitro de los F modelos desde los complejos iónicos ADN-F y los
factores que pueden contribuir a modificar la liberación, entendiendo el proceso
molecular y fisicoquímico que se da lugar, y a partir de esta base predecir el
comportamiento que los sistemas tendrán in vivo.
Es importante destacar que en todos los casos al complejo ADN-F lo
acompañan los contraiones provenientes de la macromolécula (Na+) y de la sal
del F (generalmente Cl-). En consecuencia, aun cuando el sistema de liberación
utiliza agua en el compartimento receptor, las especies con capacidad de
difundir libremente son F y FH+ acompañado del contraión Cl- u otro
correspondiente a la sal utilizada.
5.2. Materiales y Metodologías
Se utilizó el ADN-Na y ADN-NaR, se seleccionaron At, Li y Pr como F
modelos para realizar el estudio de liberación. La selección consideró abarcar
diferentes propiedades fisicoquímicas de los F, en cuanto a su hidrofilicidad y
basicidad descriptas previamente en el capítulo 3.
La metodología para la preparación de complejos ADN-Fx (X= 25, 42 y 85
moles %), tanto de la dispersión al 0,60% P/V del ADN-Na, como al 0,12% P/V
del ADN-NaR y el ajuste de pH de los sistemas, se describe en el numeral 2.2.9.
del capítulo 2.
Finalmente, los estudios de cinética de liberación in vitro de los F desde
las dispersiones de los complejos ADN-Fx, se llevaron a cabo utilizando celdas
de difusión bicompartimentales del tipo Franz (ver esquema 5.2), separadas por
una membrana semipermeable de acetato de celulosa (12,000 Da; Sigma. St
Louis.MA. USA), de la manera que se describió en el numeral 2.2.12. del
capítulo 2.

Capítulo 5 CINÉTICA Y MECANISMO DE LIBERACIÓN UNIVERSIDAD DE LOS F DESDE LOS COMPLEJOS ADN-F NACIONAL DE CÓRDOBA
92
Esquema 5.2 Celdas bicompartimentales del tipo Franz utilizada para
evaluar la cinética de liberación de los complejos ADN-F.
5.3. Resultados y discusión
5.3.1 Liberación en agua
Como se señaló en el capítulo 3, la eficacia de la membrana frente a la
difusión de la macromolécula se evaluó y controló colocando como muestra
ADN-Na en el compartimento donor y monitoreando la absorbancia en el
compartimento receptor a 260 nm, durante el ensayo no se observó migración
del PE al compartimento receptor, como se puede observar en las figuras 5.1,
5.2 y 5.3 a), dónde el perfil del ADN-Na se superpone al eje de las “x”.
Los perfiles de liberación de los F modelo desde el complejo ADN-F, en
comparación con el perfil de difusión de la solución de referencia del F (no
acomplejado) pueden observarse en las figuras 5.1, 5.2 y 5.3 a), donde se
grafica la proporción en % de F liberado en función del tiempo. La liberación
del F desde los complejos hacía el agua ocurre en forma lenta.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
93
En esas condiciones, la interacción reversible ADN-Fx genera una difusión
de la especie libre del F ( F ) hacia el compartimento receptor, acompañado de
la especie FH+ con su contraión correspondiente. La velocidad de difusión de
las especies libres anteriormente mencionadas, fue considerablemente más
lenta que la del F colocado en una concentración equivalente en el
compartimiento donor del sistema como referencia.
Esta capacidad para modular la liberación es consistente con el alto
grado de condensación contraiónica observado anteriormente. De hecho, como
se describe en la ecuación 6, tanto la especie F sin carga y la especie catiónica
FH+ junto con A- son capaces de difundir libremente y liberarse, mientras que
[R- FH+] se mantiene como un reservorio del F. En línea con estas
observaciones, las velocidades de liberación del F siguieron la correlación
clásica de Higuchi, tiempo1/2 (Visakh P, 2014) como se puede ver en las figuras
5.1, 5.2 y 5.3 b), donde se grafica la cantidad en % de F liberado en función
del tiempo1/2. Los valores de k, para las diferentes dispersiones, se informan en
la Tabla 5.1.
Se puede ver que la constante de liberación de Li desde el complejo
(4,33) es mayor que las obtenidas para los complejos de At (2,64) y Pr (1,85),
comportamiento esperado por la menor Kpi determinada para la lidocaína
Tabla 5.1 Constante de la cinética de liberación en agua: Pendiente de la ecuación de Higuchi (Ec 1), % F liberado en función de la raíz cuadrada del tiempo para F y sistemas ADN-F50.
Sistema k de
liberación ((%).h-1/2)
Δ k y-
intercepto r2
At 8,8 ± 0,2
6,1 -13,6 1,000
ADN-At 2,6 ± 0,1 -1,8 0,989 Li 7,1 ± 0,1
2,8 -10,5 0,999
ADN-Li 4,3 ± 0,2 -8,1 0,993 Pr 7,4 ± 0,3
5,6 -11,8 0,996
ADN-Pr 1,8 ± 0,1 -3,1 0,992
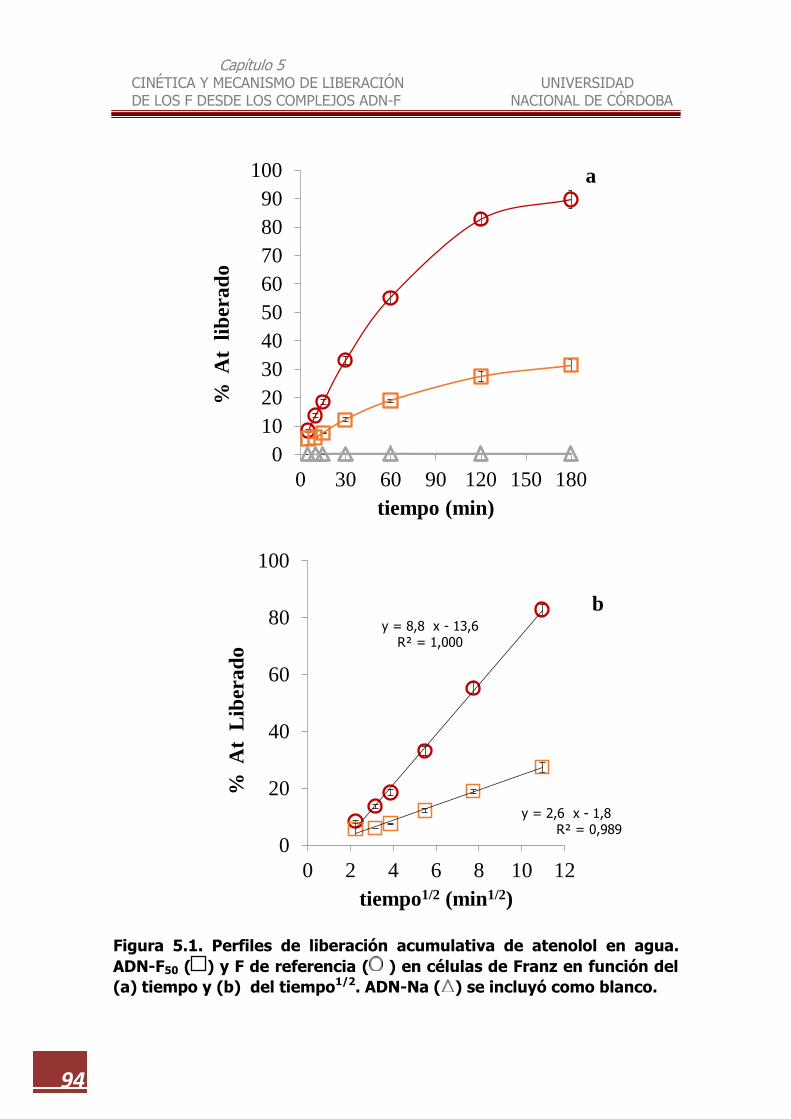
Capítulo 5 CINÉTICA Y MECANISMO DE LIBERACIÓN UNIVERSIDAD DE LOS F DESDE LOS COMPLEJOS ADN-F NACIONAL DE CÓRDOBA
94
Figura 5.1. Perfiles de liberación acumulativa de atenolol en agua.
ADN-F50 ( ) y F de referencia ( ) en células de Franz en función del
(a) tiempo y (b) del tiempo1/2. ADN-Na ( ) se incluyó como blanco.
0
10
20
30
40
50
60
70
80
90
100
0 30 60 90 120 150 180
% A
t l
iber
ad
o
tiempo (min)
a
y = 8,8 x - 13,6
R² = 1,000
y = 2,6 x - 1,8
R² = 0,989
0
20
40
60
80
100
0 2 4 6 8 10 12
% A
t L
iber
ad
o
tiempo1/2 (min1/2)
b

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
95
Figura 5.2. Perfiles de liberación acumulativa de lidocaína en agua.
ADN-F50 ( ) y F de referencia ( ) en células de Franz en función del
(a) tiempo y (b) del tiempo1/2. ADN-Na ( ) se incluyó como blanco.
0
10
20
30
40
50
60
70
80
90
100
0 30 60 90 120 150 180
%
Li
Lib
era
do
tiempo (min)
a
y = 4,3 x - 8,1
R² = 0,993
y = 7,1 x - 10,5
R² = 0,998
0
20
40
60
80
100
0 2 4 6 8 10 12
% L
i L
iber
ad
o
tiempo1/2 (min1/2)
b

Capítulo 5 CINÉTICA Y MECANISMO DE LIBERACIÓN UNIVERSIDAD DE LOS F DESDE LOS COMPLEJOS ADN-F NACIONAL DE CÓRDOBA
96
Figura 5.3 Perfiles de liberación acumulativa de propranolol en agua.
ADN-F50 ( ) y F de referencia ( ) en células de Franz en función del
(a) tiempo y (b) del tiempo1/2. ADN-Na ( ) se incluyó como blanco.
0
10
20
30
40
50
60
70
80
90
100
0 30 60 90 120
% P
r L
iber
ad
o
tiempo (min)
a
y = 1,8 x - 3,1
R² = 0,992
y = 7,4 x - 11,8
R² = 0,996
0
20
40
60
80
100
0 2 4 6 8 10 12
% P
r L
iber
ad
o
tiempo1/2 (min1/2)
b

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
97
Por otra parte, cuando se mantiene la concentración de ADN-Na
constante, y se incrementa gradualmente la proporción de F incorporado, se
observa un aumento proporcional de la velocidad de liberación del F, como se
muestra en las Figuras 5.4, 5.5 y 5.6. y en la tabla 5.2. Es decir, que los
sistemas se comportaron como portadores que liberan lentamente el F, tal
como se observa con otros sistemas PE-F similares conteniendo At, Li y Pr
(Jiménez-Kairuz A, 2003; Ardusso M, 2010).
Los resultados presentados anteriormente apoyan la hipótesis de que la
principal interacción entre el ADN y los F con grupos básicos protonables ocurre
a través de los grupos fosfato del ADN, lo que conlleva a una alta proporción de
condensación contraiónica, como se describe en la ecuación 6 y se explica en el
capítulo 4. La alta acidez de los grupos fosfato dio como resultado altas
constantes de afinidad (Kpi), incluso en la presencia de sales. A modo
comparativo, Kpi menores se han reportado para PE con grupos carboxílicos de
menor acidez (Ardusso M, 2010).
Tabla 5.2 Datos cinéticos de % F liberado en agua en función de la raíz cuadrada del tiempo para F y sistemas ADN-F50.
Complejo
Fx
[mol
%]
mg de F liberado al compartimento d
T5 T15 T30 T60 T120 T180
ADN-At
42 0,15 ± 0,06 0,198± 0,003 0,32 ± 0,02 0,49 ± 0,01 0,71 ± 0,04 0,82 ± 0,06
64 0,17 ± 0,08 0,26 ± 0,03 0,45 ± 0,04 0,75 ± 0,07 1,16 ± 0,09 1,38 ± 0,11
85 0,22 ± 0,05 0,45 ± 0,06 0,80 ± 0,08 1,3 ± 0,1 1,90 ± 0,12 2,19 ± 0,12
ADN-Li
25 0,24 ± 0,14 0,21 ± 0,04 0,28 ± 0,08 0,43 ± 0,03 0,63 ± 0,04 0,73 ± 0,07
42 0,14 ± 0,01 0,21 ± 0,01 0,31 ± 0,01 0,56 ± 0,05 0,9 ± 0,1 1,12 ± 0,15
85 0,32 ± 0,04 0,43 ± 0,07 0,79 ± 0,09 1,30 ± 0,12 1,93 ± 0,23 2,21 ± 0,21
ADN-Pr
25 0,023 ± 0,001 0,05 ± 0,01 0,08 ± 0,01 0,11 ± 0,02 0,15 ± 0,02 0,17 ± 0,02
42 0,037 ± 0,001 0,092 ± 0,002 0,17 ±0,01 0,28 ± 0,02 0,42 ± 0,04 0,52 ± 0,05
85 0,09 ± 0,01 0,22 ± 0,02 0,38 ± 0,04 0,7 ± 0,1 1,09 ± 0,16 1,36 ± 0,12
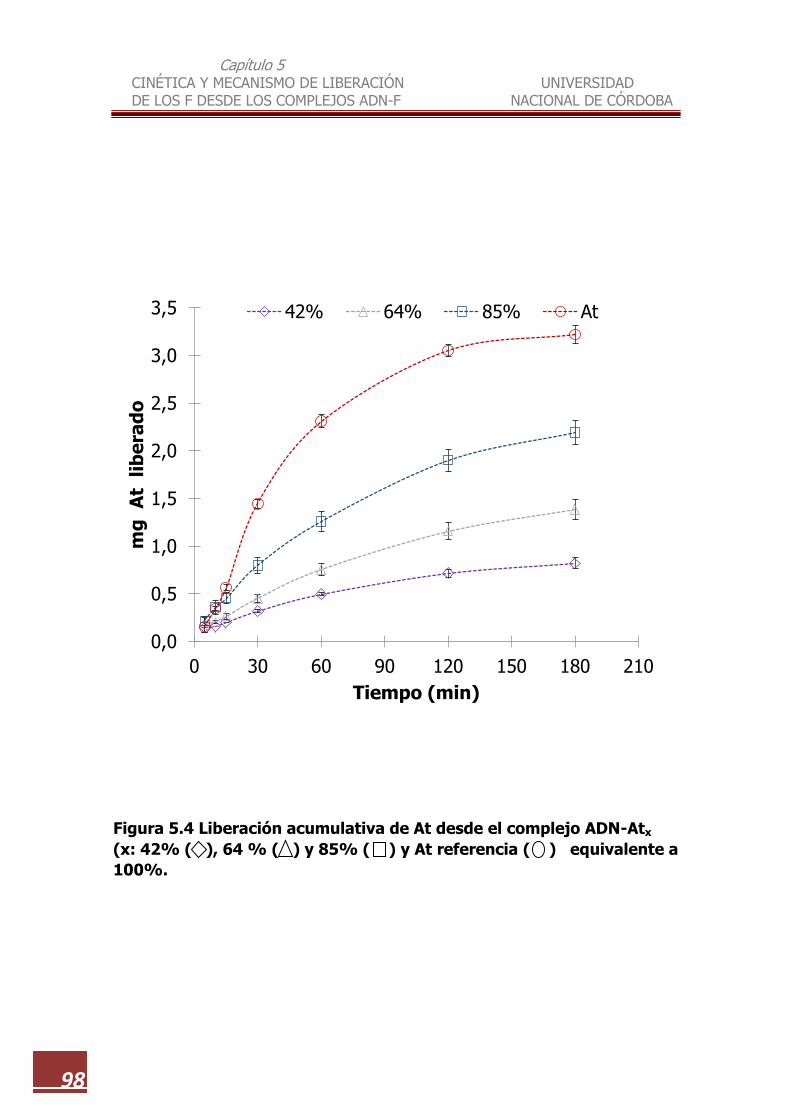
Capítulo 5 CINÉTICA Y MECANISMO DE LIBERACIÓN UNIVERSIDAD DE LOS F DESDE LOS COMPLEJOS ADN-F NACIONAL DE CÓRDOBA
98
Figura 5.4 Liberación acumulativa de At desde el complejo ADN-Atx
(x: 42% ( ), 64 % ( ) y 85% ( ) y At referencia ( ) equivalente a
100%.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
0 30 60 90 120 150 180 210
mg
A
t l
ibe
rad
o
Tiempo (min)
42% 64% 85% At
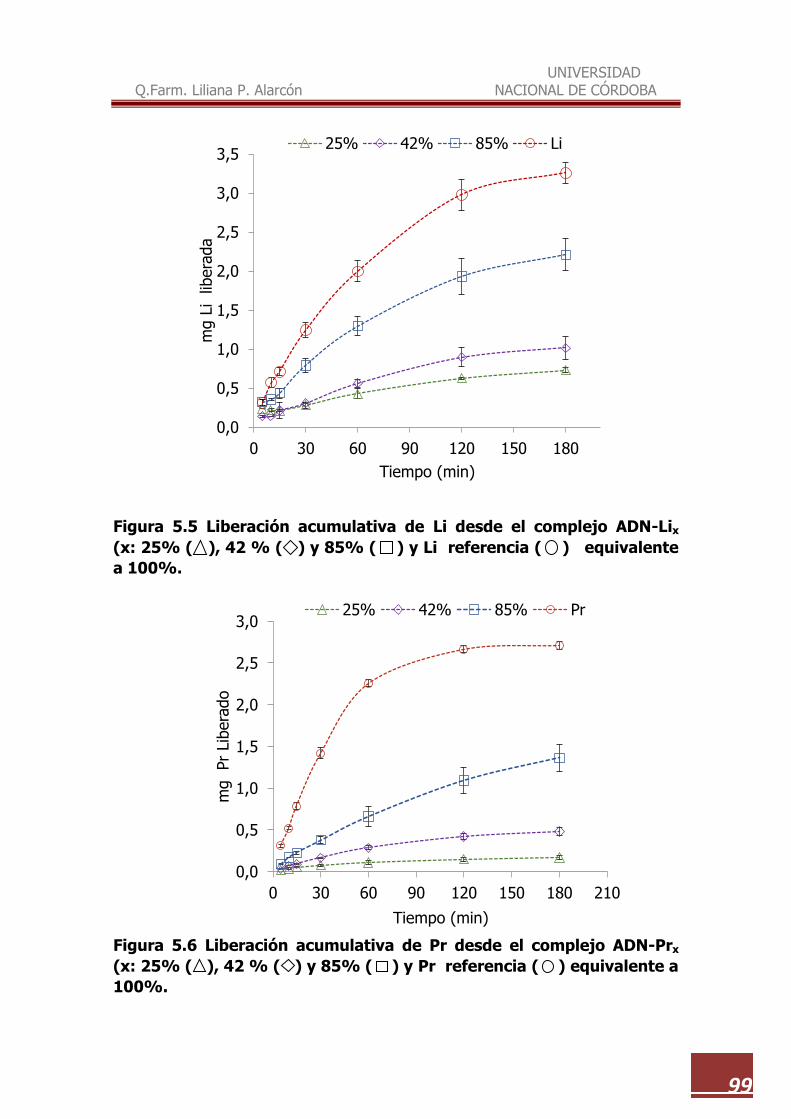
UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
99
Figura 5.5 Liberación acumulativa de Li desde el complejo ADN-Lix
(x: 25% ( ), 42 % ( ) y 85% ( ) y Li referencia ( ) equivalente
a 100%.
Figura 5.6 Liberación acumulativa de Pr desde el complejo ADN-Prx
(x: 25% ( ), 42 % ( ) y 85% ( ) y Pr referencia ( ) equivalente a
100%.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
0 30 60 90 120 150 180
mg L
i lib
era
da
Tiempo (min)
25% 42% 85% Li
0,0
0,5
1,0
1,5
2,0
2,5
3,0
0 30 60 90 120 150 180 210
mg
Pr
Lib
era
do
Tiempo (min)
25% 42% 85% Pr

Capítulo 5 CINÉTICA Y MECANISMO DE LIBERACIÓN UNIVERSIDAD DE LOS F DESDE LOS COMPLEJOS ADN-F NACIONAL DE CÓRDOBA
100
5.3. Conclusiones
La liberación de los F desde los complejos respondió en todos los casos a
una cinética interpretada como difusional.
El incremento en la proporción de F en los complejos produjo un
concomitante aumento en la velocidad de liberación.
En línea con la reversibilidad de la interacción ADN-F, la cinética de
liberación del F observada en los sistemas evaluados es consistente con
la determinada en otros PE-F modelo desarrollados previamente
(Jiménez-Kairuz A, 2002; 2003;), no se observaron diferencias
significativas en el comportamiento de la cinética de liberación de los
complejos con los diferentes F evaluados.

Efecto de la
interacción PE-F
sobre la estructura
secundaria del ADN
Capítulo 6
En este capítulo se describen los fundamentos de la técnica dicroísmo circular
y su aplicación al análisis estructural del ADN. Así como la caracterización de
la estructura secundaria del ADN-Na de Salmón y el efecto que produce la
interacción con los F sobre la estructura de la macromolécula y su
reversibilidad.


UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
103
6.1. Introducción
En los capítulos anteriores se ha determinado que la interacción ADN-F
exhibe un alto grado de condensación iónica, con constantes de afinidad del
orden de 106 y que, en concordancia a esta interacción reversible, el F es
liberado lentamente de estos complejos, respondiendo a una cinética de tipo
difusional. El comportamiento observado fue interpretado en términos de la
interacción electrostática entre los grupos fosfato del ADN y las especies
protonadas de los F modelo.
En ese orden de ideas, el objetivo de este capítulo es la caracterización
de la estructura secundaría del ADN-Na y el efecto que produce la interacción
con F sobre la estructura de la macromolécula y su reversibilidad.
A tales fines se utilizó el dicroísmo circular (DC). Las bases físicas y
químicas de esta técnica, se describen en el anexo 2.
6.1.1. Aplicaciones del DC en el análisis estructural del ADN
La señal en el perfil de DC en el ADN se debe a un efecto inducido por el
azúcar que posee un centro quiral, sobre las bases purínicas que son simétricas
y por lo tanto ópticamente inactivas. Los grupos fosfatos tienen transiciones
electrónicas a altos niveles de energía a longitudes de onda inferiores a los 170
nm, el azúcar presenta bajos niveles de intensidad de absorción electrónica
alrededor de 190 nm, longitudes donde la técnica es poco sensible; por lo
tanto se centra el análisis en la región de 300 nm donde se observa una señal
de gran intensidad debida a la transición π-π* generada por la nube electrónica
del sistema aromático de las bases purínicas, y otra de menor intensidad
debida a la transición n-π* generada por electrones no enlazantes (Johnson C,
1996; Xi Z, 2005; Zsila F, 2015).

Capítulo 6 EFECTO DE LA INTERACCIÓN PE-F SOBRE UNIVERSIDAD LA ESTRUCTURA SECUNDARIA DEL ADN NACIONAL DE CÓRDOBA
104
Figura 6.1 Espectro de DC para ARN-A dextrógiro y ADN-A, ADN-B
dextrógiro, y ADN-Z levógiro (Bloomfield V. A. et al. 2000).
Los perfiles de DC de las diferentes formas que pueden tener los AN se
muestran en la figura 6.1; de manera general, la intensidad de las señales es
baja debido a que el DC ocurre como un efecto secundario inducido por el
azúcar asimétrico. La intensidad de la señal toma valores mayores en función
del aumento de complejidad del sistema, desde el acomodamiento de las bases
formando una hélice, hasta la complementariedad entre las cadenas generando
la doble hélice o estructura secundaria helicoidal, siendo esta una estructura
asimétrica (Johnson C, 1996).

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
105
Por lo anteriormente expuesto acerca de las interacciones electrónicas
entre las bases, la técnica de DC es altamente sensible a los cambios en la
estructura secundaria helicoidal de los AN y su orientación, ya que la formación
de la doble hélice depende: del solvente en que se dispersa, el número de pb
por giro y finalmente, la inclinación y distancia de las bases con respecto al eje
central (Johnson C, 1996).
Las configuraciones que adopta el ADN tienen las siguientes
características espectrales:
Forma B: Un efecto Cotton positivo centrado cerca de 275 nm, un
efecto Cotton negativo centrado cerca de 240 nm y un punto de cruce con el
eje entre las dos bandas mencionadas alrededor de 258 nm, que coincide con
la longitud de onda de máxima absorción. En la región de longitud de onda
corta, es decir entre 180 y 200 nm, el efecto Cotton positivo cambia de manera
significativa en función del porcentaje de los enlaces G-C y de la pérdida de la
estructura secundaria, esta intensidad puede disminuir a ¼ de su señal inicial
cuando solo el 50% de las hebras se encuentran apareadas entre sí, es decir,
cuando ocurre su fusión (Tm) (Tanaka K, 1996; Kankia B, 2001; Sousa F, 2007).
Forma A: Un efecto Cotton positivo centrado alrededor de 260nm, Un
efecto Cotton negativo en 210 nm, y otro fuertemente intenso positivo en 190
nm (Johnson C, 1996).
Forma Z: Un efecto Cotton negativo en 190 nm indica que la doble
hélice fue formada hacía la izquierda. (Walker G, 1980).
6.2. Materiales y Metodologías
Se utilizaron el ADN-Na y ADN-NaR y los F descriptos descripto en el
capítulo 3. Se utilizó agua Milli-Q para la realización de los ensayos.

Capítulo 6 EFECTO DE LA INTERACCIÓN PE-F SOBRE UNIVERSIDAD LA ESTRUCTURA SECUNDARIA DEL ADN NACIONAL DE CÓRDOBA
106
La metodología para la preparación de complejos ADN-Fx (X= 25, 42 y
85 moles %), tanto de la dispersión al 0,60% P/V del ADN-Na, como al 0,12%
P/V del ADN-NaR y el ajuste de pH de los sistemas, se describe en el numeral
2.9 del capítulo 2.
Finalmente, los estudios de la estructura secundaria tanto del ADN como
de las dispersiones de los complejos ADN-Fx, se llevaron a cabo en un
espectropolarímetro Jasco 81, de la manera que se describió en los numerales
2.8 y 2.13 del capítulo 2
6.3 Resultados y Discusión
6.3.1 Efecto de la lipofilicidad de los F sobre la estructura
secundaria
Se puede observar en la Figura 6.2 que ambas muestras de ADN-Na
exhibieron espectros de DC característicos de una estructura secundaria de
doble hélice tipo-B (Johnson C, 1996; Zsila F, 2015). En la figura 6.3 se
demostró que el efecto de la temperatura conlleva a la formación de una
cadena sencilla, caracterizada por la desaparición del efecto Cotton negativo en
240 nm, la ausencia del punto de cruce con el eje cercano a 258 nm, y la
desaparición del efecto Cotton positivo en la región corta del espectro, es decir,
a longitudes < 200 nm (Walker G, 1980). El enfriamiento de la muestra
evidencia que la estructura en la mayor parte del perfil recupera su estructura
de doble hélice.
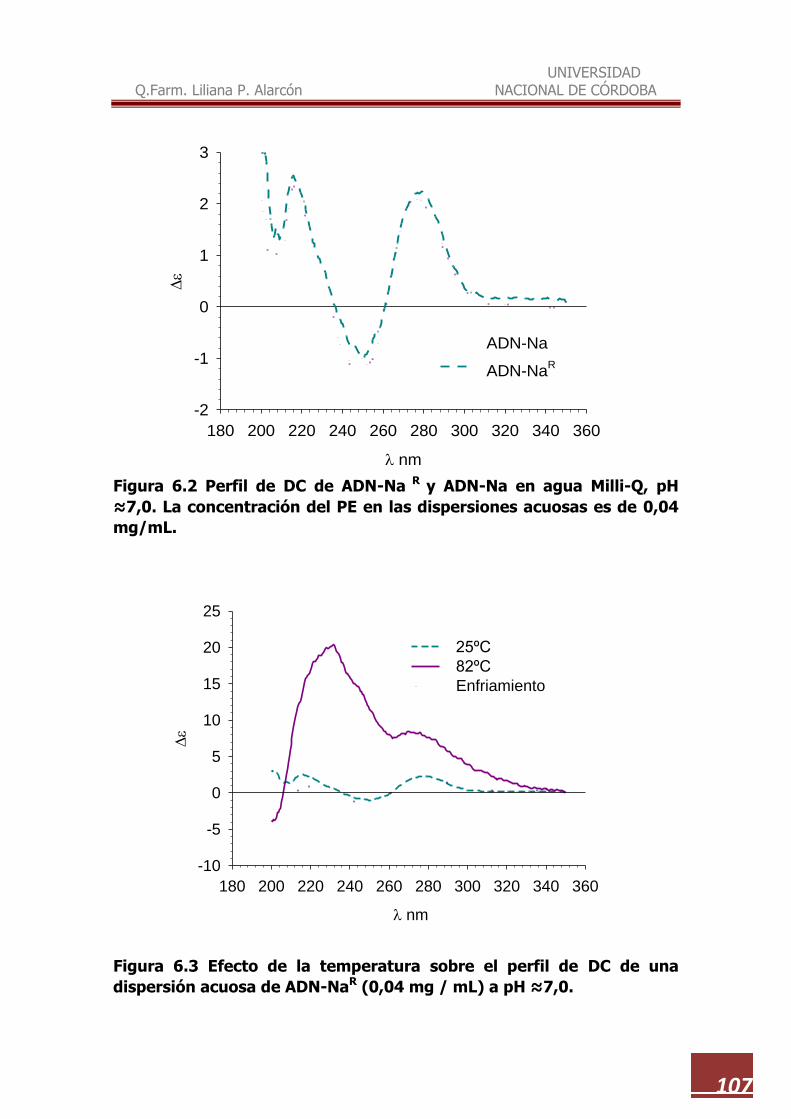
UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
107
Figura 6.2 Perfil de DC de ADN-Na R y ADN-Na en agua Milli-Q, pH
≈7,0. La concentración del PE en las dispersiones acuosas es de 0,04
mg/mL.
Figura 6.3 Efecto de la temperatura sobre el perfil de DC de una
dispersión acuosa de ADN-NaR (0,04 mg / mL) a pH ≈7,0.
nm
180 200 220 240 260 280 300 320 340 360
-2
-1
0
1
2
3
ADN-Na
ADN-NaR
nm
180 200 220 240 260 280 300 320 340 360
-10
-5
0
5
10
15
20
25
25ºC
82ºC
Enfriamiento

Capítulo 6 EFECTO DE LA INTERACCIÓN PE-F SOBRE UNIVERSIDAD LA ESTRUCTURA SECUNDARIA DEL ADN NACIONAL DE CÓRDOBA
108
Los complejos de ADN-F50 con los F modelo exhibieron diferentes perfiles
de DC. Los complejos con F de menor lipofilicidad como el atenolol (At),
lidocaína (Li) y timolol (Ti), mostraron señales características del ADN nativo
como se observa en las figuras 6.4, 6.5 y 6.6, lo que sugiere que el ADN en el
complejo mantiene la forma de doble cadena tipo-B.
Figura 6.4 Perfil de DC de ADN-Na R y ADN-At50 en agua Milli-Q, pH
≈7,0. La concentración del PE en las dispersiones acuosas es de 0,04
mg/mL.
nm
220 240 260 280 300 320
-20
-10
0
10
20
30
ADN-NaR
ADNR-At50

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
109
Figura 6.5 Perfil de DC de ADN-Na R y ADN-Li50 en agua Milli-Q, pH
≈7,0. La concentración del PE en las dispersiones acuosas es de 0,04
mg/mL.
Figura 6.6 Perfil de DC de ADN-Na R y ADN-Ti50 en agua Milli-Q, pH
≈7,0. La concentración del PE en las dispersiones acuosas es de 0,04
mg/mL.
nm
220 240 260 280 300 320
-30
-20
-10
0
10
20
30
ADN-NaR
ADNR
-Li50
nm
220 240 260 280 300 320
-30
-20
-10
0
10
20
30
ADN-NaR
ADNR
-Ti50
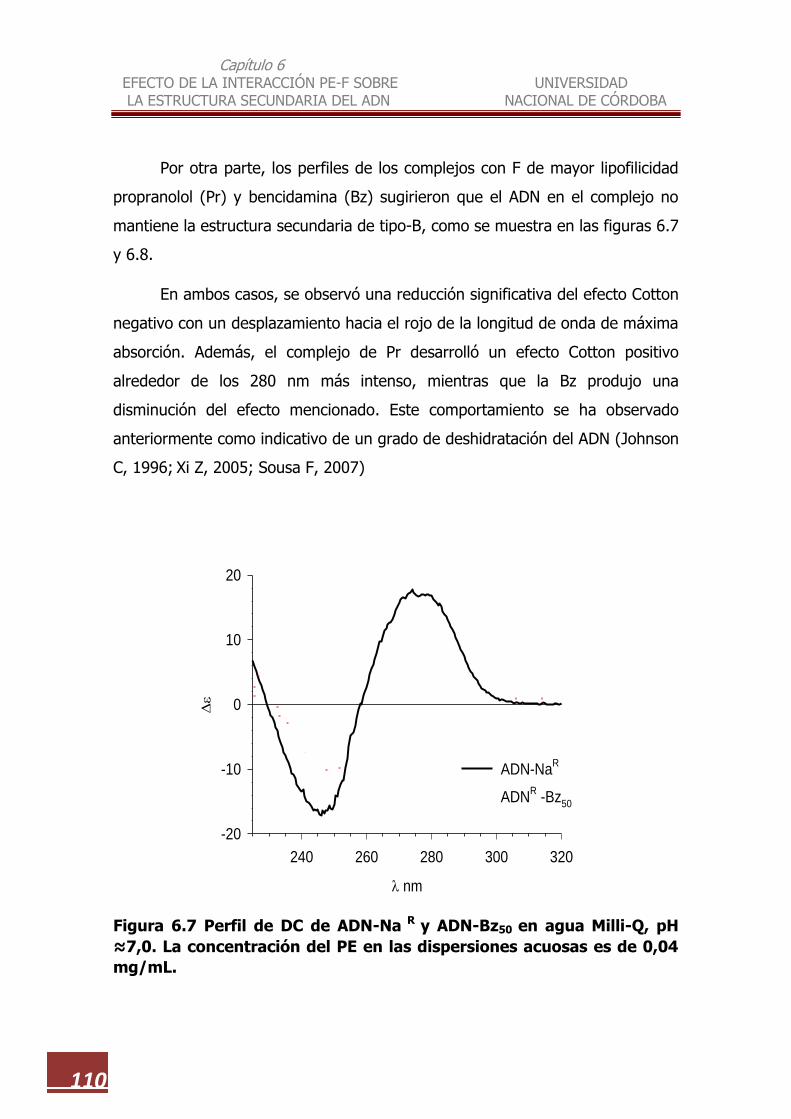
Capítulo 6 EFECTO DE LA INTERACCIÓN PE-F SOBRE UNIVERSIDAD LA ESTRUCTURA SECUNDARIA DEL ADN NACIONAL DE CÓRDOBA
110
Por otra parte, los perfiles de los complejos con F de mayor lipofilicidad
propranolol (Pr) y bencidamina (Bz) sugirieron que el ADN en el complejo no
mantiene la estructura secundaria de tipo-B, como se muestra en las figuras 6.7
y 6.8.
En ambos casos, se observó una reducción significativa del efecto Cotton
negativo con un desplazamiento hacia el rojo de la longitud de onda de máxima
absorción. Además, el complejo de Pr desarrolló un efecto Cotton positivo
alrededor de los 280 nm más intenso, mientras que la Bz produjo una
disminución del efecto mencionado. Este comportamiento se ha observado
anteriormente como indicativo de un grado de deshidratación del ADN (Johnson
C, 1996; Xi Z, 2005; Sousa F, 2007)
Figura 6.7 Perfil de DC de ADN-Na R y ADN-Bz50 en agua Milli-Q, pH
≈7,0. La concentración del PE en las dispersiones acuosas es de 0,04
mg/mL.
nm
240 260 280 300 320
-20
-10
0
10
20
ADN-NaR
ADNR
-Bz50

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
111
Figura 6.8 Perfil de DC de ADN-Na R y ADN-Pr50 en agua Milli-Q, pH
≈7,0. La concentración del PE en las dispersiones acuosas es de 0,04
mg/mL.
6.3.2 Reversibilidad de los cambios en la estructura del ADN
Las dispersiones de los complejos de Pr y Bz se sometieron a una diálisis
exhaustiva en un buffer a pH = 7,0. El ADN recuperado de la dispersión del
complejo con Bz exhibe un perfil DC similar al de la forma nativa, pero el del
complejo con Pr no volvió completamente al perfil original, figuras 6.9 y 6.10. El
Pr recuperado en el medio receptor determinado por espectrometría de UV,
sólo representa el 88,2% de la cantidad original. Este hallazgo sugiere una
contribución de otras interacciones de tipo no iónico. Por lo tanto, los cambios
observados en la estructura secundaria de ADN, como consecuencia de la
interacción con la muestra de F de mayor lipofilicidad ensayados, parecen ser
de naturaleza reversible excepto con Pr.
nm
200 220 240 260 280 300 320
-6
-4
-2
0
2
4
6
8
10
ADN-NaR
ADNR
-Pr50

Capítulo 6 EFECTO DE LA INTERACCIÓN PE-F SOBRE UNIVERSIDAD LA ESTRUCTURA SECUNDARIA DEL ADN NACIONAL DE CÓRDOBA
112
Figura 6.9 Perfil de DC de ADN-Na R y ADN-Bz50 en agua Milli-Q, pH
≈7,0; después de realizar la diálisis exhaustiva.
Figura 6.10 Perfil de DC de ADN-NaR y ADN-Pr50 en agua Milli-Q, pH
≈7,0; después de realizar la diálisis exhaustiva
nm
200 220 240 260 280 300 320 340
-6
-4
-2
0
2
4
6
8
10
ADN-NaR 24 hs
ADNR-Pr 24 hs
nm
200 220 240 260 280 300 320 340
-6
-4
-2
0
2
4
6
8
10
ADN-NaR 24 hs
ADNR
-Bz 24hs

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
113
6.3.3 Efecto de la temperatura sobre la estructura
secundaria de los complejos ADN-F.
En todos los casos el efecto de la temperatura sobre los complejos ADN-
Fx generó cambios en la intensidad de las señales, que volvieron a su estado
original. Se eligen el Ti y Pr como modelos para representar los resultados,
como se puede ver en la figura 6.11 y 6.12, el cambio de las señales entre
temperatura ambiente y temperatura de calentamiento del ADN en presencia
de F, mantiene las señales características de una estructura secundaría
helicoidal, no evidencia una apertura de la doble hélice, como ocurrió con el
ADN sin presencia del F (figura 6.3), sugiriendo de esta manera el aporte del F
a la estabilidad de la doble hélice.
nm
220 240 260 280 300 320
-30
-20
-10
0
10
20
30
ADN-Na T 25°C
ADN-Ti T 25°C
ADN-Ti T 78°C
ADN-Ti Enfriamiento
Figura 6.11 Efecto de la temperatura sobre el perfil de DC de una
dispersión acuosa de ADN-Ti50 a pH ≈7,0.
λ nm

Capítulo 6 EFECTO DE LA INTERACCIÓN PE-F SOBRE UNIVERSIDAD LA ESTRUCTURA SECUNDARIA DEL ADN NACIONAL DE CÓRDOBA
114
Figura 6.12 Efecto de la temperatura sobre el perfil de DC de una
dispersión acuosa de ADN-Pr50 a pH ≈7,0.
6.4. Conclusiones
Se evidenció que las sales de ADN presentan una estructura secundaria
nativa tipo-B, que es susceptible al efecto de la temperatura.
Se observa un efecto diferenciado de los F sobre la estructura secundaria
del ADN-Na, según sus propiedades lipofílicas.
Los complejos ADN-F con F de menor lipofilicidad (At, Li y Ti), conservan
la estructura secundaria nativa tipo-B original del ADN-Na.
nm
220 240 260 280 300 320 340
-30
-20
-10
0
10
20
30
40
50
ADN-Pr T25°C
ADN-Pr T 78°C
ADN-Pr Enfriamiento
ADN-Na T25°C

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
115
Los complejos ADN-F con F de mayor lipofilicidad (Pr y Bz), no
mantienen la estructura secundaria nativa tipo-B original del ADN-Na.
Los cambios generados a la estructura secundaria por la Bz son
reversibles.
Los cambios generados a la estructura secundaria por el Pr no son de
naturaleza reversible, lo que sugiere la posible contribución de otras
interacciones de tipo no iónico en el complejo ADN-Pr.
El efecto de la temperatura sobre la estructura de los complejos es de
naturaleza reversible, lo que sugiere un aporte a la estabilidad de la
estructura por parte del F.

Capítulo 6 EFECTO DE LA INTERACCIÓN PE-F SOBRE UNIVERSIDAD LA ESTRUCTURA SECUNDARIA DEL ADN NACIONAL DE CÓRDOBA
116

Conclusiones
Generales y
Proyecciones


UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
119
Conclusiones Generales y Proyecciones
La caracterización in vitro de las interacciones ADN-F desarrollada en
esta tesis, proporciona una base físicoquímica con proyecciones en varios
campos de interés farmacoterapéutico.
El esquema 7.1 representa las principales interacciones observadas
entre el grupo de F utilizado y el ADN modelo.
Esquema 7.1. Caracterización de las interacciones iónicas ADN-F
El conjunto de ensayos realizado sobre la interacción ADN-F puso de
manifiesto el comportamiento de los F utilizados con el AN modelo.

UNIVERSIDAD
CONCLUSIONES GENERALES Y PROYECCIONES NACIONAL DE CÓRDOBA
120
Por una parte, se observó un comportamiento de tipo general que comprende
la interacción electrostática inespecífica análoga a la que ocurre con otros PE
ácidos de menor complejidad que los AN. Los resultados obtenidos sobre los
equilibrios involucrados en la interacción ADN-F fueron satisfactoriamente
interpretados mediante la ecuación 6 del capitulo 4 y la cinética de liberación
difusional de los F fue consistente con el modelo propuesto para los PE ácidos.
1. Por otra parte, se observó un comportamiento diferenciado del efecto de
la interacción sobre la estructura secundaria del ADN, que en una
primera aproximación se clasificó en función de las propiedades
hidrofílicas- lipofílicas de los F estudiados.
En función de las altas constantes de afinidad ADN-F observadas, es
razonable pensar que, como resultado de la administración farmacoterapéutica
de F básicos con alta permeabilidad sobre las membranas celulares, en la
secuencia farmacocinética que se desarrolla, los AN (ADN y ARN), emplazados
en citoplasma y núcleo, constituyan un significativo blanco intracelular (ver
introducción página 14).
En consecuencia, si las interacciones con esta clase de F pueden afectar
la funcionalidad de los AN es una cuestión que surge como una de las
proyecciones de los resultados presentados. Más aún, teniendo en cuenta los
avances actuales en los campos de la genética y la epigenética (Narayanan et
al., 2017; Wong et al., 2017; Xiao P, 2017).
Basados en lo anterior, en una primera instancia los resultados
descriptos permitirían focalizar el estudio del potencial impacto de la interacción
ADN-F en sistemas in vivo utilizando aquellos F que afectan la conformación y

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
121
Interacción ADN-F Sobre la conformación
No
afecta Afecta
Efecto
reversible
Efecto no reversible en su totalidad
particularmente en los que evidencian que no ocurre su completa reversibilidad
como es el caso del propranolol (Esquema 7.2).
Esquema 7.2 Comportamiento diferenciado de la interacción ADN-F
sobre la estructura secundaria del ADN
Por otra parte, el comportamiento de la interacción ADN-F descripto en
el inciso 1 y los resultados sobre la obtención de los complejos en estado sólido
y su posterior redispersión, podrían contribuir al uso de ADN como portador de
F para el diseño de formulaciones de liberación modificada; por lo que el
estudio de las propiedades de relevancia biofarmacéutica y farmacotécnica de
estos sistemas constituyen otra proyección de los resultados presentados.

UNIVERSIDAD
CONCLUSIONES GENERALES Y PROYECCIONES NACIONAL DE CÓRDOBA
122
Finalmente, el estudio de la transfección de AN es un área de gran
interés. Los vectores no virales que se utilizan en la actualidad para el
direccionamiento de genes al interior celular, como los plásmidos y la
introducción de RNA de bajo peso molecular (miRNA, siRNA), requieren ser
vehiculizados en liposomas y polímeros lipídicos catiónicos capaces de modificar
la carga superficial negativa de los AN. Dado que los complejos ADN-F
presentan una reducción de la carga superficial, es de interés explorar su
potencial utilidad en el campo de la transfección intracelular de ADN y ARN,
generando una alternativa local, segura y asequible en comparación a los
polímeros utilizados actualmente.

BIBLIOGRAFÍA


UNIVERSIDAD
BIBLIOGRAFÍA NACIONAL DE CÓRDOBA
125
A
Acharya, S. Digital Comprehensive Summaries of Uppsala Dissertations
from the Faculty of Science and Technology 164. Some Aspects of
Physicochemical Properties of DNA and RNA; Acta Universitatis Upsaliensis:
Uppsala, Sweden, 2006.
Ansel HC, Allen L V, Popovich NG. Disperse Systems. Ansels´s
Pharmaceutical Dosage Forms and Drug Delivery systems. Baltimore: Williams
& Williams. 9°ed. 2011:376-430.
Ardusso MS, Manzo RH, Jimenez-Kairuz A.F. Comparative study of three
structurally related acid polyelectrolytes as carriers of basic drugs: Carbomer,
Eudragit L-100 and S-100. Supramolecular Chemistry. 2010;22(5):289-296.
doi:10.1080/10610270903469757.
Avdeef, A. Absorption and Drug Development: Solubility, Permeability,
and Charge State; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003.
B
Ban C, Ramakrishnan B, Sundaralingam M. Crystal structure of the highly
distorted chimeric decamer r (C) d (CGGCGCCG) r (G) * spermine complex -
spermine binding to phosphate only and minor groove tertiary. Nucleic Acids
Research. 1994;22(24):5466-5476.
Banker, G, Rhodes, C. Modern Pharmaceutics, 1996;4th ed. New York.
Batool H. Design And In-Vitro Evaluation Of Sustained Release Matrix
Tablets Of Eperisone Hydrochloride Using Carbopol 934 P. International Journal
of Pharmacy and Integrated Life Sciences. 2013;2(January):28-41

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
126
Battistini FD. Desarrollo de complejos Polielectrolito-Fármaco y su
utilización en el diseño de sistemas de liberación modificada. Tesis doctoral.
2015. Facultad de Ciencias Químicas, Universidad Nacional de Córdoba
Battistini FD, Flores-Martin J, Olivera ME, Genti-Raimondi S, Manzo RH.
Hyaluronan as drug carrier. the in vitro efficacy and selectivity of Hyaluronan-
Doxorubicin complexes to affect the viability of overexpressing CD44 receptor
cells. European Journal of Pharmaceutical Sciences. 2014; 65: 122-129.
doi:10.1016/j.ejps.2014.09.008.
Battistini FD, Olivera ME, Manzo RH. Equilibrium and release properties
of hyaluronic acid-drug complexes. European Journal of Pharmaceutical
Sciences. 2013;49(4):588-594. doi:10.1016/j.ejps.2013.04.023.
Blackburn, G. M.; Gait, M. J. Nucleic Acids in Tiera Chemistry and
Biology; OxfordUniversity Press, NewYork, 1996.
Bordelon JA, Feierabend KJ, Siddiqui SA, Wright LL, Petty JT. Viscometry
and Atomic Force Microscopy Studies of the Interactions of a Dimeric Cyanine
Dye with DNA. J Phys Chem B. 2002; 106:4838-4843.
Bougas A, Vlachogiannis IA, Gatos D, Arenz S, Dinos G. Dual effect of
chloramphenicol peptides on ribosome inhibition. Amino Acids. 2017.
10.1007/s00726-017-2406-5.
Braña MF, Cacho M, Gradillas A, Ramos A. Intercalators as Anticancer
Drugs. Current Pharmaceutical Desing. 2001; 7: 1745-1780.
Brown CK, Friedel HD, Barker AR, et al. FIP / AAPS Joint Workshop
Report: Dissolution / In Vitro Release Testing of Novel / Special Dosage Forms.
AAPS Pharm Sci Tech, Vol. 2011;12(2):782-794. doi:10.1208/s12249-011-9634-
x.

UNIVERSIDAD
BIBLIOGRAFÍA NACIONAL DE CÓRDOBA
127
Budhian A, Siegel SJ, Winey KI. Controlling the in vitro release profiles
for a system of haloperidol-loaded PLGA nanoparticles. International Journal of
Pharmaceutics. 2008; 346: 151-159. doi:10.1016/j.ijpharm.2007.06.011.
Burkard ME, Kierzek R, Turner DH. Thermodynamics of Unpaired
Terminal Nucleotides on Short RNA Helixes Correlates with Stacking at Helix
Termini in Larger RNAs. j Mol Biol. 1999; 290: 967-982.
C
Chamberlin SI, Merino EJ, Weeks KM. Catalysis of amide synthesis by
RNA phosphodiester and hydroxyl groups. Proc Natl AcadSci.
2002;99(23):14688-14693.
Cheng, X., Zhang, F., Zhou, G., Gao, S., Dong, L., Jiang, W., & Ding, Z.
DNA / chitosan nanocomplex as a novel drug carrier for doxorubicin. Drug
Delivery, 2009; 16: 135–144. https://doi.org/10.1080/10717540802605376
Comisión Europea. Diario Oficial de la Unión Europea, L 97, 05 de abril
de 2006. Diario Oficial de la Unión Europea. 2006;(2).
Crick, F.H.C.; Watson, J.D. The complementary structure of
deoxyribonucleic acid, Proc. Royal Soc. 1954, 223,80–96.
D
Description, Atenolol. Clinical Pharmacology. Available online:
http://www.clinicalpharmacology.com/Forms/drugoptions.aspx?cpnum=46&n=
Atenolol&t=0 (accessed on 28 December 2016).

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
128
Description, Propranolol. Clinical Pharmacology. Available online:
http://www.clinicalpharmacology.com/Forms/drugoptions.aspx?cpnum=1083&n
=Hydrochlorothiazide%2c+HCTZ%3b+Propranolol&t=0 (accessed on 28
December 2016).
Dobrynin A V, Rubinstein M. Theory of polyelectrolytes in solutions and
at surfaces. Progress in Polymer Science. 2005; 30: 1049-1118.
doi:10.1016/j.progpolymsci.2005.07.006.
E
Esteban SL, Manzo RH, Alovero FL. Azithromycin loaded on hydrogels of
carbomer: Chemical stability and delivery properties. International Journal of
Pharmaceutics. 2009;366(1-2):53-57. doi:10.1016/j.ijpharm.2008.08.043.
F
FDA. Guidance for Industry Nonsterile Semisolid Dosage Forms. Scale-Up
and Postapproval Changes: Chemistry, Manufacturing, and Controls; In Vitro
Release Testing and In Vivo Bioequivalence Documentation. SUPAC-SS. US
Department of Health and Human Services Food and Drug Administration
Center for Drug Evaluation and Research (CDER). 1997;(May).
Feigon J, Hud N V, Sklena V. Localization of Ammonium Ions in the Minor
Groove of DNA Duplexes in Solution and the Origin of DNA A-Tract Bending. J
Mol Biol. 1999; 286: 651-660.
Feuerstein, B.G.; Pattabiraman, N.; Marton, L.J. Molecular mechanics of
the interactions of spermine with DNA: DNA bending as a result of ligand
binding. Nucleic Acids Res. 1990, 18, 1271–1282.

UNIVERSIDAD
BIBLIOGRAFÍA NACIONAL DE CÓRDOBA
129
Franz TJ. The Finite Dose Technique as a Valid. Curr Probl Dermatol.
1978; 7: 58-68.
Fukushima T, Inoue Y, Hayakawa T, Taniguchi K, Miyazaki K, Okahata Y.
Preparation of and tissue response to DNA-lipid films. J Dent Res.
2001;80(8):1772-1776. doi:10.1177/00220345010800081801.
G
García JP, Marrón E, Martín VI, Moyá ML, Lopez-Cornejo P.
Conformational changes of DNA in the presence of 12-s-12 gemini surfactants
(s=2 and 10). Role of the spacer’s length in the interaction surfactant-
polynucleotide. Colloids and Surfaces B: Biointerfaces. 2014; 118: 90-100.
doi:10.1016/j.colsurfb.2014.03.040.
Getts R; Muro S. DNA-Based Drug Carriers: The Paradox of a Classical
“Cargo” Material Becoming a Versatile “Carrier” to Overcome Barriers in
Drug Delivery. Current Pharmaceutical Design. 2016; 22 (9): 1245 – 1258.
doi:10.2174/1381612822666151216151614.
Guzmán ML, Manzo RH, Olivera ME. Eudragit E100 as a drug carrier: The
remarkable affinity of phosphate ester for dimethylamine. Molecular
Pharmaceutics. 2012;9(9):2424-2433. doi:10.1021/mp300282f.
H
Hackl E.V.; Kornilova S.V.; Blagoi Y.P. DNA structural transitions induced
by divalent metal ions in aqueous solutions. Int. J. Biol. Macromol. 2005, 35,
175–191.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
130
Hajian, R.; Shams, N.; Mohagheghian, M. Study on the interaction
between doxorubicin and deoxyribonucleic acid with the use of methylene blue
as a probe. J. Braz. Chem. Soc. 2009, 20, 1399–1405.
Hasan EI, Amro BI, Arafat T, Badwan AA. Assessment of a controlled
release hydrophilic matrix formulation for metoclopramide HCl. European
Journal of Pharmaceutics and Biopharmaceutics. 2003; 55: 339-344.
doi:10.1016/S0939-6411(03)00022-5.
Hasanzadeh M, Shadjou N. Pharmacogenomic study using bio- and
nanobioelectrochemistry: Drug – DNA interaction. Materials Science and
Engineering C. 2016; 61: 1002-1017.
Higuchi T. Mechanism of Sustained- Action Medication: Theoretical
Analysis of Rate of Release of Solid Drugs Dispersed in solid matrices. Journal
of pharmaceutical sciences. 1963;52(12):1145-1149.
Holen E, Bjørge OA, Jonsson R. Dietary nucleotides and human immune
cells. II. Modulation of PBMC growth and cytokine secretion. Nutrition.
2005;21(10):1003-1009. doi:10.1016/j.nut.2005.03.003.
I
Inoue Y, Fukushima T, Hayakawa T, et al. Antibacterial characteristics of
newly developed amphiphilic lipids and DNA–lipid complexes against bacteria.
Journal of Biomedical Materials Research Part A. 2003; 65(2) :203-208.
doi:10.1002/jbm.a.10483.

UNIVERSIDAD
BIBLIOGRAFÍA NACIONAL DE CÓRDOBA
131
J
Jiménez Kairuz A.F., Ramírez M.V., Quinteros D.A., Vilches A.P., Olivera
M.E., Alovero F., Manzo R.H. Recientes contribuciones sobre la utilización de
Polielectrolitos en Sistemas portadores de F. Parte I: Dispersiones acuosas.
Revista farmacéutica de la Academia Nacional de Farmacia y Bioquímica.
2008.150: 11-25
Jiménez Kairuz A.F., Ramírez M.V., Quinteros D.A., Vilches A.P., Olivera
M.E., Alovero F., Manzo R.H. Recientes contribuciones sobre la utilización de
Polielectrolitos en Sistemas portadores de F. Parte II: Materiales Solidos.
Revista farmacéutica de la Academia Nacional de Farmacia y Bioquímica.
2008.150: 6-21
Jimenez-Kairuz AF, Llabot JM, Allemandi DA, Manzo RH. Swellable drug-
polyelectrolyte matrices (SDPM). International Journal of Pharmaceutics.
2005;288(1):87-99. doi:10.1016/j.ijpharm.2004.09.014.
Jimenez-Kairuz AF. Investigación y desarrollo de nuevos materiales con
utilidad en tecnología farmacéutica para el diseño de sistemas terapéuticos.
Tesis doctoral.2004. Facultad de Ciencias Químicas, Universidad Nacional de
Córdoba.
Jimenez-Kairuz AF, Allemandi DA, Manzo RH. The improvement of
aqueous chemical stability of a model basic drug by ion pairing with acid groups
of polyelectrolytes. International Journal of Pharmaceutics. 2004; 269(1): 149-
156. doi:10.1016/j.ijpharm.2003.09.008.
Jimenez-Kairuz AF, Allemandi DA, Manzo RH. Equilibrium properties and
mechanism of kinetic release of metoclopramide from carbomer hydrogels.
International Journal of Pharmaceutics. 2003; 250(1): 129-136.
doi:10.1016/S0378-5173(02)00525-2.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
132
Jimenez-kairuz A, Allemandi D, Manzo RH. Mechanism of Lidocaine
Release from Carbomer ± Lidocaine Hydrogels. Journal of pharmaceutical
sciences. 2002; 91(1): 267-272.
Johnson C. Determination of the Conformation of Nucleic Acids by
Electronic CD. Circular Dichroism and the Conformational Analysis of
Biomolecules. 1996; (12): 433-465.
K
Kankia BI, Buckin V, Bloomfield VA. Hexamminecobalt (III) -induced
condensation of calf thymus DNA: circular dichroism and hydration
measurements. 2001; 29(13): 2795-2801.
Kopka ML, Yoon C, Goodsell D, Pjura P, Dickerson RE. Binding of an
Antitumor Drug to DNA. J Mol Biol. 1985; 183: 553-563.
Korsmeyer R, Gurny R, Doelker E, Buri P, Peppas N. Mechanisms of
Solute Release from Porous Hydrophilic Polymers Mechanisms of solute release
from porous hydrophilic polymers. International Journal of Pharmaceutics.
1983;15(May 2016):25-35. doi:10.1016/0378-5173(83)90064-9.
L
Lehninger A, Nelson DL, Cox MM. The Citric Acid Cycle. Langmuir: the
ACS journal of surfaces and colloids. 2008: 630.
Leng F, Chaires JB, Waring MJ, Street SW. Energetics of echinomycin
binding to DNA. Nucleic Acids Research. 2003; 31(21): 6191-6197.
doi:10.1093/nar/gkg826.

UNIVERSIDAD
BIBLIOGRAFÍA NACIONAL DE CÓRDOBA
133
Liu W, Sun S, Cao Z, et al. An investigation on the physicochemical
properties of chitosan/DNA polyelectrolyte complexes. Biomaterials. 2005;
26(15): 2705-2711. doi:10.1016/j.biomaterials.2004.07.038.
Lotfipour F, Nokhodchi A, Saeedi M, Norouzi-sani S. The effect of
hydrophilic and lipophilic polymers and fillers on the release rate of atenolol
from HPMC matrices. IL FARMACO. 2004; 59: 819-825.
doi:10.1016/j.farmac.2004.06.006.
M
Manning, G.S. The molecular theory of polyelectrolyte solutions with
applications to the electrostatic properties of polynucleotides, Q. Rev. Bio-phys.
1978; 11: 179–246.
Martin, A. Colloids. En Physical Pharmacy. 4ta Ed. Williams & Wilkins.
Baltimore.USA. 1993. Pág. 406.
Martínez-Augustin O, Boza JJ, Navarro J, Martínez-Valverde A, Araya M,
Gil A. Dietary nucleotides may influence the humoral immunity in
immunocompromised children. Nutrition. 1997; 13(5): 465-469.
doi:10.1016/S0899-9007(97)00012-9.
McGregor, T.D.; Bousfield, W.; Qu, Y.; Farrell, N. Circular dichroism
study of the irreversibility of conformational changes induced by polyamine-
linked dinuclear platinum compounds. J. Inorg. Biochem. 2002, 91, 212–219.
Mizuno, Y.; Naoi, T.; Nishikawa, M.; Rattanakiat, S.; Hamaguchi, N.;
Hashida, M.; Takakur, Y. Simultaneous delivery of doxorubicin and
immunostimulatory CpG motif to tumors using a plasmid DNA/doxorubicin
complex in mice. J. Control. Release 2010, 141, 252–259.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
134
N
Navarro J, Maldonado J, Narbona E, et al. Influence of dietary
nucleotides on plasma immunoglobulin levels and lymphocyte subsets of
preterm infants. BioFactors (Oxford, England). 1999; 10(1): 67-76.
Narayanan S, Singh S, Shukla S. A saga of cancer epigenetics: linking
epigenetics to alternative splicing. Biochem. J. 2017; 474: 885–896.
doi:10.1042/BCJ20161047
Nerella A, D BR, M AD. Formulation, Optimization and In vitro
Characterization of Letrozole Loaded Solid Lipid Nanoparticles. International
Journal of Pharmaceutical Sciences and Drug Research. 2014; 6(3): 183-188.
Nielsen JE, Vriend G. Optimizing the Hydrogen-Bond Network in Poisson
– Boltzmann Equation-Based p K a Calculations. PROTEINS: Structure,
Function, and Genetics. 2001; 412: 403-412.
O
Ohmichi T, Nakano S, Miyoshi D, Sugimoto N. Long RNA Dangling End
Has Large Energetic Contribution to Duplex Stability. 2002; (17): 10367-10372.
P
PubChem Compound Database; Benzydamine CID 12555. Available
online: https://pubchem.ncbi.nlm.nih.gov/compound/12555#section=Chemical-
and-Physical-Properties (accessed 28 December 2016).

UNIVERSIDAD
BIBLIOGRAFÍA NACIONAL DE CÓRDOBA
135
PubChem Compound Database; Lidocaíne CID 3676. Available online:
https://pubchem.ncbi.nlm.nih.gov/compound/3676#section=Solubility
(accessed on: 28 December 2016).
PubChem Compound Database; racemic-Timolol CID 5478. Available
online: https://pubchem.ncbi.nlm.nih.gov/compound/5478#section=Solubility
(accessed on: 28 December 2016).
Q
Quinteros DA, Allemandi DA, Manzo RH. Equilibrium and Release
Properties of Aqueous Dispersions of Non-Steroidal Anti-Inflammatory Drugs
Complexed with. Scientia Pharmaceutica. 2012; 80: 487-496.
doi:10.3797/scipharm.1107-17.
Quinteros DA, Manzo RH, Allemandi DA. Interaction Between Eudragit R
E100 and Anionic Drugs: Addition of Anionic Polyelectrolytes and Their
Influence on Drug Release Performance. Journal of pharmaceutical sciences.
2011; 100: 4664-4673. doi:10.1002/jps.
Quinteros DA; Ramirez MV; Jiménez Kairuz AF; Olivera ME; Manzo RH;
Allemandi DA. Interaction between a cationic polymethacrylate (Eudragit E100)
and anionic drugs. European Journal of Pharmaceutical Sciences. 2008; 33: 72-
79. doi:10.1039/b000000x.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
136
R
Ramírez Rigo M V, Allemandi DA, Manzo RH. A Linear Free Energy
Relationship Treatment of the Affinity between Carboxymethylcellulose and
Basic Drugs. Molecular pharmaceutics. 2004; 1(5): 383-386.
doi:10.1021/mp0499353.
Ramírez Rigo M V, Allemandi DA, Manzo RH. Swellable drug-
polyelectrolyte matrices of drug-carboxymethylcellulose complexes.
Caracterization ande delivery properties. Drug delivery. 2009; 16(2):1 08-115.
doi:10.1016/j.ijpharm.2006.05.025.
Ramírez Rigo M V, Allemandi DA, Manzo RH. Swellable drug-
polyelectrolyte matrices (SDPM) of alginic acid. Characterization and delivery
properties. International Journal of Pharmaceutics. 2006; 322(1-2): 36-43.
doi:10.1016/j.ijpharm.2006.05.025.
Ranga K., Padmalatha K, Buri P. Cellulose matrices for zero-order release
of soluble drugs. Drug development and industrial pharmacy. 1988; 4: 2299-
2320.
Rau I, Grote JG, Kajzar F, Pawlicka A. DNA - novel nanomaterial for
applications in photonics and in electronics. Comptes Rendus Physique. 2012;
13(8): 853-864. doi:10.1016/j.crhy.2012.09.005.
Ritger PL, Peppas NA. A simple equation for description of solute release
ii. Fickian and anomalous release from swellable devices. Journal of Controlled
Release. 1987; 5: 37-42.
Robert H.; Moran,L.; Scrimgeour, G.; Perry, M. JDR. Principios de
Bioquímica. Cuarta edi. México: pearson educación; 2008; Chapter 19

UNIVERSIDAD
BIBLIOGRAFÍA NACIONAL DE CÓRDOBA
137
Rolland, A.; Sullivan, S.M. Pharmaceutical Gene Delivery Systems. Drugs
and the Pharmaceutical Science, Taylor & Francis Group: Boca Raton, FL, USA,
2003; 131.
Romero VL, Manzo RH, Alovero FL. Enhanced bacterial Uptake and
bactericidal properties of Ofloxacin loaded on bioadhesive Hydrogels Against
Pseudomonas aeruginosa. Journal of Chemotherapy. 2010; 22(5): 328-334.
S
Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory
Manual; Cold Spring Harbor Laboratory: New York, NY, USA, 1989; p. 626.
Sasaki Y, Miyoshi D, Sugimoto N. Utilization of salmon milt DNA against
UV damage. Applied Biochemistry and Biotechnology. 2010;160(8):2458-2466.
doi:10.1007/s12010-009-8697-6.
Shah VP, Elkins JS, Williams RL. Evaluation of the Test System Used for
in Vitro Release of Drugs for Topical Dermatological Drug Products.
Pharmaceutical Development and Techonology. 1999;4(3):377-385.
Shah, A.; Khan, A.M.; Usman, M.; Siddiq, M. Plex with DNA as Studied by
Conductometric and Spectroscopic Techniques. J. Chil. Chem. Soc. 2009, 2,
134–137.
Shaikh SA, Ahmed SR, Jayaram B. A molecular thermodynamic view of
DNA–drug interactions: a case study of 25 minor-groove binders. Archives of
Biochemistry and Biophysics. 2004; 429: 81-99. doi:10.1016/j.abb.2004.05.019.
Shen X, Nagai N, Murata M, Nishimura D, Sugi M, Munekata M.
Development of salmon milt DNA/salmon collagen composite for wound
dressing. Journal of Materials Science: Materials in Medicine. 2008;19(12):3473-
3479. doi:10.1007/s10856-008-3512-4.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
138
Shimanovich U, Eliaz D, Aizer A, Vayman I, Michaeli S, Shav-tal Y.
Sonochemical Synthesis of DNA Nanospheres. 2011:1678-1681.
doi:10.1002/cbic.201100009.
Sousa F, Prazeres DMF, Queiroz JA. Circular dichroism investigation of
the effect of plasmid DNA structure on retention in histidine chromatography.
Archives of Biochemistry and Biophysics. 2007;467(2):154-162.
doi:10.1016/j.abb.2007.08.027.
Stepankova, J.; Studenovsky, M.; Malina, J.; Kasparkova, J.; Liskova, B.;
Novakova, O.; Ulbrich, K.; Brabec, V. DNA interactions of 2-
pyrrolinodoxorubicin, a distinctively more potent daunosamine-modified
analogue of doxorubicin. Biochem. Pharmacol. 2011, 82, 227–235.
T
Talukdar MM, Kinget R. journal of pharmaceutics Swelling and drug
release behaviour of xanthan gum matrix tablets. International Journal of
Pharmaceutics. 1995; 120: 63-72.
Tanaka K, Okahata Y. A DNA- Lipid Complex in Organic Media and
Formation of an Aligned Cast Film1. J Am Chem Soc. 1996;118(44):10679–
10683. doi:10.1021/ja9617855.
Taylor P, Leszczynski J, Hobza P. Dynamics Hydrogen Bonding and
Stacking of DNA Bases : A Review of Quantum-chemical ab initio Studies
Hydrogen. Journal of Biomolecular Structure 2015:37-41.
doi:10.1080/07391102.1996.10508935.

UNIVERSIDAD
BIBLIOGRAFÍA NACIONAL DE CÓRDOBA
139
Tiera MJ, Shi Q, Winnik FM, Fernandes JC. Polycation-Based Gene
Therapy : Current Knowledge and New Perspec- tives. Current Gene Therapy.
2011; 11: 288-306.
U
Um, S.H.; Lee, J.B.; Park, N.; Kwon, S.Y.; Umbach, C.C.; Luo, D.
Enzyme-catalysed assembly of DNA hydrogel. Nat. Mater. 2006, 5, 97–801.
USP 34-NF 29. The United States Pharmacopoeia, 34th ed.; U.S.
Pharmacopoeial Convention., Inc., Rockville, MD, USA, 2011.
Utsuno, K.; Uluda, H. Thermodynamics of polyethylenimine-DNA binding
and DNA condensation. Biophys. J. 2010, 99, 201–207.
Uytterhoeven K, Sponer J, Meervelt L Van. Two 1: 1 binding modes for
distamycin in the minor groove of d ( GGCCAATTGG ). Eur J Biochem. 2002;
269: 2868-2877. doi:10.1046/j.1432-1033.2002.02952.
V
Van Dam L, Levitt MH. BII nucleotides in the B and C forms of natural-
sequence polymeric DNA: A new model for the C form of DNA. Journal of
molecular biology. 2000; 304(4): 541-561. doi:10.1006/jmbi.2000.4194.
Van Den Beucken J, Vos MRJ, Thüne PC, et al. Fabrication,
characterization, and biological assessment of multilayered DNA-coatings for
biomaterial purposes. Biomaterials. 2006; 27(5): 691-701.
doi:10.1016/j.biomaterials.2005.06.015.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
140
Vilches AP, Jimenez-Kairuz A, Alovero F, Olivera ME, Allemandi DA,
Manzo RH. Release kinetics and up-take studies of model fluoroquinolones from
carbomer hydrogels. International Journal of Pharmaceutics. 2002; 246(1-2):
17-24. doi:10.1016/S0378-5173(02)00333-2.
Visakh, P.M.; Bayraktar, O. Polyelectrolytes Thermodynamics and
Rheology; Springer: Heidelberg, Germany, 2014.
W
Wahyuni E, Tjahjono D, Yoshioka N, Inoue H. Spectroscopic studies on
the thermodynamic and thermal denaturation of the ct-DNA binding of
methylene blue. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2010; 77: 528–
534.
Walker G, Stone M, Krugh T. Interaction of Drugs with Z-DNA:
Cooperative Binding of Actinomycin D or Actinomine to the Left-Handed Forms
of Poly( dG-dC).Poly( dG-dC) and Poly( dG-m5dC)*Poly( dG-m5dC) Reverses
the Conformation of the Helix. Biochemistry. 1985; 24(1980): 7471-7479.
Wang L, Yoshida J, Ogata N, Sasaki S, Kajiyama T. Self-assembled
supramolecular films derived from marine deoxyribonucleic acid (DNA) -
Cationic surfactant complexes: Large-scale preparation and optical and thermal
properties. Chemistry of Materials. 2001; 13(4): 1273-1281.
doi:10.1021/cm000869g.
Watson J, Crick F. Molecular structure of nucleic acids. A structure for
deoxyribose nucleic acid, Nature. 1953; 171: 737–738.
Wong C, Qian Y, Yu J. 2017. Interplay between epigenetics and
metabolism in oncogenesis: mechanisms and therapeutic approaches.
Oncogene.2017; 1–16. doi:10.1038/onc.2016.485

UNIVERSIDAD
BIBLIOGRAFÍA NACIONAL DE CÓRDOBA
141
X
Xi Z, Ouyang D, Mu HT. Interaction of bulged DNA with leucine-
containing mimics of NCS-chrom. Bioorganic and Medicinal Chemistry Letters.
2006; 16(5): 1185-1190. doi:10.1016/j.bmcl.2005.11.084.
Xiao Pfister S, Ashworth A. Marked for death: targeting epigenetic
changes in cancer. Nat. Publ. Gr. 2017; 1-23 doi:10.1038/nrd.2016.256
Y
Yamada M, Yokota M, Kaya M, et al. Preparation of novel bio-matrix by
the complexation of DNA and metal ions. Polymer. 2005; 46(23): 10102-10112.
doi:10.1016/j.polymer.2005.07.086.
Yoshida J, Wang LL, Kobayashi S, Zhang GJ, Ikeda H, Ogata N. Optical
properties of photochromic-compound-doped marine-biopolymer DNA-
surfactant complex films for switching applications. Organic Photonic Materials
and Devices Vi. 2004; 5351: 260-267 ST-Optical properties of photochromic-c.
doi:10.1117/12.528781.
Z
Zsila F. Apparent circular dichroism signature of stirring-oriented DNA
and drug-DNA complexes. International Journal of Biological Macromolecules.
2015; 72: 1034-1040. doi:10.1016/j.ijbiomac.2014.10.009.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
142

PUBLICACIONES y
PRESENTACIONES


UNIVERSIDAD
PUBLICACIONES Y PRESENTACIONES NACIONAL DE CÓRDOBA
145
Publicaciones en revistas especializadas
Alarcón LP, Baena Y, Manzo RH. Interaction between DNA and Drugs
Having Protonable Basic Groups: Characterization through Affinity Constants,
Drug Release Kinetics, and Conformational Changes. Scientia Pharmaceutica.
2017;85(1):1-14. doi:10.3390/scipharm85010001.
Participación en reuniones científicas
“Interaction between nucleic acids and drugs having protonable basic
groups. Characterization through affinity constants, drug release kinetics and
conformational changes.” RICIFA Cuarta Reunión Internacional de Ciencias
Farmacéuticas Rosario, Argentina Facultad de Ciencias Químicas UNC Oct 2016
“Efectos sobre la estructura secundaria del ADN generado por el
acomplejamiento de los grupos fosfato con fármacos ionizables. Estudio por
dicroísmo circular y espectroscopia de correlación fotónica.” COIFFA III
Congreso Sudamericana de Biofarmacia y Farmacocinética. Córdoba, Argentina
Facultad de Ciencias Químicas UNC. Nov. 2015
“Study of the formation of ion pairs between the phosphate groups of
deoxyribonucleic acid (DNA) and ionizable drugs therapeutically useful”. RICIFA
Tercera Reunión Internacional de Ciencias Farmacéuticas Córdoba, Argentina
Facultad de Ciencias Químicas UNC. Sep 2014

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA

ANEXO I
MANEJO DEL ELECTRODO DE SODIO


Anexo I UNIVERSIDAD
MANEJO DEL ELECTRODO DE SODIO NACIONAL DE CÓRDOBA
149
1. OBJETIVO: Establecer el procedimiento a seguir para la operación del electrodo selectivo
de Sodio marca METTLER TOLEDO perfectIONTM comb Na+.
2. ALCANCE: Este instructivo aplica a todas las mediciones de actividad de Sodio y
concentración de Sodio en sistemas acuosos. Ver Anexo 1
3. RESPONSABLES:
3.1. DE REALIZARLO: Tesistas de Doctorado y de Posdoctorado, practicantes del laboratorio.
3.2. DE VERIFICARLO: Profesores Encargados del Laboratorio
4. FRECUENCIA: Cada vez que se realice una medición.
5. PRECAUCIONES Y RECOMENDACIONES:
5.1. No se recomienda la calibración por debajo de 1 x 10-4 M, ya que las repuestas pueden
mostrar cierta curvatura.
5.2. La temperatura de todas las muestras a medir no debe tener entre ellas una diferencia
mayor a los 2°C.
5.3. Ver Anexo 2
6. SIMBOLOS Y/O ABREVIATURAS:
6.1. O: Operador del equipo
6.2. PEL: Profesor encargado del Laboratorio
6.3. ISE: Ion Selective Electrode: Electrodo de Ion Selectivo
6.4. ISAS: Ionic Strehgth Adjuster Solution: Solución de ajuste de la fuerza iónica.
7. CALCULO
7.1 Un punto de Calibración
Cu = Cs * 10ΔE/S, dónde Cu = concentración desconocida, Cs= Concentración de la solución
estándar, ΔE= diferencia de potencial muestra y estándar, S = Pendiente de la recta de
calibración.
8. DIAGRAMA DE FLUJO:
8.1. FLUJOGRAMA DE ACTIVIDADES:

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
150
INICIO
Determinación de Concentración de Na Verificación de material de vidrio limpio
1. Etapa A: Preparación de ISAS Pesar 200 g NH4 Cl disolver en un matraz con 500 mL de agua destilada, adicionar 50 mL de NH3 concentrado y completar a 1L con agua destilada. 2. Etapa B: Preparación de la solución estándar Hacer diluciones: 1 x 10
-1, 1 x 10
-2, 1 x 10
-
3, 1 x 10
-4 NaCl en agua destilada,
previamente secar el NaCl a 105°C por 2h y dejar enfriar en desecador. Adicionar por cada 100 mL de solución, 2 mL de ISAS. 3. Etapa C: Preparar muestras a valorar de igual manera que la Etapa B
Tomar correctivos posibles.
Revisar posibles causas de desviación Anexo 2
FINAL
NO
¿La pendiente es la esperada: 59.16 ± 2-8%
mV/pH?
SI
Preparación de las soluciones estándar (SS) de calibración y
muestras
Conectar el electrodo al módulo de pH (izq) y ubicar el módulo de
unidades en mV
Sumergir el electrodo en las SS de < a >
concentración. Tomar el potencial.
Aceptar la calibración
Solicitar asesoría al proveedor, si ha extendido el tiempo de vida útil, gestionar compra
Realizar las determinaciones de
las muestras problema.
Lavar y acondicionar el electrodo.
Agite suavemente y lave el
electrodo con agua destilada
entre cada muestra.
Verificar información con el PEL

Anexo I UNIVERSIDAD
MANEJO DEL ELECTRODO DE SODIO NACIONAL DE CÓRDOBA
151
NORMA(S) COMPLEMENTARIA(S):
10.1 . Serie de Informes Técnicos de la OMS 823. Informe 32. Anexo 1 Prácticas adecuadas de
fabricación de productos farmacéuticos. Capítulo 13 y 14. Ginebra 1992
9. FORMATOS ANEXOS:
Número Anexo
Código Nombre Responsable del
Archivo Localización
Tipo de Archivo
1
Ficha técnica de mettler toledo
perfectionTM comb Na+.
Director de Subsidio LAB 212 Físico
Magnético
2 Precauciones y
Recomendaciones Director de Subsidio LAB 212
Físico
Magnético
ELABORADO POR REVISADO POR REVISIÓN ESTRUCTURAL POR
APROBADO POR
LILIANA ALARCÓN MARIA EUGENIA OLIVERA ALVARO KAIRUZ DANIEL ALLEMANDI Tesista Doctorado Profesor Encargado Profesor Encargado Director de unitefa
FIRMA:_________________
FECHA: FABIANA ALOVERO Profesor Encargado FIRMA: FECHA:
FIRMA:_________________
FIRMA:________________
FIRMA:________________
FECHA: FECHA: FECHA:

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
152

ANEXO II
BASES TEÓRICAS DE DICROÍSMO
CIRCULAR


Anexo II UNIVERSIDAD
BASES TEÓRICAS DICROÍSMO CIRCULAR NACIONAL DE CÓRDOBA
155
2.1 Actividad óptica
EL DC es una técnica espectropolarimétrica basada en dos fenómenos, la
rotación óptica y la elipticidad, que son dos formas de manifestación de una
misma propiedad física que presentan algunas sustancias: la actividad óptica.
En términos generales se dice que la actividad óptica es un
fenómeno físico por el que una sustancia hace rotar el plano de polarización de
un haz de luz plano-polarizada que la atraviesa.
La luz es una onda electromagnética consistente en un campo
eléctrico (E) y un campo magnético (B) oscilantes. Ambos son perpendiculares
entre si y perpendiculares a la dirección de propagación. El plano que contiene
el vector E se conoce como plano de polarización.
Luz (ondas) Linealmente-Polarizada
Si el vector del campo eléctrico oscila a lo largo de una línea recta, la luz
(ondas) se denomina plano-polarizada o linealmente polarizada. La luz
puede polarizarse en planos diferentes, en la figura 1 se muestra una onda
linealmente-polarizada en un plano horizontal vista a lo largo de la dirección de
propagación y frontalmente. En la figura 2 se observa una onda linealmente-
polarizada en un plano vertical.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
156
Figura 1. Luz linealmente-polarizada en plano horizontal.
Figura 2. Luz linealmente-polarizada en plano vertical.
Superposición de Ondas Linealmente-Polarizadas con igual Fase
Cuando se superponen dos ondas electromagnéticas linealmente-
polarizadas en dos planos perpendiculares sus campos eléctricos se suman
vectorialmente. Las propiedades de la onda electromagnética resultante
dependen de las intensidades y diferencia de las ondas componentes.
Anexo II UNIVERSIDAD
BASES TEÓRICAS DICROÍSMO CIRCULAR NACIONAL DE CÓRDOBA
157
En la figura 3 puede verse la superposición de dos ondas que tienen la
misma amplitud y longitud de onda, que están polarizadas en dos planos
perpendiculares y oscilan con la misma fase. Estar en fase significa que las dos
ondas alcanzan sus máximos (picos) y el cruce por el punto cero en el mismo
momento a lo lardo de su propagación. Las ondas componentes se visualizan
en rojo y verde, y la onda resultante en azul.
Como se puede observar, el resultado de esta superposición es otra onda
linealmente-polarizada, su plano de polarización forma un ángulo de 45° con
los planos de polarización de las ondas componentes.
Figura 3. Superposición de dos ondas linealmente-polarizadas que
tienen la misma amplitud y longitud de onda y oscilan con la misma
fase.
Superposición de Ondas Linealmente-Polarizadas fuera de Fase
Cuando dos ondas linealmente-polarizadas (en dos planos
perpendiculares) se superponen estando fuera de fase, la onda que resulta de

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
158
dicha superposición no es una onda linealmente-polarizada o plano-polarizada.
En la figura 4 se presenta la superposición de dos ondas que tienen la
misma amplitud y longitud de onda, están polarizadas en dos planos
perpendiculares, pero tienen una diferencia de fase de 90°. Una diferencia de
fase de 90° significa que cuando una onda está en su máximo respecto a la
línea de propagación, la otra atraviesa el punto cero. Las ondas componentes
se visualizan en color rojo y verde y la resultante en color azul.
Figura 4. Superposición de dos ondas linealmente-polarizadas que
tienen la misma amplitud y longitud de onda y oscilan con una
diferencia de fase de 90°.
El resultado de esta superposición es una onda electromagnética
especial. En todos los puntos a lo largo de la línea de propagación, el vector
campo eléctrico rota en círculo y su longitud se mantiene constante. Dicha onda
se denomina luz (onda) circularmente-polarizada. Como se observa en la
figura 4, la onda se propaga describiendo una espiral en lugar de describir una
curva sinusoidal como la observada en la figura 3 (azul).

Anexo II UNIVERSIDAD
BASES TEÓRICAS DICROÍSMO CIRCULAR NACIONAL DE CÓRDOBA
159
Si las ondas que se superponen tienen una diferencia de fase de -90°
(desplazadas una respecto a la otra en ¾ de longitud de onda) la onda
resultante también será circularmente-polarizada, figura 5.
Una diferencia de fase de 90° genera una onda de luz circularmente-
polarizada que gira en sentido horario y una diferencia de -90° una onda que
gira en sentido anti horario. Debido a ello es necesario distinguirlas y se les
denomina luz circularmente-polarizada derecha y luz circularmente-
polarizada izquierda.
Figura 5. Superposición de dos ondas linealmente-polarizadas que
tienen la misma amplitud y longitud de onda y oscilan con una
diferencia de fase de -90°.
Superposición de Ondas Circularmente-Polarizadas
Anteriormente se describió lo que ocurre cuando se superponen dos ondas
linealmente-polarizadas. Dos ondas circularmente-polarizadas también pueden
encontrarse y superponerse, la onda resultante es aquella que se obtiene a
partir de la suma vectorial del campo eléctrico de las ondas componentes.
UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
160
Cuando se superpone una onda de luz circularmente-polarizada derecha
con una circularmente-polarizada izquierda (ambas con igual amplitud y
longitud de onda) la onda resultante es una onda linealmente-polarizada,
figura 6.
Una conclusión importante que se extrae de esto es: la luz linealmente-
polarizada puede obtenerse por superposición de una onda circularmente-
polarizada derecha y una circularmente-polarizada izquierda cuyas amplitudes
sean idénticas.
Figura 6. Superposición de dos ondas circularmente-polarizadas que
tienen la misma amplitud y longitud de onda.
Interacción de la Luz con la Materia
Cuando la luz interacciona con la materia sus propiedades cambian,
intensidad (amplitud), polarización, velocidad, longitud. Los dos fenómenos más
importantes que se producen como consecuencia de la interacción son la
absorción y disminución de su velocidad.

Anexo II UNIVERSIDAD
BASES TEÓRICAS DICROÍSMO CIRCULAR NACIONAL DE CÓRDOBA
161
Luz Linealmente-Polarizada en un Medio Absorbente
En la figura 7 se observa lo que ocurre cuando una onda lienalmente-
polarizada ingresa a un medio que absorbe luz y no refracta (índice de
refracción n=1). Cuando la onda sale del material el vector campo eléctrico
oscila de la misma manera que cuando ingresó, pero su amplitud es menor que
la inicial.
Figura 7. Interacción de luz linealmente-polarizada con un material
que absorbe (n=1).
Lo mismo ocurre cuando luz circularmente-polarizada interacciona con un
medio absorbente. Cuando el vector campo eléctrico abandona el medio rota
igual que antes de entrar, pero su longitud es menor figura 8.
Figura 8. Interacción de luz circularmente-polarizada con un material
que absorbe (n=1).

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
162
Luz Circularmente-Polarizada en un Medio que Produce Dicroísmo
Circular
Algunas moléculas poseen una propiedad particular: absorben luz
circularmente-polarizada derecha en diferente magnitud que luz circularmente-
polarizada izquierda. Este fenómeno se denomina Dicroísmo Circular (DC). Se
observa por ejemplo en compuestos quirales, compuestos que presentan algún
tipo de asimetría (como los atropoisómeros o el ADN) o bien puede ser inducido
por ejemplo por un solvente quiral o un receptor quiral.
Como se demostró anteriormente la luz linealmente-polarizada puede
obtenerse por superposición de ondas circularmente-polarizadas derecha e
izquierda.
Si la luz linealmente-polarizada atraviesa un medio que presenta DC sus
propiedades van a cambiar porque el medio absorbe las dos componentes
circularmente-polarizadas en diferente extensión. En la figura 8.9 se muestra lo
que ocurre cuando una onda de luz linealmente-polarizada (el plano de
polarización mostrado es vertical) atraviesa un medio que no absorbe la
componente circular izquierda (roja) y absorbe la componente circular derecha
(verde).
Debido a que la componente izquierda (roja) no es absorbida, atraviesa
el medio sin cambios. La intensidad de la componente derecha (verde)
disminuye respecto al valor inicial debido a la absorción. La superposición de las
dos componentes a lo largo de la línea de propagación no es más una onda
linealmente-polarizada. El vector campo eléctrico resultante (azul) no oscila más
a lo largo de una recta si no que rota describiendo una elipse. Esta onda se
denomina luz elípticamente-polarizada.

Anexo II UNIVERSIDAD
BASES TEÓRICAS DICROÍSMO CIRCULAR NACIONAL DE CÓRDOBA
163
Figura 8. Interacción de luz linealmente-polarizada con un material
que absorbe (n=1) y presenta dicroísmo circular.
El DC hace que la luz linealmente-polarizada se transforme en luz
elípticamente-polarizada.
Un parámetro que se define en este tipo de polarización es la
Elipticidad (θ), que es igual al arco de la tangente de la relación entre los
semiejes de la elipse siendo b, el semieje menor y a el mayor, figura 9 y
ecuación 1.
θ = arctg 𝐛
𝐚 ΔA Ecuación 1

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
164
Figura 9. Representación gráfica de la elipticidad (θ) y rotación (α) de
la luz linealmente-polarizada, al emerger de un medio ópticamente
activo, en la región de las longitudes de onda de una banda de
absorción.
Cuando un material absorbe luz circularmente-polarizada derecha en
diferente magnitud que la luz circularmente-polarizada izquierda los coeficientes
de extinción derecho (ƐR) e izquierdo (ƐL) serán diferentes, lo que conduce a:
ΔƐ = ƐL-ƐR que representa al fenómeno de dicroísmo circular. En los
espectropolarímetros se registran cambios de absorbancia en función de la
longitud de onda, ecuación 2.
El cambio de absorbancia (ΔA) puede expresarse como elipticidad empleando la
ecuación 3.

Anexo II UNIVERSIDAD
BASES TEÓRICAS DICROÍSMO CIRCULAR NACIONAL DE CÓRDOBA
165
𝚫Ɛ = 𝐈
𝐜𝐥 ΔA Ecuación 2
θ = 33 ΔA Ecuación 3
Los valores de elipticidad y de rotación óptica dependen de la longitud de
onda de la radiación que interacciona con la materia. Cuando se grafica la
elipticidad (θ) en función de la longitud de onda se obtiene un espectro de DC;
si se grafica el ángulo con el que se desvía el plano de la luz linealmente-
polarizada (α) en función de la longitud de onda se obtiene una curva de
dispersión rotatoria óptica (DRO); El primer fenómeno es de tipo absortibo en
tanto que el segundo es dispersivo.
A los siguientes efectos: DC y elipticidad se los conoce como efecto
Cotton.
Para un compuesto que presenta actividad óptica pero que no contiene
ningún cromóforo, esto es, no absorbe luz a la longitud de onda empleada en la
determinación, la rotación óptica decrece en valor absoluto cuando aumenta la
longitud de onda. De esta manera se obtienen las curvas plano-positivas o
plano-negativas. Una curva plana es positiva si la rotación óptica aumenta al
disminuir la longitud de onda. En el caso contrario es negativa.
Si una sustancia además de ser ópticamente activa presenta absorción
en la zona de longitudes de onda empleada, se obtiene curvas con efecto
Cotton.
En conclusión, para obtener espectros de DC un compuesto debe
presentar la característica de ser quiral y poseer un cromóforo que presente
transiciones electrónicas cuyas energías se encuentren en el intervalo de 190
nm a 800 nm, que es la zona accesible a los espectropolarímetros
convencionales.

UNIVERSIDAD Q.Farm. Liliana P. Alarcón NACIONAL DE CÓRDOBA
166