
Construcción de aljibe con placas de cemento en Gancedo, Chaco ...

Capítulo 6: Hidratos de CarbonoNelio E. Bazán, Emilse Paccotti y María E. Gancedo
Los hidratos de carbono son la principal fuente de energía para el organismo durante el ejercicio intenso. El nombre de glúcido deriva del vocablo griego glykys que significa dulce, aunque solamente lo son algunos monosacáridos y disacáridos. Podemos resumir las funciones de los hidratos de carbono (HC), en los siguientes puntos:
Aportan energía a corto plazo proporcionando aproximadamente 4 Kcal. por gramo.
Esta energía puede almacenarse en forma de glucógeno hepático y muscular o transformarse en grasa y utilizarse cuando el cuerpo necesite energía.
La glucosa constituye la única fuente energética del sistema nervioso y de las células sanguíneas.
La ribosa y la desoxirribosa participan en la síntesis de material genético (ADN, ARN) y otros compuestos.
Forman parte de proteínas, glicoproteínas, en el tejido cartilaginoso. En la membrana celular algunos azúcares como fucosa, manosa y ácido siálico
están relacionados con fenómenos de reconocimiento celular.
Están compuestos por átomos de carbono, oxígeno e hidrógeno, es decir por carbono y agua, es por ello que también se los llama carbohidratos aunque a veces tienen N, S, o P. La mayor parte de los carbohidratos se degradan a sus monosacáridos constituyentes durante la digestión. Las moléculas de almidón se degradan parcialmente convirtiéndose en cadenas más pequeñas denominadas dextrinas. Según el número de moléculas que poseen, se pueden clasificar en monosacáridos (de absorción más rápida), disacáridos o polisacáridos (de absorción más lenta). Los alimentos fuente de hidratos de carbono se pueden dividir en diversos grupos. Uno de ellos está compuesto por azúcar, que es el nombre común de la sacarosa pura, y los productos elaborados como dulces, mermeladas y frutas confitadas. La fructosa está presente en grandes cantidades en frutas y en miel, siendo el más dulce de los azúcares sencillos. El almidón es la forma más familiar del polisacárido vegetal y sirve como un almacén de energía para su uso por las plantas. Se encuentra en los granos de los diversos cereales, con los que se hacen el pan y las pastas, en las chauchas, las papas y algunas raíces. Las legumbres, además de proporcionar hidratos de carbono digeribles contienen fibras y proteínas. Los frutos secos contienen un porcentaje algo menor de hidratos de carbono. La lactosa, un hidrato de carbono de origen animal, se encuentra de forma natural sólo en la leche siendo el menos dulce de los disacáridos.

MONOSACARIDOS
Están compuestos por una molécula que posee de 3 a 8 carbonos, son sólidos, incoloros, dulces, cristalinos y solubles en agua. Entre los monosacáridos, los más importantes son las pentosas (como ejemplo la ribosa) y las hexosas. Entre ellas se encuentran la glucosa, galactosa, manosa y fructosa. Forman parte de los ácidos nucleicos, de las coenzimas y del principal compuesto que otorga energía: el ATP (la ribosa forma parte del nucleótido). Algunos antibióticos como la eritromicina contienen amino azúcares. Químicamente son polihidroxialdehídos o polihidroxicetonas, responden a la fórmula (CH2O)n, en la que n tiene un valor igual o mayor que 3, aunque los más frecuentes son los de 5 y 6 átomos de carbono. Presentan en uno de sus carbonos un grupo carbonilo y el resto grupos hidroxilos. Si el grupo carbonilo se encuentra al final de la cadena, el monosacárido es un aldehído (aldosa) y una cetona si se encuentra en un carbono secundario (cetosa).
Tabla 6.00: Monosacáridos.
La glucosa es el monosacárido más conocido y es la principal fuente de energía para el sistema nervioso central y músculos. Cuando se ingieren hidratos de carbono a través de los alimentos, las enzimas del aparato digestivo atacan las moléculas de polisacáridos y liberan los diferentes monosacáridos dando por resultado principalmente glucosa. Esta glucosa es transportada por el torrente sanguíneo hacia el hígado donde puede seguir varios caminos: conversión a grasas, sostén de la glucemia o almacenamiento en forma

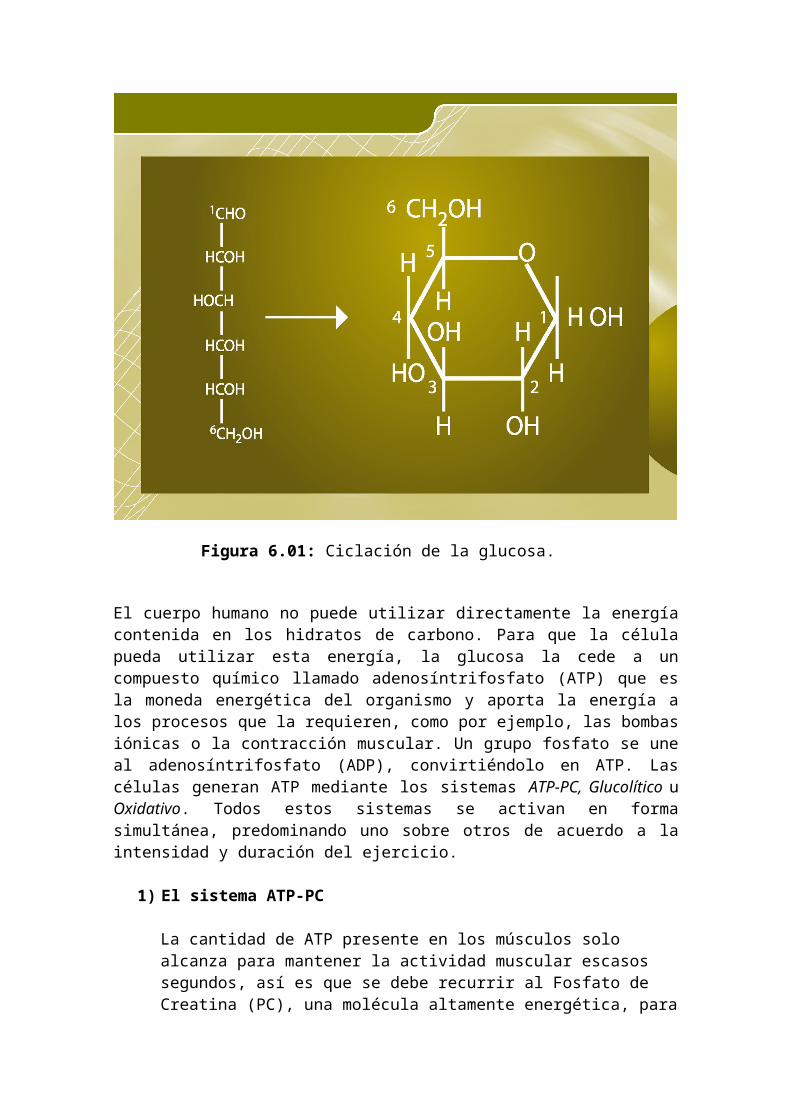
de glucógeno hepático o muscular. Puede presentar una estructura cíclica al reaccionar los grupos aldehídos (cetonas en otros monosacáridos) con un hidroxilo de la misma molécula convirtiéndola en anillo. Si el aldehído reacciona con el -OH se forma un hemiacetal.
Figura 6.01: Ciclación de la glucosa.
El cuerpo humano no puede utilizar directamente la energía contenida en los hidratos de carbono. Para que la célula pueda utilizar esta energía, la glucosa la cede a un compuesto químico llamado adenosíntrifosfato (ATP) que es la moneda energética del organismo y aporta la energía a los procesos que la requieren, como por ejemplo, las bombas iónicas o la contracción muscular. Un grupo fosfato se une al adenosíntrifosfato (ADP), convirtiéndolo en ATP. Las células generan ATP mediante los sistemas ATP-PC, Glucolítico u Oxidativo. Todos estos sistemas se activan en forma simultánea, predominando uno sobre otros de acuerdo a la intensidad y duración del ejercicio.
1) El sistema ATP-PC
La cantidad de ATP presente en los músculos solo alcanza para mantener la actividad muscular escasos segundos, así es que se debe recurrir al Fosfato de Creatina (PC), una molécula altamente energética, para sintetizar ATP. El ADP resultante de la utilización del ATP se combina con el ión fosfato de alta energía provisto por la creatina, manteniendo constante el suministro de ATP. Con el aporte de este sistema se pueden sostener las demandas energéticas por unos 10 a 12 segundos, utilizándose para períodos breves de potencia máxima como en
una carrera de 100 metros, un salto o en el levantamiento de pesas. Más allá de este tiempo comienza a predominar otro sistema.
2) Sistema glucolítico
Para que la glucosa pueda ingresar en la vía energética debe convertirse en glucosa-6-fosfato que es finalmente degradada a dos moléculas de ácido pirúvico. En este proceso se produce una ganancia neta de 3 moles de ATP por cada mol de glucógeno descompuesto. Si el compuesto inicial es glucosa en lugar de glucógeno la ganancia neta es de 2 moles ya que se utiliza un ATP para convertirla en glucosa-6-fosfato. La glucólisis produce una acumulación de ácido láctico que puede dosarse en sangre a los pocos minutos de iniciado el ejercicio, ya que sale de las células musculares hasta el líquido intersticial y la sangre. El sistema glucógeno - ácido láctico puede sostener la actividad muscular hasta 1.6 minutos como sucede en carreras de 200 a 500 m o en natación en 100 m.
3) Sistema oxidativo
El ácido pirúvico en presencia de oxígeno es transformado en acetil coenzima A (acetil CoA) ingresando al ciclo del Ácido Cítrico (Krebs) con separación de átomos de hidrógeno captados por las coenzimas NAD y FAD siendo conducidos hacia la cadena de transporte de electrones donde se producen 39 moléculas de ATP.
La glucosa absorbida por la mucosa intestinal, llega a través del torrente sanguíneo a las células del organismo para satisfacer sus necesidades energéticas; la cantidad restante, que no es empleada por el metabolismo celular, y por lo tanto no es necesario en ese momento, se deposita en el hígado y en el tejido muscular en forma de glucógeno, lo cual se lleva a cabo mediante el procedimiento metabólico conocido como glucógeno génesis. Este polímero de HC puede llegar a representar aproximadamente el 5% del peso total del hígado, siendo la capacidad total de almacenamiento de glucógeno por parte de la víscera hepática de alrededor de 70g.El glucógeno es un polisacárido formado por una gran cantidad de moléculas de glucosa, el cual, cada vez que el organismo lo demande, sé ira desdoblando liberando moléculas de glucosa a fin de poder ser utilizadas en la obtención de energía. Esto se realiza a través de un mecanismo contrario a la glucogénesis, denominado glucógeno lisis. Estos dos procesos, que interactúan permanentemente para mantener equilibradas las cifras de glucosa en la sangre, están mediados por una serie de hormonas, tales como la insulina, el glucagon, ciertas catecolaminas y la hormona de crecimiento.Cuando en el organismo se produce un exceso de glucosa que no es utilizada para las funciones energéticas celulares, ni se almacena en forma de glucógeno hepático o muscular, va a sufrir una serie de procesos metabólicos mediante los cuales se convertirá en grasa, la cual va a ser transportada a través de la sangre hacia el tejido adiposo, lugar en el que se almacenará en forma de reserva energética para una utilización posterior.

La glucosa debe encontrarse de forma permanente en la sangre en una concentración, la glucemia, que ha de oscilar entre unos márgenes relativamente estrechos: 70 y 110mg por cada 100 cc de sangre, en condiciones basales del individuo.
Transportadores de glucosa a través de la membrana
Los monosacáridos sólo atraviesan las membranas celulares ayudados por proteínas específicas de transporte. Este paso lo pueden realizar contra gradiente (en enterocitos y algunas células renales) o a favor de gradiente (difusión facilitada). El movimiento de difusión facilitada tiene como característica la activación inmediata del glúcido para asegurarse que el gradiente siempre sea favorable a la entrada. Existen dos sistemas de transporte de glucosa y de otros monosacáridos: los transportadores de sodio y glucosa llamados SGLT (sodium-glucose transporters) y los transportadores de glucosa llamados GLUT (glucosa transporters). Los transportadores SGLT son proteínas con una estructura secundaria de catorce dominios que efectúan un transporte acoplado sodio y glucosa, o galactosa, localizadas en la membrana luminal de las células epiteliales encargadas de la absorción (intestino delgado) y la reabsorción (riñón, túbulo contorneado proximal) de nutrientes. Aprovechan el ingreso de sodio a favor del gradiente electroquímico, entre el exterior y el interior de la célula, para transportar la glucosa en contra de un gradiente químico. Se han identificado tres transportadores SGLT que se diferencian en la afinidad por la glucosa y el sodio, la capacidad para transportar glucosa o galactosa y la ubicación tisular. El SGLT 1 posee una alta afinidad por la glucosa, y transporta dos moléculas de sodio por una de glucosa o galactosa, y 260 moléculas de agua. Se encuentra en intestino y en el segmento S3 de la nefrona proximal. El SGLT 2 transporta una molécula de sodio por una de glucosa; en los segmentos S1 y S2 reabsorbiendo el 90% de la glucosa filtrada por el riñón. El SGLT 3 transporta dos moléculas de sodio por una de glucosa. Los GluT son glicoproteínas de 45 a 55 kDa con doce dominios transmembrana en estructura α-hélice. Transportan glúcidos por difusión facilitada presentando por lo menos dos conformaciones intercambiables inducidas por el sustrato. El receptor capta la presencia de glucosa y la proteína toma a la molécula para ingresar en el canal. Inmediatamente se produce un intercambio de conformación de la proteína transportadora y el canal se abre hacia el lado opuesto. Dado que la fuerza de unión del receptor es débil, el movimiento térmico de la molécula unida hace que este se libere. Los GluT se encuentran en trece formas posibles, numerados del 1 al 13. El GluT4 es una proteína de 509 aminoácidos, codificada por un gen ubicado en el cromosoma 17 y posee una Km para la glucosa de 5 mM. Está expresada en tejido muscular esquelético y cardíaco y en tejido adiposo. Tienen gran afinidad por el sustrato, pero el 90 % se encuentra en vesículas intracelulares que están sometidas a un ciclo continuo de exocitosis-endocitosis. Cuando la insulina se une al receptor se induce un cambio conformacional en éste, que estimula la actividad tirosina-quinasa. El receptor activado se autofosforila y, a su vez, fosforila proteínas en secuencia como las IRS (Insuline receptor substrate) que desencadenan una cascada de eventos moleculares, incluyendo, entre otros, la translocación de las vesículas. Esta exocitosis de las vesículas incrementa el número de GluT 4 en la membrana del miocito o del adipocito y la entrada de glucosa. Cuando el estímulo cesa se produce la endocitosis, con formación

de trisqueliones de clatrina y la participación del citoesqueleto celular. El calcio utilizado para la contracción muscular produce también la movilización de las vesículas y la incorporación de GluT4 a la membrana en forma independiente de la insulina. Los valores basales de GluT4 se recuperan luego de dos horas de reposo. Por otro lado, el GluT 12 es una proteína de 617 aminoácidos y se considera un segundo sistema de transporte mediado por insulina. Una característica de este tipo de transporte es su saturación. La cantidad de GluT4
presentes en la membrana celular es la que va a determinar el ritmo de entrada de la glucosa al músculo esquelético. Como se explicó anteriormente, la migración de las vesículas que contienen GluT4 hacia la membrana está regulada por la insulina y el calcio. Ante un aumento de glucemia (por ejemplo, luego de una ingesta de alimentos) la insulina es liberada por las células β del páncreas. La insulina circulante se une a receptores de membrana, quienes darán la señal para la migración y expresión de GluT4. De esta manera la glucosa es captada por la célula y sacada de la circulación, volviendo la glucemia a valores normales. Durante la actividad física, el músculo esquelético necesita incorporar glucosa para metabolizarla y así producir energía. El calcio utilizado en el proceso de contracción muscular (se une a la subunidad C de la troponina para desdoblarla y exponer los sitios activos de la actina) también sirve como estímulo para la expresión de GluT4. De esto se pueden inferir dos puntos importantes: la prescripción de actividad física a personas con problemas de hiperglucemia (por ejemplo, diabéticos) y lo fundamental de ingerir hidratos de carbono antes y durante la actividad para evitar hipoglucemias. La cantidad de transportadores GluT4 es directamente proporcional a la cantidad de actividad física realizada. La vida media de los GluT4 es corta, por lo que su expresión puede variar dependiendo del período de actividad de cada persona, pero puede estar entre las 18 y 24 horas. Esto explica la necesidad de realizar actividad física diariamente.

Figura 6.02: Translocación de transportadores GluT a la membrana

Tabla 6.03: Transportadores de monosacáridos.
OLIGOSACARIDOS
Están formados por la unión de 2 a 20 monosacáridos, límite establecido arbitrariamente, pudiendo encontrarse en forma lineal o ramificada. La unión de los monosacáridos tiene lugar mediante enlaces glicosídicos. Los compuestos con grupos OH, NH2 y SH pueden reaccionar con el OH de un monosacárido, con pérdida de una molécula de agua para formar los compuestos llamados glicósidos. Este es el enlace glicosídico susceptible a la hidrólisis ácida y a la acción de las enzimas llamadas

glicosidasas. Se distinguen O-glicósidos (a partir de un OH), N-glicósidos (a partir de un NH2) y S-glicósidos (a partir de un SH).
Figura 6.04: El enlace O-glicosídico. Realizado a partir de un OH.
El grupo que reacciona con el monosacárido recibe el nombre genérico de aglicona, entonces un glicósido es un monosacárido unido a una aglicona por medio de un enlace glicosídico y si la aglicona es otro monosacárido, se trata de un holósido o disacárido, y heterósido si es un compuesto distinto. En los disacáridos reductores el carbono anomérico de un monosacárido reacciona con un OH alcohólico de otro, entonces el segundo azúcar presenta libre su carbono anomérico, y seguirá teniendo propiedades reductoras, y podrá presentar el fenómeno de la mutarrotación. A este grupo pertenecen la maltosa, la isomaltosa, la celobiosa y la lactosa: La maltosa está formada por dos glucosas unidas por el OH del C1 en posición a de una y el OH del C4 de otra y se obtiene a partir de la hidrólisis del almidón (polisacárido de reserva en vegetales). La isomaltosa también está formada por dos glucosas, y difiere de la anterior en que el enlace glicosídico se forma entre el OH del C1 en posición a de una y el OH del C6 de la otra. La celobiosa se obtiene a partir de la hidrólisis de la celulosa. La lactosa está formada por glucosa y galactosa, donde el OH del C1 en posición de la galactosa está unido al OH del C4 de la glucosa. Este azúcar se encuentra en la leche.En los disacárido no reductores el carbono anomérico de un monosacárido reacciona con el carbono anomérico del otro monosacárido, así no queda ningún carbono anomérico libre y tampoco podrá presentar mutarrotación. A este grupo pertenecen los disacáridos sacarosa y trehalosa: La sacarosa es el azúcar de caña y es una forma de reserva de muchas plantas. Se encuentra también en el néctar de las flores, siendo entonces un componente básico para la elaboración de la miel. Está formada por glucosa y fructosa unidas por sus carbonos anoméricos.
Los nombres genéricos indican el número de monosacáridos que intervienen en la unión. Son importantes los disacáridos y los trisacáridos. Los más abundantes son los disacáridos que están formados por dos monosacáridos, iguales o distintos, entre ellos

encontramos la maltosa (glucosa y glucosa), sacarosa (glucosa y fructosa) y la lactosa (glucosa y galactosa). A la unión de tres monosacáridos se la denomina trisacáridos. La rafinosa (fructosa, glucosa y galactosa) y la maltotriosa forman parte de los trisacáridos. Este último, muy utilizado por maratonistas.
Figura 6.05: Lactosa.
Figura 6.06: Sacarosa.

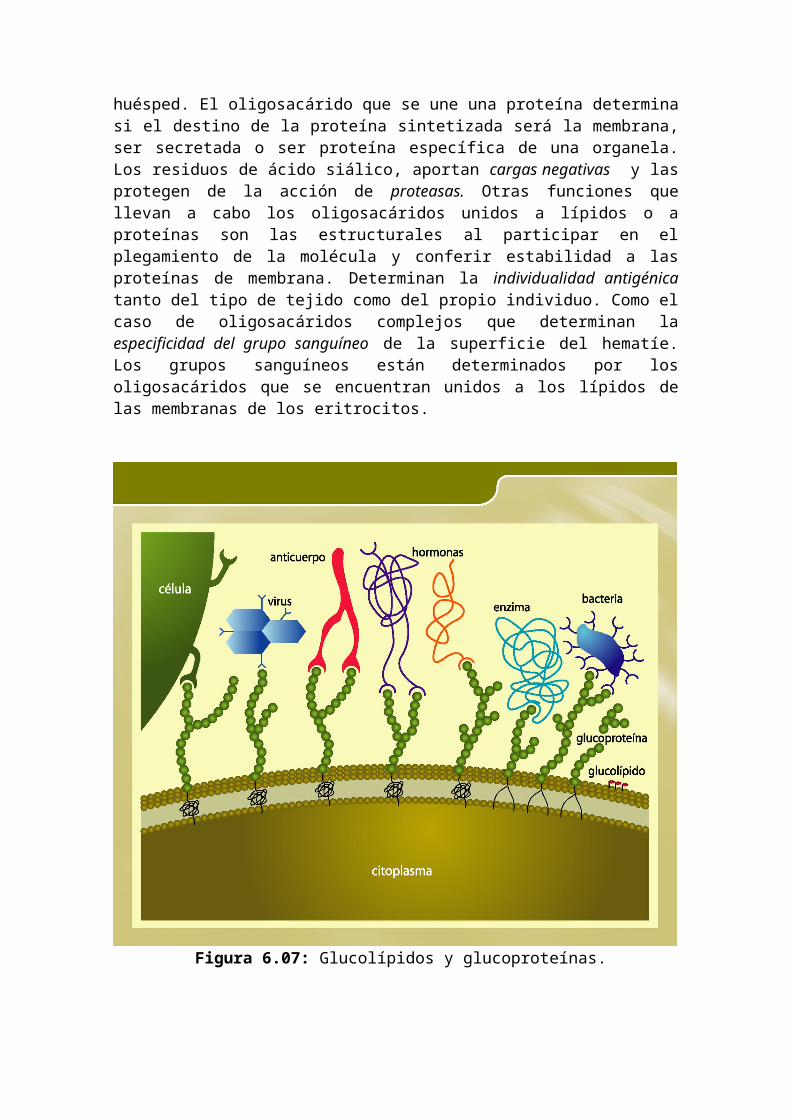
Los oligosacáridos son parte integrante de los glucolípidos y glucoproteínas que se encuentran en la superficie externa de la membrana plasmática y tienen una gran importancia en las funciones de reconocimiento en superficie como en la interacción de ciertos bacterias y virus con las células huésped. El oligosacárido que se une una proteína determina si el destino de la proteína sintetizada será la membrana, ser secretada o ser proteína específica de una organela. Los residuos de ácido siálico, aportan cargas negativas y las protegen de la acción de proteasas. Otras funciones que llevan a cabo los oligosacáridos unidos a lípidos o a proteínas son las estructurales al participar en el plegamiento de la molécula y conferir estabilidad a las proteínas de membrana. Determinan la individualidad antigénica tanto del tipo de tejido como del propio individuo. Como el caso de oligosacáridos complejos que determinan la especificidad del grupo sanguíneo de la superficie del hematíe. Los grupos sanguíneos están determinados por los oligosacáridos que se encuentran unidos a los lípidos de las membranas de los eritrocitos.
Figura 6.07: Glucolípidos y glucoproteínas.

POLISACARIDOS
Están compuestos por numerosas unidades de monosacáridos. Algunos de ellos son polímeros de un solo tipo de monosacáridos (homopolisacaridos), mientras que otros dan más de una clase de monosacáridos (heteropolisacáridos). Se los puede clasificar también en simples y complejos. El almidón, un homopolímero, cumple el papel de reserva de nutrientes en los vegetales. Se deposita en las células formando gránulos cuya forma y tamaño varían según el vegetal de origen. Constituye la fuente más importante de HC en los vegetales y es el principal hidrato de carbono de la alimentación humana. Es abundante en los cereales, tubérculos como papa y batata y legumbres. Formado por cadenas helicoidales, la 12nterna, y cadenas ramificadas, la amilopectina. Los glucosaminoglucanos o mucopolisacáridos están constituidos por HC complejos y ácidos urónicos. Si se encuentran unidos a una proteína componen el proteoglucano. Son importantes componentes del tejido conectivo como el acido hialurónico, el ácido condrointilsulfúrico, cemento de contacto de los cartílagos y los huesos y la heparina, un anticoagulante por excelencia. Las glucoproteínas o mucoproteínas son proteínas que contienen hidratos de carbono y están presentes en líquidos corporales y diversos tejidos. Por otro lado, los HC también están presentes en estructuras lipídicas como las membranas celulares o en algunas lipoproteínas como las LDL (low-density-protein).
Glucógeno
El glucógeno que también forma parte de los polisacáridos simples, es la unión de más de 30.000 moléculas de glucosa en forma muy ramificada y cuya característica es la de ser la principal reserva de energía en células animales. Es un polímero muy ramificado y de gran peso molecular siendo el hígado y el músculo los tejidos más ricos en glucógeno. Está formado por moléculas de glucosa unidas por medio de enlaces glicosídicos 4 que son lineales y ramificaciones unidas por enlaces 6, con un extremo reductor y otro no reductor. Esta estructura posee ciertas ventajas como el poseer muchos extremos no reductores a la vista y ser más soluble. Forma gránulos en el citosol en los que se encuentran las enzimas que lo metabolizan.
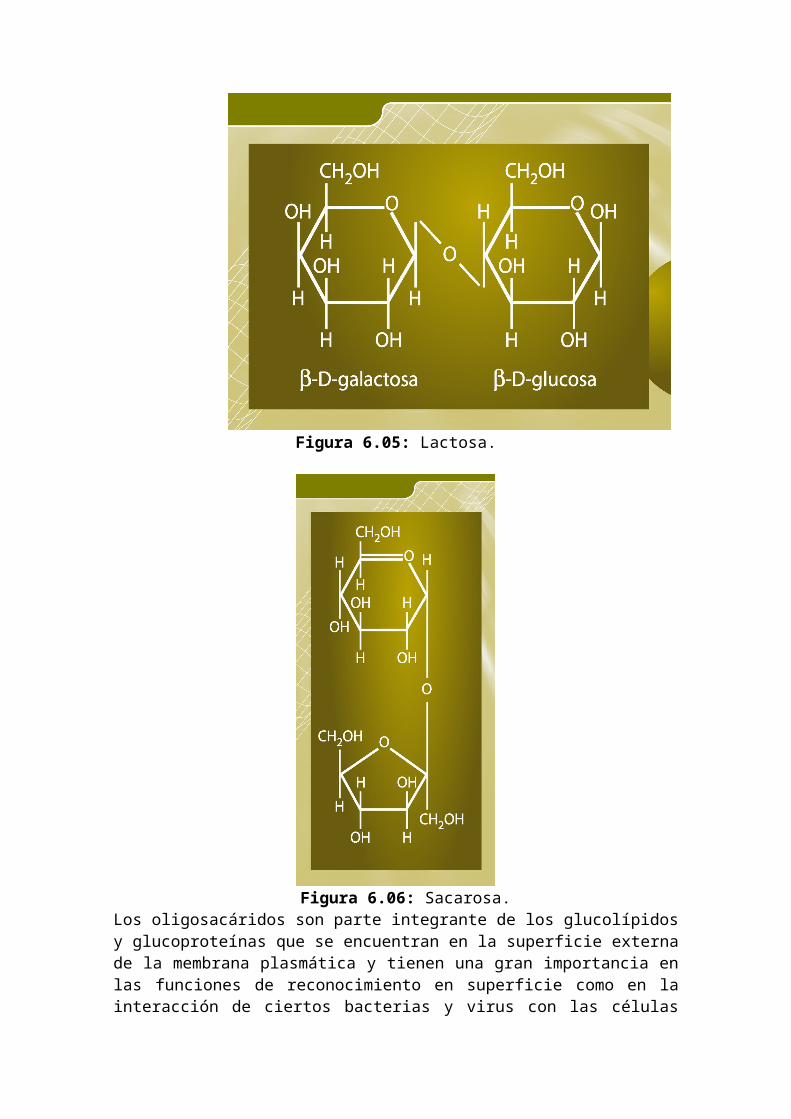
Figura 6.08: Glucógeno.
El contenido de glucógeno en el hígado es de aproximadamente 70 a 100 g, que proveen de 280 a 400 Kcal, variando en función de la cantidad de glucógeno que se degrade para sostener la glucemia y de la cantidad de glucosa que pueden aportar los alimentos. La reserva hepática disminuye especialmente durante la noche, y durante el ejercicio físico, por aumento de la captación de glucosa sanguínea por parte de los músculos que trabajan. El glucógeno hepático constituye el factor clave para el mantenimiento de un nivel de glucosa en sangre normal durante el ejercicio. Al agotarse los depósitos de glucógeno del hígado se producirá la utilización de proteínas y una movilización de grasas. Un individuo con dieta convencional posee una concentración de 15 a 18 g de glucógeno por kg de tejido muscular, lo que supone una reserva total de 350 g hasta 550 g en sujetos muy entrenados mediante la combinación de ejercicio y una dieta rica en
HC. Los HC totales almacenados en el tejido muscular alcanzan, por lo tanto, una equivalencia energética que oscila entre las 1.200 y las 2.000 kcal. El glucógeno muscular se forma con características bifásicas. Existe una fase rápida, 14nternac independiente, que ocurre las primeras 24 hs seguida por una fase lenta, 14nternac dependiente, que dura varios días. El ejercicio y la insulina estimulan la proteína transportadora GLUT4 en la membrana del músculo aumentando la captación de glucosa, esta hormona es también importante en la síntesis de glucógeno al actuar sobre la enzima 14nternaciona. La proteína glucogenina es la enzima iniciadora del polímero glucógeno, de 37 kDa, se glicosila sobre un residuo específico de tirosina, así una glucosa se une a los OH de Ser y Thr formando un enlace glicosídico. En el músculo la 14nternacio permanece adherida en el centro de la molécula de glucógeno. Esta proteína actúa como núcleo de la molécula de glucógeno (primer) y a la vez como una enzima estimuladora de la glicosilación. La acumulación inicial de glucosa a la 14nternacio forma un tipo de glucógeno llamado pro-glucógeno, relativamente pequeño, y su almacenamiento es sensible a la provisión de hidratos de carbono de la dieta. Durante la segunda fase se produce principalmente el almacenamiento de macroglucógeno, molécula con mayor cantidad de glucosa para cada núcleo de 14nternacio. Cuando la reserva de proglucógeno ha alcanzado un límite crítico en un medio de hidratos de carbono favorable durante 2 o 3 días de ingesta, una porción es sintetizada en macroglucógeno. El extremo no reductor incorpora glucosa activada combinada con un nucleótido derivado de la uridina, UTP). La activación implica el pasaje de glucosa a glucosa – 6P, catalizada por la hexoquinasa (en el hígado por la glucokinasa), y luego la transformación a glucosa – 1P catalizado por la fosfoglucomutasa. Luego G1P + UTP → UDP-glucosa + Ppi, catalizado por la UDP glucosapirofosforilasa. La enzima glucogenosintetasa agrega esa glucosa formando (Glu)n+1, liberando UDP. Cuando la cadena se ha alargado unos 11 residuos de glucosa, la enzima ramificante ( 1→4 a 1→6 amilotransglucosidasa) toma un grupo de unos 7 monómeros y lo transfiere entero en posición 6 a una glucosa ya unida. La separación entre ramas debe ser de por lo menos 4 glucosas para permitir luego la degradación. La glucógeno sintasa siempre necesita cebador (extremo no reductor) que puede ser el glucógeno sin degradar o la glucogenina. La regulación de la glucogénesis implica control alostérico, llevado a cabo por metabolitos y por enzimas bajo control hormonal. El glucógeno se encuentra en el citosol junto a las enzimas encargadas de su degradación y síntesis que tienen una regulación contrapuesta. La síntesis implica la activación de la glucogenosintasa frente a abundancia de glucosa y la degradación ocurre al bajar dicho nivel.El punto de regulación de la glucogenogénesis muscular es la glucógeno sintasa, que existe en dos estados conformacionales diferentes: sintasa B (muy poco activa, fosforilada) y sintasa A (muy activa, desfosforilada). Es un homotratrámero que posee nueve sitios de fosforilación y cuanto más fosforilada está, menos activa es. En la regulación por modificación covalente la inactivación por fosforilación se produce cuando la enzima fosforilasa quinasa activa la glucógeno fosforilasa por fosforilación inactivando la glucógeno sintasa. La activación se produce cuando las fosfatasas desfosforilan a la glucógeno fosforilasa y la glucógeno sintasa, que de la forma B, fosforilada e inactiva, pasa a la A, desfosforilada y activa. La glucógeno sintasa es activada también por la presencia de insulina. La adrenalina y la insulina son antagonistas en las células musculares, ya que la insulina disminuye los niveles de AMPc, compuesto intermediario a través del que actúan diversas hormonas, elevados

por la adrenalina o glucagon (en hígado y músculo esquelético) al estimular una fosfodiesterasa. En el hígado la fosfoproteína fosfatasa-I (PP1), es la encargada de desfosforilar el sistema, uniéndose en principio fuertemente a la glucógeno fosforilasa-A cuando sobra glucosa, luego ésta se libera y actúa sobre la glucógeno sintasa, desfosforilándola y activándola. Por eso cuando se suministra glucosa, la actividad de la glucógeno fosforilasa-A hepática disminuye rápidamente y, luego de un tiempo de latencia aumenta rápidamente la actividad glucógeno sintasa.
La glucogenolisis se realiza en en presencia de glucógenofosforilasa y se obtiene glucosa ya fosforilada: (Glu)n + Pi G1P + (Glu)n-1. El glucógeno se degrada por el extremo reductor rompiendo enlaces glicosídicos 4 hasta cerca de cuatro residuos de la ramificación. Allí se necesita otro enzima, la transferasa α→4, que transfiere los 2 primeros a un extremo no reductor. Luego la glicosidasa toma el residuo del punto de ramificación con enlace 6 y lo hidroliza quedando una glucosa sin fosforilar. El punto de regulación la glucogenolisis es la glucógeno fosforilasa, que existe en dos estados diferentes: fosforilasa b (muy poco activa) y fosforilasa a (muy activa). Debido al diferente papel del glucógeno muscular y el hepático, su regulación es diferente. En el músculo esquelético suministra glucosa para que sea degradada obteniéndose ATP. Cuando se precisa realizar trabajo muscular se produce la activación por fosforilación, para ello el SNC estimula la médula adrenal y así segrega adrenalina. La adrenalina utiliza como segundo mensajero al cAMP, que es sintetizado por la adenilato ciclasa.La desfosforilación de las enzimas fosforiladas provoca su inactivación. En el músculo la fosfoproteína fosfatasa-I (PP1) desfosforila las enzimas fosforiladas de la cascada metabólica y detiene la glucogenolisis. La PP1 tiene, un inhibidor específico, el cual se une a la enzima cuando está fosforilado. La PP1 actúa únicamente cuando se encuentra unida al glucógeno, a través de su subunidad G. Cuando la cascada metabólica cAMP –dependiente se encuentra activada, dicha subunidad se encuentra fosforilada y no posee afinidad por la PP1 y ésta es inactiva. Cuando cesa el impulso del SNC disminuye la actividad fosforilasa quinásica y la relación quinasas/fosfatasas disminuye, entonces el sistema se desfosforila y se detiene la glucogenolisis. La glucógeno fosforilasa posee, además, un sistema de regulación alostérica que responde inmediatamente a las condiciones celulares en las que existe una baja carga energética y es independiente de la respuesta hormonal. Las señales que determinan la contracción muscular son cambios en la concentración de iones. Al aumentar la concentración de Ca2+ dentro de la célula se activa la degradación de glucógeno (coordinando los dos procesos de lisis y síntesis). El Ca2+ activa la glucógeno fosforilasa quinasa uniéndose a la subunidad calmodulina.

Figura 6.09: Control de la glucógeno sintasa en músculo. GSC: glucógeno sintasa kinasa.
Por otro lado el glucógeno hepático sirve como fuente de glucosa para los tejidos manteniendo la glucemia en valores adecuados. La regulación de la glucogenolisis hepática puede ser realizada por modificación covalente que consiste en la modificar la actividad de la glucógeno fosforilasa mediante fosforilación: la fosforilasa B (poco activa) no está fosforilada, mientras que la fosforilasa A (muy activa) se encuentra fosforilada. La cascada metabólica que dispara las fosforilaciones está activada por el glucagón, y aunque la cascada es sensible a la adrenalina sus mecanismos son diferentes. El glucagón impulsa una cascada de fosforilaciones utilizando el cAMP como segundo mensajero. El cAMP actúa de modulador para la proteínquinasa (PK)

que está formada por 2 subunidades catalíticas y 2 reguladoras cuando está inactiva (L2R2). El cAMP se une a las R que se separan de las catalíticas y la molécula queda activada (PKA). La PKA modifica a la glucógeno fosforilasa quinasa, activándola, y fosforila la glucógenosintasa, inactivándola. La adrenalina tiene receptores α y β adrenérgicos. El receptor α1-adrenérgico tiene tiene al Ca2+ como desencadenante de la cascada metabólica de fosforilaciones, necesitando, a su vez, al inosotol 1,4,5-trifosfato (IP3) como segundo mensajero para poder ser liberado al citoplasma. El receptor β2-adrenérgico al cAMP como segundo mensajero desencadenante de la cascada metabólica de fosforilaciones. La inactivación se produce por desfosforilación al actuar la PP1 sobre los fosfatos de la Ser de la glucógeno fosforilasa-A cuando pasa al estado tenso (T) y los expone, eliminándolos, transformando la enzima en la forma B, inactiva. La glucosa inhibe la glucógeno fosforilasa-A, desplazando su equilibrio alostérico hacia el estado T.
Fibras
Dentro de los simples encontramos la celulosa o fibra vegetal. Cumple funciones estructurales en los vegetales, en los cuales forma las paredes celulares. La pulpa de madera contiene un alto porcentaje de celulosa, y el algodón es prácticamente celulosa pura. Este polisacárido es el compuesto orgánico más abundante en la naturaleza. No todos los HC de la alimentación humana son digeribles, este es el caso de la celulosa, porque el humano, al igual que muchos mamíferos, no puede hidrolizarla. La fibra es entonces resistente a los procesos digestivos y se divide en dos grupos:Las fibras hidrosolubles, pectinas, algunas hemicelulosas, gomas y mucílagos que se encuentra en verduras, legumbres y frutas como la manzana, zanahoria, coliflor, batata, avena, salvado de avena o arveja. Retrasan el vaciamiento gástrico, 17nternacio la absorción de glucosa, colesterol y otras grasas en sangre. Las fibras no hidrosolubles como la celulosa, lignina, algunas hemicelulosas se encuentran en vegetales y cereales como berenjenas, peras, rábanos, todo tipo de salvado, brócoli y cereales enteros. Se caracterizan por su efecto de absorción de agua, reblandeciendo el bolo fecal y aumentando la velocidad de tránsito intestinal. Se aconseja una ingesta variada de las mismas diariamente, que se considera habitualmente recomendable entre los 25 y 30 gramos al día. Se consideraba que los fructanos inulina y oligofructosa poseían muchas de las características de la fibra dietaria pero no eran incluidos en las listas porque no precipitaban en etanol al 78% como se indicaba en los métodos de detección de fibras de la AOAC (Association of 17nterna Analytical Chemists) 17nternacional. Desde 1995 se cuenta con un método analítico de cálculo de fructanos, que utiliza un tratamiento enzimático de los alimentos en estudio con amiloglucosidasa e inulinasa, estudiándose los azúcares liberados por cromatografía de intercambio iónico. Los fructanos son compuestos de moléculas de fructosa unidas por uniones ß (2 1). Las moléculas de glucosa pueden unirse al final de la cadena por uniones (1 2) como en la sacarosa. El grado de polimerización varía de dos a varios cientos, siendo los mayores componentes de los fructanos la inulina y la oligofructosa, que se encuentran en frutas y vegetales.
Estructura del grano de trigo

El grano de trigo está compuesto por 3 partes: las envolturas, la semilla y el germen.Las envolturas representan apenas el 15% del peso del grano y están formadas por el pericarpio y los tegumentos externos e internos. A su vez, el pericarpio consta de 3 capas: epicarpio, mesocarpio y endocarpio. Estas envolturas comúnmente llamadas salvado, afrecho o afrechillo son ricas en fibras (celulosa y lignina), minerales, vitaminas y proteínas. Estos nutrientes se pierden luego de la molienda.El endospermo es la parte interna de la semilla y represente entre el 80% y el 85% del peso del grano. Está compuesto por una parte externa de origen proteico llamada aleurona y una parte interna rica en almidón denominada zona central. La capa proteica determina los diferentes tipos de harinas desde las féculas que no aportan proteínas hasta la sémola (harina gruesa de trigo).El germen, representa el 2% al 3% y es el encargado de dar origen a otra planta. Es rico en minerales, proteínas, vitaminas (Complejo B y Vitamina E) y grasas polinsaturadas (omega 3 y 6).
Figura 6.10: Estructura del grano de trigo.
REQUERIMIENTOS

Es recomendable que luego de la infancia, al menos la mitad de los requerimientos diarios de energía, sean proporcionados por los carbohidratos, especialmente los complejos. En una dieta habitual equilibrada el requerimiento diario puede ser del 55% del valor calórico total del día (VCT), aunque en deportistas oscila entre el 60% y el 70%, dependiendo del tipo de deporte que se realice y el momento de la planificación deportiva en la que se encuentre. En deporte también se acostumbra a relacionar los gramos de hidratos de carbono con los kilos del peso corporal (Pc), así entre 7 a 10 g por Kg de peso corporal son habituales en una estrategia de carga de HC. Veamos un ejemplo con un deportista masculino de 70 kg con un plan de 3.000 kcal (VCT):
Tabla 6.11: Hidratos de carbono en la dieta.
Recordemos que si se ingiriere este nutriente en forma deficitaria, tanto los aminoácidos como el glicerol de las grasas, pueden convertirse en glucosa para poder nutrir al sistema nervioso central. No obstante, es probable que una dieta que no contenga entre 50 y 100 g de hidratos de carbono diarios provoque cetosis. Esto es la formación de cuerpos cetónicos como el aceto acetato, el betahidroxibutirato y la acetona en mayor cantidad a la habitual y que comienzan a observarse en sangre (cetonemia) y en orina (cetonuria). La excreción continua de estos ácidos provocará pérdida de cationes y deshidratación. Si bien el cuadro no reviste la gravedad de la cetoacidosis en diabéticos, seguramente compromete el equilibrio interno y la perfomance física. Se desestima cualquier dieta de inanición o exagerada en grasas y proteínas como las llamadas cetogénicas.
INDICE GLICEMICOCuando ingerimos cualquier alimento que contiene glúcidos, los niveles de glucosa en sangre se van incrementando a medida que se produce la asimilación de sus almidones y otros HC. El tiempo que tardan en digerirse los nutrientes de los alimentos depende de varios factores como los tipos de HC y el contenido de fibra. La velocidad a la que se absorben los diferentes alimentos depende del tipo de nutrientes que los componen, de la cantidad de fibra presente y de la composición del resto de alimentos presentes en el estómago e intestino durante la digestión y absorción. Estos aspectos se valoran a través del índice glucémico de un alimento.El índice glucémico es el método utilizado para evaluar la velocidad de difusión de la glucosa de los alimentos hacia la sangre. Dicho de un modo simple, el índice se determina comparando el área bajo la curva del nivel de glucosa en sangre luego de la ingesta de una porción de 50 g de glucosa pura, con la obtenida utilizando la misma porción de otro alimento. El proceso consiste en tomar repetidas muestras de sangre a una persona a la que se le ha hecho consumir soluciones de glucosa pura (o pan blanco)

unas veces y el alimento en cuestión otras. A pesar de ser bastante complicado de determinar, su interpretación es que los índices elevados implican una rápida absorción, mientras que los índices bajos indican una absorción pausada.
Existen varios factores que modifican el índice glucémico:
1. Concentración y solubilidad de los hidratos de carbono: Una alta concentración de hidratos de carbono simples aumentan el índice glucémico.
2. Tamaño de la partícula: Cuanto menor sea el tamaño de las partículas, mayor será el IG ya que son más fácilmente digeridas.
3. Cocción y grado de gelatinización del almidón: Cuanto mayor sea el grado de gelatinización del almidón, mayor será el IG. Es decir que los fideos al dente tendrán menor IG que los fideos más cocidos.
4. Cocción y formación de almidón resistente: Una vez que un alimento rico en almidón se enfría, éste se transforma químicamente (almidón resistente) y genera un mayor trabajo digestivo. Por lo tanto una ensalada de papa fría tendrá menor IG que una ensalada de papa tibia o caliente.
5. Mezcla digestiva: Las proteínas y las grasas retardan el vaciamiento gástrico, con lo cual se demora la digestión y absorción, disminuyendo así el IG.
6. Fibra dietética: estas estructuras retrasan el vaciado gástrico, generando un efecto similar al de las proteínas y las grasas.
Dentro de los distintos grupos de diferentes índices glicémicos, los alimentos pueden necesitar a su vez, diferentes cantidades para aportar 50 g de de hidratos de carbono. Por ejemplo para igualar los 50 g que aporta la glucosa son necesarios 70 g de miel, 60 g de copos de maíz, 90 g de pan francés, 75 g de chocolate, 250g de papa hervida, 260 g de banana, 170 g de arroz blanco, 370 g de tallarines, 100 g de uva blanca y 200g de uva negra.. Entre los de moderado índice glicérico tenemos 500 g de naranja y 195 g de arroz integral. Entre los de bajo índice tenemos a las manzanas que necesitan de 400 g para aportar los 50 g de hidratos de carbono, 450g de durazno o ciruelas, 295 g de lentejas, 1.100 ml de leche entera o 1.000 ml de descremada y 800 ml de yogur descremado.
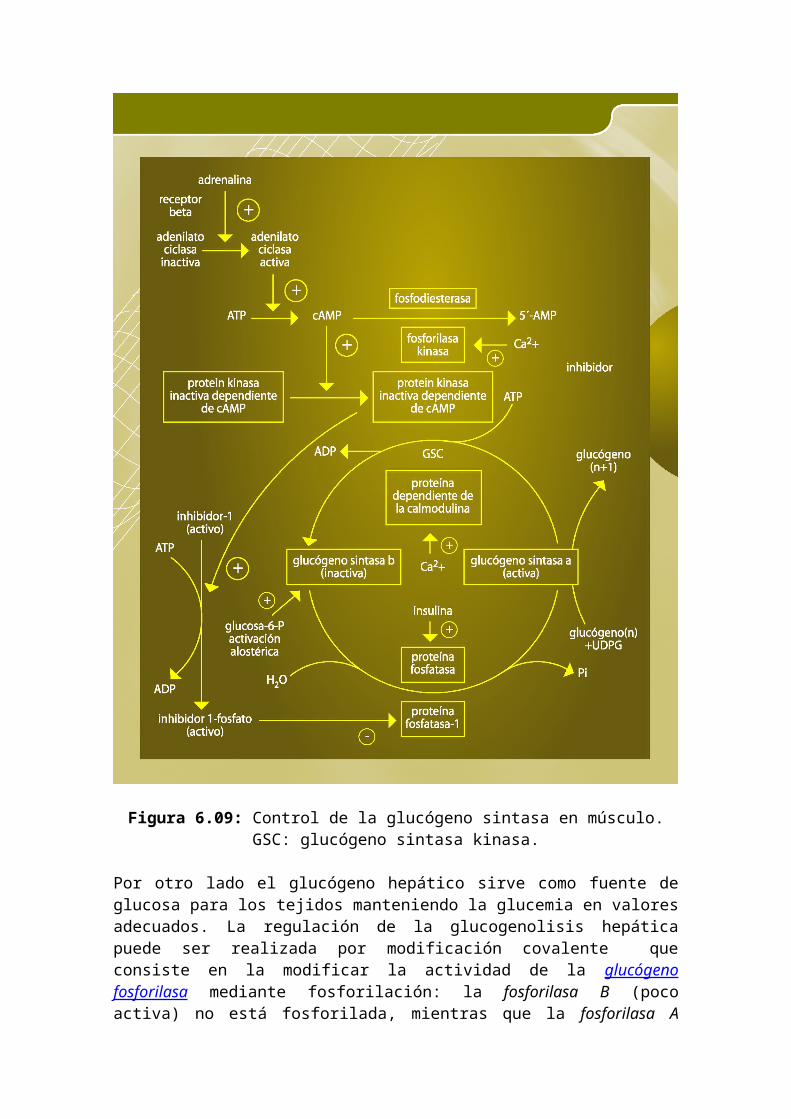
Tabla 6.12: Índice glucémico de los alimentos.
Podemos pensar en el uso del índice glicémico como una referencia para la selección de diferentes alimentos a fin de optimizar la estrategia nutricional. Los alimentos con elevado índice glicémico podrían ser útiles en situaciones como entrenamientos y competencias o inmediatamente después de un ejercicio agotador (1 g x kg de peso corporal). Los alimentos con moderado índice glicémico estarán indicados antes de trabajos de resistencia a fin de sostener los valores de glucemia durante tiempos prolongados.

HIDRATOS DE CARBONO Y EJERCICIO
Las dietas para deportistas deben aportar nutrientes necesarios para el ejercicio y para una adecuada recuperación. Cuanto más elevada es la cantidad de glucógeno almacenado en los músculos, mayor es la intensidad del esfuerzo que puede realizarse y mayor su duración, su reserva está condicionada por las características de la dieta y el volumen y tipo de actividad física que desarrolla en los días previos a la competencia. Si el glucógeno se utiliza en una proporción superior a la que se regenera, su concentración se reduce y el trabajo físico es cada vez menos eficiente. Entonces resulta difícil realizar los ejercicios habituales a la intensidad normalmente tolerada. Usualmente, los valores de glucógeno muscular en músculos entrenados son de 100 -120 mMol/Kg de músculo húmedo. Si la tasa de síntesis es de 5 mMol/Kg. por hora, los atletas necesitan 24 a 36 horas luego de la última sesión de entrenamiento para normalizar sus reservas de glucógeno, con una ingesta alta de hidratos de carbono (8 a 10 g/kg/día). Los hidratos de carbono ingeridos 30 minutos antes de un ejercicio causan una reducción en los niveles de glucosa en sangre antes de comenzar la prueba, pero estos efectos son transitorios y no afecten al rendimiento.
Figura 6.13: Fuentes energéticas durante el ejercicio.
Durante la fase inicial del ejercicio, la mayor parte de la energía de los hidratos de carbono proviene del glucógeno muscular. Al continuar el ejercicio, el glucógeno muscular disminuye su contribución a la demanda de hidratos de carbono. Pero hay mayor dependencia de la glucosa sanguínea como fuente de hidratos de carbono. Luego de tres horas de ejercicio, la mayor parte de la energía obtenida de los hidratos de carbono procede del metabolismo de la glucosa. Luego de dos o tres horas de ejercicio, la concentración de la glucosa sanguínea cae y debido al agotamiento de las reservas de glucógeno hepático el hígado disminuye su producción de glucosa. Los hidratos de carbono aportados durante el ejercicio retrasan la fatiga tanto en ejercicios continuos como intermitentes.

Una estrategia puede consistir en la utilización de bebidas deportivas con diferentes concentraciones de hidratos de carbono. En las bebidas para reposición de hidratos de carbono se deben tener en cuenta variables como su concentración, tipo, osmolaridad, sabor y temperatura. Es posible ingerir una solución que contenga 0.5 a 1.5 g/kg de peso corporal disuelto en un volumen de 500 a 1.000.ml antes de comenzar el ejercicio, continuando con 150 a 250.ml de líquidos al 8.% cada 15 a 30.minutos, aportando un mínimo de 0,2 g de hidratos de carbono por kilo de peso. El ritmo de ingestión de los hidratos de carbono debe ser en promedio de 30 a 70 g/h. Ello asegura que los tejidos recibirán un gamo de hidratos de carbono por minuto en el momento en que comience la fatiga.Si bien algunos individuos pueden tolerar elevadas concentraciones de hidratos sin molestias, por lo general el contenido debe ser de 8% ya que las bebidas con esa concentración pasan a sangre a la misma velocidad que el agua. Las soluciones de más del 10% demoran su pasaje a intestino delgado, al precisar un aporte de líquidos para disminuir su osmolaridad.
Tabla 6.14: Aporte de carbohidratos en ejercicios.
Los polímeros de glucosa aumentan el contenido de hidratos de carbono sin incrementar la osmolaridad y no afectando el sabor de la bebida. Por su parte la fructosa es absorbida y metabolizada con mayor lentitud, ya que necesita un paso hepático para convertirse en glucosa, constituyendo una fuente de hidratos de carbono de utilidad en eventos de larga duración, aunque son interesantes las mezclas de hidratos de carbono (glucosa, sacarosa, fructosa y maltodextrinas) siempre que la fructosa no sea predominante. Las formas sólidas o liquidas son igualmente eficaces en cuanto al aporte de hidratos de carbono, pero las bebidas deportivas tienen como importante objetivo las reposición de líquidos. En competencias prolongadas pueden ser preferibles los sólidos que por sus características físicas (textura, mordida, sabor) producen un efecto de saciedad. Recordar siempre entrenar la ingesta de estos productos durante los entrenamientos.

Recuperación
Los atletas pueden realizar varias sesiones de entrenamiento o incluso competir en diferentes pruebas al día, necesitando un eficaz reposición de los depósitos de glucógeno. La síntesis de glucógeno muscular es más rápida si los hidratos de carbono son consumidos inmediatamente después del ejercicio ya que la tasa de resíntesis es en promedio de 5 a 7mMol/kg/h, pero luego de unas horas cae a la mitad debido a la insulinoresistencia muscular, mecanismo protector que previene el desarrollo de hipoglucemia cuando hay disponible una inadecuada cantidad de glucosa. El aporte de hidratos de carbono posterior al ejercicio mejora el proceso de recuperación del glucógeno muscular, mantiene la glucemia y previene el desarrollo de insulinoresistencia.La tasa de formación de glucógeno muscular luego del ejercicio es proporcional a los hidratos de carbono ingeridos y si se consumen de 150 a 650 g de hidratos de carbono durante las 24 horas posteriores la síntesis es óptima. La recomendación es consumir 1.2 a 1.5g/kg/h después de haber terminado el ejercicio y continuar el aporte a intervalos de 2 horas por lo menos durante las 6. La combinación de proteína e hidratos de carbono puede acelerar la formación de glucógeno muscular post ejercicio en tres veces la cantidad en reposo por los mayores efectos en la secreción de insulina, pero es efectiva si el aporte de hidratos de carbono no es mayor a 1g/kg/h. La ingestión de 0.8 g de carbohidratos y 0.4 g de proteínas produce similar concentración de glucógeno muscular en 4 horas de recuperación que la ingesta de 1.2g/kg/h de hidratos de carbono solamente. La ingesta de hidratos de carbono con IG alto produce una mayor síntesis de glucógeno que la ingesta de hidratos de carbono de IG bajo. En ese sentido la glucosa y sacarosa son más rápidas que la fructosa, aunque ésta sí lo es para la síntesis de glucógeno hepático.

Bibliografía
Burke LM y Deakin V. (2000). Clinical Sport Nutrition. Sydney: Mc Graw-Hill.Burke LM, Claassen A, Hawley JA, Noakes TD (1998). Carbohydrates intake during prolonged cycling minimizes effect of glycemic index of pre-exercise meal. Journal Applied Physiology, 85: 2220-2226.Burke LM, Claassen A, Noakes TD. (1998). No effect of glycemic index of pre-exercise meals with carbohydrate intake during exercise. Medicine & Science en Sport & Exercise, 30, S82. (Abstract 471).Burke LM, Cox G, Cumming NK, Desbrow B. (2001). Guidelines for daily carbohydrate intake. Do athletes achieve them? Clinical Sport Medicine, 31 (4) 267-299.Burke LM, Hawley JA. (1999). Carbohydrate and exercise. Current Opinion in Clinical Nutrition and Metabolic Care;2:515-520.Costill DL, Hargeaves M. (19992). Carbohydrate nutrition and fatigue. Clinical Sport Medicine, 13 (2): 86-92.Coyle E. (1994). Suplementación de carbohidratos durante el ejercicio. Revista Actualización Ciencias del Deporte, 2 (5).Davis JM, Brown AS. (2001). Carboidratos, hormônios e performance em exercicios de resistencia. Gatorade Sport Science Institute, 31 (10-12).Díaz Hernández DP y Burgos Herrera LC. (2002). ¿Cómo se transporta la glucosa a través de la membrana celular? IATREIA, 15 (3).Hawley JA, Schabort EJ, Noakes TD, Dennis, SC. (1991). Carbohydrate-loading and exercise performance. Clinical Sport Medicine, 24: 73-81.Ivy JL. (2001). Dietary strategies to promote glycogen synthesis after exercise. Can. Journal Applied Physiology, 26 (suppl): 236-5245.Ivy JL. (1999). Role of carbohydrate in physical activity. Clinical Sport Medicine, 18 (3): 469-484.Prosky L y Hoebregs H. (1999). Methods to determine food inulin and oligofructose. Journal of Nutrition,129:1418S-1423S.Rankin J. (2000). Los carbohidratos en las dietas y la performance de ejercicios intensos de corta duración. Gatorade Sports Science Institute. Sport Science Exchange, 13(4).Romijn JA, Coyle EF, Sidossis LS y otros. (1993). Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J Physiol Endocrinology, 265: E380-E391.Salas-Salvadó, J. (2000). Nutrición y dietética clínica. Barcelona: Masson.Skinner RE, Coleman E y Rosenbloom CA. (2000). Ergogenic aids en Rosenbloom C.A. (ed) Sports Nutrition: A Guide for the Professional Working with Active People. 3rd Ed. Chicago: The American Dietetic Association, 107-146.Tarnopolsky M, Zawada C, Richmond LB y otros. (2001). Gender differences in carbohydrate loading are related to energy intake. Journal Appl. Physiol, 91(1): 225-230.Wee SL, Williams C, Gray S, Horabin J. (1999). Influence of high and low glycemic index meals on endurance running capacity. Med. Sci. Sport Exercises, 31: 393-399.



















