
Oviedo L. (2008). Análisis del uso de hábitat del delfín manchado pantropical Stenella attenuata...
115
UNIVERSIDAD NACIONAL SISTEMA DE ESTUDIOS DE POSTGRADO ESCUELA DE CIENCIAS BIOLÓGICAS ESTACIÓN DE BIOLOGÍA MARINA PROGRAMA DE MAESTRÍA EN CIENCIAS MARINA Y COSTERAS ANÁLISIS DEL USO DE HÁBITAT DEL DELFÍN MANCHADO PANTROPICAL Stenella attenuata (Cetacea: Delphinidae) EN EL GOLFO DULCE, COSTA RICA Por: Lenin E. Oviedo Correa Puntarenas, Costa Rica 2008 Tesis presentada para optar al grado académico de Magíster Scientiae en Ciencias Marinas y Costeras, con énfasis en Manejo de los Recursos Marinos y Costeros. Cumple con los requisitos establecidos por el Sistema de Estudios de Postgrado de la Universidad Nacional, Heredia, Costa Rica
-
Upload
centro-de-investigacion-de-cetaceos-ceic-costa-rica -
Category
Documents
-
view
209 -
download
2
description
Oviedo L. (2008). Análisis del uso de hábitat del delfín manchado pantropical Stenella attenuata (Cetacea: Delphinidae) en el Golfo Dulce, Costa Rica. Master's Thesis, Universidad Nacional Costa Rica PROCMAR
Transcript of Oviedo L. (2008). Análisis del uso de hábitat del delfín manchado pantropical Stenella attenuata...

UNIVERSIDAD NACIONAL SISTEMA DE ESTUDIOS DE POSTGRADO
ESCUELA DE CIENCIAS BIOLÓGICAS ESTACIÓN DE BIOLOGÍA MARINA
PROGRAMA DE MAESTRÍA EN CIENCIAS MARINA Y COSTERAS
ANÁLISIS DEL USO DE HÁBITAT DEL DELFÍN MANCHADO PANTROPICAL Stenella attenuata (Cetacea: Delphinidae) EN EL GOLFO DULCE, COSTA RICA
Por: Lenin E. Oviedo Correa
Puntarenas, Costa Rica
2008 Tesis presentada para optar al grado académico de Magíster Scientiae en Ciencias Marinas y Costeras, con énfasis en Manejo de los Recursos Marinos y Costeros. Cumple con los requisitos establecidos por el Sistema de Estudios de Postgrado de la Universidad Nacional, Heredia, Costa Rica

1
UNIVERSIDAD NACIONAL SISTEMA DE ESTUDIOS DE POSTGRADO
ESCUELA DE CIENCIAS BIOLÓGICAS ESTACIÓN DE BIOLOGÍA MARINA
PROGRAMA DE MAESTRÍA EN CIENCIAS MARINA Y COSTERAS
ANÁLISIS DEL USO DE HÁBITAT DEL DELFÍN MANCHADO PANTROPICAL Stenella attenuata (Cetacea: Delphinidae) EN EL GOLFO DULCE, COSTA RICA.
Por: Lenin E. Oviedo Correa
Puntarenas, Costa Rica 2008
Tesis presentada para optar al grado académico de Magíster Scientiae en Ciencias Marinas y Costeras, con énfasis en Manejo de los Recursos Marinos y Costeros. Cumple con los requisitos establecidos por el Sistema de Estudios de Postgrado de la Universidad Nacional, Heredia, Costa Rica

2
La presente Tesis de Maestría se _______________ por el Comité de Gestión Académico del Programa de Maestría en Ciencias Marinas y Costeras de la Escuela de Ciencias Biológicas de la Universidad Nacional, como requisito parcial para optar por el grado académico de Magíster Scientiae con énfasis en manejo de los recursos marinos y costeros.
__________________________________ M.Sc. Luis Villalobos Chacón
Director Programa de Maestría en Ciencias Marinas y Costeras
_______________________________
Ph.D. Ricardo Jiménez Montealegre Director
Escuela de Ciencias Biológicas
______________________________
Ph.D. Luis Sierra Sierra Decano
Facultad de Ciencias Exactas y Naturales
___________________
Ph.D. Daniel Odell Tutor
___________________
M.Sc. Dora Ingrid Rivera Luther Asesor
_____________________
M.Sc. Miguel Iñiguez
Asesor
_____________________
Representante del SEPUNA

3
“…porque los chicos y muchos hombres no cuentan lo que ven, sino lo que han leído
sobre lo mismo que acaban de ver…”
Juan Darién, Horacio Quiroga

4
A mi madre Corina, porque en cada una de
estas páginas se refleja todo el amor y
dedicación que inviertes en tus hijos…
A mi hermana katiuska, porque fuiste la bebe
más linda del mundo, y hoy eres un hermoso
ser humano que enorgullece cada día de mi
vida…
A Idhalmis, Kis y Herlin, porque en esta etapa
de mi vida siento que mis logros son de
ustedes, el cariño y la admiración que
sentimos y nos profesamos va más allá del
tiempo y la distancia…
A las tribus de los Océanos, porque
continúan inspirando cada paso que doy…

5
AGRADECIMIENTOS
El desarrollo de este trabajo involucró a mucha gente que de manera directa o indirecta dieron su valioso aporte: Agradezco el apoyo institucional de Programa de Maestría en Ciencias Marinas y Costeras (PROCMAR - UNA); en particular a su secretaria María Antonieta Parker, y especialmente a su director Luis Villalobos Chacón, quien siempre fue un impulso clave no solo de este trabajo sino de la formación y evolución de sus estudiantes. Mucho de la planificación de este trabajo y gran parte de una evaluación preliminar se realizó en las instalaciones de la Estación de Biología Marina (EBM) en Puntarenas, y en la Estación de Ciencias del Mar (ECMAR) en Punta Morales. Quisiera agradecer especialmente a Inés Lostalo (EBM), Ramiro Segura y Oscar Pacheco (ECMAR). Agradezco la colaboración de la Asociación para el Desarrollo del Pacífico Sur (ADEPAS), y a su coordinador Rómulo León, por su valiosa colaboración al prestar las instalaciones de “Casa Loma”, en Rincón de Osa como la base del trabajo de campo. Un agradecimiento especial al encargado de “Casa Loma” Don Jorge Medina. Un agradecimiento especial a las instituciones que dieron el soporte financiero de esta investigación: Fundación Gran Mariscal de Ayacucho (Fundayacucho) y a su programa de becas de postgrado de excelencia académica. A la Sociedad para el Estudio de Mamíferos Marinos (Society of Marine Mammalogy) quienes otorgaron dos subsidios de investigación del programa “small grant in aid”. Un aporte financiero crucial para el desarrollo de este estudio fue el subsidio de investigación de ISV-Costa Rica (International Student Volunteers), les agradezco a mis compañeros Wagner Quirós, Manuel de la Cruz Ramírez y Kattia Garro, no solo por el interés y la preocupación sobre el bienestar financiero del proyecto, sino por su entusiasmo y motivación. Deseo especialmente honrar y agradecer a mi comité de tesis: a mi tutor Daniel Odell, quien a sido una influencia importante en mi desarrollo dentro del campo de la cetología, y a quien no le ha faltado consejos sabios y palabras de aliento, para dar en los momentos más oportunos. A mi asesor Miguel Iñíguez por todo ese ejemplo de profesionalismo, y por el cariño con que aporta su amplio conocimiento. A mi asesora y profesora Dora Ingrid Rivera por su calidez y paciencia, por la enseñanza del valor de los detalles y la buena labor. A mi tutora ad honoren Hilda María Víquez, por todo su empeño, motivación, conocimiento y cariño, por supuesto también por toda esa gama cromática que tanto resalta el sentido de la ciencia. Quisiera extender un agradecimiento particular a mis profesores: Pre-Grado; Alex Ferguson, Renato Di Nobrega, Francisco Provenzano, Carmen Ferreira, Joaquín Buitriago, Irene Astor, Andrés Rodríguez y Juan capelo, por tanta Biología, Ecología, Estadística y Oceanografía, es bonito poder apreciar su buena labor a esta etapa de mi desarrollo profesional y académico. Postgrado; Dora Ingrid Rivera, Hilda Víquez, Luis Villalobos, Farid Tabash, Laura González, y José Luis Araya, Les agradezco el cariño y la dedicación a lo largo de toda su enseñanza.

6
No puedo dejar de mencionar y agradecer a Hedelvys Guada, por sus sabios consejos y su buen ejemplo. Debo agradecer a todos esos profesionales en el campo de la cetología que ha influido en mi motivación y formación; Erich Hoyt, Janet Mann, John Ford, Willian F. Perrin, Steven Swartz, Alexander Zerbini, Fernando Félix, Alejandro Acevedo, Steve Dawson, Ana Cañadas, Hal Whitehead, David Lusseau, Kathleen Dudzinski, Leszek Karczmarski, Dagmar Ferlt, Jeanette Thomas, Giovanni Bearzi. También tengo un agradecimiento especial a mis amigos y colegas, a mi generación de cetólogos: Romina Acevedo, María Alejandra Esteves, Noemi Silva quienes desde fuera de Costa Rica han motivado constantemente el desarrollo de esta investigación, he tenido el honor de aprender con ustedes y de ustedes. A mis compañeros de Fundación Vida Marina, especialmente Marc Fernández, Denise Echeverria y Sierra Goodman, quien ha sido fuente de inspiración de muchos de mis colegas a través de sus fotos, y con quien comparto su amor al mar de Osa y sus cetáceos. A mis asistentes de campo y co-investigadores, Juan Diego Pacheco, María Gabriela Silva y David Herra. A todos los voluntarios de ISV que han ayudado en las labores de campo y quienes han aportado su material fotográfico. Una parte importante del análisis de los datos colectados en campo fue posible gracias a la valiosa ayuda de Dulce Natalie Arocha, en el desarrollo de los modelos estadísticos, y de Juan Eudes en la elaboración de los mapas y SIG. Agradezco inmensamente a mi gente de Golfo Dulce y de Rincón de Osa, especialmente a la familia Medina-Herra; Don Jorge, Doña Azucena, David, Estefany y Erick, por brindarme ese cariño de familia tan importante cuando se esta lejos de casa. A mi capitán Marcos Loaiciga “Taboga”, quien con cariño comparte todo su basto conocimiento sobre los mares de Osa. Tengo un agradecimiento especial a mis compañeros de Maestría; Ricardo (Tato) Hernández, Luis Adrian Vega, Helven (Leven) Naranjo, Rodney (Baulo) Piedra, Jonathan Chacón, y Ulises (Licho) Arrieta, me siento afortunado de haber pertenecido a la mejor generación de PROCMAR. Debo también mencionar a la nueva generación; a mis amigos Fernando Rincón y Ricardo (Negro) Torres, quienes han traído consigo todo el buen bagaje de mi alma mater FLASA. Una mención muy especial a mis amigos y hermanos Ricardo (Tato) Hernández y María Alejandra Esteves, quienes no solo me han dado el regalo de la amistad y han compartido conmigo el amor al mar, sino que con su cariño y afecto me han hecho parte de su familia. Finalmente mi gratitud a dos países; a Costa Rica por la bonita experiencia, por Osa, sus cetáceos y la posibilidad de crecimiento profesional… y a mí amada Venezuela, porque todos los días agradezco el haber nacido en tu tierra de gracias.

7
INDICE GENERAL CONTENIDO PÁGINA
Hoja de aprobación 2 Índice de contenidos 7 Índice de cuadros 9 Índice de figuras 10 Índice de anexos 11 Resumen ejecutivo y palabras claves 12 CAPITULO 1. INTRODUCCIÓN 13 1 Introducción 13 1.a Problema 15 1.b Hipótesis 15 1.c. Objetivos 16
1.c Objetivo General 16 1.c.1. Objetivos específicos 16
1.d Justificación 16 1.e. Antecedentes 17
1.e.1 Marco socio-económico relacionado con las perspectivas de manejo 21 1.e.2 Contexto legal de los cetáceos como recursos en Costa Rica. 23
CAPITULO 2. MARCO TEÓRICO 25 2.a. Uso de Hábitat 25 2. b. Alimento 27
2.b.1. La teoría del consumidor óptimo 28 2.c Cobertura en el medio marino 29 2.d. Abundancia 30 CAPITULO 3. MATERIAL Y MÉTODOS 32 3.a. Área de estudio 32 3.b. Muestreos en campo 34 3.c Abundancia Relativa 35 3.d Muestreo Etológico 37
3.d.1 Análisis de datos etológicos 39 CAPITULO 4. RESULTADOS 45 4.a. La abundancia relativa del delfín manchado en Golfo Dulce 45 4.b. Conducta del delfín manchado en Golfo Dulce 49 4.c. Uso de hábitat del delfín manchado en Golfo Dulce 51
4.c.1 Variables eco-geográficas que definen el hábitat de alimentación 58

8
CAPITULO 5. DISCUSIÓN 66 5.a. La abundancia relativa del delfín manchado en golfo dulce 66 5.b. La conducta del delfín manchado en golfo dulce 68 5.c. El uso de hábitat del delfín manchado en golfo dulce 71
5.c.1 Localización espacial de las áreas de alimentación 74 5.c.2 Caracterización del nicho ecológico de alimentación 75
CAPITULO 6. CONCLUSIONES 79 CAPITULO 7. IMPLICACIONES PARA EL MANEJO Y CONSERVACIÓN 82 7.a Delfines manchado como sostén de la industria de observación de cetáceos en golfo dulce 82 7.b La cuenca interna de Golfo Dulce: Un área marina protegida (AMP) 86 CAPITULO 8. LITERATURA CITADA 89 CAPITULO 9. ANEXOS 102

9
LISTA DE CUADROS
CUADRO PÁGINA Cuadro 1. Esfuerzo de investigación en campo en muestreos con S. attenuata en Golfo Dulce, Costa Rica: Temporada de lluvia 2007 – Temporada seca 2008
46
Cuadro 2. Índices y estimados de abundancia relativa de S. attenuata en Golfo Dulce, Costa Rica: Temporada de lluvia 2007 – Temporada seca 2008.
47
Cuadro 3. Frecuencias de avistamiento de delfines manchados, S. attenuata en Golfo Dulce, Costa Rica: Temporada de lluvia 2007 – Temporada seca 2008.
48
Cuadro 4. Comparación del consumo de biomasa de S. attenuata en Golfo Dulce y Golfo de Papagayo, Costa Rica.
48
Cuadro 5. Índices de Uso de Área y Actividad para S. attenuata en la cuenca interna de Golfo Dulce durante la temporada de lluvia 2007
54
Cuadro 6. Índices de Uso de Área y Actividad para S. attenuata en la cuenca interna de Golfo Dulce durante la temporada seca 2008
56
Cuadro 7. Variables eco-geográficas asociadas al nicho ecológico de alimentación de S. attenuata en la cuenca interna de Golfo Dulce: temperatura superficial y distancia de ríos
58
Cuadro 8. Variables eco-geográficas asociadas al nicho ecológico de alimentación de S. attenuata en la cuenca interna de Golfo Dulce: profundidad máxima y pendiente.
60 - 61
Cuadro 9. Análisis de Factores de Nicho Ecológico de la variable eco-geográficas profundidad máxima (Zmax) y pendiente (IP), para S. attenuata en Golfo Dulce, durante la temporada de lluvia 2007.
62 - 63
Cuadro 10. Análisis de Factores de Nicho Ecológico de la variable eco-geográficas profundidad máxima (Zmax) y pendiente (IP), para S. attenuata en Golfo Dulce, durante la temporada seca 2008.
64 -65

LISTA DE FIGURAS
FIGURAS PÁGINA Figura 1. Área de estudio, Golfo Dulce, Área de Conservación de Osa (ACOSA), Puntarenas, Costa Rica.
33
Figura 2. Presupuesto conductual de S. attenuata en Golfo Dulce Costa Rica temporada de lluvia 2007.
49
Figura 3. Presupuesto conductual de S. attenuata en Golfo Dulce Costa Rica temporada seca 2008.
50
Figura 4. Localización espacial de las conductas registradas para S. attenuata en Golfo Dulce Costa Rica.
51
Figura 5. Localización espacial de las actividades de alimentación registradas para S. attenuata en Golfo Dulce, Costa Rica en la temporada de lluvia.
52
Figura 6. Localización espacial de las actividades de alimentación registradas para S. attenuata en Golfo Dulce, Costa Rica en la temporada seca.
53
Figura 7. Clases de uso para S. attenuata durante la temporada de lluvia en Golfo Dulce.
53
Figura 8. Clases de uso para S. attenuata durante la temporada seca en Golfo Dulce.
54
Figura 9. Distribución espacial de la probabilidad de ocurrencia de la actividad de alimentación para S. attenuata en la temporada de lluvia, Golfo Dulce, Costa Rica.
57
Figura 10. Distribución espacial de la probabilidad de ocurrencia de la actividad de alimentación para S. attenuata en la temporada seca, Golfo Dulce, Costa Rica.
57
Figura 11. Clases de temperatura superficial del mar para S. attenuata en Golfo Dulce.
59
Figura 12. Clases de temperatura superficial del mar para S. attenuata en Golfo Dulce.
59

11
LISTA DE ANEXOS
ANEXOS PÁGINA Anexo 1. Descripción del delfín manchado pantropical (Stenella attenuata).
102
Anexo 2. Clave de Muestreo Conductual (García y Dawson 2003, desarrollada por Schneider 1999).
106
Anexo 3. Definiciones de los estados conductuales a evaluar para la estimación del presupuesto conductual de S. attenuata en el Golfo Dulce.
107
Anexo 4. Modelo Aditivo Generalizado (MAG) para la temporada de lluvia y temporada seca.
108
Anexo 5. Distribución espacial de la probabilidad de ocurrencia de las actividades de movilización (verde), merodeo (azul), socialización (violeta) y descanso (gris) en la cuenca interna de Golfo Dulce, Costa
110
Anexo 6. Líneas de acción sugeridas para el manejo y conservación de los cetáceos en Golfo Dulce
112

12
RESUMEN EJECUTIVO
Golfo Dulce es un ecosistema particular donde convergen características neríticas y
oceánicas. La productividad se asume como limitada, sin embargo, sostiene una
población de más de 80 delfines nariz de botella Tursiops truncatus, y cerca de 600
delfines manchados pantropicales Stenella attenuata. El objetivo de esta contribución
es evaluar el uso de hábitat de S. attenuata para determinar los requerimientos
necesarios para asegurar su permanencia en el Golfo Dulce, Península de Osa,
Costa Rica. El uso del hábitat del delfín manchado se determinó por medio de
técnicas de ecología conductual y espacial, también se estableció el nicho ecológico
por medio de un modelo mecanicista, donde la variable respuesta es la alimentación,
condicionada por el gradiente de de recursos (presas) disponibles ante factores eco-
geográficos que ejercen una influencia en la ocurrencia de la alimentación. La
profundidad y la pendiente como variables físicas, diferencian el nicho ecológico de
los delfines manchados, con influencia de los ríos Tigre, Agujas, Rincón, Esquinas y
Coto Colorado, en los episodios de captura y consumo de presa. La presente
investigación provee de una línea base que puede sustentar la determinación de la
capacidad de carga, ante la presión de las embarcaciones turísticas en la
observación de cetáceos en Golfo Dulce . La delimitación espacial de zonas claves
de captura y consumo de presas, permite identificar el hábitat crítico de alimentación.
Lo anterior es un argumento sustentable para el establecimiento de áreas de
protección marina en la cuenca interna del Golfo Dulce
Palabras clave: Golfo Dulce, Hábitat, Uso de Hábitat, Estado Conductual,
Alimentación, Stenella attenuata.

13
INTRODUCCIÓN
La definición de hábitat para organismos acuáticos, en términos espaciales
representa un reto particular, específicamente para especies pelágicas, ya que los
limites geográficos en ecosistemas marinos están ausente (Hooker et al. 2002). Esto
implica que muchos de los conceptos para la evaluación y manejo de hábitat son
modificados con el fin de adaptarlos al ámbito acuático. Otra consideración
importante para la compresión de procesos de vida en ecosistemas marinos es la
necesidad de su entendimiento, no obstante dichos procesos de vida también
ocurren en una dimensión no tan perceptible como los ambientes terrestres. La
evaluación y el análisis de hábitat no son un fin en si, sino que el valor de esta labor
esta en la información y utilidad que estas aportan para el manejo de ecosistemas y
vida silvestre (Ojasti 2000). La evaluación de hábitat se parcializa por las
necesidades de la fauna, y se condiciona por la manera de como detectan los
animales su entorno, lo que implica elementos intrínsecos de la biología del
organismo como su estatura, estrato de ubicación, hábitos y capacidad sensorial.
Golfo Dulce sostiene una población de más de 80 delfines nariz de botella
Tursiops truncatus (Acevedo y Matthew 2005), y una población considerable de más
de 300 delfines manchados pantropicales, Stenella attenuata (Oviedo 2006). Este
golfo es un ecosistema particular, un mar interno de cuenca semi-cerrada donde
convergen características neríticas y oceánicas. La productividad es aparentemente
limitada, con una contribución significativa de las descargas de varios ríos como el
Rincón, Esquinas, Coto Colorado, Agujas y Tigre. Ciertamente T. truncatus reside
dentro del golfo y las actividades claves de supervivencia como la alimentación y
reproducción han sido documentadas. El uso del hábitat del delfín manchado
representó una interrogante durante el progreso de esta investigación; se
sospechaba de tránsitos de la especie entre el golfo y el océano abierto
regularmente. Igualmente, se estima que S. attenuata podría usar el golfo como
refugio de la presión de depredadores en el océano Pacífico abierto (Acevedo &
Buckhart 1998, Cubero-Pardo 1998a). Esta contribución es una evaluación ecológica

14
que intenta determinar la utilización de hábitat del delfín manchado pantropical en
términos de los requerimientos básicos para sustentar su presencia dentro de la
cuenca interna del Golfo Dulce.
Esta investigación se apoya en enfoques de ecología espacial y conductual
para determinar el uso de hábitat (Lusseau y Higham 2004) y en el concepto de
hábitat crítico aplicado a mamíferos marinos (Hoyt 2005), haciendo énfasis primero
en la descripción detallada de la distribución y abundancia relativa de la especie,
como indicador clave del uso de hábitat. La estimación de abundancia permitió
hacer una aproximación ecológica del consumo de energía de la especie en el
ecosistema (Hooker et al. 2002, Read & Brownstein 2003), y la comparación del
consumo con otras poblaciones en Costa Rica (May - Collado 2001, May - Collado y
Morales 2005) en base al nivel trófico estimado de S. attenuata como depredador
(Pauly et al., 1998).
Los aspectos conductuales claves que definen el uso de hábitat se analizaron
a la luz de la “teoría del consumidor optimo”, de acuerdo a los elementos implícitos a
dicha teoría descritos en Begon et al. (2006). Por medio de modelos teóricos
mecanicistas se diferenciaron las estrategias de conducta alimentaria en dos
tendencias básicas; la especialización y la generalización, en el contexto específico
de Golfo Dulce y adicionando factores importantes como la competencia inter-
específica e intra-específica.
También se describen las características y factores ambientales que se
relacionan con la ocurrencia de los aspectos conductuales claves, que definen el uso
de hábitat; y que permiten la determinación del área en estudio como un hábitat
crítico de alimentación. Por medio de los índices del Análisis de Factores de Nicho
Ecológico (Hirzel et al. 2001,2002; Compton 2004) se definió el nicho ecológico de
alimentación, en donde la conducta alimentaria fue la variable respuesta ante los
descriptores eco-geográficos (profundidad máxima y pendiente del fondo). El
conjugar las características de un modelo empírico predictivo de uso del hábitat con

15
aspectos conductuales, que de manera indirecta reflejan el efecto de un gradiente
de recursos tróficos; derivaron finalmente en un modelo mecanicista descriptivo que
ilustra el uso de hábitat considerando la dinámica ecológica que implican las
respuestas fisiológicas ante factores ambientales igualmente dinámicos (Guisan y
Zimmermann 2000).
Finalmente se exploran las perspectivas de manejo basadas en la información
generada. Dos enfoques se ponderan para las recomendaciones de manejo: el
enfoque ecosistémico que identifica al Golfo Dulce como unidad de conservación
clave, y un enfoque de especie unitaria para delfines manchados como recurso no
consuntivo y de gran importancia para el eco-turismo local.
1. a Problema Golfo dulce presenta una población de delfines manchados pantropicales, la
utilización de hábitat por este depredador no es clara, ya que los indicios anteriores
de eventos que implicaran conducta alimentarias no fueron concluyentes, sumado
esto al hecho de que las aguas del Golfo son aparentemente oligotróficas, es decir
las condiciones ambientales podrían no ser propicias para sostener una abundancia
de presas potenciales. El registro de elementos de la biología de la especie como las
conductas en función al contexto conductual predominante donde estas respuestas
se desencadenan, definirían la calidad de hábitat y por lo tanto su uso por parte de la
especie.
¿Cumple el Golfo Dulce con los requerimientos de hábitat en términos de
disponibilidad de recursos que el delfín manchado pantropical necesita para su
permanencia?
1. b Hipótesis El Golfo Dulce es un hábitat adecuado que sustenta la permanencia del delfín
manchado pantropical debido a que este cumple con los requerimientos
indispensables para la supervivencia de esta especie.

16
1. c Objetivo general Analizar el uso de hábitat de Stenella attenuata con el fin de determinar los
requerimientos necesarios que aseguran su permanencia en el Golfo Dulce,
Península de Osa, Costa Rica
1.c.1 Objetivos específicos
• Determinar la abundancia relativa de la población de Stenella attenuata en el
Golfo Dulce.
• Describir las pautas conductuales de Stenella attenuata.
• Caracterizar el hábitat según el uso por parte de Stenella attenuata.
1. d Justificación El análisis y la evaluación de hábitat no son fines en si mismos sino que su
valor recae en la utilidad para el manejo. Golfo Dulce es un ecosistema marino -
costero clave donde confluyen atributos ambientales neríticos y oceánicos. La
productividad es en apariencia limitada, típica de ambientes con ausencia de
surgencias. Sin embargo, sus aguas supuestamente oligotróficas albergan una
población de depredadores pelágicos importante. La presencia y la utilización de
hábitat del delfín manchado pantropical es una interrogante. Se han identificado
áreas de presencia significativas de la especie dentro del Golfo Dulce (Acevedo y
Buckhart 1998, Cubero-Pardo 1998ab, 2007, Oviedo 2007) que pueden ser
acordadas como prioridad para la investigación y el manejo intensivo.
En el proceso de evaluación de hábitat, el muestreo sistemático del
comportamiento puede proporcionar la información clave en las estrategias y el tipo
preferido de la presas en el aprovisionamiento (epipelágicas contra demersales), así
como ayudaría a determinar el tiempo dedicado a la alimentación y la inversión
energética relacionados a variables temporales y ambientales. Pocos modelos de

17
uso de hábitat consideran las respuestas dinámicas condicionadas a cambios en
factores ambientales, más aun en estudios ecológicos concernientes a mamíferos
marinos (Kaschner 2004), por lo tanto solo aportan una visión estática de la
distribución geográfica. Este estudio al incorporar la respuesta etológica, la cual
indirectamente es una medida del efecto del gradiente de recursos tróficos (Hastie et
al. 2004), convierte un enfoque empírico, en uno mecanicista que relaciona el uso del
hábitat a factores causales y dinámicos inherentes a la auto-ecología de la especie
en estudio (Guisan y Zimmermman 2000).
La información disponible sobre el Golfo Dulce, no permite la puesta en
práctica de modelos eco-tróficos que permitan una visión y predicción de eventos
estocástico potenciales. Esta investigación pretende recoger información en el área
de estudio que se pueda utilizar en modelos más exactos y más actualizados del
ecosistema en el futuro cercano, con la meta de comprender la diversidad de
relaciones y funciones de los organismos dentro del ecosistema. De hecho, un
enfoque ecosistémico que incluya la descripción cuidadosa de la naturaleza y de la
escala de las interacciones tróficas en hábitats marinos es necesaria para el
planeamiento riguroso de la conservación de los mismos (Hooker et al. 2002). La
mejor herramienta para optimizar y crear políticas de manejo es el conocimiento de
aspectos claves de la ecología de especies (consumo energético, reproducción,
interacciones sociales inter.-específicas e intra-específicas) y los hábitat que podrían
estar vinculados a la principal actividad económica de la nación: eco-turismo, más
aun cuando las acciones centradas a desarrollar la industria de la observación
turística de cetáceos están en una etapa inicial.
1.e Antecedentes
En Costa Rica, la investigación en el orden cetácea se ha centrado sobre todo
en aspectos de la biología de dos especies residentes y una migratoria: el delfín nariz
de botella, T. truncatus, (Acevedo-Gutierrez 1996, Acevedo- Gutierrez y Burkhart
1998, Cubero-Pardo 1998a, 2007), el delfín manchado pantropical; con particular

18
énfasis en la sub-especie costera, S. attenuata graffmani (Cubero-Pardo 1998a,
2007, Rodríguez Sáenz y Rodríguez-Fonseca 2004, May-Collado y Morales 2005), y
la ballena jorobada Megaptera novaeangliae (Acevedo y Smultea 1995,
Calambokidis et. al. 2000, 2002, Rasmussen et. al. 2001, 2003, Oviedo y Solis 2008).
Los estudios en los delfines nariz de botella indican que en algunas áreas
como el Golfo Dulce, estos tienen patrones muy restrictivos de movimiento y por lo
tanto dependen del hábitat local para reproducirse y para alimentarse (Acevedo-
Gutierrez y Burkhart 1998, Cubero-Pardo 1998a,b, 2007, Pacheco y Oviedo 2007).
Estudios recientes indican que las ballenas jorobadas observadas en diversas
estaciones del año, a lo largo de la costa del Pacífico de Costa Rica, están
emigrando de las áreas de aprovisionamiento alimentario de California (Enero-Abril)
y de la Península de la Antártica (Junio a Octubre) para criar y para dar a luz en
aguas Costarricenses (Steiger et. al. 1991, Calambokidis et. al. 2000, 2002,
Rasmussen et. al. 2001, 2003). En base a lo anterior, se han identificado los
parámetros ambientales relevantes en la determinación del hábitat crítico de
reproducción y cría de M. novaeangliae en aguas de la Península de Osa (Oviedo &
Solís en prensa).
El delfín manchado pantropical es probablemente el cetáceo odontoceto más
extensamente distribuido y sin duda el más representativo de las aguas territoriales
de Costa Rica. Los registros en la especie son abundantes, con un período de más
de 20 años de colecta de datos de avistamientos (May et. al. 2005). En el norte
Costa Rica, la mayoría de los registros vienen del Golfo de Papagayo,
particularmente de la Bahía Culebra y de Islas Murciélago (525 avistamientos, para
12.311 individuos). En la zona meridional del Pacífico Costarricense los primeros
avistamientos son divulgados por Acevedo (1996) y Acevedo & Buckhart (1998) en la
cuenca del Golfo Dulce (200 avistamientos). La información disponible sugiere que
los factores ambientales en las costas de Pacífico norte de Costa Rica, tales como
transparencia del agua y las concentraciones de oxígeno disueltos desempeñan un

19
papel importante en la abundancia de la población local de los delfines manchados
costeros. Además, las actividades de alimentación de esta población se asocian a la
estacionalidad del área (May-Collado y Morales 2005).
Recientemente, un mapa de la distribución de los delfines manchados
pantropicales presentado por May et al. (2005), sugirió un estatus desconocido a
nivel de subespecie para los delfines manchados avistados en Golfo Dulce y la costa
Pacífica meridional. S. attenuata, en esta cuenca, parece ser morfológicamente
similar y con el mismo patrón de manchas de S. a. graffmani en la bahía de Drake (el
Pacífico meridional abierto), sin embargo pareciese estar asociado con la batimetría
típica de los delfines manchados de hábitat oceánicos S. attenuata attenuata. A
pesar de que el conocimiento en la ecología de mamíferos marinos en aguas
costarricenses se ha incrementado notablemente en la última década, todavía hay
vacíos de información relevantes para propósitos de conservación; tales como
patrones de abundancia, utilización de hábitat e incluso taxonomía regional de
morfotípos locales potenciales.
En términos específicos del Golfo Dulce, la distribución de delfines
(manchados y nariz de botella) en relación a la profundidad y a las características
fisiográficas del golfo fueron consideradas previamente por Acevedo y Burkhart
(1998), y Cubero-Pardo (1998a,b, 2007), quienes relacionaron la distribución de los
delfines manchados con aguas profundas y el correspondiente aumentó de la
distancia desde la orilla. Mientras que el delfín nariz de botella fue asociado con
niveles bajos de salinidad y batimetría. Observaciones previas en el área de estudio
aportan detalles sobre la influencia de una isobata particular (100 brazadas) asociada
a una característica fisiográfica que condiciona el uso del hábitat por parte de estos
delfines y promueve la co-existencia (Oviedo 2007).
Al examinar la distribución espacial de S. attenuata en la cuenca interna del
Golfo Dulce, se aprecian consideraciones claves a ponderar: a) Las cualidades
ambientales del Golfo Dulce, se asemejan más a un ecosistema de aguas oceánicas,

20
más que a un mar interno. b) Los delfines manchados en el área del estudio
pareciesen agregarse usando la isobata de 100 brazas como referencia batimétrica.
Delfínidos, sobre todo las especies de hábitos teutívoros, entre ellos S. attenuata con
una composición de dieta que incluye al calamar en proporciones considerables
(Robertson y Chivers 1997; Wang et. al. 2003) tienden a tener éxito en aguas
oceánicas más allá de las isobatas de transición que delimitan el borde de la
plataforma continental, ése es el caso en el Mar Caribe y el Golfo de México (Davis
et. al. 2002; Mignucci et. al. 2003), así como en otras localidades del Pacífico como
Hawaii (Lammers et. al. 2000).
Escorza Trevino et. al. (2002) encontró evidencias de seis unidades regionales
distintivas de manejo en poblaciones costeras de S. attenuata en América Central
(entre estas una forma costera distintiva en las aguas de Costa Rica). La carencia en
definición de formas sub-específicas reportada por May et. al. (2005) para los
delfines manchados en la costa Pacífica meridional, incluyendo Golfo Dulce,
pareciese dejar abierto la interrogante sobre una forma geográfica distinta de S.
attenuata, posiblemente una forma con las características morfológicas de S. a.
graffmani, pero con los hábitos pelágicos de S. a. attenuata; como la agregación en
grupos grandes y la asociación con aguas profundas.
Cubero-Pardo (1998b, 2007) hizo un estudio específico sobre el
comportamiento (incluyendo aspectos relacionados a la alimentación) de ambas
especies del delfines en el Golfo Dulce. Sin embargo, este autor reportó apenas un
6% de tiempo dedicado a la alimentación para S. attenuata, y 10 % para T.
truncatus. Oviedo (2007) registró observaciones de comportamiento alimentario que
parecen evidenciar una proporción importante de eventos de alimentación, el 40% de
los avistamientos de delfines manchados mostraron conductas de alimentación. Sin
embargo, ambos estimados no son contrastables, ya que los registros de Oviedo
(2007) no se pueden asumir como proporciones de un presupuesto conductual; al
derivarse estas de observaciones ad libitum. Montero (2005) registró una proporción
más elevada de comportamiento alimentario para S. attenuata en la bahía de Drake,

21
62 % colectado con el muestreo por barrido (Altman 1974). García y Dawson (2003)
en Bahía Honda, Panamá, también documentaron una frecuencia más elevada de
eventos de alimentación (38%) en delfines manchados con el mismo método.
Inicialmente la baja frecuencia de eventos de alimentación dentro del Golfo
Dulce, en adición a la supuesta baja productividad de este ecosistema, motivó la
hipótesis de que el Golfo Dulce era utilizado como área de refugio más que una zona
de alimentación, por lo menos para los delfines manchados pantropicales. Sin
embargo, aunque tal asunción no debe ser desechada en el área de estudio. Las
observaciones recientes indican que la captura de presas y la alimentación pudiesen
ser más relevantes para S. attenuata en el Golfo Dulce que lo documentado
previamente. Un aspecto interesante de las observaciones preliminares, es la
relación evidente de eventos de alimentación con los ciclos del nivel de marea. Las
mareas de flujo y reflujo pareciesen proveer de ventajas en la captura de presas.
Esto sería lógico para T. truncatus, ya que la agregación de esta especie se asocia
comúnmente a las bocas de ríos (Cubero – Pardo 2007, Pacheco y Oviedo 2007).
Pero también fue observado en avistamientos de S. attenuata donde se registraron
eventos de alimentación en áreas relativamente llanas como el río de Esquinas y
otras zonas en la costa suroeste de la cuenca interna del Golfo (Oviedo 2007).
1. e.1 Marco socio-económico relacionado con las perspectivas de manejo
De acuerdo a Sierra et al. (2006), de una población de 6.102 habitantes del
distrito de Puerto Jiménez en el cantón de Golfito, 300 habitantes (mayormente
hombres > 95%) se dedican a la pesca, 814 personas (55% mujeres, 45% hombres)
están empleadas en hoteles y restaurantes, mientras que 533 (Hombres >90%)
laboran en transporte y comunicaciones. La fuerza de trabajo se caracteriza por una
taza de desempleo más alta que la nacional. Esta fuerza de trabajo se vio
particularmente afectada con la culminación de actividades de la compañía
bananera, sin embargo, el cultivo de la palma y el arroz han alivianado la situación de
desempleo, ya que el 34% de la población esta empleada en el sector agropecuario.

22
No obstante, no hay estabilidad en cuanto a la demanda de empleo, y la misma
presenta variaciones anuales significativas. Un ejemplo es la demanda laboral en el
sector turismo; el cual a pesar de haber aumentado en la última década, tiene un
carácter muy estacional y por lo tanto inestable.
La pesca en ACOSA tiene poca trascendencia en el abastecimiento nacional,
de 4.823 embarcaciones inscritas y no inscritas en Costa Rica, el 6% de las
registradas se localizan en ACOSA. Para el año 2000 INCOPESCA contaba con 355
embarcaciones en orden, de las cuales el 77% se dedicaban a la pesca artesanal y
el resto (27%) a la pesca deportiva. La pesca deportiva, la cual esta en aumento y se
lleva a cabo en áreas especiales como Golfo Dulce y aguas abiertas, la mayoría de
la captura de esta operación es liberada (sin evidencias de que permanecen vivas).
La actividad turística, y la correspondiente necesidad de establecer
infraestructura, han evolucionado desde la década de los 80, para ese entonces las
actividades de turismo ecológico estaban dirigidos a eco-naturalistas duros, y los
requerimientos de habitación eran básicos sin estrellas ni comodidades. En la
década de los noventas se incluyeron una gran porción de no solo eco-naturalistas
suaves, sino también de un mercado receptivo de turismo de sol y playa, por lo que
los requerimientos en el alojamiento incrementaron las comodidades y la
categorización se ubicó entre una a cinco estrellas. Finalmente la década del 2000
diversificó aun más el mercado incluyendo turismo de salud y belleza, turismo de
aventura y turismo cultural, aparte de las modalidades de turismo natural.
Aparecieron además ofertas hoteleras de entre cuatro y cinco estrellas destinadas a
operadores locales y extranjeros. Para 1999 – 2000, se estimó una oferta de 1600
habitaciones para los cantones Golfito y Osa, con el 72% de estas en el Cantón de
Golfito (Sierra et al., 2006).
Los impactos negativos de este crecimiento se relacionan con la carencia en
sistemas efectivos de tratamiento de aguas servidas y una infraestructura sanitaria
poco óptima. Otro aspecto importante de resaltar es el dominio del capital extranjero

23
en sector turismo de la región, relacionado con la poca posibilidad de adquirir
terrenos y propiedad de los nacionales por el alto costo de la tierra. También la falta
de preparación local en experiencias turísticas y el manejo del idioma Ingles, hace
perentorio la asistencia extranjera en las operaciones turísticas locales.
Sierra et al. (2006) hace especial mención sobre el deterioro de los factores
económicos y sociales en conjunto con los recursos naturales promovido por el
predominio de una economía extractiva, la falta de industrialización, la baja demanda
de empleo y la taza alta de desempleo. Lo anterior tiene implicaciones directas en la
extracción ilegal de recursos para la subsistencia del sector más afectado
económicamente de la población, y trae como consecuencia; que las áreas
protegidas y las poblaciones silvestres que estas albergan estén gravemente
amenazadas por las condiciones imperantes fuera de sus límites.
1. e.2 Contexto legal de los cetáceos como recursos en Costa Rica.
El manejo, control, regulación de la caza marítima, la investigación;
conservación y protección de los recursos marinos de Costa Rica le corresponde al
Instituto Nacional de Pesca INCOPESCA. También es obligación de este ente
gubernamental el fomentar la utilización sostenible de dichos recursos, bajo estrictos
criterios técnicos y científicos, como lo establece la Ley No 7384.
Los cetáceos como recurso marino son expresamente protegidos por la Ley de
Pesca y Acuicultura No 8436, específicamente en el artículo 39, capitulo VI el cual
dice:
“Prohíbanse la caza marítima, la captura de cetáceos, pinnípedos y quelonios, así
como el aprovechamiento de sus lugares de cría, salvo lo establecido en los
convenios o tratados internacionales debidamente ratificados por Costa Rica”.
Al artículo citado anteriormente se le adiciona el Reglamento Para la Operación de
Actividades Relacionadas con Cetáceos en Costa Rica No 32495, publicado en La

24
Gaceta el 28 de Julio del 2005. Este reglamento consta de 21 artículos que norman
la observación directa de cetáceos silvestres en campo, tanto a nivel comercial como
científico. También se establece la prohibición expresa sobre la tenencia en
cautiverio de estos mamíferos marinos; con las correspondientes excepciones para
casos de varamiento que requieran del tratamiento del animal en condiciones ex-situ.
Otras provisiones legales que se derivan o relacionan con acuerdos
internacionales, establecen discrecionalidad directa sobre los cetáceos:
• La Convención sobre el Comercio Internacional de Especies Amenazadas
(CITES) de flora y fauna silvestre incluye a las especies de cetáceos a nivel
mundial dentro de los apéndices 1 y 2, lo que implica que las 29 especies de
cetáceos que están presentes en aguas de Costa Rica son protegidas por
este convenio; por lo tanto su uso y comercio esta regulado por la Ley de
Conservación de Vida Silvestre. Esta ley asigna al MINAE competencias para
el otorgamiento de permisos de vida silvestre y de todas aquellas especies
que se encuentren en las listas de los anexos de CITES.
• Costa Rica fomenta el Acuerdo sobre el Programa Internacional para la
Conservación de los Delfines entre la República de Costa Rica y Estados
Unidos de América, bajo la Ley Nº 7938.
• Costa Rica es signatario desde 1982 de la Comisión Ballenera Internacional,
la cual actualmente vela internacionalmente por la actividad de observación de
delfines y ballenas.
• En relación a los aspectos inherentes a la pesca de atún, las provisiones
relacionadas a cetáceos se rigen por lo a lo estipulado en la Convención de la
Comisión Interamericana del Atún Tropical (CIAT).

25
MARCO TEÓRICO
2.a Uso de Hábitat
De acuerdo a Krausman (1999) y Ojasti (2000), el concepto del hábitat es una
piedra angular en el manejo de la fauna. Las decisiones sobre manejo del hábitat
serán de poco valor a menos que los biólogos primero determinen los patrones del
uso del hábitat de un animal dentro de un ambiente específico y consideren los
disturbios evolutivos y humanos que lo influencian. Los hábitat son los recursos y las
condiciones presentes en un área que producen la ocupación, incluyendo
supervivencia y la reproducción por un organismo dado. El hábitat implica más que la
vegetación o la estructura de la vegetación. Es la suma de los recursos específicos
que son necesitados por los organismos. Estos recursos incluyen el alimento,
cubierta, agua, y los factores especiales necesarios para la subsistencia de una
especie y el éxito reproductivo de la misma. Dondequiera que a un organismo se le
proporcione los recursos que permiten que sobreviva, ése es su hábitat. Así, los
corredores de migración y de dispersión, así como las zonas que los animales
ocupan durante estaciones de crianza y no-crianza son hábitat (Krausman 1999).
El uso del hábitat es la manera que un animal utiliza los recursos físicos y
biológicos en un hábitat (Krausman 1999). El hábitat puede ser utilizado para
aprovisionamiento, refugio, anidación, escape, u otros rasgos inherentes a la historia
de vida de la especie. Estas categorías dividen el hábitat zonificándolo, pero el
solapamiento puede ocurrir en algunas áreas. Unas o más categorías pueden existir
dentro de la misma área, pero no es una condición única y necesaria, ya que las
varias actividades vitales de un animal requieren de componentes ambientales
específicos que pueden variar sobre una base estacional o anual. El uso de hábitat
implica un proceso de discriminación, la selección del hábitat es por lo tanto un
proceso de comportamiento activo de un animal. Cada especie busca de sustentarse
en medio de las características dentro de un ambiente que se asocian directamente o

26
indirectamente a los recursos que un animal necesitaría para reproducirse,
sobrevivir, y persistir (Krausman 1999, Ojasti 2000).
La selección del hábitat es una compilación de los comportamientos naturales
y aprendidos que se encuentran en una serie continua programas genéticos. El éxito
reproductivo y la supervivencia de la especie son las razones de mayor importancia
que influencian a determinada especie para seleccionar un hábitat. Rosenwieg
(1981) sustenta que el aprovisionamiento es el factor determinante en la selección de
hábitats. Sin embargo, el aprovisionamiento es solamente uno de los
comportamientos que conduce a la selección del hábitat. El hábitat se puede
seleccionar según la disponibilidad de refugio, calidad y cantidad de
aprovisionamiento, y sitios de reproducción. Cada uno de estos factores puede variar
estacionalmente.
Si un individuo o una especie demuestran el uso no proporcional de cualquier
factor, entonces la selección se deduce en base a esos criterios. Varios factores que
interactúan recíprocamente tienen una influencia en la selección del hábitat para un
individuo (competición, refugio, y depredación). La competición está implícita porque
cada individuo esta involucrado en relaciones intra - específicas e inter.-específicas
que dividen los recursos disponibles dentro de un ambiente. La competición y la
depredación pueden determinar que la especie no pueda seleccionar un hábitat
conveniente en abundancia de otros recursos necesarios o puede determinar la
distribución espacial dentro del hábitat (Ricklefs 2001, Begon et al., 2006).
El análisis de hábitat implica el estudio de los ambientes presentes y sus
patrones espaciales, es inherente al cuestionamiento sobre cuan adecuada es un
área y sus ambientes para la fauna local. La evaluación de hábitat relaciona los
requisitos ecológicos de las especies presentes (o que habitan ambientes similares
en áreas cercanas) con las condiciones de sus respectivos hábitat. Caracterizar un
hábitat no es lo mismo que describir los ambientes en un área, ya que el caracterizar
se parcializa por las necesidades intrínsecas de la especie (espacio, cobertura,

27
alimento y agua), es decir por los recursos necesarios para la sobrevivencia de la
especie (Krausman 1999, Ojasti 2000).
Un recurso puede definirse como un elemento particular requerido y utilizado
por los organismos; puede variar en calidad y cantidad, ser escaso o incluso
agotarse (Ricklefs 2001). Entre los recursos mas determinantes tenemos la cobertura
y el alimento. Para prosperar en un hábitat una población requiere tanto de cobertura
como de alimento, coexistentes o ubicados en parches aledaños. La evaluación de
disponibilidad de alimento se solapa con el levantamiento de la cobertura. Sin
embargo las características de la cobertura salta a la vista, mientras que los
elementos inherentes al alimento pueden ser más escasos y ocultos (Ojasti 2000)
2. b Alimento
De acuerdo a Ojasti (2000) el alimento se cuantifica en términos de producción
(tasa de cambio de biomasa por unidad de tiempo o espacio), existencia o
abundancia (cantidad presente en el momento por unidad de área), disponibilidad
(cantidad instantánea al alcance efectivo de un cierto tipo de animal por unidad de
área) y la utilización (tasa de consumo de un reglón durante un lapso de tiempo). La
abundancia de una especie de presa para un depredador particular podría ser
cuantificada, sin embargo no siempre esta cuantificación es completa, ya que no toda
la presas en el hábitat están disponible al depredador porque puede haber factores,
(como disponibilidad de cubierta amplia) que restringen su accesibilidad (Krausman,
1999).
El alimento expresado en cualquiera de sus formas cuantificables, representa
el principal gradiente de recursos que condiciona la presencia o no de una especie
(Guisan y Zimmermman 2000), y generan respuestas ante cambios en la calidad y
cantidad del mismo, esto es particularmente relevante en la distribución de especies
de cetáceos como el delfín nariz de botella (Hastie et al, 2004).

28
2.b.1 La teoría del consumidor óptimo
La teoría del consumidor optimo (Begon et al., 2006) pondera la estrategia de
alimentación más eficaz en determinadas condiciones específicas, con el fin ultimo
de incrementar la aptitud del organismo en términos de sobrevivencia y mediante la
optimización del consumo de energía. Bajo el enfoque de esta teoría, se espera que
el consumo promedio de energía sea el de mayor valor nutricional y el que promueve
la aptitud. Por lo tanto, la ecuación:
E /(S+H) (1)
Representa el cociente entre el valor nutricional “E ” y el esfuerzo constituido por la
búsqueda “S ” y el manejo del alimento “H ”.
Si equiparamos la búsqueda con la actividad de movilización (M ) para el caso
de S. attenuata en Golfo Dulce, y la manipulación con la actividad de alimentación
(A) en el contexto de captura y consumo de presa, entonces generamos un modelo
mecanicista que nos permite predecir la estrategia de conducta alimentaría esperada
en dos situaciones básicas:
Conducta alimentaría especialista: el esfuerzo de captura de la presa se constituye
básicamente en una mayor inversión de tiempo y energía en captura y consumo
selectivo, en contraposición a la movilización por búsqueda. Por lo que la ecuación 1
se establecería bajo las siguientes condiciones: 0 ≤ M < A ≤ E (1a). El valor menor de
A en relación a E, maximizaría el cociente entre el valor nutricional y el esfuerzo
Conducta alimentaría generalista: el esfuerzo de captura se constituye de mayor
adjudicación de tiempo y energía a la movilización por búsqueda de presas, siendo
mucho menor la captura y el consumo selectivo. La ecuación 1 se plantearía bajo la
condición siguiente: 0 ≤ A < M ≤ E (1b), una vez más el cociente entre el valor
nutricional y el esfuerzo es optimo por el menor valor de M en relación a E.

29
Las situaciones 1a y 1b ocurren asumiendo que la presión de depredación
sobre los delfines manchados es mínima y despreciable, al igual que el efecto
negativo de la competencia intra-específica y inter-específica con delfines nariz de
botella (T. truncatus). Los implícitos que se asumen anteriormente poseen algún
sentido empírico. No hay evidencias sustanciales de depredadores como tiburones
atacando delfines en Golfo Dulce (sin que esto implique la ausencia de
elasmobranquios) y los niveles críticos de competencia inter-especie se minimizan
por repartición de habitat (Bearzi 2005 a,b, Oviedo 2007).
2. c Cobertura en el medio marino
La definición de cobertura en medio marino, en contraposición con el caso de
análisis de ecosistemas terrestres no esta determinada por factores relacionado a la
vegetación, sino a disposición de elementos físicos como la batimetría y topografía
en correlación a otras variables como la temperatura y salinidad (Ricklefs 2001).
Ojasti (2000) definen la cobertura como un factor del hábitat que alberga a los
animales y los resguarda de extremos climáticos y de depredadores (incluyendo al
hombre).
La oferta de alimento para los depredadores de vertebrados es obviamente
proporcional a las de sus respectivas presas, por lo que factores que afectan
directamente la distribución de presas potenciales como productividad primaria,
temperatura y cobertura ejercen un efecto indirecto en la permanencia de dichos
depredadores en un hábitat. La depredación es siempre una interacción entre
poblaciones por lo que el cociente abundancia de presas / abundancia de
depredadores es fundamental en el estudio de poblaciones silvestres. Los carnívoros
tope son considerados como indicadores de la salud de un ecosistema, aunque estos
pueden consumir una porción sustancial de la producción de presas silvestres e
incluso domesticas. Las presas además de sufrir mortalidad, pueden confinarse en
coberturas protectoras y quedar privadas de los demás hábitat y recursos por efecto
de la presión ejercida por los depredadores (Ojasti 2000).

30
A mayor diversidad de hábitat se puede esperar una mayor diversidad
fáustica, es decir mayor diversidad β, la cual expresa la variedad de organismos en
una región como resultado de un recambio de especies entre hábitat (Ricklefs 2001).
El impacto ecológico de los límites, eco-tonos o fajas de transición entre dos tipos de
hábitat sobre las poblaciones animales es denominado efecto borde. Una especie
que es localiza en el borde tiene acceso simultaneo a dos tipos de hábitat, el efecto
borde depende de los tipos y extensión de hábitat colindantes. Se podrán apreciar
dos tendencias; si los hábitat colindantes son parte del ámbito de distribución de la
especie se espera un efecto positivo, ya que la especie requiere más de un tipo de
hábitat para su permanencia y sobrevivencia. La tendencia contraria implica que la
diferencia de habita representa un limite o barrera que se reflejará en el ámbito de
distribución de dicho organismo.
2.d La Abundancia
También se define como cantidad de individuos o de biomasa, es un atributo
poblacional variable en el tiempo y en el espacio. Esta variable es de singular
importancia en el manejo de la fauna silvestre. Su estimación es crucial para el
manejo práctico porque:
• Indica el estado de una población en un momento dado.
• Permite compararla con otras poblaciones.
• Es útil como criterio de evaluación de la calidad de hábitat.
La estimación de abundancia es una herramienta versátil y valiosa en la toma de
decisiones, en el seguimiento de planes de manejo y es un criterio rector en las
investigaciones sobre dinámica de poblaciones. Por medio de la abundancia y de la
evaluación de hábitat se puede apreciar la existencia particular de meta-poblaciones
en un área particular (Ojasti 2000). Una meta-población es un conjunto de
poblaciones semi-aisladas con cierto grado de dispersión entre poblaciones locales

31
(Ricklefs 2001). En el evento de una extinción local, las meta-poblaciónes ofrecen la
posibilidad de recolonización del área vacante por las poblaciones vecinas.

32
MATERIAL Y MÉTODOS 3.a Área de Estudio
Golfo Dulce (8º 30’ N, 83º16’W) (Figura 1) es un mar interno de origen
tectónico usualmente denominado fiordo tropical. El Golfo Dulce tiene 50 Km. de
largo y entre 10-15 Km. de ancho aproximadamente; el área total esta calculada en
682 Km2 (Quesada et al. 2007).
Este mar interno se subdivide en dos sub-áreas importantes: una cuenca
interna profunda de pendiente inclinada escarpada, con un fondo plano de
aproximadamente 215 m de profundidad máxima, y una cuenca externa llana con
una profundidad de 70 m (Wolff et al. 1996, Quesada et al. 2007). La línea costera
dominante en el norte es litoral rocoso escarpado con abundancia de bosque tropical.
En el sur se presentan playas arenosas, particularmente en la zona sureste,
específicamente en la Bahía de Pavones, así como al suroeste desde Playa
Platanares hasta Cabo Matapalo.
Svendsen et al. (2006) consideran que la circulación del Golfo esta
condicionada por la morfología tipo fiordo y en cierto grado por los vientos locales,
estos inducen gradientes de velocidad en la capa de agua superficial, mientras que
los gradientes de temperatura y salinidad serían relativamente constantes a lo largo
del mismo (salinidad baja y una temperatura alta), por el aporte fluvial de cuatro ríos;
Tigre, Rincón, Esquinas y Coto, en su parte interna. Debido a la cercanía con el
ecuador, el efecto de la fuerza de Coriolis sobre la circulación se considera
despreciable. La circulación sigue un sentido anticiclónico (en dirección a las agujas
del reloj) y el flujo es hacia la zona externa del Golfo.
Hay una gama de mareas considerable: 2-3 m aproximadamente, con una
gran influencia en el litoral y los cursos de agua dulce. Quirós (2003) adiciona como
factor relevante en la circulación las mareas. El transporte de agua, tendría un
máximo durante la media marea, con una distribución desigual entre el umbral y la

33
fosa interna del golfo: en el tercio exterior se tienen flujos del orden de los
15.000m3/s. En el canal principal sobre el eje de la fosa, los flujos se caracterizan por
una velocidad entre 10 a 20 veces menor.
Figura 1. Área de estudio, Golfo Dulce, Área de Conservación de Osa (ACOSA), Puntarenas, Costa Rica.
Aproximadamente 200 hectáreas de manglares están asociadas a los ríos que
fluyen al golfo. La productividad es aparentemente baja, con niveles nutrientes
pobres cerca de la superficie, típica de aguas con ausencia de procesos de
surgencia. De acuerdo a Wolff et al. (1996), Golfo Dulce sigue siendo un estuario
prístino que actúa diferente a la mayoría de los ecosistemas costeros tropicales, al
ser dominado por una biomasa y un flujo de energía dentro del dominio pelágico, con
semejanzas mayores a un ecosistema oceánico que a un estuario.

34
3. b Muestreos en Campo La colecta de datos comprendió 157 horas en campo, en dos períodos claves;
la temporada de lluvias 2007 y la temporada seca 2008. Se utilizaron muestreos por
recorridos sistemáticos en embarcaciones pequeñas, los cuales se centraron en
áreas de presencia significativa, determinadas previamente en los pre-muestreos de
esta investigación, basado en lo descrito por Acevedo & Burkhart (1998) y Cubero
Pardo (1998a, 1998b, 2007).
A lo largo de las zonas de alta densidad referidas anteriormente se siguió la
misma ruta de muestreo, partiendo del mismo punto, hasta que se iniciase el
encuentro con el grupo de delfines a seguir. Este protocolo permitió un muestreo
completo de la cuenca interna de Golfo Dulce traducido en un esfuerzo espacial
homogéneo.
La fuente principal de datos de este estudio fueron los seguimientos grupales,
con la finalidad de cuantificar una unidad de esfuerzo durante y antes de los
seguimientos, se totalizaron el número de estaciones ambientales (Gowans &
Whitehead 1995), registradas durante cada muestreo y seguimiento grupal: cada 30
min, se colectaron datos sobre: temperatura superficial del mar, fuerza del viento; por
medio de la escala Beaufort, y localización espacial en coordenadas geográficas
mediante lecturas del GPS. Por lo tanto, cada muestreo inició con la toma de la
primera estación ambiental. Como medida relativa del esfuerzo en campo y de
presencia de la especie en el área de estudio, se calcularon índices de frecuencia de
avistamiento (FA= No Avistamiento/ esfuerzo), para cada 100 Km recorridos y por
horas de búsqueda invertidas.

35
3. c Abundancia relativa La estimación de la abundancia relativa de S. attenuata en el área de estudio,
no siguió la metodología de transectos lineales, ya que logísticamente no era
compatible en términos de costos/beneficio con los muestreos etológicos. Por lo
tanto, se recurrió a la estimación de índices de abundancia relativa, alternativos y
compatibles con el protocolo de seguimiento grupal.
En base a lo anterior, dos índices se presentan:
1) un índice de abundancia relativa que considera la totalidad de individuos
observados durante el muestreo en campo, en función a una unidad de esfuerzo
(APUE), dicha unidad de esfuerzo es la totalización del número de estaciones
ambientales (Gowans & Whitehead 1995), registradas durante cada muestreo y
seguimiento grupal.
2) la abundancia relativa en base a un área de detección efectiva (Henrys 2005), este
índice es una adaptación de algunos de los factores del método de transectos
lineales para datos obtenidos por muestreos no azarosos, modificados en función al
protocolo de seguimiento grupal, el cual considera:
a) la distancia de espera (li); que es la distancia desde el comienzo del muestreo
(primera estación ambiental) al avistamiento inicial del grupo a seguir, si durante el
protocolo de seguimiento se debe reiniciar el mismo por avistamiento de un nuevo
grupo (lo que implica pérdida del grupo seguido anteriormente), se debe calcular una
nueva distancia. La distancia de espera es un factor novel para el cálculo de
abundancia relativa con el programa “Distancia” (Distance) usando datos de
recorridos, que por relación costo/beneficio no siguen las reglas del azar.
b) la distancia de detección (μ), la cual es fija (μ= 500 m) debido al protocolo de
seguimiento y al método de muestreo etológico, los cuales requieren de una
observación relativamente cercana, que permita el registro efectivo de conductas.

36
c) el estimado de densidad del área de estudio (D = ns/as) donde “ns” es el número
total de individuos observados durante los seguimientos grupales, y “as” es el área
total de seguimiento, este estimado es particularmente aplicable a áreas bien
delimitadas como golfos y bahías, para el caso particular de esta investigación, este
valor se calculó con ayuda de un SIG (ArcGIS 9.2), y corresponde a 343 Km2 de la
cuenca interna de Golfo Dulce.
La estimación requiere primeramente establecer el cálculo del área de
detección, mediante la relación de la distancia de espera con la distancia de
detección en la expresión li2(μ), la cual genera el área de detección de cada
seguimiento grupal. En la fómula de densidad se ingresa el valor de cada área de
detección calculada y se obtiene un estimado de abundancia (ns=Dxli2(μ)) para cada
seguimiento grupal, por medio de un bootstrap no-paramétrico (100.000 repeticiones,
95% de confianza) se obtienen los intervalos de confianza. Las diferencias entre
estimados de abundancia, y entre estimadores como la distancia de espera para la
temporada de lluvia y la temporada seca se definió con la prueba no-paramétrica
“Kruskal Wallis”, ya que los datos generados no siguieron un comportamiento normal
(Zar 1996), el nivel de significancia de la prueba 95%, lo establece el paquete
estadístico (Analyze It versión 2,09).
A partir del estimado de abundancia relativa para la población de S. attenuata,
en Golfo Dulce, se calculó un índice de densidad, considerando el área efectiva de
distribución de los delfines manchados durante el período de observaciones en la
temporada de lluvia (143Km2). De manera similar se calculó el mismo valor para S.
attenuata en Golfo de Papagayo, en base a las observaciones registradas en May -
Collado (2001) con la finalidad de establecer comparaciones.
El consumo de biomasa por delfín, por día, se estimó de acuerdo a Read y
Brownstein (2003) con la expresión:
IB (Biomasa Ingerida)= 0,123M0,80

37
Para la ecuación anterior, “M” representa la masa corporal del delfín, usando como
parámetro clave la correlación entre el peso y la longitud total para S. attenuata
(Perrin 2001). Se calculó el consumo para las poblaciones de delfines manchados en
Golfo Dulce y Golfo de Papagayo, extrapolando el estimado con el índice de
densidad anteriormente mencionado.
3. d Muestreo etológico El muestreo etológico de esta investigación se realizó por medio de censos
sistemáticos visuales desde una embarcación pequeña, que permitieron establecer
estados conductuales geo-referenciados mediante la observación directa de la
población en estudio. Esta contribución generó datos etológicos asociados
geográficamente a extensiones espaciales, por medio del protocolo de seguimiento
grupal. La técnica de registro etológico seguida fue el muestreo por barrido de grupo
focal, descrito por Altman (1974) y Mann (1999, 2000), Con el fin de minimizar el
sesgo, el barrido grupal se intercaló con un barrido constante de un individuo
positivamente identificado (GD001, anexo 1), presente en todos los seguimientos del
grupo observado.
Los períodos de observación por barridos fueron intervalos de 2 minutos,
intercalados por períodos de 5 minutos. Al inicio y al final de cada período de barrido
se registró una lectura de coordenadas geográficas inicial, y otra adicional al culminar
el período de barrido. La posición del grupo de delfines se equiparó a la posición
geográfica del bote de observación. La definición de grupo para esta contribución
considera a cualquier agregación de delfines a menos de 10 m de sus con-
específicos, y que manifiestan el mismo estado conductual. La definición anterior es
consecuente con la utilizada por Smolker et al. (1992) y Shane (1990).
La colecta de datos involucró a tres observadores, dos que realizaron el
seguimiento visual, incluyendo al investigador principal (con experiencia y
entrenamiento previo en muestreo etológico), un tercer observador documentó los
avistamientos junto con las conductas observadas de S. attenuata, por medio de

38
fotos y videos del grupo, así como de los individuos claves para el reconocimiento de
grupos específicos. Este registro fotográfico se realizó por medio de una cámara
digital sony DCH con un zoom óptico de 12X, la cual también permitió tomar
secuencias de videos como soporte a las observaciones.
Cada episodio conductual registrado se consideró independiente al efecto del
bote de muestreo durante la observación, esto se logró mediante una conducción del
bote apropiada, poco intrusiva y mediante un tiempo de familiarización de los grupos
en seguimiento con el bote, de manera tal que siguieran con sus actividades sin
signos de perturbación, una vez que los grupos continuaban sus actividades
independientes de la presencia de los investigadores, se iniciaba el muestreo por
barrido.
Los estados conductuales documentados, definidos en los anexos 2 y 3,
derivaron de una clave utilizada por García y Dawson (2003) y desarrollada por
Scheneider (1999). Estos estados conductuales son consecuentes con aquellos
analizados en otros estudios etológicos (García y Dawson 2003, Lusseau 2003) y se
consideran mutuamente exclusivos y cumulativamente inclusivos; es decir en
conjunto son descriptores óptimos del presupuesto conductual integral de cada grupo
de delfines (Lusseau y Higham 2004). Es importante mencionar que dichos estados
conductuales no consideran a los observadores (ni al bote de muestreo) como una
variable interviniente, ya que intentan hacer una aproximación a las condiciones
naturales diarias de los grupos en seguimiento.
Es importante resaltar que este tipo de muestreo conductual se ha utilizado en
otras investigaciones en áreas de estudio en el Pacífico de Costa Rica y Panamá con
la misma especie, lo que permite establecer una comparación en patrones y
proporciones conductuales con otras localidades diferentes a Golfo Dulce (García y
Dawson 2003, May-Collado y Morales 2005).

39
3.d.1 Análisis de datos etológicos
El razonamiento de los datos se basó en técnicas de análisis de ecología
espacial y conductual, mediante la utilización de tres enfoques claves; la
determinación de un presupuesto conductual (Lusseau 2003, Lusseau y Higham
2004), un análisis espacial en base al presupuesto conductual (Karczmarski et al.
2000, Lusseau y Higham 2004, Garrafo et al. 2007) y la determinación del nicho
ecológico real en base a factores eco-geográficos, por medio de un modelo empírico
con propiedades mecanicista (Hirzel et al., 2002; Oviedo 2007):
a) Los datos etológicos se analizaron por medio del cálculo de la frecuencia y
proporción de cada estado conductual utilizando estadística descriptiva,
obteniéndose un presupuesto conductual expresado en porcentaje (Mann 2000,
García y Dawson 2003, Lusseau 2003). Cada actividad registrada en el presupuesto
conductual fue transformada en un cociente (cociente de observación = Co) en base
al período de barrido. Lo anterior tiene como finalidad el cuantificar las
observaciones, de manera tal, que sean una variable continua que exprese no solo
la ocurrencia de una actividad conductual específica, sino que también son una
referencia cualitativa de la actividad. Por lo tanto un registro de barrido en un período
de 2 minutos, con una actividad “X” tiene un cociente con un rango de 0 a 1; siendo 1
un período de observación integral de la actividad “X”. Cualquier período de
observación menor a uno, representa un episodio conductual mixto en donde la
actividad “X” tiene cierto nivel de dominancia en la ocurrencia, bajo un criterio a priori
de ≥ 0,60 como dominante.
Las diferencias entre estados conductuales dominantes en una misma
temporada y entre temporadas se evaluaron por medio de la prueba no-paramétrica
Kruskal Wallis (Zar 1996), a un nivel de significancia del 95%, con el paquete
estadístico “Analyze It” versión 2,09.

40
b) El análisis espacial basado en el presupuesto conductual se realizó mediante un
SIG en donde se geo-referenciaron las 5 actividades documentadas en el
presupuesto conductual por temporadas (alimentación, movilización, socialización,
merodeo y descanso), con el fin de asociar dichas actividades con un patrón espacial
de distribución. Se procedió primeramente a establecer una matriz con la la
información referente a la localización geográfica de las observaciones (valores [x,y]
en coordenadas de grados decimales). El comportamiento observado se representó
de forma binaria dentro de la matriz; una columna por cada categoría de
comportamiento en donde 1 corresponde a la actividad conductual observada y 0
cuando la misma no fue observada. Mediante el software ArcGIS 9.2 se procedió a
generar los elementos espaciales a partir de los datos en la matriz mencionada
anteriormente. Para la representación de los datos se utilizó un formato vectorial con
geometría de puntos de localización única.
El área de estudio; la cuenca interna de Golfo Dulce se dividió en celdas de 1
min de latitud X 1 min de longitud, equivalentes a un área de 3,24 km2. En cada celda
se estimaron dos índices; el Uso de Área (UA) de acuerdo a Garrafo et al. (2007) y el
Índice de Actividad (IA) según Karczmarski et al. (2000). Ambos índices fueron
adaptados para el uso de los datos generados en los seguimientos grupales. Por lo
tanto el uso de área se determinó mediante la expresión:
UA= ΣCoX / CoGD
ΣCoX es la sumatoria de todos los cocientes de observación en la celda X, y CoGD
es suma total de todos los cocientes durante el período de muestreo.
El índice de actividad fue estimado mediante la siguiente ecuación:
IA= Coα / ΣCoX

41
Donde Coα es la totalización de todos los cocientes de observación en la actividad
conductual específica (α), en la celda “X”. Las clases de uso de área por temporadas
fueron obtenidas por medio de un análisis de agrupamiento por distancia euclidiana
al cuadrado, usando el paquete estadístico Statgraphic plus (V 5.0).
Con la finalidad de ilustrar espacialmente la información contenida en el
análisis anterior, se establecieron los polígonos de probabilidad que ilustran la mayor
ocurrencia de la actividad clave, que define el hábitat crítico (Lusseau y Higham
2004, Hoyt 2005). Se realizó la interpolación de los valores 0 y 1 descritos en la
matriz binaria, con la finalidad de ser consecuente con los cocientes de observación
generados (Co), se estableció como criterio que 1 representó una probabilidad del
100% de ocurrencia de la actividad conductual específica, y 0 representó la ausencia
de dicha conducta. Mediante el paquete geo-estadístico de ArcGIS 9.2, se utilizó el
método de interpolación del valor ponderado inverso a la distancia IDW (inverse
distance weighted) para generar mapas con los polígonos de probabilidad por
temporadas. Este método permite evaluar la probabilidad de ocurrencia de un
fenómeno a partir de observaciones, tomando como premisa, que a medida que se
aumenta la distancia entre la estimación y el punto observado, la estimación adquiere
un comportamiento inverso.
c) Para establecer la relación entre las variables eco-geográficas y la ocurrencia de
episodios de alimentación, se analizaron la profundidad máxima, pendiente,
temperatura, distancia a río más cercano, río de mayor influencia y temperatura
como las variables descriptoras más relevantes en relación a la captura y consumo
de presa. La temperatura y distancia de ríos cercanos, se agruparon en clases
mediante un análisis de agrupamiento de media de grupos, por medio de la distancia
euclidiana al cuadrado, tal como se ilustra con las figuras 11 y 12 (Statgraphic plus v
5.0). En el caso de la profundidad y la pendiente, se establecieron las clases de
pendiente (muy baja, baja, medio baja, media, medio alta, alta, y empinada) y
profundidad (aguas llanas, aguas media, aguas profundas y aguas muy profundas)

42
de acuerdo a Oviedo (2007), con el fin de estratificar el área de estudio en los
ámbitos costero y oceánico.
Se aplicó la prueba Kruskal Wallis (Zar, 1996), al comparar las sub-áreas
cercanas a la costa - bocas de ríos y área central, así como las variables asociadas
con la actividad determinante del hábitat crítico de S. attenuata. Las diferencias entre
la temporada de lluvia y la temporada seca también se determinaron con la misma
prueba no-paramétrica. El nivel de significancia para la prueba Kruskal Wallis se
estableció en 95%.
El análisis y caracterización del nicho ecológico real fue consecuente con el
realizado por Oviedo (2007), en la definición de la repartición de hábitat como
estrategia de coexistencia entre T. truncatus y S. attenuata en Golfo Dulce. Sin
embargo, al usar como datos claves los cocientes de observación de la actividad
conductual determinante del hábitat crítico, se estableció un modelo de
características mecanicistas; ya que la variable respuesta fue la conducta ante el
efecto de los factores eco-geográficos evaluados.
La profundidad máxima y la pendiente se escogieron como las variables eco-
geográficas a incluir en el análisis, ya que ambas son la que mejor definen la relación
entre las variables descriptoras de hábitat, la presencia y distribución de de cetáceos
(Hui 1985, Davis et al. 2002; Cañadas et al. 2002, 2005; , Hooker et al. 2002,
Azzelino et al. 2008, Gómez de Segura et al. 2008), y son particularmente importante
en la definición de hábitat de delfines del género Stenella (Griffin y Griffin 2003). Así
como las de mayor influencia en la distribución de S. attenuata en Golfo Dulce
(Acevedo y Burkhart 1998, Cubero Pardo 1998a).
Se determinó la topografía submarina en función a las lecturas de profundidad
máxima y mínima en una carta batimétrica del área de estudio (Carta Batimétrica No
21562, US Navy 1998) por medio del Índice de Pendiente (IP) de Mignucci-Giannoni
(1998) y modificado por Oviedo et al. (2005):

43
Donde PM y Pm son la máxima y mínima profundidad encontrada en una celda de
1,8 Km (equivalente a 1 min de latitud) por 1,8 Km (equivalente a 1 min de longitud),
mientras que PMa es la profundidad máxima del área considerada.
Dos índices de comparación de promedios y de dispersión de la muestra
permitieron establecer una relación numérica entre las variables estudiadas y los
cocientes de observación de la actividad conductual clave para la definición del
hábitat crítico; la Marginalidad y la Tolerancia (Oviedo 2007), este tratamiento
derivado del Análisis de Factores del Nicho Ecológico (ENFA) desarrollado por Hirzel
et al. (2002), especialmente minimiza el sesgo relacionado con la falta de ausencias
reales que caracterizan a los datos de presencia:
Marginalidad; M = μGD- μSa / 1.96XθGD
Tolerancia; Tol= θSa/θGD
Donde μGD es el promedio global del área de estudio por factor en cada celda de 1
min de latitud X 1 min de longitud, μSa es el promedio específico para la distribución
de los grupos de clases de S. attenuata en relación a cada factor en cada celda, θDG
es la dispersión global de los datos, expresada por la desviación estándar, y
finalmente θSa es la dispersión específica de la distribución de los grupos en relación
a cada factor evaluado. Por lo tanto, un valor de marginalidad o tolerancia mayor o
igual a uno es considerado alto, el signo negativo refleja una marginalidad o
tolerancia con tendencia opuesta al parámetro evaluado, mientras que el signo
positivo refleja una tendencia acorde al del parámetro evaluado.
Con el fin de corroborar las tendencias observadas, se estableció un modelo
de regresión no lineal (Modelo Aditivo Generalizado) utilizando una matriz con las
100×=PMa
Pm - PMIP

44
variables eco-geográficas mencionadas anteriormente. El modelo de regresión
comprendió al cociente de observación (Co) asociado a la alimentación, como
variable respuesta, y a las variables eco-geográficas como regresores (Redfern et al.
2006). Sin embargo, este modelo solo se estimó de manera exploratoria y
confirmativa, ya que los modelos de regresión de uso de hábitat; específicamente los
modelos aditivos generalizados, son difíciles de interpretar ecológicamente por su
complejidad estructural, a pesar de ser comunes en la evaluación de relaciones entre
presencia y distribución de cetáceos en sus hábitats. El modelo se calculó mediante
el paquete estadístico “R, 2.5.1”

45
RESULTADOS
4 a Abundancia relativa del delfín manchado en Golfo Dulce
La presencia importante de S. attenuata en el área de estudio se refleja con un
porcentaje de encuentro > 80% (Cuadro 1), para la cuenca interna del Golfo Dulce:
durante 157 horas invertidas en la búsqueda, seguimiento y registro de conducta de
grupos de delfines manchados, se recorrieron 649,84 Km de áreas marinas, y se
registraron 218 estaciones ambientales. Para todo este esfuerzo de campo se logró
colectar datos de 26 encuentros, de los cuales en 23 de ellos se realizó un
seguimiento grupal para muestreos etológicos. El tamaño promedio de grupo no
superó los 70 individuos, y aproximadamente el 9% eran crías (incluyendo neonatos).
Los índices de frecuencia de avistamiento (FA) y abundancia relativa (IA)
estimados a partir del esfuerzo mencionado anteriormente (Cuadro 2), reflejan como
tendencia general una presencia mayor de delfines manchados en el área de estudio
para la época de lluvias en contraposición a la época seca, no obstante estas
diferencias no son estadísticamente significativas, al menos en los tamaños de
grupos observados (Prueba Kruskal Wallis, X2:0,06, GL: 1, p >0,05). Sin embargo,
factores relevantes al estimado de los índices de abundancia, como la distancia de
detección (para el caso de esta investigación en particular, la distancia de detección
y el área de detección son iguales numéricamente), si reflejan diferencias
estadísticamente significativas entre temporadas (Prueba Kruskal Wallis, X2:4,02, GL:
1, p <0,05).
Los estimados de abundancia reflejados en el cuadro 2, derivaron de la
extrapolación a los valores máximos de área de detección y de estaciones
ambientales para cada temporada; a partir de los índices calculados con los valores
promedio de ambas variables. Es decir, en la cuenca interna de Golfo Dulce hay 319
delfines manchados en 115 Km2 durante la temporada de lluvia, y 110 individuos
para 85,22 Km2 durante la temporada seca. La misma tendencia anterior es valida
para los estimados con estaciones ambientales (1044 delfines - 147 estaciones, y
497 delfines - 71 estaciones respectivamente).

46
Cuadro 1. Esfuerzo de investigación en campo en muestreos con S. attenuata en Golfo Dulce, Costa Rica: Temporada de lluvia 2007 – Temporada seca 2008.
Esfuerzo de Investigación en Campo 2007 - 2008 Muestreos en Bote Numero total de campañas de observación 32 Campañas con avistamientos de delfines 26 Campañas con seguimientos grupales de delfines Porcentaje de encuentro de delfines manchados
23 81%
Porcentaje de encuentros con seguimiento grupal
72%
Esfuerzo de Investigación (en horas) Horas totales de muestreo en campo 157,3 Horas con observaciones y seguimientos de delfines
63,62
Tiempo de observación (seguimiento) promedio (+/- DE) temporada de lluvia 2007
2,94 (+/- 1,50)
Tiempo de observación (seguimiento) promedio (+/- DE) temporada seca 2008
2,36 (+/- 1,29)
Esfuerzo de Investigación en No de Estaciones Ambientales (30 min)
No total de estaciones 218 No de estaciones promedio (+/- DE) temporada de lluvia 2007
9,19 (+/- 2,34)
No de estaciones promedio (+/- DE) temporada seca 2008
10,14 (+/- 1,77)
Esfuerzo de Investigación en distancia recorrida (Km)
No total de Km 649,84 Distancia recorrida promedio (+/- DE) temporada de lluvia 2007
24,24 (+/- 12,96)
Distancia recorrida promedio (+/- DE) temporada seca 2008
37,52 (+/- 14,97)
Esfuerzo de Investigación en periodos de barridos (2 min)
No total de periodos de barridos 367 Temporada de lluvia 2007 261 Temporada seca 2008 106

47
Cuadro 2. Índices y estimados de abundancia relativa de S. attenuata en Golfo Dulce, Costa Rica: Temporada de lluvia 2007 – Temporada seca 2008.
Abundancia relativa Stenella attenuata Area de detección (Km2)
Promedio (+/- DE) temporada de lluvia 2007 7,24 (+/- 5,14)Promedio (+/- DE) temporada seca 2008 11,89 (+/- 3,75) Temporada de lluvia (n=16)
Promedio (+/- DE) tamaño de grupos 59,06 (+/- 18,68)Total de individuos observados 945 Promedio de crías (incluyendo neonatos) 5,31 (+/- 2,12)Porcentaje promedio de crías por grupo 9% (+/- 3) Temporada Seca (n=7)
Promedio (+/- DE) tamaño de grupos 65 ( +/- 12,58) Total de individuos observados 455 Promedio de crías (incluyendo neonatos) 5,29 ( +/- 2,06)Porcentaje promedio de crías por grupo 9% (+/- 4) Índice de Abundancia (Individuos Totales/Área de detección promedio)
Promedio temporada de lluvia (+/- DE) 19,98 ( +/- 14,19)Intervalo de confianza 95% 13,59 – 27,02Promedio temporada seca (+/- DE) 15,77 ( +/- 4,97) Intervalo de confianza 95% 11,98 – 19,22 Abundancia relativa (No de individuos)
Temporada de lluvia para un área total de detección de 115 Km2
319,5
Intervalo de confianza 95% 217,38 –432,38Temporada seca para un área total de detección de 85,22 Km2
110,39
Intervalo de confianza 95% 83,89 – 134,52 Índice de Abundancia (Individuos Totales/Estaciones ambientales promedio)
Promedio temporada de lluvia (+/- DE) 7,10 ( +/- 4,07)Intervalo de confianza 95% 5,43 – 9,28 Promedio temporada seca (+/- DE) 6,56 ( +/- 1,72)Intervalo de confianza 95% 5,29 – 7,83 Abundancia relativa (No de individuos)
Temporada de lluvia para un total de 147 estaciones
1043,62
Intervalo de confianza 95% 798,06 – 1363,53Temporada seca para un total de 71 estaciones 497 Intervalo de confianza 95% 375,29 – 555,60

48
El cuadro 4, ilustra el valor estimado de densidad a partir de la abundancia relativa
de delfines manchados reportadas anteriormente y el consumo de biomasa asociado.
Cuadro 3. Frecuencias de avistamiento de delfines manchados, S. attenuata en Golfo Dulce, Costa Rica: Temporada de lluvia 2007 – Temporada seca 2008.
Cuadro 4. Comparación del consumo de biomasa de S. attenuata en Golfo Dulce y Golfo de Papagayo, Costa Rica.
* Se considera para este estimado un estándar de 180 días de duración de la temporada
Localidad Golfo Dulce-Lluvia*
(esta contribución)
Golfo de Papagayo-Seca*
(May – Collado 2001)
Densidad:
No individuos/Km2
2,16 1,24
Consumo de Biomasa
(Ton/Año/Km2)
260,20 187,8
Frecuencia de avistamiento (FA) Lluvia 2007 No total Avistamientos/Tiempo Observación Promedio
6,46 (Avistamientos/Horas)
No total Avistamientos/Estaciones Ambientales Promedio
2,07 (Avistamientos/Estaciones)
No total Avistamientos/Distancia Recorrida Promedio
0,78 (Avistamientos/Km)
Seca 2008 No total Avistamientos/Tiempo Observación Promedio
2,96 (Avistamientos/Horas) No total Avistamientos/Estaciones Ambientales Promedio
0,70 (Avistamientos/Estaciones)
No total Avistamientos/Distancia Recorrida Promedio
0,19 (Avistamientos/Km)

49
4. b Conducta del delfín manchado en Golfo Dulce
De 26 encuentros con S. attenuata, en 23 de ellos se realizó un seguimiento
grupal para muestreos etológicos, equivalente a 367 períodos de barrido con
registros de estados conductuales (Cuadro1). Dado a la distribución del esfuerzo de
campo en dos temporadas (lluvia y seca) los resultados se presentan en
presupuestos conductuales por temporadas (Figuras 2 y 3 respectivamente).
Para ambas temporadas los estados conductuales dominantes fueron la
alimentación y movilización, totalizando en conjunto el 91% del tiempo de
observación de la temporada de lluvia y el 89% de la temporada seca. La
socialización, el descanso y el merodeo fueron observados en menor ocurrencia.
Para la temporada de lluvia, el 53% de los períodos de barrido colectados durante el
seguimiento grupal, se invirtieron en actividades de persecución, captura y consumo
de presas. La movilización, y en particular la movilización lenta, ocurrieron en 38 %
del tiempo invertido en los seguimientos. Las diferencias entre las conductas
dominantes referidas anteriormente fueron sustentadas estadísticamente (Prueba
Kruskal Wallis, X2:5,50, GL: 1, p <0,05).
Figura 2. Presupuesto conductual de S. attenuata en Golfo Dulce Costa Rica temporada de lluvia 2007: Alimentación incluyendo persecución y captura de presa (Rojo), Movilización (Verde), Descanso (Gris), Merodeo (Azul) y Socialización (Violeta).
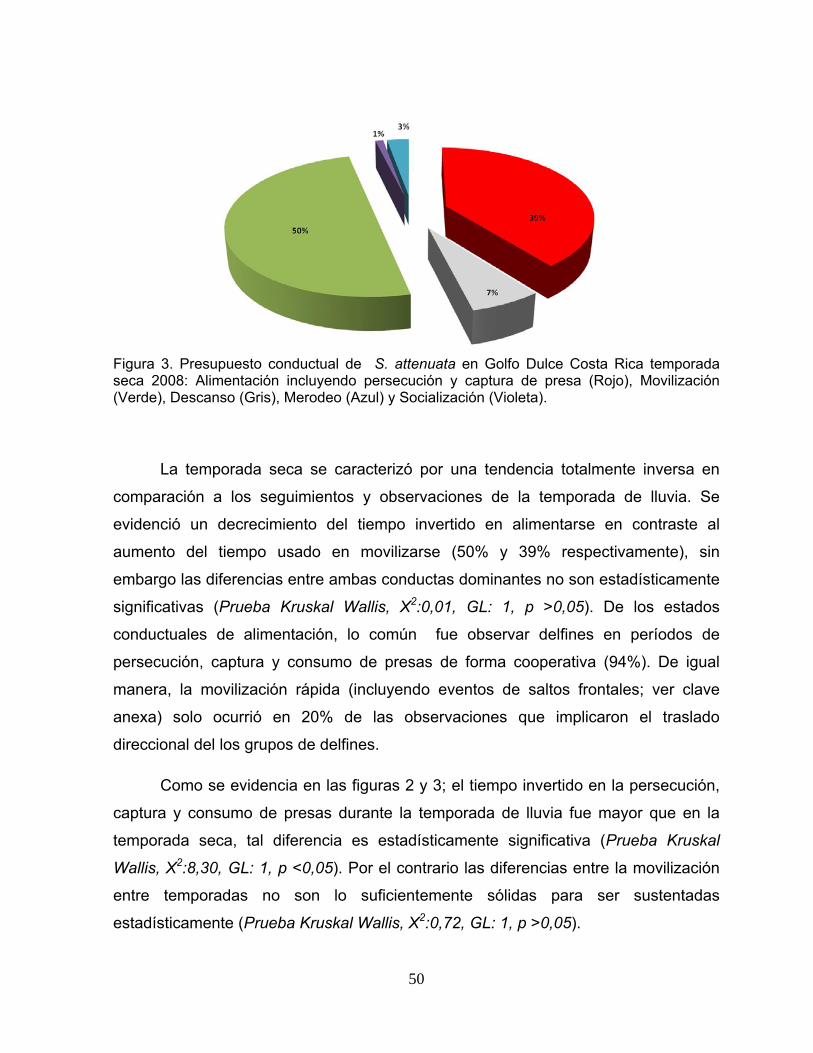
50
Figura 3. Presupuesto conductual de S. attenuata en Golfo Dulce Costa Rica temporada seca 2008: Alimentación incluyendo persecución y captura de presa (Rojo), Movilización (Verde), Descanso (Gris), Merodeo (Azul) y Socialización (Violeta).
La temporada seca se caracterizó por una tendencia totalmente inversa en
comparación a los seguimientos y observaciones de la temporada de lluvia. Se
evidenció un decrecimiento del tiempo invertido en alimentarse en contraste al
aumento del tiempo usado en movilizarse (50% y 39% respectivamente), sin
embargo las diferencias entre ambas conductas dominantes no son estadísticamente
significativas (Prueba Kruskal Wallis, X2:0,01, GL: 1, p >0,05). De los estados
conductuales de alimentación, lo común fue observar delfines en períodos de
persecución, captura y consumo de presas de forma cooperativa (94%). De igual
manera, la movilización rápida (incluyendo eventos de saltos frontales; ver clave
anexa) solo ocurrió en 20% de las observaciones que implicaron el traslado
direccional del los grupos de delfines.
Como se evidencia en las figuras 2 y 3; el tiempo invertido en la persecución,
captura y consumo de presas durante la temporada de lluvia fue mayor que en la
temporada seca, tal diferencia es estadísticamente significativa (Prueba Kruskal
Wallis, X2:8,30, GL: 1, p <0,05). Por el contrario las diferencias entre la movilización
entre temporadas no son lo suficientemente sólidas para ser sustentadas
estadísticamente (Prueba Kruskal Wallis, X2:0,72, GL: 1, p >0,05).

51
4. c Uso de hábitat del delfín manchado en Golfo Dulce
La distribución espacial de la totalidad de las actividades conductuales
registradas durante los seguimientos grupales de ambas temporadas, se ilustra en la
figura 4. De acuerdo a las tendencias mencionadas anteriormente en el presupuesto
conductual, la alimentación es la actividad clave que define el uso de hábitat en la
cuenca interna de Golfo Dulce. Las figuras 5 y 6 corresponden a la distribución
espacial de los episodios de alimentación durante los seguimientos grupales en la
temporada de lluvia y la temporada seca respectivamente.
Figura 4. Localización espacial de las conductas registradas para S. attenuata en Golfo Dulce Costa Rica: Alimentación incluyendo persecución y captura de presa (Rojo), Movilización (Verde), Descanso (Gris), Merodeo (Azul) y Socialización (Violeta).
Punta Adela
Río Rincón
Río Tigre Río Agujas
Río Esquinas Esquinas
Punta Gallardo

52
De la distribución espacial anterior se puede identificar áreas con patrones de
agregación significativas, en las actividades de alimentación. Las figuras 7 y 8
representan el análisis de agrupamiento de las clases de uso de área en cuatro
categorías (bajo, medio, alto y muy alto) para la temporada de lluvia y seca
respectivamente. De acuerdo al cuadro 5, aproximadamente 13 Km2 (4 celdas, con
un índice de área > 0,05) concentran el área núcleo donde ocurren la mayoría de las
actividades de alimentación durante la temporada de lluvia. Durante la temporada
seca están se reducen a 9,72 Km2 (3 celdas, UA > 0,05) pero agregadas
espacialmente de manera mas contigua (Cuadro 6). Las figuras 9 y 10, muestran la
distribución espacial de la probabilidad de ocurrencia de la actividad de alimentación
para ambas temporadas de muestreo en la cuenca interna del Golfo Dulce.
Figura 5. Localización espacial de las actividades de alimentación registradas para S. attenuata en Golfo Dulce, Costa Rica en la temporada de lluvia.
Punta Adela
Río Rincón
Río Tigre Río Agujas
Río Esquinas
Punta Gallardo
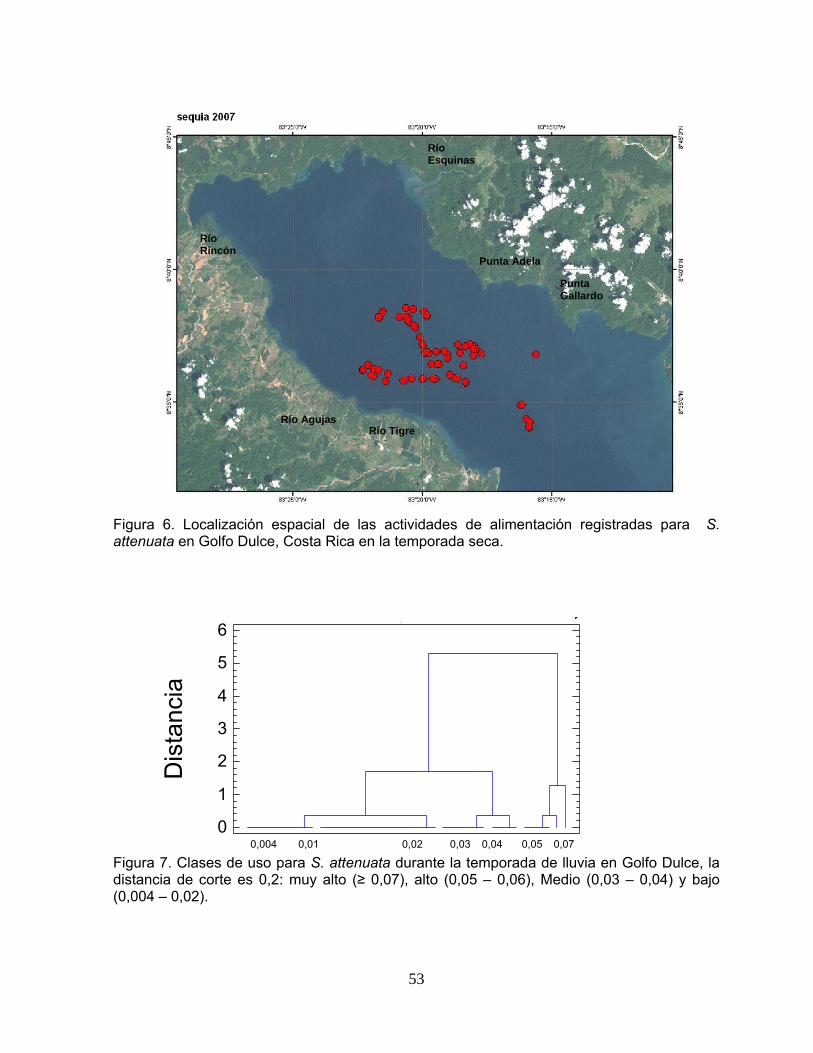
53
Figura 6. Localización espacial de las actividades de alimentación registradas para S. attenuata en Golfo Dulce, Costa Rica en la temporada seca.
Figura 7. Clases de uso para S. attenuata durante la temporada de lluvia en Golfo Dulce, la distancia de corte es 0,2: muy alto (≥ 0,07), alto (0,05 – 0,06), Medio (0,03 – 0,04) y bajo (0,004 – 0,02).
Método Media de Grupo,Euclidean Cuadrado
Dis
tanc
ia
0
1
2
3
4
5
6
0,07 0,05 0,04 0,03 0,020,004 0,01
Punta Adela
Río Rincón
Río Tigre Río Agujas
Río Esquinas
Punta Gallardo
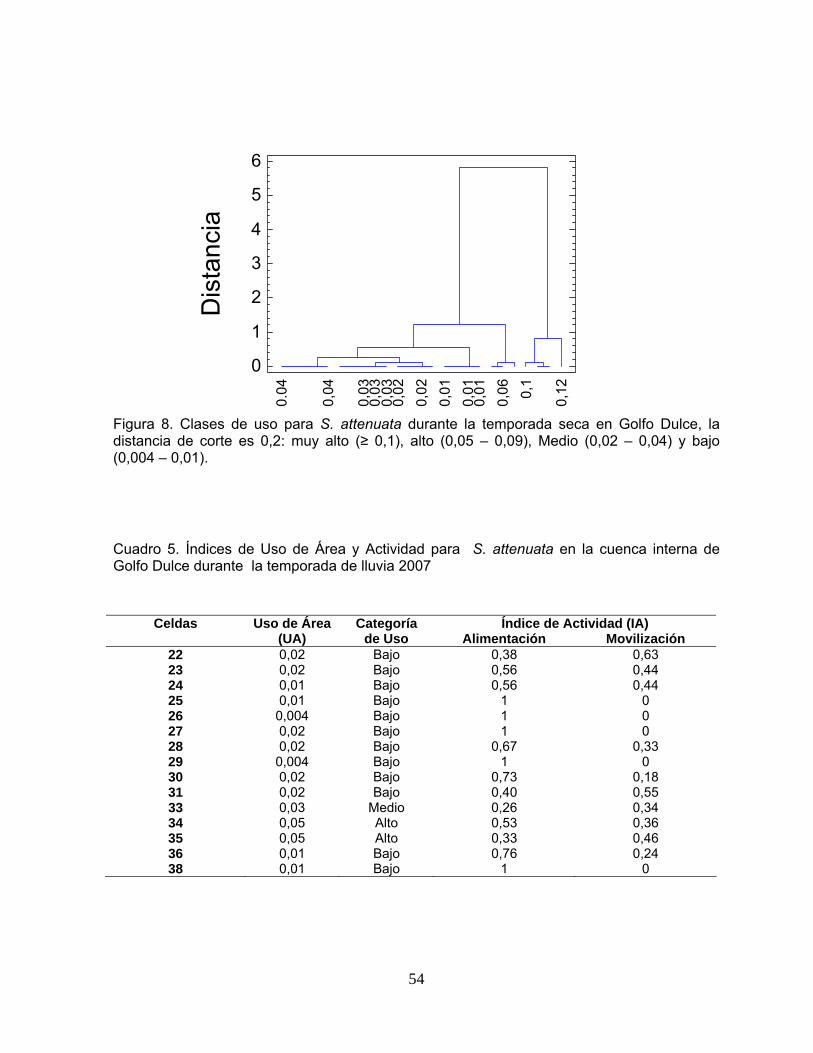
54
Dis
tanc
ia
0
1
2
3
4
5
6
0.04
0,02
0,03
0,03
0,02 0,
1
0,12
0,01
0,01
0,04
0,01
0,03
0,06
Figura 8. Clases de uso para S. attenuata durante la temporada seca en Golfo Dulce, la distancia de corte es 0,2: muy alto (≥ 0,1), alto (0,05 – 0,09), Medio (0,02 – 0,04) y bajo (0,004 – 0,01).
Cuadro 5. Índices de Uso de Área y Actividad para S. attenuata en la cuenca interna de Golfo Dulce durante la temporada de lluvia 2007
Celdas Uso de Área Categoría Índice de Actividad (IA) (UA) de Uso Alimentación Movilización
22 0,02 Bajo 0,38 0,63 23 0,02 Bajo 0,56 0,44 24 0,01 Bajo 0,56 0,44 25 0,01 Bajo 1 0 26 0,004 Bajo 1 0 27 0,02 Bajo 1 0 28 0,02 Bajo 0,67 0,33 29 0,004 Bajo 1 0 30 0,02 Bajo 0,73 0,18 31 0,02 Bajo 0,40 0,55 33 0,03 Medio 0,26 0,34 34 0,05 Alto 0,53 0,36 35 0,05 Alto 0,33 0,46 36 0,01 Bajo 0,76 0,24 38 0,01 Bajo 1 0
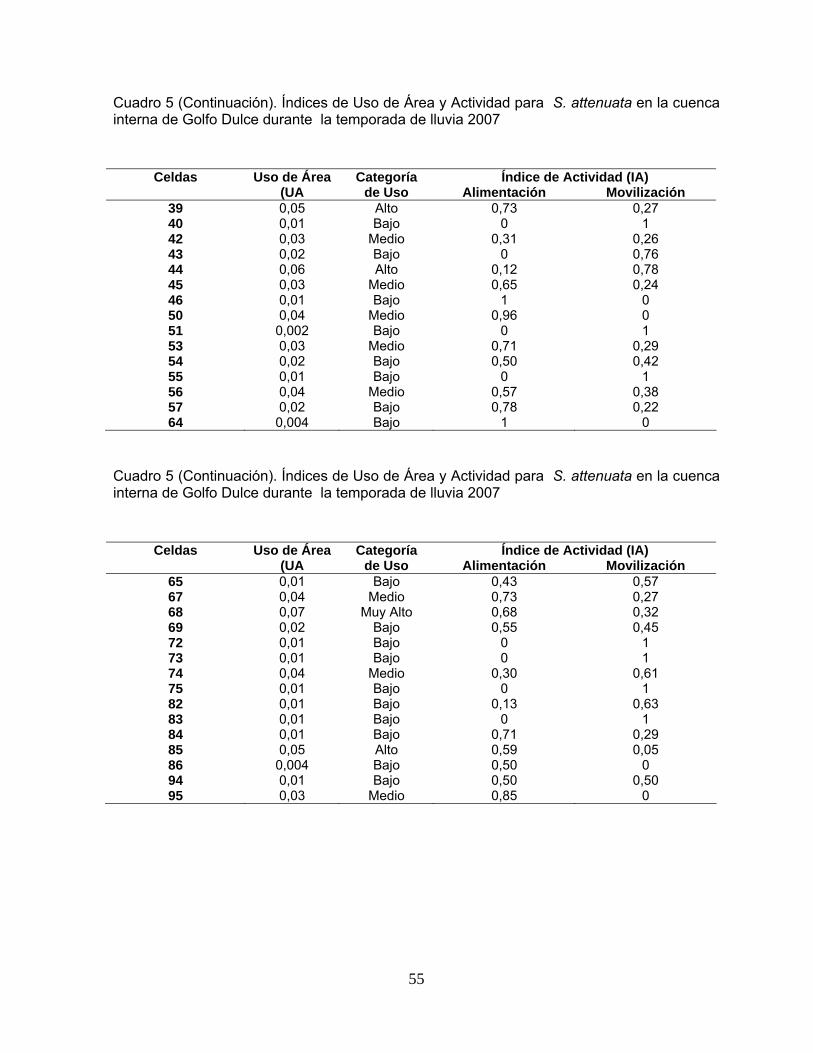
55
Cuadro 5 (Continuación). Índices de Uso de Área y Actividad para S. attenuata en la cuenca interna de Golfo Dulce durante la temporada de lluvia 2007
Cuadro 5 (Continuación). Índices de Uso de Área y Actividad para S. attenuata en la cuenca interna de Golfo Dulce durante la temporada de lluvia 2007
Celdas Uso de Área Categoría Índice de Actividad (IA) (UA de Uso Alimentación Movilización
39 0,05 Alto 0,73 0,27 40 0,01 Bajo 0 1 42 0,03 Medio 0,31 0,26 43 0,02 Bajo 0 0,76 44 0,06 Alto 0,12 0,78 45 0,03 Medio 0,65 0,24 46 0,01 Bajo 1 0 50 0,04 Medio 0,96 0 51 0,002 Bajo 0 1 53 0,03 Medio 0,71 0,29 54 0,02 Bajo 0,50 0,42 55 0,01 Bajo 0 1 56 0,04 Medio 0,57 0,38 57 0,02 Bajo 0,78 0,22 64 0,004 Bajo 1 0
Celdas Uso de Área Categoría Índice de Actividad (IA) (UA de Uso Alimentación Movilización
65 0,01 Bajo 0,43 0,57 67 0,04 Medio 0,73 0,27 68 0,07 Muy Alto 0,68 0,32 69 0,02 Bajo 0,55 0,45 72 0,01 Bajo 0 1 73 0,01 Bajo 0 1 74 0,04 Medio 0,30 0,61 75 0,01 Bajo 0 1 82 0,01 Bajo 0,13 0,63 83 0,01 Bajo 0 1 84 0,01 Bajo 0,71 0,29 85 0,05 Alto 0,59 0,05 86 0,004 Bajo 0,50 0 94 0,01 Bajo 0,50 0,50 95 0,03 Medio 0,85 0
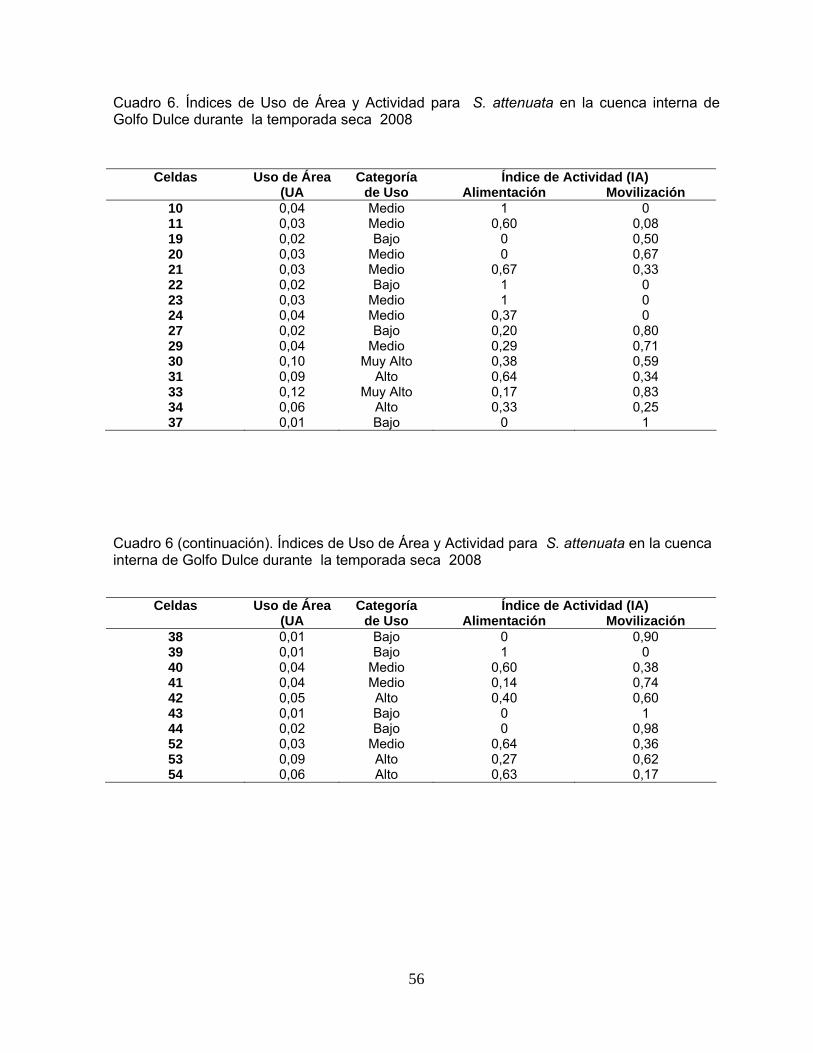
56
Cuadro 6. Índices de Uso de Área y Actividad para S. attenuata en la cuenca interna de Golfo Dulce durante la temporada seca 2008
Cuadro 6 (continuación). Índices de Uso de Área y Actividad para S. attenuata en la cuenca interna de Golfo Dulce durante la temporada seca 2008
Celdas Uso de Área Categoría Índice de Actividad (IA) (UA de Uso Alimentación Movilización
10 0,04 Medio 1 0 11 0,03 Medio 0,60 0,08 19 0,02 Bajo 0 0,50 20 0,03 Medio 0 0,67 21 0,03 Medio 0,67 0,33 22 0,02 Bajo 1 0 23 0,03 Medio 1 0 24 0,04 Medio 0,37 0 27 0,02 Bajo 0,20 0,80 29 0,04 Medio 0,29 0,71 30 0,10 Muy Alto 0,38 0,59 31 0,09 Alto 0,64 0,34 33 0,12 Muy Alto 0,17 0,83 34 0,06 Alto 0,33 0,25 37 0,01 Bajo 0 1
Celdas Uso de Área Categoría Índice de Actividad (IA) (UA de Uso Alimentación Movilización
38 0,01 Bajo 0 0,90 39 0,01 Bajo 1 0 40 0,04 Medio 0,60 0,38 41 0,04 Medio 0,14 0,74 42 0,05 Alto 0,40 0,60 43 0,01 Bajo 0 1 44 0,02 Bajo 0 0,98 52 0,03 Medio 0,64 0,36 53 0,09 Alto 0,27 0,62 54 0,06 Alto 0,63 0,17
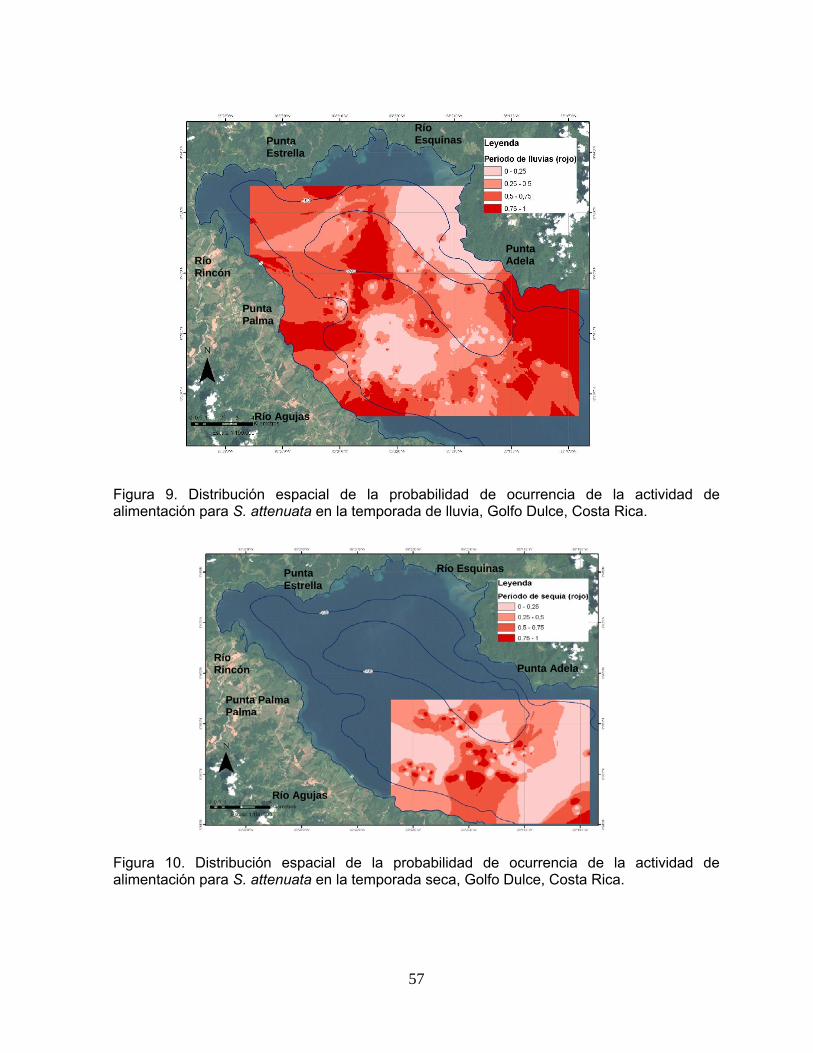
57
Figura 9. Distribución espacial de la probabilidad de ocurrencia de la actividad de alimentación para S. attenuata en la temporada de lluvia, Golfo Dulce, Costa Rica.
Figura 10. Distribución espacial de la probabilidad de ocurrencia de la actividad de alimentación para S. attenuata en la temporada seca, Golfo Dulce, Costa Rica.
Punta Adela Río
Rincón
Punta Estrella
Río Agujas
Río Esquinas
Punta Palma
Punta Adela Río Rincón
Punta Estrella
Río Agujas
Río Esquinas
Punta Palma Palma

58
4.c.1 Variables eco-geográficas que definen el hábitat de alimentación
El cuadro 7 y 8 muestra los valores promedios de las variables eco-
geográficas asociadas con los episodios de alimentación en la cuenca interna de
Golfo Dulce:
Temperatura: Los episodios de captura y consumo de presas no parecen estar
asociados a una temperatura superficial del mar en particular, durante la temporada
de lluvia (Prueba Kruskal Wallis, X2:0,35, GL: 2, p >0,05). Sin embargo, durante la
temporada seca, los eventos parecen relacionarse a temperaturas igual o mayores a
los 29 grados (Prueba Kruskal Wallis, X2:12,46, GL: 2, p < 0,05).
Distancia a río más cercano: Durante la temporada de lluvia las distancias máximas
(2,16 Km, 5,04 Km, 7,20 Km, y 8, 64 Km) en relación a la cercanía a los ríos Tigre,
Agujas, Barrigones, Caballero, Rincón y Esquinas, pareciesen ser relevantes a los
episodios de alimentación de S. attenuata (Prueba Kruskal Wallis, X2:9,20, GL: 3, p <
0,05). En la temporada seca la captura y consumo de presas pareciese ocurrir
mayormente a distancias de 5,04 y 7,20 Km de los ríos que desembocan en la
cuenca interna del Golfo Dulce (Prueba Kruskal Wallis, X2:9,39, GL: 3, p < 0,05).
Cuadro 7. Variables eco-geográficas asociadas al nicho ecológico de alimentación de S. attenuata en la cuenca interna de Golfo Dulce: temperatura superficial y distancia de ríos
Variable eco-geográfica Alimentación Lluvia
Alimentación Seca
Clases de Temperatura A (29,5 – 30,5 °C) 0,88 (+/-0,24) 0,85 (+/-0,21) B (29 - 29,4 °C) 0,86 (+/-0,27) 0,84 (+/-0,22) C (28 – 28,9 °C) 0,85 (+/-0,27) 0,56 (+/-0,16) Diferencias entre clases (Kruskal Wallis)
P > 0,05 (X2 = 0,35) p < 0,002 (X2 = 12,46)
Clases de Distancia de Ríos A (0,90 – 2,16 Km) 0,95 (+/-0,14) 0,94 (+/-0,09) B (2,17 - 5,04 Km) 0,80 (+/-0,29) 0,78 (+/-0,19) C (5,05 - 7,20 Km.) 0,89 (+/-0,25) 0,99 (+/-0,001) A (7,21 - 8,64 Km) 0,98 (+/-0,07) 0,83 (+/-0,29) Diferencias entre clases (Kruskal Wallis)
p< 0,001 (X2 = 139,47) p < 0,001 (X2 = 35,58)
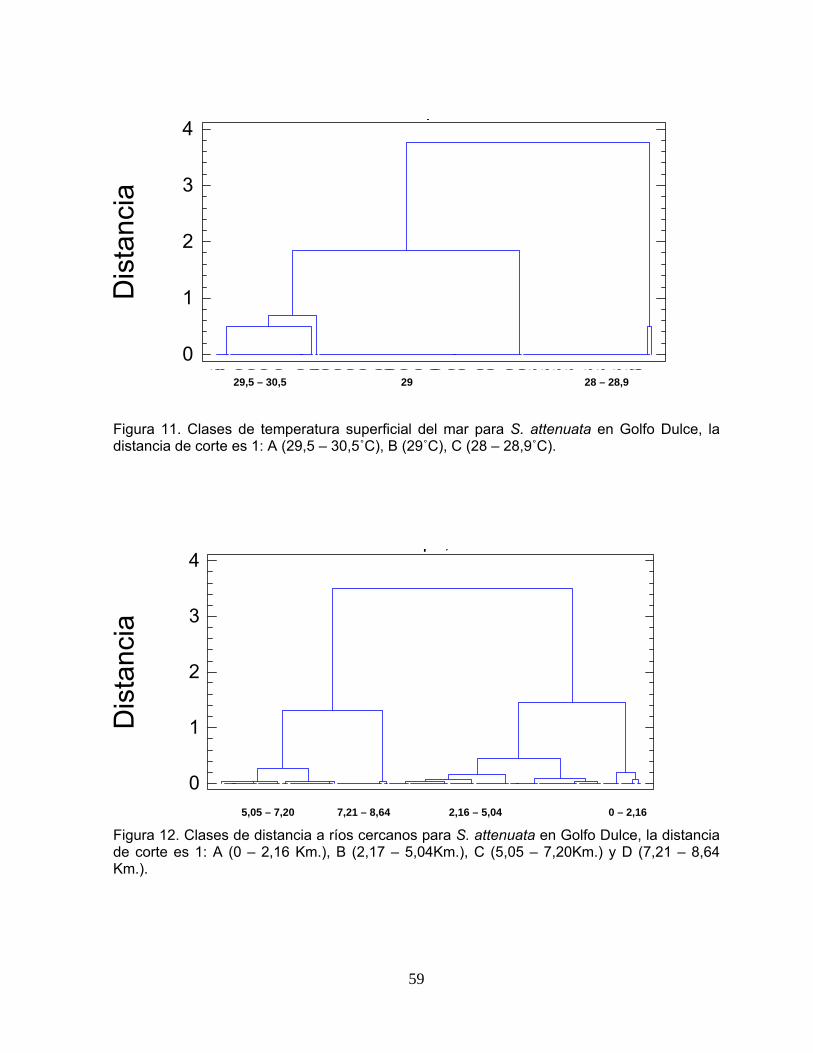
59
Figura 11. Clases de temperatura superficial del mar para S. attenuata en Golfo Dulce, la distancia de corte es 1: A (29,5 – 30,5˚C), B (29˚C), C (28 – 28,9˚C).
Figura 12. Clases de distancia a ríos cercanos para S. attenuata en Golfo Dulce, la distancia de corte es 1: A (0 – 2,16 Km.), B (2,17 – 5,04Km.), C (5,05 – 7,20Km.) y D (7,21 – 8,64 Km.).
pD
ista
ncia
0
1
2
3
4
29.5 29 29 29 29
29,5 28 28 28 2829 29 29 29 282930 30 30 29 2829 28 2829 2930 28 2829 29
30,530 30 29 2929 28 – 28,9 29,5 – 30,5
p ,
Dis
tanc
ia
0
1
2
3
4
0 – 2,16 2,16 – 5,04 5,05 – 7,20 7,21 – 8,64
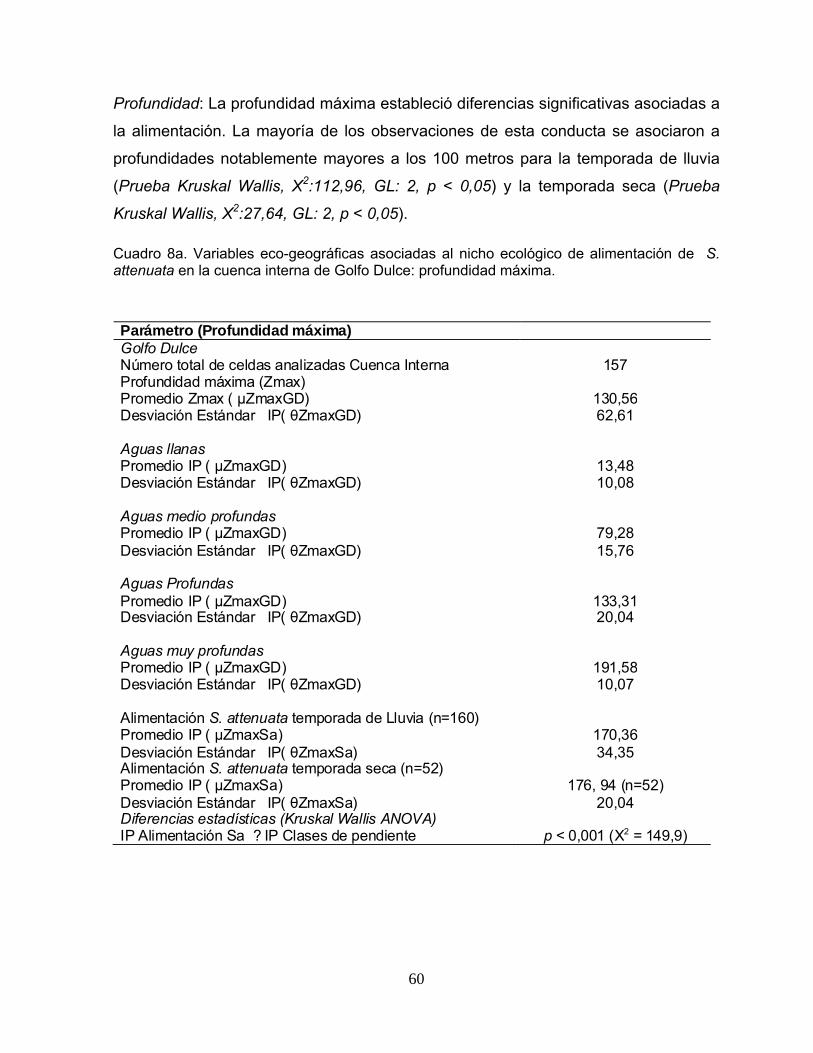
60
Profundidad: La profundidad máxima estableció diferencias significativas asociadas a
la alimentación. La mayoría de los observaciones de esta conducta se asociaron a
profundidades notablemente mayores a los 100 metros para la temporada de lluvia
(Prueba Kruskal Wallis, X2:112,96, GL: 2, p < 0,05) y la temporada seca (Prueba
Kruskal Wallis, X2:27,64, GL: 2, p < 0,05).
Cuadro 8a. Variables eco-geográficas asociadas al nicho ecológico de alimentación de S. attenuata en la cuenca interna de Golfo Dulce: profundidad máxima.
Parámetro (Profundidad máxima) Golfo Dulce Número total de celdas analizadas Cuenca Interna 157 Profundidad máxima (Zmax) Promedio Zmax ( μZmaxGD) 130,56 Desviación Estándar IP( θZmaxGD) 62,61 Aguas llanas Promedio IP ( μZmaxGD) 13,48 Desviación Estándar IP( θZmaxGD)
10,08
Aguas medio profundas Promedio IP ( μZmaxGD) 79,28 Desviación Estándar IP( θZmaxGD)
15,76
Aguas Profundas Promedio IP ( μZmaxGD) 133,31 Desviación Estándar IP( θZmaxGD)
20,04
Aguas muy profundas Promedio IP ( μZmaxGD) 191,58 Desviación Estándar IP( θZmaxGD)
10,07
Alimentación S. attenuata temporada de Lluvia (n=160) Promedio IP ( μZmaxSa) 170,36 Desviación Estándar IP( θZmaxSa) 34,35 Alimentación S. attenuata temporada seca (n=52) Promedio IP ( μZmaxSa) 176, 94 (n=52)Desviación Estándar IP( θZmaxSa) 20,04 Diferencias estadísticas (Kruskal Wallis ANOVA) IP Alimentación Sa ? IP Clases de pendiente p < 0,001 (X2 = 149,9)

61
Cuadro 8b. Variables eco-geográficas asociadas al nicho ecológico de alimentación de S. attenuata en la cuenca interna de Golfo Dulce: pendiente del fondo.
Pendiente: La pendiente representó un descriptor importante en la ocurrencia de
episodios de alimentación, la captura y consumo de presas se asociaron a fondos de
pendientes muy poco inclinadas (bajas y medias bajas) durante ambas; la temporada
de lluvia (Prueba Kruskal Wallis, X2:145,3, GL: 5, p < 0,05), y la temporada seca
(Prueba Kruskal Wallis, X2:37,15, GL: 1, p < 0,05).
Parámetro (Pendiente fondo oceánico) Golfo Dulce Número total de celdas analizadas Cuenca Interna 157 Índice de Pendiente (IP) Promedio IP ( μIPGD) 23,17 Desviación Estándar IP( θIPGD) 24,95 Pendiente muy baja Promedio IP ( μIPGD) 1,92 Desviación Estándar IP( θIPGD) 1,34 Pendiente baja Promedio IP ( μIPGD) 10,42 Desviación Estándar IP( θIPGD) 5,40 Pendiente medio baja Promedio IP ( μIPGD) 24,53 Desviación Estándar IP( θIPGD) 3,85 Pendiente media Promedio IP ( μIPGD) 34,33 Desviación Estándar IP( θIPGD) 2,02 Pendiente medio alta Promedio IP ( μIPGD) 43,02 Desviación Estándar IP( θIPGD) 3,27 Pendiente alta Promedio IP ( μIPGD) 58,61 Desviación Estándar IP( θIPGD) 8,31 Pendiente empinada Promedio IP ( μIPGD) 86,32 Desviación Estándar IP( θIPGD) 6,24 Alimentación S. attenuata Temporada de Lluvia (n=160) Promedio IP ( μIPSa) 18,96 Desviación Estándar IP( θIPSa) 22,08 Alimentación S. attenuata Temporada Seca (n=52) Promedio IP ( μIPSa) 5,03 Desviación Estándar IP( θIPSa) 4,11 Diferencias estadísticas (Kruskal Wallis ANOVA) IP Alimentación Sa Lluvia ? IP Clases de pendiente p < 0,001 (X2 = 149,90)IP Alimentación Sa Seca ? IP Clases de pendiente p < 0,001 (X2 = 174,76)

62
Cuadro 9a. Análisis de Factores de Nicho Ecológico de la variable eco-geográficas profundidad máxima (Zmax), para S. attenuata en Golfo Dulce, durante la temporada de lluvia 2007.
Los índices de marginalidad y de tolerancia para la profundidad máxima
(cuadros 9a y 10a) y la pendiente (Cuadros 9b y 10b) en las observaciones de
episodios de alimentación para ambas temporadas reflejan como tendencia principal,
índices muy alto (> 1) que favorecen la diferencia con tendencia opuesta en valores
de profundidad máxima donde ocurren episodios de alimentación, con profundidades
llanas, medias y aguas profundas. Por el contrario la marginalidad en aguas muy
profundas es alta (1 para la temporada de lluvia, 0,74 para la temporada Seca).
Adicionalmente la tolerancia a alimentarse en aguas de poca profundidad (o de
batimetría mayor a la máxima profundidad del área de estudio: 215 m) es
relativamente baja en comparación a la tolerancia a profundidades medias y altas.
Análisis de Factores de Nicho Ecológico Área de Estudio(GD)
Marginalidad -0,33 Tolerancia >1 Profundidad Máxima Llana
Marginalidad > -1 Tolerancia 0,29 Profundidad Máxima Media
Marginalidad > -1 Tolerancia 0,48 Profundidad Máxima Aguas Profundas
Marginalidad -1 Tolerancia 0,58 Profundidad Máxima Aguas Muy Profundas
Marginalidad 1 Tolerancia 0,29

63
Cuadro 9b. Análisis de Factores de Nicho Ecológico de la variable eco-geográficas pendiente (IP), para S. attenuata en Golfo Dulce, durante la temporada de lluvia 2007.
En relación a la pendiente, la marginalidad es opuesta a fondos planos, y
diferente a fondos medianamente inclinados y muy inclinados (marginalidad > 1),
pero asociada con fondos de pendientes bajas y medio bajas. Hay un incremento de
la tolerancia a pendientes mayores de un 40% para la temporada de lluvia. En la
temporada seca la marginalidad favorece mucho los fondos de pendientes medio
baja, casi planos (< 2%), pero con alta tolerancia a fondos de inclinación diferente.
Análisis de Factores de Nicho Ecológico Área de Estudio(GD)
Marginalidad 0,09 Tolerancia >1 Pendiente Muy Baja
Marginalidad > -1 Tolerancia 0,06 Pendiente Baja
Marginalidad 0,81 Tolerancia 0,24 Pendiente Media Baja
Marginalidad 0,74 Tolerancia 0,17 Pendiente Media
Marginalidad >1 Tolerancia 0,09 Pendiente Media Alta
Marginalidad >1 Tolerancia 0,15 Pendiente Alta
Marginalidad >1 Tolerancia 0,38 Pendiente Muy Alta
Marginalidad >1 Tolerancia 0,28

64
Cuadro10a. Análisis de Factores de Nicho Ecológico de la variable eco-geográficas profundidad máxima (Zmax), para S. attenuata en Golfo Dulce, durante la temporada seca 2008.
Los resultados del modelo de regresión (anexo 4) establecen que durante la
temporada de lluvia, la profundidad y los ríos mencionados anteriormente describen
significativamente la ocurrencia de episodios de alimentación. Durante la temporada
seca los regresores más relevantes son la distancia a los ríos y la pendiente. Estas
tendencias son acordes con los resultados presentados anteriormente, y confirman la
relevancia de la profundidad, la pendiente y los ríos en la determinación del nicho
ecológico de alimentación de S. attenuata.
Análisis de Factores de Nicho Ecológico Área de Estudio(GD)
Marginalidad -0,38 Tolerancia > -1 Profundidad Máxima Llana
Marginalidad >-1 Tolerancia 0,42 Profundidad Máxima Media
Marginalidad >-1 Tolerancia 0,66 Profundidad Máxima Profunda
Marginalidad -1 Tolerancia 0,83 Profundidad Máxima Muy Profunda
Marginalidad 0,74 Tolerancia 0,42
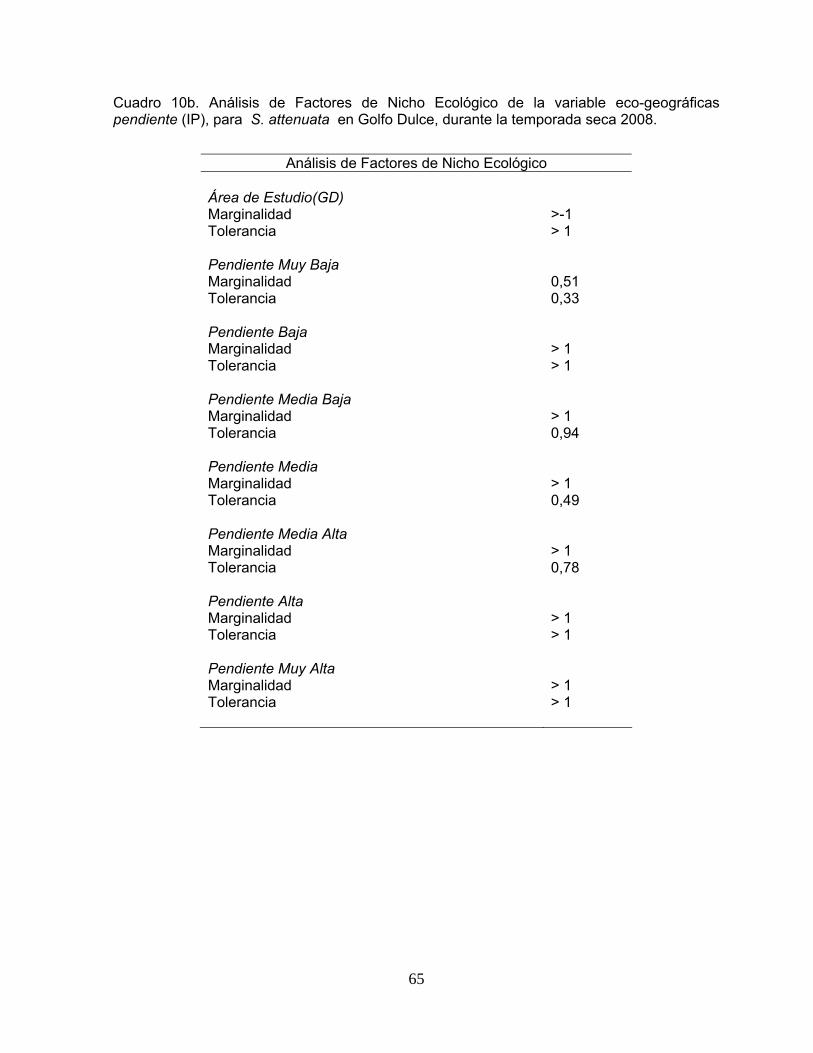
65
Cuadro 10b. Análisis de Factores de Nicho Ecológico de la variable eco-geográficas pendiente (IP), para S. attenuata en Golfo Dulce, durante la temporada seca 2008.
Análisis de Factores de Nicho Ecológico Área de Estudio(GD)
Marginalidad >-1 Tolerancia > 1 Pendiente Muy Baja
Marginalidad 0,51 Tolerancia 0,33 Pendiente Baja
Marginalidad > 1 Tolerancia > 1 Pendiente Media Baja
Marginalidad > 1 Tolerancia 0,94 Pendiente Media
Marginalidad > 1 Tolerancia 0,49 Pendiente Media Alta
Marginalidad > 1 Tolerancia 0,78 Pendiente Alta
Marginalidad > 1 Tolerancia > 1 Pendiente Muy Alta
Marginalidad > 1 Tolerancia > 1

66
DISCUSIÓN
5. a Abundancia relativa del delfín manchado en Golfo Dulce
Los resultados obtenidos en esta investigación muestran algunas tendencias
claras que caracterizan la población en estudio, Los tamaños de grupo son
relativamente grande y la presencia de la especie en la cuenca interna es importante,
así lo refleja la posibilidad de encuentros (81%), en lo que pareciese ser un rango de
distribución sumamente restringido, obviamente condicionado por el carácter semi-
cerrado de la cuenca interna del Golfo Dulce.
Igualmente alta es la frecuencia de avistamiento durante el período de
muestreo de esta investigación al compararse con trabajos anteriores (Acevedo &
Burkhart, 1998; Cubero Pardo 1998a, 1998b, 2007) en la misma localidad de
muestreo y con la misma especie. Tales diferencias, más que reflejar alguna
tendencia en el incremento poblacional, lo cual no es enteramente descartable,
ilustran ventajas metodológicas al concentrarse los muestreo a una sola especie.
Acevedo y Burkhart (1998) y Cubero Pardo (1998a, 1998b, 2007), establecieron
como objetivo estudiar a T. truncatus y S. attenuata como especies simpátricas, lo
que implicó una dispersión no equitativa de las unidades de esfuerzos entre las dos
especies, con una mayor cantidad de encuentro para delfines nariz de botella, que
repercutió en un porcentaje medio (< 50 %, Acevedo & Burkhart, 1998) a bajo (<
20%, Cubero Pardo 1998a, 1998b, 2007) para delfines manchados, a pesar de un
mayor tiempo invertido en campo.
Hay evidencias que podrían sustentar la suposición de haber estado
observando el mismo grupo durante todo el período de muestreo, como la presencia
de un individuo positivamente identificado en todos los registros de seguimiento
grupal. Esto podría ser una característica única y local de esta población en estudio,
generalmente las agrupaciones de delfines oceánicos, y particularmente las de
delfines del género Stenella, son sociedades tipo fisión- fusión, donde la estabilidad
de las unidades sociales es muy comprometida (Perrin 2001). Sin embargo, unidades
poblacionales geográficamente aisladas, tienden a desarrollar particularidades

67
locales, como el adoptar un patrón de agregación social discreto y estable
(Karczmarski et al., 2005). La tendencia anterior podría explicar el patrón de
agregación en grupos numerosos de delfines manchados, que potencialmente se
constituiría de los mismos individuos, con evidencias claras de un comportamiento
residente dentro del área de estudio.
En el caso de los estimados en base a la unidad de esfuerzo (estaciones
ambientales), las estaciones ambientales cubrieron todo el muestreo en campo,
incluyendo ocasiones en donde no hubo registros de seguimiento grupal, de manera
que al extrapolar el índice de abundancia a lo que corresponde al numero total de
estaciones, el resultado de 1044 delfines en la temporada de lluvia y 497 en la seca,
constituye probablemente una sobre-estimación.
El estimado de abundancia relativa en base a la distancia de espera (319
delfines, lluvia - 110 delfines, seca) es un valor conservador, corresponde con una
población discreta muy local, donde en promedio cada grupo avistado se constituye
en menos del 10% de crías. La diferencia en los valores de abundancia relativa en
ambos métodos, pareciese mostrar alguna tendencia de respuesta al efecto de
factores denso-independiente relacionados a la estacionalidad. Dicha tendencia
pareciese coincidir con lo documentado para la costa norte del pacífico en Costa
Rica (May-Collado 2001, Rodríguez Saenz & Rodríguez Fonseca 2004 May-Collado
& Morales 2005) pero en si, la situación de la estacionalidad en dichas localidades
(Golfo de Papagayo - Bahía Culebra) es poco conclusiva, mientras Rodríguez Saenz
& Rodríguez Fonseca (2004) afirman que el incremento en el número de animales es
en la época de lluvia, May-Collado & Morales (2005) atribuyen un numero mayor de
delfines observados a la finalización de las lluvias y el inicio del efecto de los vientos
alisios (que inciden en la surgencia costera) en la abundancia de presas, en esencia
el incremento reportado en May Collado & Morales (2005) también podría
relacionarse a factores denso-independiente (mayor abundancia de presas) y se
sustenta con una mayor incidencia de comportamientos de alimentación. Las
observaciones reportadas en este estudio, en relación a un mayor número de
delfines en la cuenca interna del Golfo Dulce, no son estadísticamente significativas

68
en relación a la cantidad de delfines observados en las temporadas de lluvia y seca,
pero son paralelas a la incidencia de niveles mayores en actividades alimentarias
durante las lluvias, que disminuyen gradualmente en la época seca.
La comparación entre el consumo de biomasa de las poblaciones de Golfo
Dulce y Golfo de Papagayo a partir de una abundancia relativa conocida, pareciesen
reflejar patrones de consumo similares. Anteriormente se mencionó que las
diferencias entre la abundancia relativa entre temporadas podrían estar
condicionadas a la abundancia de el recurso trófico, ya que coinciden con el
incremento de conductas inherentes a actividades de alimentación y captura de
presas, el nivel relativamente alto de consumo de biomasa pareciese reforzar la
tendencia, que si bien estadísticamente no es significativa (en la diferencia estacional
de los tamaños de grupos), lo es biológicamente.
La similitud en los niveles de consumo de presas de ambas poblaciones de
delfines manchados, no solo es una referencia de línea base para el manejo y
conservación de la especie, también es relevante como indicador de productividad en
Golfo Dulce (un ecosistema aparentemente oligotrófico): es poco probable sustentar
una población discreta de depredadores, con un nivel de consumo de presas similar
al de poblaciones localizadas en ecosistemas marinos caracterizados por alta
productividad (Golfo de Papagayo) y con preferencia por pequeños pelágicos como
presas (Belonidae y Exocoetidae), en un ecosistema de baja productividad biológica.
Es claro que el subsidio a la productividad primaria proveniente de los ríos, es mucho
más importante de lo que se ha documentado anteriormente en Golfo Dulce
(Quesada Alpizar & Cortés 2006), también es evidente que en estas áreas es donde
se deberían focalizar las estrategias de manejo y conservación del hábitat.
5.b Conducta del delfín manchado en Golfo Dulce
La actividad de alimentación en conjunto con la movilización, son los aspectos
etológicos más resaltante de los estados conductuales registrados para S. attenuata
en Golfo Dulce. De manera indirecta esta tendencia podría ser un reflejo del efecto
de un gradiente de recursos tróficos, que ilustra el uso de hábitat considerando la

69
dinámica ecológica, que implican las respuestas fisiológicas ante factores
ambientales igualmente dinámicos. La factibilidad de la captura, la accesibilidad de
presas potenciales y el éxito en la captura de presas, son cruciales en la formación,
por parte del depredador, de estrategias basadas en la relación costo-beneficio
(Daura Jorge, 2007).
La dominancia de ambas actividades y el hecho de que durante la época de
lluvia, aumenten significativamente los episodios de captura y consumo de presas,
pareciese indicar una subordinación de conductas en torno a una de mayor
relevancia (Garrafo et al., 2007): capturar y consumir presas es inherente al consumo
de energía, el consumo a su vez afecta la aptitud (sobrevivencia). Por otro lado la
búsqueda de presas estaría condicionada a los procesos de captura y consumo,
también a que tan lejos y que tan densos son los parches de presas. Lo anterior
deviene en secuencias de alimentación y movilización, así como en el balance de
ambas actividades en función a la disponibilidad de alimento.
Las observaciones etológicas de Cubero Pardo (1998a, 1998b) establecieron
un presupuesto conductual constituido (en orden de relevancia) por la socialización
(54%), desplazamiento (25%), Merodeo (15%) y por ultimo la alimentación (6%). Los
niveles de tiempo invertidos en las actividades mencionadas anteriormente
contrastan con las de esta investigación. En base a sus observaciones Cubero Pardo
(1998a, 1998b) concluyó que S. attenuata extendía su rango de distribución fuera de
Golfo Dulce, con el fin de evitar competencia, y por la baja productividad del sistema
(que se traduce en poca disponibilidad de presas).
Al analizar la información anterior bajo la luz de la “teoría del consumidor
óptimo”, las conclusiones de Cubero Pardo (1998a, 1998b) tendrían una gran lógica,
tal como se plantean las condiciones en base a la ecuación 1 (Pág. 28 del Marco
Teórico): la movilización por búsqueda de presas dentro del Golfo Dulce, es mayor a
la ganancia por valor nutricional (M > E), por lo que la inversión en captura de presas,
es tan poco relevante que es casi igual a cero (A ≥ 0). Esto implicaría que la
inversión energética en busca de mayor ganancia nutricional se enfocaría a otras
zonas con mayor potencialidad en parches de alimentos, ya que no tiene sentido

70
para el incremento de la aptitud, el invertir en la búsqueda, captura y consumo si la
ganancia nutricional es mínima. Adicionalmente, la seguridad relativa del golfo como
refugio a posibles depredadores, potenciaría el uso energético en las actividades de
socialización, incluyendo el apareamiento.
La relevancia de la alimentación y la movilización como actividades
conductuales en esta investigación plantea un escenario muy diferente al observado
por Cubero - Pardo (1998a, 1998b): la disponibilidad de presas en la temporada de
lluvia promueve una inversión mayor en la captura y consumo de presas que
maximiza el valor del cociente entre el valor nutritivo de la presa y el esfuerzo de
captura (A ≤ E), al mismo tiempo, el valor de la movilización entre parches es
significativamente menor al tiempo usado en la captura (0 ≤ M < A). En la temporada
seca se presenta un balance entre los niveles de captura de presas y movilización
entre parches (M = A ≤ E). En esencia, S. attenuata presentaría una conducta
alimentaria que concuerda con un consumidor especialista en ámbitos y temporadas
de abundancia de presas (tal como se predice en la situación 1a, pág.28 del Marco
Teórico), que durante la época seca oscila su conducta entre tendencias generalistas
y especialistas.
Un aspecto relevante a considerar en la explicación del modelo anterior
basado en la “teoría del consumidor óptimo”, es la influencia de la competición intra-
específica en la conducta alimentaria de S. attenuata. La competencia inter-
específica con T. truncatus como especie simpátrica es mínima (Acevedo y Burkhart
1998, Cubero Pardo 1998a, 1998b, 2007) y se debe a la repartición de hábitat como
estrategia de coexistencia (Oviedo 2007). Sin embargo en el nicho de alimentación
real del delfín manchado, el consumo de energía per cápita podría ser influenciado
por competición de individuos de la misma población (Begon et al. 2006).
Como se puntualizó en los resultados, la captura y consumo de presas en
forma cooperativa fue particularmente importante y se reflejó en la ocurrencia de
círculos de alimentación. Esta conducta se ha documentado como la estrategia
principal de captura de presas en Nueva Zelanda por delfines comunes (Delphinus
delphis); denominada carouseling (Neumann y Orams 2003), también es análoga a la

71
técnica de acorralamiento mencionada en May - Collado (2001) y May - Collado y
Morales (2005). La cooperación en la captura y consumo de presa no solo afecta la
eficacia en el consumo de energía, al incrementar la eficiencia en la captura (Gallo-
Reynoso 1991, Neumann y Orams 2003), sino que debe reducir considerablemente
la competencia entre con-específicos al balancear el valor nutricional de la presa
capturada per cápita, con un esfuerzo de captura colectivo. Las estrategias de
captura de presas que implican cooperación en el acorralamiento de presas son
comunes en ecosistemas caracterizados por alta productividad (Gallo-Reynoso
1991), tal como ocurre con S. attenuata en el Golfo de Papagayo (May - Collado
2001, May Collado y Morales 2005).
Los delfines manchados en Golfo Dulce pareciese invertir mayor tiempo y
energía en capturar presas y alimentarse de ellas mediante el comportamiento típico
de un consumidor especialista durante períodos de abundancia, y de manera mixta
(especialista - generalista) en la época seca, reduciendo notablemente el efecto
negativo de la competencia entre con-específicos, al adoptar como estrategia la
captura cooperativa.
5.c Uso de hábitat del delfín manchado en Golfo Dulce
La alimentación como actividad conductual dominante determina el uso de
hábitat de S. attenuata en el área de estudio, y define la cuenca interna de Golfo
Dulce como un hábitat crítico para la captura y consumo de presas (Hoyt 2005). En
el Golfo de Papagayo (costa del Pacífico norte en Costa Rica), el uso de hábitat
pareciese ser igualmente definido por la mayoría notable de actividades de
alimentación (May - Collado 2001, May - Collado & Morales 2005). Sin embargo, hay
un contraste notable con lo que correspondería al uso de hábitat en Golfo Dulce,
basado en las observaciones con la misma especie en 1996 - 1997 (Cubero - Pardo
1998a), básicamente el presupuesto conductual de Cubero Pardo (1998a)
establecería un área clave para la reproducción y de muy poca importancia para el
consumo de energía.

72
Adicionalmente, el mismo autor reporta la ocurrencia de la mayoría de los
episodios de captura y consumo de presas en la temporada seca. De acuerdo a lo
observado en este estudio, en el período 2007 - 2008, la alimentación se incrementa
en la temporada de lluvia, como respuesta a una mayor disponibilidad de presas, que
a su vez podría asociarse a un mayor caudal y aporte de nutrientes de los ríos del
Golfo. Estas diferencias en niveles de ocurrencia de actividad dentro de una misma
localidad, no reflejarían cambios notables en la productividad y disponibilidad de
presas, sino a diferencias metodológicas, particularmente en la definición de las
variables asociadas al muestreo etológico.
Se podría argumentar el uso de la cuenca interna de Golfo Dulce como área
de transito de delfines manchados, esto en base a la relevancia de la movilización
como estado conductual, particularmente en la temporada seca. Lo refutable de esta
apreciación es que Golfo Dulce es primeramente una cuenca semi-cerrada, una
especie de mar interno, si habría una movilización direccionada en particular, ¿por
qué habría de ocurrir por una ruta que termina en una cuenca sin salida alterna?
Adicionalmente, ¿por qué invertiría S. attenuata energía y tiempo en movilizarse en
un mar interno?, aparentemente la respuesta a esta ultima interrogante la podría
explicar la “teoría del consumidor optimo” (Begon et al., 2006); la movilización se
justifica, solo si se obtiene como beneficio la optimización del consumo de energía
para promover la aptitud, es decir solo si el esfuerzo de movilizarse (búsqueda),
garantiza que se obtendrá más energía (presas de alto valor nutritivo) de la que se
gastó.
Una explicación muy sólida a la ocurrencia importante de la movilización
dentro del presupuesto conductual, es que esta actividad es sub-ordinada a la
alimentación, tal como lo plantea Garrafo et al (2007) con el traslape de áreas
utilizadas para la alimentación y movilización en delfines oscuros (Lagenorhynchus
obscurus), en una cuenca semi-cerrada; el Golfo Nuevo (Patagonia, Provincia del
Chubut, Argentina). El aspecto determinante en este traslape es que ambas

73
actividades son secuenciales y conjugan como elementos de sustento la densidad de
los parches de presas y la distancia entre ellos.
De acuerdo a lo descrito anteriormente, se esperaría que la ocurrencia
significativa de episodios de captura y consumo de presas se asociara a la
disponibilidad de presas potenciales, ya que el uso de hábitat definido en un contexto
etológico, refleja la respuesta (conducta de alimentación) ante un gradiente de
recursos y factores ambientales influenciando dichos recursos. En esencia, la
importancia de los episodios de alimentación es indirectamente un reflejo de presas
disponibles. ¿Que tipo de presas consumiría S. attenuata en la cuenca interna de
Golfo Dulce?
Basado en los niveles tróficos y las dietas descritas por Pauly et al. (1998),
Oviedo (2007) ofrece un acercamiento teórico al consumo de presas y la divergencia
de dietas de ambos: T. truncatus y S. attenuata como especies simpátricas en Golfo
Dulce. Este autor teoriza que la dieta de los delfines manchados es básicamente
constituida por presas (particularmente peces) meso-pelágicas, y estos serian incluso
mucho más importantes (> 60%) en la dieta de S. attenuata de lo que plantean los
estimados de Pauly et al. (1998).
En los registros de episodios de captura y consumo de presas del presente
estudio, se identificaron visualmente dos presas particularmente relevantes: el pez
volador (Exocetidae) y el Ballyhoo (Hemiramphus sp), el cual es frecuentemente
utilizado como carnada en la importante operación turística de pesca deportiva en
Golfo Dulce. A estos dos elementos habría que agregarle al pez aguja (Belonidae)
como componente de la dieta, de acuerdo a lo observado por Cubero-Pardo (1998a).
La importancia de otras presas como peces meso-pelágicos linternas (Myctophidae)
es una posibilidad latente pero requeriría, debido a la migración vertical característica
de este tipo de peces, de observaciones nocturnas de episodios de alimentación. Los
aspectos discutidos anteriormente, reflejan evidencias cualitativas de la
disponibilidad de presas en la cuenca interna de Golfo Dulce, en concordancia con la

74
importancia de los episodios de alimentación que definen el uso de hábitat, más
adelante se hará énfasis en detalles relacionados con la distribución y agregación de
las presas potenciales en el Golfo Dulce.
5.c.1 Localización espacial de las áreas de alimentación
La información que proveen los índices de uso de área y actividad, permite
identificar cuatro núcleos importantes en la ocurrencia de episodios de alimentación
durante la temporada de lluvia. Para la temporada seca estos núcleos se reducen a
tres. Dichos núcleos concentran en promedio más del 60% de los episodios de
captura y consumo de presas durante la temporada de lluvia y más del 50% en la
temporada seca. Cubero-Pardo (2007) no identifica patrones espaciales discernibles
en sus observaciones con S. attenuata (1996 - 1997). Por el contrario, los registros
de T. truncatus de la misma autora, muestran un patrón espacial definido en las
actividades de alimentación entre otras.
La focalización de los episodios de alimentación que se plantea en esta
contribución, establece ventajas en la identificación de hábitats críticos promoviendo
la conceptualización y zonificación de áreas importantes y áreas críticas
Las áreas importantes para la alimentación contendrían el 40% de los
episodios de captura y consumo de presas (intervalo 0,25 - 0,50 en las figuras 8 y 9),
mientras que las áreas críticas concentrarían mas del 50% de dichos episodios
(intervalos 0,50 - 0,75 y 0,75 - 1 en las figuras 8 y 9), si bien esta zonificación y sus
umbrales, es primeramente arbitraría, de acuerdo a lo establecido por Lusseau &
Higham (2004) en la determinación del hábitat crítico de descanso para T. truncatus
en Doubtful Sound (New Zealand), lo propuesto anteriormente tiene un basamento
biológico significativo, ya que deriva del presupuesto conductual de la población en
estudio. Para el caso de Golfo Dulce, la cuenca interna en su totalidad representaría
un hábitat crítico de alimentación con áreas núcleos (importantes y críticas) que
promueven la ocurrencia de episodios de captura y consumo de presas. En otras

75
palabras toda la cuenca contendría parches de presas potenciales, con
conglomerados importantes de presas en áreas focales claves.
5.c.2 Caracterización del nicho ecológico de alimentación
El nicho ecológico de alimentación del delfín manchado pantropical (S.
attenuata) esta definido por una batimetría típica de aguas profundas (> 100 m) y
fondos de pendiente muy suaves (< 20 %). Estas características claves se mantienen
durante la temporada de lluvia y la seca. Igualmente, hay una tolerancia a un
espectro de profundidades relativamente limitado, hasta aguas de profundidad
media. La captura y el consumo de presas no necesariamente se limitan por fondos
con pendientes empinadas. Evidentemente, las características fisiográficas
relacionadas a pendientes considerablemente empinadas, deben repercutir en
ventajas en la captura de presas para S. attenuata, tal como ocurre con otras
especies de odontocetos (Baumgarther 1997, Wilson et al. 1997, Ingram & Rogan
2002). Oviedo (2007) estableció que la isobata de los 100 m, era determinante en la
repartición de hábitat de S. attenuata y T. truncatus como estrategia de coexistencia
y supresión del efecto negativo de la competencia.
La información de los índices de marginalidad y tolerancia se sustentan con
las diferencias estadísticamente significativas evidenciadas con la prueba Kruskal
Wallis (Zar 1996), específicamente para la profundidad y la pendiente como factores
claves en la definición de hábitat. Lo anterior es consecuente con muchas
evaluaciones de hábitat en mamíferos marinos (Hui 1985, Davis et al. 2002; Cañadas et
al. 2002, 2005; , Hooker et al. 2002, Azzelino et al. 2008, Gómez de Segura et al. 2008,
Oviedo y Solís 2008). Al parecer el carácter estable de los atributos topográficos
(profundidad y pendiente del fondo oceánico) del ambiente marino, promueve la
relevancia de estos factores en la definición de áreas de distribución claves para
cetáceos, en contraposición a otras variables como la temperatura y salinidad, la
cuales pueden ser afectadas a nivel estacional, interanual, o incluso a nivel de micro-
escala temporal.

76
Otro aspecto con una relevancia relativa, es la distancia de los ríos más
cercanos a las localidades donde se desarrollaron los episodios de alimentación.
Acevedo & Burkhart (1998) y Cubero-Pardo (1998) consideraron el incremento de la
distancia a la costa como factor ambiental importante en la distribución espacial de
delfines manchados en Golfo Dulce. De acuerdo a las observaciones de este estudio,
más que la distancia relativa a la costa, la cercanía de los ríos que desembocan en la
cuenca interna de Golfo Dulce pareciesen afectar las áreas focales de alimentación
(efecto directo en la distribución de las presas potenciales) en ambas temporadas.
Los núcleos de ocurrencia de episodios de alimentación estarían relacionados por un
rango de distancia de 2 - 8 Km con los ríos Tigre, Agujas, Rincón y Esquinas
principalmente, a un nivel secundario con los ríos Barrigones y Caballero.
El mecanismo de afectación de la pluma de los ríos a las áreas focales de
alimentación no esta claro y requiere de mayor esfuerzo investigativo. Sin embargo,
si consideramos la información sobre circulación en la cuenca interna generada por
los modelos de Quirós (2003) y Svendsen et al. (2006) se esperaría que la marea de
flujo incrementara el efecto de la pluma del río Coto Colorado en la cuenca interna,
particularmente cerca de la costa noreste en las adyacencias de Punta Adela - Punta
Gallardo. Alternativamente, durante la marea de reflujo, y dada la velocidad de la
corriente, junto con el patrón de circulación anticiclónico, las plumas de los ríos Tigre
y Agujas afectarían las adyacencia de Punta Palma, la pluma del río Rincón ejercería
efectos sobre las zonas cercanas a Punta Estrella y Mogos, mientras que el río
Esquinas afectaría la costa noreste.
Un elemento importante a considerar son las implicaciones de las
características del nicho ecológico de alimentación, con lo que debería ser la
distribución y abundancia de presas. Las aguas profundas con pendientes suaves
deberían asociarse positivamente con la disponibilidad de presas pelágicas. Según lo
planteado por Giannoulaki et al. (2003), las cuencas semi-cerradas como el Golfo
Dulce, promueven agregaciones de pequeños pelágicos mucho más densas que en

77
áreas abiertas. Por otro lado las agregaciones densas de peces pelágicos tienden a
estructurarse en conglomerados compactos en aguas profundas. Las áreas de
batimetría llana inducen el “efecto nugget” el cual se caracteriza por la pérdida de la
estructuración de las agregaciones de pequeños pelágicos (Giannoulaki et al., 2003).
Lo anterior concuerda notablemente con la ocurrencia de los episodios de
alimentación cooperativa; si las presas se agregan en parches densos y
estructurados en aguas profundas, la forma más óptima de capturar y consumir tales
agregaciones, es por medio de un esfuerzo cooperativo donde el consumo per cápita
se balancea con la eficacia en la captura (Neumann y Orams 2003), con el beneficio
adicional de reducir la competencia entre con-específicos, tal como se mencionó en
secciones anteriores. Es importante mencionar que la ocurrencia de círculos de
alimentación como estrategia de captura y consumo de presas es notablemente
común en las observaciones de Golfo Dulce (cuenca cerrada y profunda), mientras
que en áreas externas de la Península de Osa, como la Bahía de San Josecito
(adyacencias al Parque Nacional Corcovado: cuenca abierta y de batimetría llana)
los episodios de alimentación son frecuentemente en grupos dispersos sin signos de
cooperación (observaciones del autor no publicadas).
Los delfines manchados utilizan el hábitat que representa la cuenca interna de
Golfo Dulce como un área de alimentación, de acuerdo a la relevancia de las
actividades que implican captura y consumo de presas en relación a otros contextos
etológicos en el presupuesto conductual. Los episodios de alimentación forman
patrones espaciales discernibles con focos de ocurrencia significativa, que permiten
establecer la localización de áreas de importancia y áreas críticas de alimentación.
Las áreas importantes y críticas componen el nicho ecológico de alimentación, este
se caracteriza por ubicarse en aguas profundas de fondos con pendientes
relativamente suaves, con algún grado relevante de influencia de las descargas de
los ríos del Golfo. El nicho de alimentación contendría presas pelágicas disponibles,
que constituyen más del 60% del consumo de presas de S. attenuata. Estas presas
forman parches de agrupaciones densas y compactas, dichos conglomerados

78
favorecen la captura de presas mediante estrategias cooperativas. Todo esto en
conjunto determinaría la categorización de Golfo Dulce como un hábitat crítico de
alimentación, debido a la relevancia de la zona para la subsistencia de la especie, en
específico por ser un área importante para el consumo de energía (Hoyt 2005). Lo
descrito anteriormente tiene implicaciones claves para el manejo, considerando que
esta contribución conceptualiza al hábitat como la unidad fundamental de
conservación por encima del nivel de especie.

79
CONCLUSIONES
Esta investigación contribuye al conocimiento disponible sobre la ecología del
delfín manchado pantropical, mediante el entendimiento de cómo estos delfines se
relaciona y usan su hábitat físico en Golfo Dulce, detallando la influencia de los
factores determinantes en la presencia de la especie en este importante ecosistema
marino, pero de una manera dinámica en donde se exploran las respuestas a los
estímulos directos e indirectos del medio.
La presencia de S. attenuata en la cuenca interna de Golfo Dulce es
notablemente significativa, considerando que la abundancia relativa, y los tamaños
de grupos observados, en relación al tipo de topografía del área de estudio como
cuenca semi-cerrada, deberían corresponder con el patrón de agregación de delfines
en áreas oceánicas abiertas. También es importante mencionar que el nivel de
consumo para sustentar la abundancia relativa de la especie en Golfo Dulce se
asemeja más a la de un ecosistema marino productivo, como el Golfo de Papagayo
al norte de la costa Pacífica de Costa Rica.
Los delfines manchados utilizan el hábitat que representa la cuenca interna de
Golfo Dulce como un área de alimentación, de acuerdo a la relevancia de las
actividades que implican captura y consumo de presas en relación a otros contextos
etológicos en el presupuesto conductual. S. attenuata en Golfo Dulce invierte mayor
tiempo y energía en capturar presas y alimentarse de ellas, mediante el
comportamiento típico de un consumidor especialista durante periodos de
abundancia, y de manera mixta (especialista - generalista) en la época seca.
La captura cooperativa de peces pelágicos, reduce notablemente el efecto
negativo de la competencia entre con-específicos. Esto en conjunto con la repartición
de hábitat, minimiza la presión por la competencia de recursos; tanto a nivel inter-
específico como intra-específico, un factor clave que puede prevenir la aptitud y
regular el crecimiento poblacional.

80
Los episodios de alimentación forman patrones espaciales discernibles con
focos de ocurrencia significativa, que permiten establecer la localización de áreas de
importancia y áreas críticas de alimentación. Lo anterior tiene implicaciones
importantes en la categorización de la cuenca interna de Golfo Dulce como hábitat
crítico de alimentación, debido a la relevancia de la zona para la subsistencia de la
especie, en específico por ser un área importante para el consumo de energía (Hoyt
2005).
Las áreas importantes y críticas componen el nicho ecológico de alimentación,
este se caracteriza por ubicarse en aguas profundas de fondos con pendientes
relativamente suaves. Se estima que existe un grado relevante de influencia en el
nicho ecológico de las descargas de los ríos del golfo.
El nicho de alimentación contendría presas pelágicas disponibles, que
constituyen más del 60% del consumo de presas de S. attenuata. Estas presas
forman parches de agrupaciones densas y compactas, dichos conglomerados
favorecen la captura de presas mediante estrategias cooperativas.
Lo anterior no solo refleja el uso de hábitat como área crítica de alimentación,
también aporta evidencias directas (abundancia relativa, nivel de consumo de
presas, tipo de presas consumidas) e indirectas (conducta alimentaría, agregación de
los parches de presas) de una mayor productividad en Golfo Dulce, de la que se
había documentado anteriormente (Wolff et al. 1996, Quesada-Alpizar y Cortés
2006).
La presencia en números importantes de depredadores esta intrínsecamente
asociado con una abundancia de presas que sustente el consumo de energía
requerido por la población local. Estos consumidores efectivamente tienden a
desarrollar conductas especializadas en ecosistemas productivos (Begon et al.,
2006). En el caso de Golfo Dulce, tal productividad pareciese estar ligada a un mayor

81
caudal y trasporte de nutrientes en los ríos en la temporada de lluvia, que disminuye
en la época seca.

82
IMPLICACIONES PARA EL MANEJO Y CONSERVACIÓN
Este estudio es una evaluación ecológica de una especie en interacción
dinámica con su hábitat, que tiene como finalidad consolidar una línea base para
sustentar acciones de manejo. El delfín manchado pantropical no es solo relevante
como ente biológico que aporta información del estado del ecosistema, también es
un recurso importante para una economía cada vez más dependiente del turismo.
Dos líneas básicas se proponen preliminarmente como estrategias, una
relacionada con un enfoque de manejo y conservación de la especie, desde la
perspectiva de los delfines manchados como recurso marino costero con potencial
para uso no consuntivo (observación de cetáceos). El otro aspecto se basa en un
enfoque más ecosistémico, que promueve el manejo y conservación del hábitat
crítico de la especie en Golfo Dulce.
7.a Delfines manchado como sostén de la industria de observación de cetáceos en Golfo Dulce
De acuerdo a Hoyt y Iñíguez (2008), el crecimiento de la observación de
cetáceos como actividad turística en la zona del sudoeste del Pacífico de Costa Rica,
se relaciona al hecho de que ciertos hoteles y albergues para huéspedes tenían sus
propias embarcaciones para ofrecer avistamiento de cetáceos. Adicionalmente, los
pescadores adaptaron sus botes para la actividad. Según los mismos autores, en
Puerto Jiménez; el principal centro focal de operadores turísticos en la zona
correspondiente a Golfo Dulce, aproximadamente siete botes generan el grueso del
beneficio de la actividad (> 900.000 $).
Las excursiones de observación de cetáceos se focalizan en los encuentros
con S. attenuata, y de manera complementaria con avistamientos de T. truncatus.
Aparentemente, la preferencia la sustenta el número mayor de delfines por
avistamiento de delfines manchados, y el mayor grado de eventos conductuales
notablemente llamativos (saltos). Efectivamente se ha observado un incremento

83
aparente de conductas percusivas durante las excursiones de avistamiento de
delfines en Golfo Dulce (observaciones y datos no publicados del autor). Las
conductas percusivas generan sonidos por choques entre la masa corporal del delfín
y el agua, probablemente como una forma importante de comunicación no vocal.
Si el costo energético de realizar demostraciones percusivas se relaciona con
actividades diarias inherentes a la historia de vida de la especie, estas son
directamente relacionadas con la subsistencia y la aptitud. Si el costo se relaciona
con una respuesta inducida por perturbación de origen antropogénico, entonces
habría una inversión de energía que debería ser dirigida hacia aspectos con mayor
valor intrínseco para la subsistencia, es decir habría un costo energético en la
interacción de los delfines con botes turísticos que no se justifica en términos
biológicos de supervivencia (Lusseau 2006).
Más importante aun sería la afectación del contexto relacionado con el
consumo de energía (alimentación). Coscarella et al. (2006) documentaron la
alteración de episodios de captura y consumo de presas en los delfines oscuros (L.
obscurus) en Golfo Nuevo, Argentina. Particularmente por interacción con botes
turísticos. Episodios similares se observan en Golfo Dulce, en repetidas ocasiones
los círculos de alimentación de delfines manchados han sido interrumpidos por botes
de turismo tratando de inducir un nivel más alto de interacción con el bote, o con
nadadores en el agua; actividad turística ilegal según la norma establecida en el
artículo 8 del reglamento No 32495.
Se podría argumentar el desconocimiento tanto de aspectos biológicos claves
de la especie, como del marco legal en los casos mencionados anteriormente. Sin
embargo, el conocimiento empírico de capitanes y dueños de embarcaciones locales
es amplio, por lo que es probable que el reconocimiento de conductas inherentes a
actividades como la alimentación sea efectivo, igualmente no han escaseados los
entrenamientos relativos a la regulación de la observación de cetáceos como
actividad turistica, ONGs (como “Fundación Keto”, “Fundación PROMAR”) han

84
ofrecido talleres al respecto, auspiciados por entes estatales como las oficinas de las
áreas de conservación del Ministerio del Ambiente y Energía (MINAE).
El aspecto que no se ha considerado a profundidad, es el efecto del
consumidor y la demanda en la oferta del servicio (observación de cetáceos no
regulada con nado como extra). Las embarcaciones responden a operadores
(hoteles y alberges), que ofertan la actividad bajo parámetros no concordante con el
marco legal existente a un consumidor totalmente ignorante de la existencia de las
regulaciones. Lo que deviene en turistas que esperan nadar y ver delfines saltando
muy cerca, agregando una presión adicional causada principalmente por la
desvirtualización del ecoturismo como actividad contemplativa no intrusiva.
Hoyt y Iñíguez (2008) ponderan la necesidad de revisar la capacidad de carga
de muchas operaciones locales de turismo relacionadas a cetáceos, principalmente
por el crecimiento acelerado de la actividad en Costa Rica. Un elemento base para
la determinación de la capacidad de carga es el presupuesto conductual (Lusseau
2003), el cual determina la línea base para establecer comparaciones con
presupuestos generados a posteriori y considerando como variable interviniente la
interacción con botes. Por lo tanto, de acuerdo a los resultados presentados
anteriormente, una reducción menor al 40% de la ocurrencia de actividades de
alimentación, ante la presencia de embarcaciones turísticas, representaría un indicio
claro de perturbación antropogénica.
En este análisis no se descarta el efecto de la línea base desplazada (Pauly
1995), Golfo Dulce no es un ecosistema enteramente prístino, y las poblaciones de
delfines han tenido contacto con las comunidades locales históricamente, mucho
antes de la realización de esta investigación. Sin embargo el marco metodológico del
presente trabajo, particularmente condicionado por las variables consideradas en el
diseño de los muestreos etológicos, las cuales excluyen la interacción con
embarcaciones, incluso con la embarcación de investigación, otorga solides y
representatividad a los estimados de los presupuestos conductuales generados.

85
Como se ha mencionado inicialmente, la finalidad de esta contribución es
aportar línea base, la determinación del grado de afectación por interacción
antropogénica escapa a los objetivos planteados en la investigación. Sin embargo la
metodología usada es reproducible y adaptable para evaluaciones ante la presión de
embarcaciones turísticas. Por lo que se recomienda a partir de la información
generada en este estudio, proseguir en los análisis de conducta aplicados a la
determinación de la capacidad de carga.
La utilidad de las licencias y permisos para la observación de cetáceos son
elementos establecidos dentro del marco legal existente en Costa Rica. Sin embargo,
necesitan de ser reivindicados en términos de aplicabilidad. Tal como lo plantean
Lusseau y Higham (2004), la zonificación espacial en áreas importantes y áreas
críticas de zonas de relevancia para la conservación, es un paso clave en la
promoción de la aplicabilidad efectiva de permisos y licencias. Si el acceso a áreas
catalogadas en base al uso diferencial de hábitat, es garantizado solo a
embarcaciones regularizadas con el sistema de licencias, se optimiza el control y por
lo tanto, el manejo del recurso. Esta no es la situación actual en Golfo Dulce, no hay
discriminación en el transito (ni en la velocidad de transito) a las diversas áreas de
este ecosistema marino. Cualitativamente la cantidad de embarcaciones en
circulación, es en apariencia baja, pero esto requiere de una revisión exhaustiva. Es
importante considerar los planes de desarrollo de marinas e instalaciones náuticas
proyectados para la localidad de Golfito, es de esperarse entonces el incremento de
embarcaciones turísticas en transito, de manera proporcional al incremento de
visitantes.

86
7. b La cuenca interna del Golfo Dulce: una Área Marina Protegida (AMP)
En los capítulos anteriores se destacó enfáticamente que los atributos y
características ecológicas de la población de delfines estudiada, no concuerdan con
las que habitarían un ecosistema poco productivo. La productividad es relevante en
el mantenimiento de la diversidad biológica (Ricklefs 2001) y esto es particularmente
cierto para Golfo Dulce. Por ejemplo, datos de carácter anecdóticos aportados por
lugareños, dan referencia de Golfo Dulce como zona de cría de tiburones.
Igualmente, es del conocimiento común y local (incluso a nivel del SINAC), la
relevancia del Golfo como área de desove de peces agujas (Belonidae), los cuales
son primordial como recurso trófico (Pacheco y Oviedo 2007).
Las Áreas Marinas Protegidas (AMPs) son espacios delimitados del ambiente
marino que son manejados para la conservación de la biodiversidad (Hoyt 2005). La
importancia de las AMPs está principalmente asociada con la conservación de la
biodiversidad, los recursos pesqueros, y como herramientas de investigación y
manejo. Complementariamente, las AMPs aportan beneficios inherentes a aspectos
recreativos, estéticos y educativos. En base a lo anterior, las pesquerías se pueden
beneficiar por la retroalimentación de una productividad óptima hacia las adyacencias
externas de las Áreas Marinas Protegidas (AMP), debido el incremento en la
dispersión de huevos y larvas. Edgar et al. (2007), pondera que dicho incremento, es
en muchos aspectos un seguro contra un colapso en los stocks pesqueros.
Los resultados presentados previamente ilustran claramente la importancia de
las zonas de influencia de los ríos mayores que desembocan en el Golfo como
hábitat crítico de alimentación para delfines manchados. Las acciones de
conservación y manejo, particularmente aquellas dirigidas al establecimiento de
AMPs, deben basarse en la determinación de hábitats críticos. Este concepto en si
mismo, aplicado a la conservación de cetáceos, es relativamente reciente, aun por
ser explorado y escasamente implementado (Hoyt 2005).

87
Lo discutido anteriormente tiene un efecto importante en el valor del delfín
manchado como una especie bandera – sombrilla, ya que su distribución en el
ecosistema marino de Golfo Dulce incrementa las posibilidades de cohabitar con
otras especies simpátricas que podrían compartir el mismo hábitat o diferentes
porciones del mismo. Dichas especies simpátricas se beneficiarían de cualquier
medida de manejo y conservación que se le otorguen a S. attenuata. Sin embargo
este papel como especie bandera y sombrilla se optimizaría y complementaría a
través de un conjunto de especies marinas paisajes, tal como se plantea en el
enfoque de las especies paisajes (Wilkie 2004, Oviedo 2005).
La factibilidad de establecer una AMP en Golfo Dulce es significativa.
Esfuerzos de ONGs nacionales e internacionales han generado una propuesta de
decreto de creación de dos nuevas categorías de manejo para áreas marinas, más
acorde con las necesidades de gestión actuales de las áreas marinas protegidas
existentes y en proyecto (como la AMP Pacífico Sur). Entre estas nuevas categorías
está la Reserva Marina, que busca la conservación de los ecosistemas y el hábitat
para la protección de especies. La categoría de reserva marina no descarta la
satisfacción de las necesidades de las comunidades locales (y en general), así como
los elementos que determinan su calidad de vida. Por lo que comprende los usos
sostenibles de los recursos marinos costeros y oceánicos (a través de la educación,
la investigación científica, y el ecoturismo de bajo impacto). Según lo estipulado en el
Artículo 32 y 36 de la Ley Orgánica del Ambiente, las categorías de manejo, son
factibles de establecer por medio de decreto, si se requiere la creación, ampliación, o
cambio de categoría de algún área.
En base a lo anterior es importante considerar que se requiere actualizar y
reforzar la información de línea base a nivel ecosistémico en Golfo Dulce, se debe
incrementar el conocimiento sobre el aporte y subsidio a la productividad de origen
fluvial. De lo contrario, se corre el riesgo de prevenir una estrategia efectiva de
manejo del Golfo Dulce como ecosistema. De acuerdo a lo planteado en párrafos

88
anteriores, el anexo 6 presenta de manera esquematizada, las líneas de acciones
sugeridas en términos de manejo y conservación.

89
LITERATURA CITADA
Acevedo-Gutierrez, A. and Smultea, M.A. 1995. First records of humpback whales
including calves at Golfo Dulce and Isla del Coco, Costa Rica, suggesting
geographical overlap of northern and southern Hemisphere populations. Marine
Mammals Science 11: 554 - 560.
Acevedo-Gutierrez, A. 1996. Lista de mamíferos marinos en Golfo Dulce e Isla del Coco,
Costa Rica. Revista de Biología Tropical 44: 933 - 934.
Acevedo-Gutierrez, A. and Burkhart, S. 1998. Seasonal distribution of bottlenose
(Tursiops truncatus) and pan-tropical spotted (Stenella attenuata) dolphins (Cetacea:
Delphinidae) in Golfo Dulce, Costa Rica. Revista de Biología Tropical 46: 91-101.
Acevedo-Gutiérrez, A., and Matthew, A. 2005. Association pattern of bottlenose dolphins
in Costa Rica: Constant Companions and Casual Acquaintances. Book of abstract
XVI Biennial Conference on the Biology of Marine Mammals, San Diego, California.
215 pp.
Altmann, J. 1974. Observational study of behavior: sampling methods. Behavior 49: 227 -
267.
Aragones, L., Jefferson T.A. and Marsh H.1997. Marine mammals survey techniques
applicable in developing countries. Asian Marine Biology 14:15 – 39.
Au, D.W.K. and W.L. Perryman. 1985. Dolphin habitats in the Eastern Tropical Pacific.
Fishery Bulletin 83: 623 - 643.
Azzelinno, A., Gaspari, S., Airoldi, S. and B. Nani. 2008. Habitat use and preferences of
cetaceans along the continental slope and the adjacent pelagic waters in the western
Ligurian Sea. Deep-Sea Research I 55: 296 – 323.

90
Baumgarther, M.F. 1997 The distribution of Risso´ dolphin (Grampus griseus) with
respect to the physiography of the northern gulf of Mexico. Marine Mammals Science
13 (4):614 – 638.
Bearzi, M. 2005a. Habitat Partitioning by Three Species of Dolphins in Santa Monica
Bay, California. Bulletin of Southern California Academic of Science 104: 113 -124.
Bearzi, M., 2005b. Aspects of the ecology and behaviour of bottlenose dolphins (Tursiops
truncatus) in Santa Monica Bay, California. Journal Cetacean Research and
Management 7(1): 75 -8 3.
Begon, M., Townsend, C. R. and J. L. Harper. 2006. Ecology: from individuals to
ecosystems.4th ed. Blackwell Publishing, U.K. 740 pp.
Calambokidis, J., K. Rasmussen and G.H. Steiger. 2002. Humpback whales and other
marine mammals off Costa Rica, 1996-1999. Report of research during Oceanic
Society Expeditions in 2002 in cooperation with Elderhostel volunteers. Cascadia
Res. Collec. 33 p.
Cañadas, A., Sagarminaga, R. and S. García-Tiscar. 2002. Cetacean distribution related
with depth and slope in Mediterranean waters off sourthern Spain. Deep Sea
Research I 49: 2053 – 2073.
Cañadas, A., Sagarminaga, R., De Stephanis, R., Urquiola, E. and P.S. Hammond 2005.
Habitat preference modelling as a conservation tool: proposal for marine protected
areas for cetaceans in southern Spanish waters. Aquatic Conservation: Marine and
Freshwater Ecosystems 15: 495 – 521.

91
Coscarella, M.A., Dans, S.L., Crespo, E.A. and S.N. Pedraza. 2003. Potential impact of
dolphin watching unregulated activities in Patagonia. Journal of Cetacean Research
and Management 5(1):77 – 84.
Compton R.C. 2004. Predicting key habitat and potential distribution of northern
bottlenose whales (Hyperoodon ampullatus) in the northwest Atlantic ocean. MSc
Thesis. University of Plymouth. U.K. 40 p.
Cubero Pardo, P. 1998a. Distribución y patrones de actividad del bufeo (Tursiops
truncatus) y el delfín manchado (Stenella attenuata) en el Golfo Dulce. Tesis de
maestría. Universidad de Costa Rica. 102p.
Cubero-Pardo, P. 1998b. Patrones de comportamiento diurnos y estacionales de
Tursiops truncatus y Stenella attenuata (Mammalia: Delphinidae) en el Golfo Dulce,
Costa Rica. Revista de Biología Tropical 46: 103 - 110.
Cubero Pardo, P. 2007. Distribución y condiciones ambientales asociadas al
comportamiento del delfín bufeo (Tursiops truncatus) y el delfín manchado (Stenella
attenuata) (Cetacea: Delphinidae) en el Golfo Dulce, Costa Rica. Revista de Biología
Tropical 55:549 - 557.
Darling. 2000. Migratory destinations of humpback whales that feed off California,
Oregon and Washington. Marine Ecology Progress Series 192: 295 - 304.
Daura Jorge, F. G. 2007. “A dinâmica predador-presa e o comportamento do boto-cinza,
Sotalia guianensis (cetacea, delphinidae), na baía norte da ilha de santa catarina, sul
do brasil. Tesis de Maestría, Universidad Federal de Paraná. Curitiba. pp 104.
Davis, R. W., Ortega Ortiz, J.G., Ribie, C. A., Evans, W.E., Biggs, C.D., Ressler, P.H.,
Cady, R.B., Leben R.R., Mullin D.K and Würsig, B. 2002 Cetacean habitat in the
northern oceanic Gulf of Mexico. Deep Sea Research I 49: 121 – 142

92
Edgar, G.F., G.R. Russ and R.C. Babcock. 2007. Marine Protected Areas In: S.D.
Connell and B.M. Gillanders Editors. Marine Ecology. Oxford University Press,
Melbourne, Australia.
Escorza-Trevino, S., A. Lang and Dizon, A.E. 2002. Genetic differentiation and
intraspecific structure of eastern tropical Pacific spotted dolphins, Stenella attenuate
revealed by mitochondrial and microsatellite DNA analyses. SWFSC Administrative
Report, La Jolla, LJ-02-38. 20 p.
Evans, P.G.H and Hammond, P. 2004 Monitoring cetacean in European waters.
Mammals Review 34: 131 - 156.
Fiedler, P. C., And Reilly, S. B. 1994. Interannual variability of dolphin habitats in the
eastern tropical Pacific. II: effects on abundances estimated from tuna vessel
sightings, 1975–1990. Fishery Bulletin 92: 451 – 463.
Gallo, J. P. 1991. Group behavior of Common dolphins Delphinus delphis during prey
capture. Anales del Instituto de Biología Universidad Nacional Autónoma de México,
Serie Zoología 62: 253 – 262.
García, C. and Dawson, S. 2003 Distribution of pantropical spotted dolphins in Pacific
coastal waters of Panama. Latin American Journal of Aquatic Mammals 2: 29 – 38
Garaffo, G., S. L, Dans, S. N., Pedraza, E. A., Crespo and M., Degrati. 2007. Habitat use
by dusky dolphin in patagonia: how predictable is their location. Marine Biology 152:
165 - 177
Giannoulaki, M., Machias, A. Koutsikopoulos, C. Haralabous, J., Somarakis, S. and N.
Tsimenides. 2003. Effect of coastal topography on the spatial structure of the
populations of small pelagic fish. Marine Ecology Progress Series 265: 243 – 253.

93
Gowans, S. and H. Whitehead. 1995. Distribution and habitat partitioning by small
odontocetes in the Gully a submarine canyon in the Scotian Shelf. Canadian Journal
of Zoology 73: 1599-1608.
Guisan A. and N. Zimmermman. 2000. Predictive habitat distribution models in ecology.
Ecological Modelling 135: 146 - 186.
Grerg, E.J. and Trites A. W. 2001. Prediction of critical habitat for five whales speciesin
the waters of coastal British Columbia. Canadian Journal of Fishery and Aquatic
Science 58: 1265–1285.
Griffin, R.B. and N.J. Griffin. 2003. Distribution, habitat partitioning, and abundance of
atlantic spotted dolphins, bottlenose dolphins, and loggerhead sea turtles on the
eastern gulf of mexico continental shelf. Gulf of Mexico Science 2003(1): 23 – 34.
Hastie, G.D., Wilson, B., Wilson, L.J., Parsons, K.M., and Thompson, P.M. 2004.
Functional mechanisms underlying cetacean distribution patterns: Hotspots for
bottlenose dolphins are linked to foraging. Marine Biology 144: 397- 403.
Henrys P. 2005. Spatial distance sampling modeling of cetaceans observed from
platforms of opportunity. Mimiografiado. 52 pp. (Disponible del autor).
Hirzel, A.H., Helfer, V., and F. Metral. 2001. Assessing habitat-suitability models with a
virtual species. Ecological Modelling 145: 111 - 121.
Hirzel, A.H., Hausser, J., Chessel, D. and N. Perrin. 2002. Ecological-niche factor
analysis: How to compute habitat-suitability maps without absence data? Ecology 83:
2027 - 2036.

94
Hohn, A. A., Chivers, S. J. and Barlow, J. 1985. Reproductive maturity and seasonality of
male spotted dolphins, Stenella attenuata, in the eastern tropical Pacific. Marine
Mammals Science 1: 273 – 293.
Hooker, S.K., Whitehead, H. and Gowans, S. 2002. Ecosystem consideration in
conservation planning: energy demand of foraging bottlenose whales (Hyperoodon
ampullatus) in a Marine Protected Area. Biological Conservation 104: 51 - 58.
Hoyt, E. 2005. Marine Protected Areas for whales, dolphins and porpoises: a world
handbook for cetacean habitat conservation. Earthscan, U.K. 512 pp.
Hoyt, E. and M. Iñíguez. 2008. Estado del Avistamiento de Cetáceos en América Latina.
WDCS, Chippenham, UK; IFAW, East Falmouth, EE.UU.; y Global Ocean, Londres,
60p.
Hui, C. 1979. Undersea topography and distribution of dolphins of the genus Delphinus in
the southern California Bight. Journal of Mammalogy 60 (3): 521 – 527.
Hui, C.A. 1985. Undersea topography and the comparative distributions of two pelagic
dolphins. Fishery Bulletin 83: 472 – 475
Ingram, S. and E. Rogan. 2002. Identifying critical areas and habitat preferences of
bottlenose dolphins Tursiops truncatus. Marine Ecology Progress Series 244: 247 -
255.
Karczmarski, L., Cockcroft, V.G. and A. McLachlan. 2000. Habitat use and preferences of
Indo-Pacific humpback Sousa chinensis in Algoa Bay, South Africa. Marine Mammals
Science 16(1): 65 – 79

95
Karczmarski, L., Wursig, B., Gailey, G., Larson, K.L.c and C. Vanderlip. 2005. Spinner
dolphins in a remote Hawaiian atoll: social grouping and population structure.
Behavioral Ecology : 675 - 685.
Kaschner, K. 2004. Modelling and mapping resource overlap between marine and
fisheries on a global scale. PhD Thesis. University of British Columbia. Canada. 225p.
Kasuya, T. 1976. Reconsideration of life history parameters of the spotted and striped
dolphins based on cemental layers. Scientific Reports of the Whales Research
Institute, Tokyo 28: 73 – 106.
Kasuya, T., Miyazaki, N. and Dawbin, W. H. 1974. Growth and reproduction of Stenella
attenuata in the Pacific coast of Japan. Scientific Reports of the Whales Research
Institute Tokyo 26: 157 – 226.
Krausman, P. 1999.Some basic principles of habitat use in Grazing Behavior of Livestock
and Wildlife. Idaho Forest, Wildlife and Range. (K.L. Launchbaugh,K.D. Sanders, J.C.
Mosley.eds) Univ. of Idaho, Moscow, ID.
Lammers, M.O., Au, W.W.L. and Feinholz, D. 2000. The occurrence and distribution of
marine mammals along Oahu’s Ewa/Honolulu coast: a study to assess the potential
interactions between high-speed ferry traffic and local populations. MMRP/HIMB
Technical Report 20001. 20 pp.
Lusseau D. effects of tour boats on the behavior of bottlenose dolphins: using Markov
chains to model anthropogenic impact. Conservation Biology 17(6):1785 - 1793.
Lusseau, D. and J.E.S. Higham. 2004. Managing the impacts of dolphin-based tourism
through the definition of critical habitats: the case of Doubtful Sound. New Zealand.
Tourism Management 25(6): 657 – 667

96
Lusseau, D., Slooten, E., and R.J. Currey. 2006. Unsustainable dolphin watching
activities in Fiordland, New Zealand. Tourism in Marine Environments 3(2): 173 -178.
Mann, Janet.1999. Behavioral Sampling methods for Cetacean: a Review and Critique.
Marine Mammals Science 15(1): 102 – 122.
Mann, Janet. 2000. Unraveling the dynamics of social life: long-term studies and
observational methods. In: Mann, J., Connor, R.C., Tyack, P.L., Whitehead, H.
Editors. Cetacean Societies. University of Chicago Press, London.
May-Collado, L.J. and A.R. Morales. 2005. Presencia y patrones de comportamiento del
delfin manchado costero, Stenella attenuata graffmani (Cetacea: Delphinidae) en el
Golfo de Papagayo, Costa Rica. Revista de Biología Tropical 53: 265 - 276.
May-Collado, L., T. Gerrodette, J. Calambokidis, K. Rasmussen, and Sereg, I. 2005.
Patterns of cetacean sighting distribution in the Pacific Exclusive Economic Zone of
Costa Rica, based on data collected from 1979-2001. Revista de Biología Tropical
53: 249 -263.
Mignucci, A. 1998. Zoogeography of cetaceans off Puerto Rico and the Virgin Islands.
Caribbean Journal of Science 34(3-4): 173 – 190.
Mignucci-Giannoni, A. Swartz, S. L., Martínez , A. Burks, C. M. and Watkins, W. A. 2003.
First records of the pantropical spotted dolphin (Stenella attenuata) for the Puerto
Rican Bank, with a review of the species in the Caribbean. Caribbean Journal of
Science 39(3): 381 - 392.
Montero, A. 2005 Potential effect of whale-watching tours and other recreational activities
on dolphins at Bahía Drake, Costa Rica: a case for the spotted dolphin, Stenella
attenuata. Marine Mammals News 134: 17 - 18.

97
Neumann D. and M. Orams. 2003. Feeding behaviours of short-beaked common
dolphins, Delphinus delphis, in New Zealand. Aquatic Mammals 29: 137 – 149.
Ojasti, J. 2000. Manejo de Fauna Silvestre Neotropical. F.Dallmeier (ed.). SIMAB Serie
No.5. Smithsoniam Intitution/MAB Program, Washington. D.C.
Oviedo, L., Silva, N., Bermúdez, L. and Odell, D.K. (2005). Distribution of bottlenose
dolphins (Tursiops truncatus) on the east coast of Isla margarita and the Los Frailes
Archipelago, Venezuela. Aquatic Mammals 31: 442 - 446.
Oviedo, L. 2005. Cetaceans as seascape species in the northeast coast of Venezuela:
preliminary assessment based on the seascape species approach. Book of Abstract,
XVI Conference on the Biology of Marine Mammals, San Diego, California. 215 pp.
Oviedo, L. 2006. Ecological aspects of pantropical spotted dolphins Stenella attenuata in
Golfo Dulce, Costa Rica: behavior ecology, distribution and abundance. Marine
Mammals News 142: 7.
Oviedo, L. 2007. Dolphin sympatric ecology in a tropical fjord: habitat partitioning by
bathymetry and topography as a strategy to coexist. Journal of Marine Biological
Association of the United Kingdom 87: 1 - 9.
Oviedo, L. and M. Solís. 2008 Underwater topography determines critical breeding
habitat for humpback whales near Osa Peninsula, Costa Rica: implications for Marine
Protected Areas. Revista de Biología Tropical 56: 591 - 602.
Palka, D.L. and Hammond, P.S. 2001. Accounting for responsive movement in line
transect estimates of abundance. Canadian Journal of Fishery and Aquatic Science
58: 777 – 787.

98
Pauly, D. 1995. Anecdotes and the shifting baseline syndrome of fisheries. Trends in
Ecology and Evolution 10(10): 430.
Pauly, D., Trites, A. W., Capuli, E. andV. Christensen. 1998. Diet composition and trophic
levels of marine mammals. ICES Journal of Marine Science 55: 467– 481.
Perrin, W. F. 1970. Color pattern of the eastern Pacific spotted porpoise Stenella
graffmani Lönnberg (Cetacea, Delphinidae). Zoologica 54:135 – 149.
Perrin, W. F. 2001. Stenella attenuata. Mammalian Species 683:1–8.
Perrin, W. F., Coe, J. M. and Zweifel, J. R. 1976. Growth and reproduction of the spotted
porpoise, Stenella attenuata, in the offshore eastern tropical Pacific. Fishery Bulletin
74: 229 – 269.
Perrin, W. F. and Reilly S.B. 1984. Reproductive parameters of dolphins and small
whales of the family Delphinidae. Report of the International Whaling Commission
Special Issue 6: 97 – 133.
Perrin, W. F. and Hohn, A. A. 1994. Pantropical spotted dolphin Stenella attenuata. In: S.
H. Ridgway and R. Harrison Editors. Handbook of marine mammals: the first book of
dolphins. Academic Press, London, United Kingdom.
Perrin, W. F., Schnell, G. D., Hough, D. J. Gilpatrick, Jr. J. W. and Kashiwada, J. V. 1994.
Reexamination of geographic variation in cranial morphology of the pantropical
spotted dolphin, Stenella attenuata, in the eastern Pacific. Fishery Bulletin 92: 324 –
346.
Quesada M.A. and J. Cortes. 2006. Los ecosistemas marinos del Pacifico sur de Costa
Rica: estado del conocimiento y perspectivas de manejo. Revista de Biología
Tropical 54 (Suppl. 1): 101 - 145.

99
Quesada, M.A., J. Cortes, J.J. Alvarado and A.C. Fonseca. 2007. Características
hidrográficas y biológicas de la zona marino-costera del área de conservación Osa.
The Nature Conservancy, Serie Técnica 2, San José. 79 p.
Quiros, G. 2003. Circulación del Golfo Dulce: un fiordo tropical. Topografía Meteorología
y Oceanografía 10 (2): 75 - 83
Rasmussen, K., J. Calambokidis and G. Steiger. 2001. Humpback whales and other
marine mammals off Costa Rica and surrounding waters, 1996-2002. Report of the
Oceanic Society 2001 field season in cooperation with Elderhostel volunteers. Dec.
2001. 21 p.
Rasmussen, K., J. Calambokidis and G. Steiger. 2004. Humpback whales and other
marine mammals off Costa Rica and surrounding waters, 1996-2002. Report of the
Oceanic Society 2001 field season in cooperation with Elderhostel volunteers. Apr.
2004. 21 p.
Read, A. J. and C. R. Brownstein. 2003. Considering other consumers:fisheries,
predators, and Atlantic herring in the Gulf of Maine. Conservation Ecology 7(1): 2.
[online] URL: http://www.consecol.org/vol7/iss1/art2
Redfern JV, Ferguson MC, Becker EA, Hyrenbach KD, Good C, Barlow J, Kaschner K,
Baumgartner MF, Forney KA, Ballance LT, Fauchald P, Halpin P, Hamazaki T,
Pershing AJ, Qian SS, Read A, Reilly SB, Torres L. and F. Werner 2006. Techniques
for cetacean-habitat modeling. Marine Ecology Progress Series 310: 271 – 295.
Reilly, S. B. 1990. Seasonal changes in distribution and habitat differences among
dolphins in the eastern tropical Pacific. Marine Ecology Progress Series 66:1 – 11.
Ricklefs, R. 2001. Invitación a la Ecología: la economía de la naturaleza. Editorial
Panamericana, 4ta edición. Madrid. 681pp.

100
Robertson, K.M. and Chivers, S.J. 1997. Prey occurrence in pantropical spotted dolphins,
Stenella attenuata from the eastern tropical Pacific. Fishery Bulletin 95: 334 -348
Rodríguez Sáenz, K. and J. Rodríguez-Fonseca. 2004. Avistamientos del delfín
manchado, Stenella attenuata (Cetacea: Delphinidae) en Bahía Culebra, Costa Rica,
1999-2000. Revista de Biología Tropical 52 (Suppl. 2): 189 - 193.
Rosenweig, M. L. 1981. A theory of habitat selection. Ecology 62: 327 - 335.
Salkind, N.J. 1999. Métodos de Investigación, 3era Edición. Prentice may. México. pp 4 -
14.
Scheneider, K. 1999. behaviour and ecology of bottlenose dolphin in Doubtful Sound,
Fiorland, New Zealand. PhD Thesis. University of Otago, New Zealand. 211p.
Gómez de Segura, A., Hammond, P.S. and J.A. Raga. 2008. Influence of environmental
factors on small cetacean distribution in the Spanish Mediterranean Journal of Marine
Biological Association of the United Kingdom 88: 1 - 8.
Shane, S. 1990. Behavior and ecology of bottlenose dolphin at Sanibel Island, Florida. In:
S. Leatherwood and R. R. Reeves Editors. The bottlenose dolphin. San Diego:
Academic Press.
Sierra, C., Vartanián, D. and J. Polimeni. 2006. Caracterización social, económica y
ambiental del Área de Conservación de Osa. Dirección de Sociedad Civil, Ministerio
del Ambiente y Energía, San José, Costa Rica. 2da edición. 200pp.
Smolker, R. A., Richards, A. F., Connor R. C., and Pepper, J. W. 1992. Sex differences
in patterns of association among Indian Ocean bottlenose dolphins. Behavior 123: 38
- 69.

101
Steiger, G.H.; J. Calambokidis, R. Sears; K.C. Balcomb and J.C. Cubbage.1991.
Movements of humpback whales between California and Costa Rica. Marine
Mammals Science 7: 306 - 310.
Svendsen, H., R. Rosland, S. Myking, J.A. Vargas, O.G. Lizano and E.J. Alfaro. A
physical-oceanographic study of Golfo Dulce, Costa Rica. Revista de Biología
Tropical 54 (Suppl. 1): 147-170.
Wang, M., Walker, W.A., Shao, K. and Chou, L. 2003. Feeding habits of pantropical
spotted dolphins, Stenella attenuata, off the eastern coast of Taiwan. Zoological
Studies 42: 368 - 378.
Wilkie, D. 2004. Belize Threats Assessment and Mapping - Highlights. Wildlife
Conservation Society, Marine Program, Belize City. 6p.
Wilson, B., Thompson, P.M. and P.S. Hammond. 1997 Habitat use by bottlenose
dolphins: seasonal distribution and stratified movement patterns in the Moray Firth,
Scotland. Journal of Applied Ecology 34: 1365 – 1374.
Wolff, M., Hartman, H.J. and Koch, V. 1996. A pilot trophic model for Golfo Dulce, a
tropical fjord-like embayment, Costa Rica. Revista de Biología Tropical 44, suppl.3:
215 – 231.
Zar, J.H. 1996. Biostatistical analysis, 3rd edn. Prentice-Hall International, U.K. 262pp.

102
ANEXOS
Anexo 1. Descripción del delfín manchado pantropical (Stenella attenuata).
Comparación de los patrones de coloración de delfines manchados Stenella
attenuata (parte superior) y Stenella frontalis (Delfines en la parte inferior). Ilustración
de Pieter Arends Folkens (tomado de Mignucci et al. 2003).
El delfín manchado pantropical es un delfín pequeño y delgado que se
caracteriza por manchas conspicuas a lo largo de su anatomía corporal. En
individuos notablemente manchados de la subespecie costera del Pacífico oriental, el
patrón de coloración de fondo se puede obscurecer en la región dorsal. El patrón de
manchas se puede extender a la aleta dorsal y aletas pectorales. El patrón de
coloración en la etapa de nacimiento es gris oscuro arriba y marfil abajo; las
extremidades son gris oscuro (Perrin 1970). La aleta dorsal es fuertemente falcada y
relativamente estrecha. Como se muestra en la figura anterior.
El delfín manchado pantropical varía geográficamente de tamaño así como en
el grado de manchas y es sexualmente dimorfo en tamaño (Perrin y Hohn 1994;
Perrin y Reilly 1984). En el Pacífico oriental la longitud promedio de machos adultos

103
de la subespecie costera y oceánica es de 223 cm y 200 cm respectivamente. Para
hembras las longitudes de la subespecie costera es de 207 Cm y 182 cm para la
subespecie oceánica. En el Golfo de México, Pacífico noroeste, Océano indico
suroeste y en el Atlántico norte las medidas de longitud total de machos están en un
rango de 173 – 215 cm para hembras, y 193 – 234 para machos. El peso máximo
registrado es de 119 kilogramos, en un ejemplar macho de 257-centímetros
reportado en la bahía de Panamá (Perrin y Hohn 1994). En el Atlántico tropical
oriental y occidental la forma costera de S. attenuata es reemplazada
ecológicamente por Stenella frontalis.
El dimorfismo sexual en el cráneo es leve; el número de dientes no diferencia
en promedio a machos y hembras. La variación geográfica en el tamaño del cráneo
es pronunciada (Perrin y Hohn 1994). Los cráneos de delfines manchados costero
del Pacífico tropical oriental son los más grandes, los de delfines manchados
oceánicos son los más pequeños, mientras que los cráneos del Pacífico occidental,
El Océano Índico, y Atlántico son intermedios.
De acuerdo a Perrin et al. (1994), las variables oceanográficas: profundidad
del agua, exposición solar, temperatura de la superficie del mar, salinidad superficial,
y profundidad de la termoclina inciden en la similitud de los cráneos de delfines
oceánicos de áreas con mayor porción de batimetría oceánica del Pacífico oriental,
con aquellos que se distribuyen al sur del ecuador.
GD001, Individuo con aleta lacerada presente en todos los seguimientos de grupo en Golfo Dulce

104
La longitud de madures sexual en los adultos es a partir de 162 a 257
centímetros aproximadamente. Las hembras alcanzan madurez sexual a los 9-11
años, dependiendo de la población, y los machos 3 años más tarde (Kasuya 1976;
Kasuya et. al. 1974). La gestación es 11,2 - 11,5 meses. La tasa de ovulación para
las hembras adulta es 0,4 - 0,6 ovulaciones por año en el Pacífico oriental. La tasa
anual del embarazo es 0,3 variante de acuerdo con la población. El intervalo entre
crías es de 2 - 4 años. La duración promedio de la lactancia es 1 - 2 años, siendo
más corto en el Pacífico tropical oriental que en el Pacífico occidental. La
reproducción es estacional, con 2 - 3 estaciones definidas en el Pacífico tropical
oriental (Hohn et. al. 1985; Perrin et. al. 1976).
El delfín manchado costero. S a. graffmani en el Pacífico tropical oriental
ocurre en el agua llanas a menos de 200 kilómetros de la costa. El delfín manchado
oceánico S. a. attenuata frecuenta las aguas caracterizadas por condiciones
inusuales de capas mezcladas llanas, presencia de bajos y termoclina aguda, y baja
variación relativa anual en la temperatura superficial (Au y Perryman 1985). En este
tipo de hábitat S. attenuata se alimenta de presas que incluyen principalmente
pequeños epipelágicos y algunos organismos mesopelágicos. Cambios estacionales
ocurren en el hábitat preferido de los delfines manchados pantropicales así como
Grupo de delfines manchados en descanso, durante los seguimientos grupales.

105
variaciones interanuales en la distribución en dichos hábitat (Fiedler y Reilly 1994;
Reilly 1990). En otros océanos S. attenuata es sobre todo un delfín de hábitos
oceánicos, aunque puede ocurrir en aguas profundas cercanas a la costa, como en
las Filipinas.

106
Anexo 2. Clave de Muestreo Conductual (García y Dawson 2003, desarrollada por Schneider 1999).
1) El grupo se encuentra disperso con una distancia entre individuos de más de 10 delfines (en longitud).........................................................................
2)
El grupo esta compacto con una distancia entre individuos de menos de 5 delfines (en longitud)...................................................................................
3)
2) Se observa peces o aves asociadas.............................................................. Alimentación individual No se observan peces o aves asociadas....................................................... Merodeo 3) Se observa peces o aves asociadas.............................................................. 4) No se observan peces o aves asociadas....................................................... 5) 4)
Nado en círculos o frontal hacia cada delfín...............................................
Alimentación cooperativa
Ninguna de las observaciones anteriores.....................................................
Alimentación Grupal
5) Dirección de movimientos constantes por un periodo de 2 minutos........... 6) Dirección de movimientos constantes por un periodo de menos de 2
minutos o sin patrón constante..................................................................... 7)
6) Saltos frontales continuos............................................................................ Movilización rápida No hay saltos frontales continuos................................................................ Movilización 7)
Contacto físico, opcionalmente conductas aéreas como atisbar, nado lateral o con vientre arriba.........................................................................................
Socialización
Contacto físico moderado, el grupo se mantiene estático o a menor velocidad que la mínima del motor del bote…………………………………….
Descanso
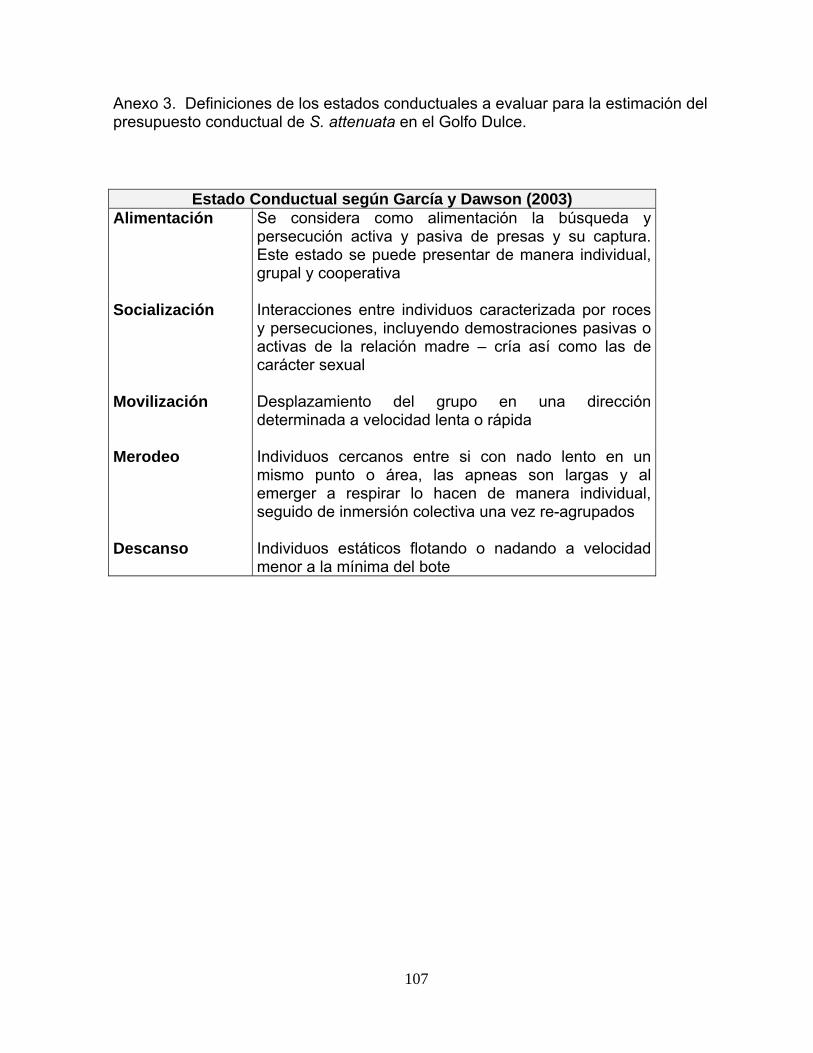
107
Anexo 3. Definiciones de los estados conductuales a evaluar para la estimación del presupuesto conductual de S. attenuata en el Golfo Dulce.
Estado Conductual según García y Dawson (2003) Alimentación Se considera como alimentación la búsqueda y
persecución activa y pasiva de presas y su captura. Este estado se puede presentar de manera individual, grupal y cooperativa
Socialización Interacciones entre individuos caracterizada por roces y persecuciones, incluyendo demostraciones pasivas o activas de la relación madre – cría así como las de carácter sexual
Movilización Desplazamiento del grupo en una dirección determinada a velocidad lenta o rápida
Merodeo Individuos cercanos entre si con nado lento en un mismo punto o área, las apneas son largas y al emerger a respirar lo hacen de manera individual, seguido de inmersión colectiva una vez re-agrupados
Descanso Individuos estáticos flotando o nadando a velocidad menor a la mínima del bote
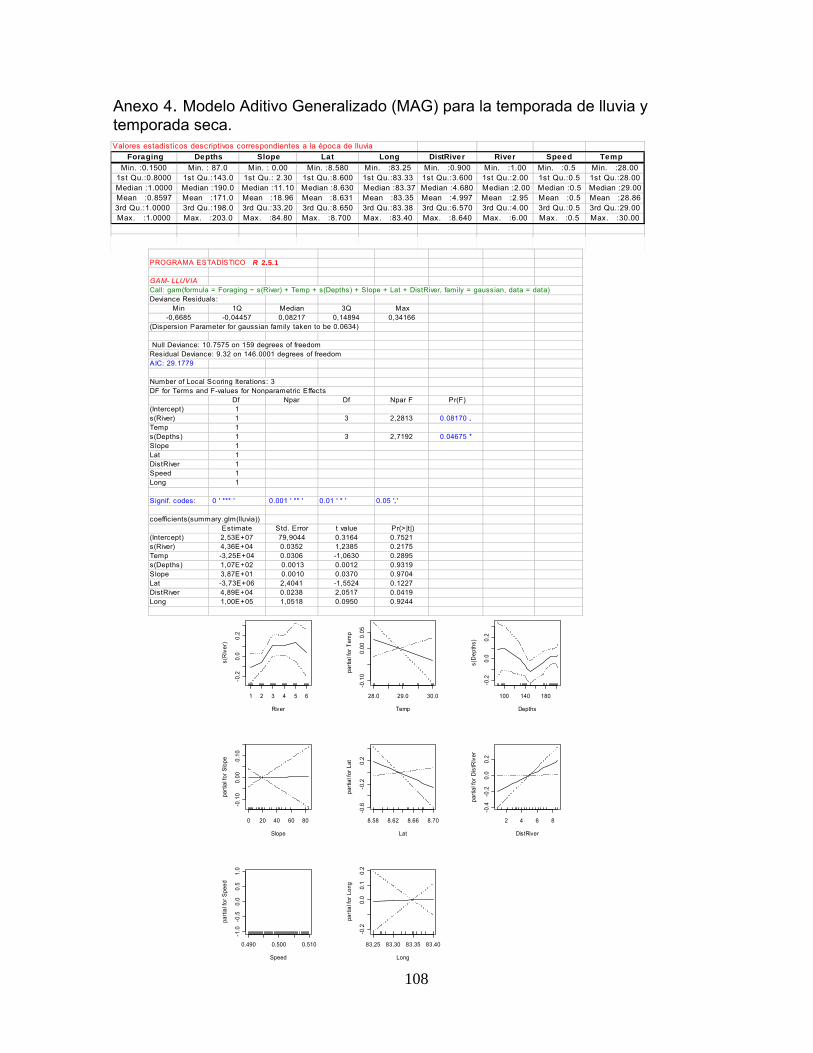
108
Anexo 4. Modelo Aditivo Generalizado (MAG) para la temporada de lluvia y temporada seca.
Valores estadisticos descriptivos correspondientes a la época de lluvia Foraging Depths Slope Lat Long DistRiver River Speed Temp
Min. :0.1500 Min. : 87.0 Min. : 0.00 Min. :8.580 Min. :83.25 Min. :0.900 Min. :1.00 Min. :0.5 Min. :28.001st Qu.:0.8000 1st Qu.:143.0 1st Qu.: 2.30 1st Qu.:8.600 1st Qu.:83.33 1st Qu.:3.600 1st Qu.:2.00 1st Qu.:0.5 1st Qu.:28.00Median :1.0000 Median :190.0 Median :11.10 Median :8.630 Median :83.37 Median :4.680 Median :2.00 Median :0.5 Median :29.00Mean :0.8597 Mean :171.0 Mean :18.96 Mean :8.631 Mean :83.35 Mean :4.997 Mean :2.95 Mean :0.5 Mean :28.863rd Qu.:1.0000 3rd Qu.:198.0 3rd Qu.:33.20 3rd Qu.:8.650 3rd Qu.:83.38 3rd Qu.:6.570 3rd Qu.:4.00 3rd Qu.:0.5 3rd Qu.:29.00Max. :1.0000 Max. :203.0 Max. :84.80 Max. :8.700 Max. :83.40 Max. :8.640 Max. :6.00 Max. :0.5 Max. :30.00
PROGRAMA ESTADÍSTICO R 2.5.1
GAM- LLUVIACall: gam(formula = Foraging ~ s(River) + Temp + s(Depths) + Slope + Lat + DistRiver, family = gaussian, data = data)Deviance Residuals:
Min 1Q Median 3Q Max-0,6685 -0,04457 0,08217 0,14894 0,34166
(Dispersion Parameter for gaussian family taken to be 0.0634)
Null Deviance: 10.7575 on 159 degrees of freedomResidual Deviance: 9.32 on 146.0001 degrees of freedomAIC: 29.1779
Number of Local Scoring Iterations: 3DF for Terms and F-values for Nonparametric Effects
Df Npar Df Npar F Pr(F)(Intercept) 1s(River) 1 3 2,2813 0.08170 .Temp 1s(Depths) 1 3 2,7192 0.04675 *Slope 1Lat 1DistRiver 1Speed 1Long 1
Signif. codes: 0 ' *** ' 0.001 ' ** ' 0.01 ' * ' 0.05 '. '
coefficients(summary.glm(lluvia))Estimate Std. Error t value Pr(>|t|)
(Intercept) 2,53E+07 79,9044 0.3164 0.7521s(River) 4,36E+04 0.0352 1,2385 0.2175Temp -3,25E+04 0.0306 -1,0630 0.2895s(Depths) 1,07E+02 0.0013 0.0012 0.9319Slope 3,87E+01 0.0010 0.0370 0.9704Lat -3,73E+06 2,4041 -1,5524 0.1227DistRiver 4,89E+04 0.0238 2,0517 0.0419Long 1,00E+05 1,0518 0.0950 0.9244
1 2 3 4 5 6
-0.2
0.0
0.2
River
s(R
iver
)
28.0 29.0 30.0
-0.1
00.
000.
05
Temp
parti
al fo
r Tem
p
100 140 180
-0.2
0.0
0.2
Depths
s(D
epth
s)
0 20 40 60 80
-0.1
00.
000.
10
Slope
parti
al fo
r Slo
pe
8.58 8.62 8.66 8.70
-0.6
-0.2
0.2
Lat
parti
al fo
r Lat
2 4 6 8
-0.4
-0.2
0.0
0.2
DistRiver
parti
al fo
r Dis
tRiv
er
0.490 0.500 0.510
-1.0
-0.5
0.0
0.5
1.0
Speed
parti
al fo
r Spe
ed
83.25 83.30 83.35 83.40
-0.2
0.0
0.1
0.2
Long
parti
al fo
r Lon
g
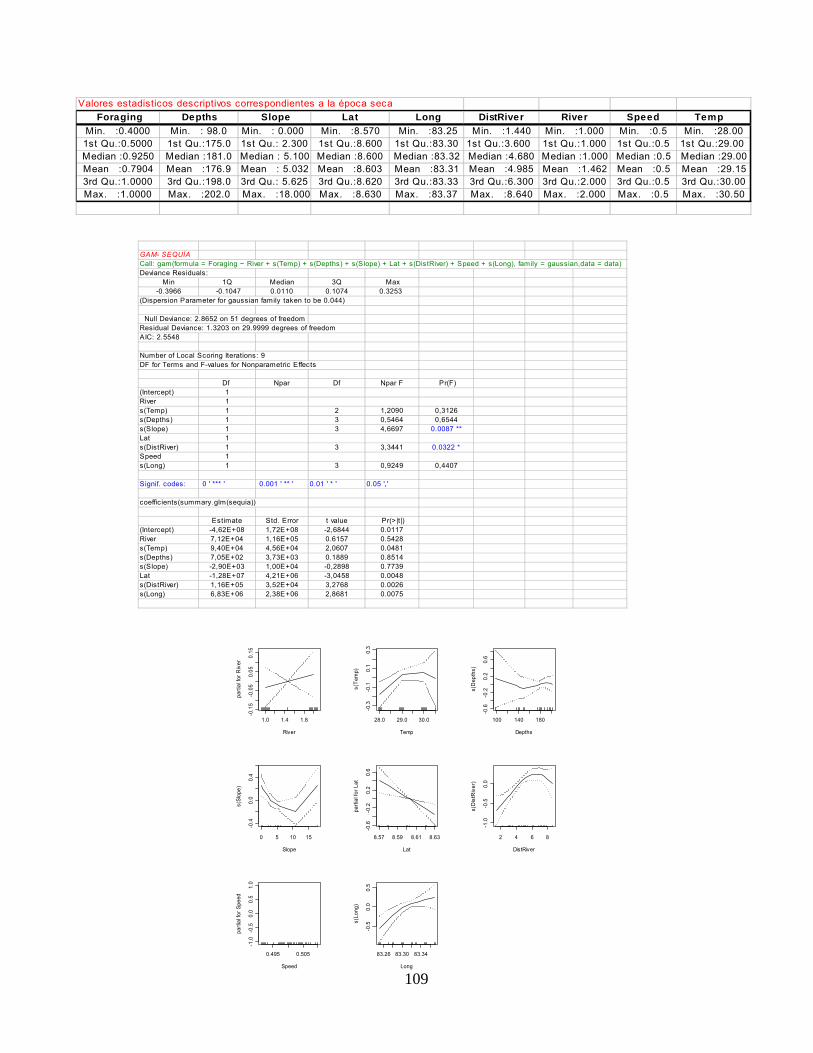
109
Valores estadisticos descriptivos correspondientes a la época seca Foraging Depths Slope Lat Long DistRiver River Speed Temp
Min. :0.4000 Min. : 98.0 Min. : 0.000 Min. :8.570 Min. :83.25 Min. :1.440 Min. :1.000 Min. :0.5 Min. :28.001st Qu.:0.5000 1st Qu.:175.0 1st Qu.: 2.300 1st Qu.:8.600 1st Qu.:83.30 1st Qu.:3.600 1st Qu.:1.000 1st Qu.:0.5 1st Qu.:29.00 Median :0.9250 Median :181.0 Median : 5.100 Median :8.600 Median :83.32 Median :4.680 Median :1.000 Median :0.5 Median :29.00Mean :0.7904 Mean :176.9 Mean : 5.032 Mean :8.603 Mean :83.31 Mean :4.985 Mean :1.462 Mean :0.5 Mean :29.153rd Qu.:1.0000 3rd Qu.:198.0 3rd Qu.: 5.625 3rd Qu.:8.620 3rd Qu.:83.33 3rd Qu.:6.300 3rd Qu.:2.000 3rd Qu.:0.5 3rd Qu.:30.00Max. :1.0000 Max. :202.0 Max. :18.000 Max. :8.630 Max. :83.37 Max. :8.640 Max. :2.000 Max. :0.5 Max. :30.50
GAM- SEQUÍACall: gam(formula = Foraging ~ River + s(Temp) + s(Depths) + s(Slope) + Lat + s(DistRiver) + Speed + s(Long), family = gaussian,data = data)Deviance Residuals:
Min 1Q Median 3Q Max-0.3966 -0.1047 0.0110 0.1074 0.3253
(Dispersion Parameter for gaussian family taken to be 0.044)
Null Deviance: 2.8652 on 51 degrees of freedomResidual Deviance: 1.3203 on 29.9999 degrees of freedomAIC: 2.5548
Number of Local Scoring Iterations: 9 DF for Terms and F-values for Nonparametric Effects
Df Npar Df Npar F Pr(F)(Intercept) 1River 1s(Temp) 1 2 1,2090 0,3126s(Depths) 1 3 0,5464 0,6544s(Slope) 1 3 4,6697 0.0087 **Lat 1s(DistRiver) 1 3 3,3441 0.0322 *Speed 1s(Long) 1 3 0,9249 0,4407
Signif. codes: 0 ' *** ' 0.001 ' ** ' 0.01 ' * ' 0.05 '.'
coefficients(summary.glm(sequia))
Estimate Std. Error t value Pr(>|t|)(Intercept) -4,62E+08 1,72E+08 -2,6844 0.0117River 7,12E+04 1,16E+05 0.6157 0.5428s(Temp) 9,40E+04 4,56E+04 2,0607 0.0481s(Depths) 7,05E+02 3,73E+03 0.1889 0.8514s(Slope) -2,90E+03 1,00E+04 -0,2898 0.7739Lat -1,28E+07 4,21E+06 -3,0458 0.0048s(DistRiver) 1,16E+05 3,52E+04 3,2768 0.0026s(Long) 6,83E+06 2,38E+06 2,8681 0.0075
1.0 1.4 1.8
-0.1
5-0
.05
0.05
0.15
River
parti
al fo
r Riv
er
28.0 29.0 30.0
-0.3
-0.1
0.1
0.3
Temp
s(Te
mp)
100 140 180
-0.6
-0.2
0.2
0.6
Depths
s(D
epth
s)
0 5 10 15
-0.4
0.0
0.4
Slope
s(Sl
ope)
8.57 8.59 8.61 8.63
-0.6
-0.2
0.2
0.6
Lat
parti
al fo
r Lat
2 4 6 8
-1.0
-0.5
0.0
DistRiver
s(D
istR
iver
)
0.495 0.505
-1.0
-0.5
0.0
0.5
1.0
Speed
parti
al fo
r Spe
ed
83.26 83.30 83.34
-0.5
0.0
0.5
Long
s(Lo
ng)
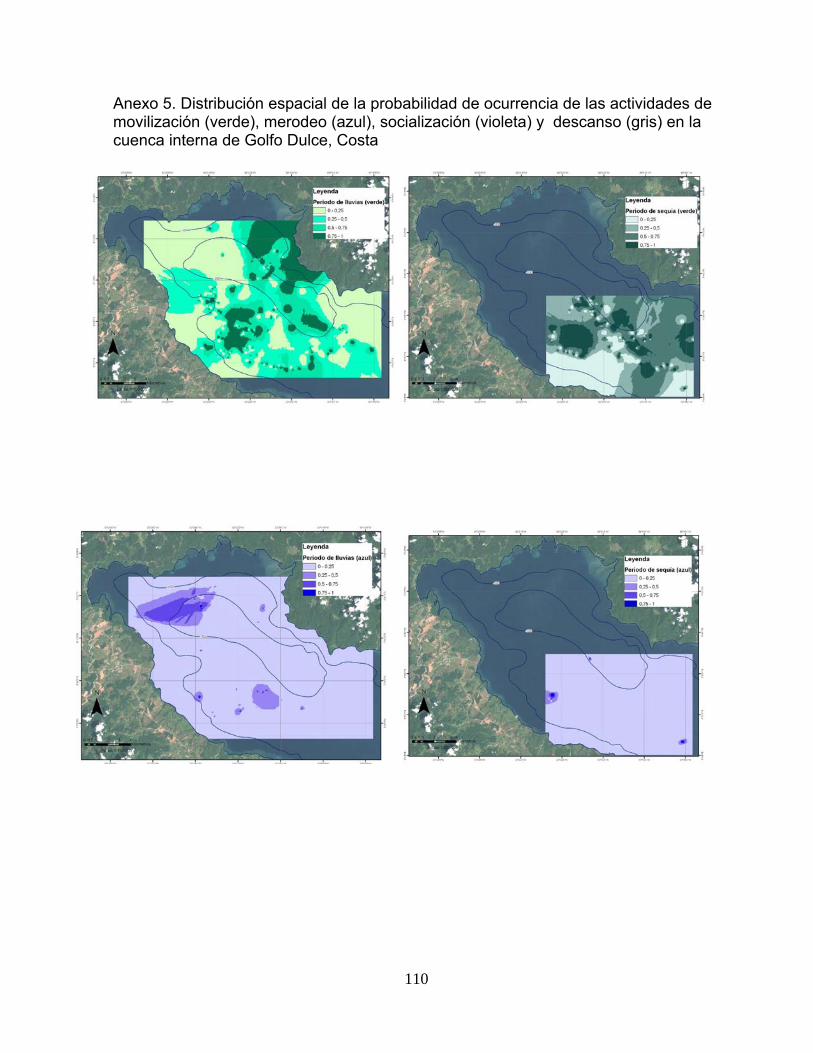
110
Anexo 5. Distribución espacial de la probabilidad de ocurrencia de las actividades de movilización (verde), merodeo (azul), socialización (violeta) y descanso (gris) en la cuenca interna de Golfo Dulce, Costa
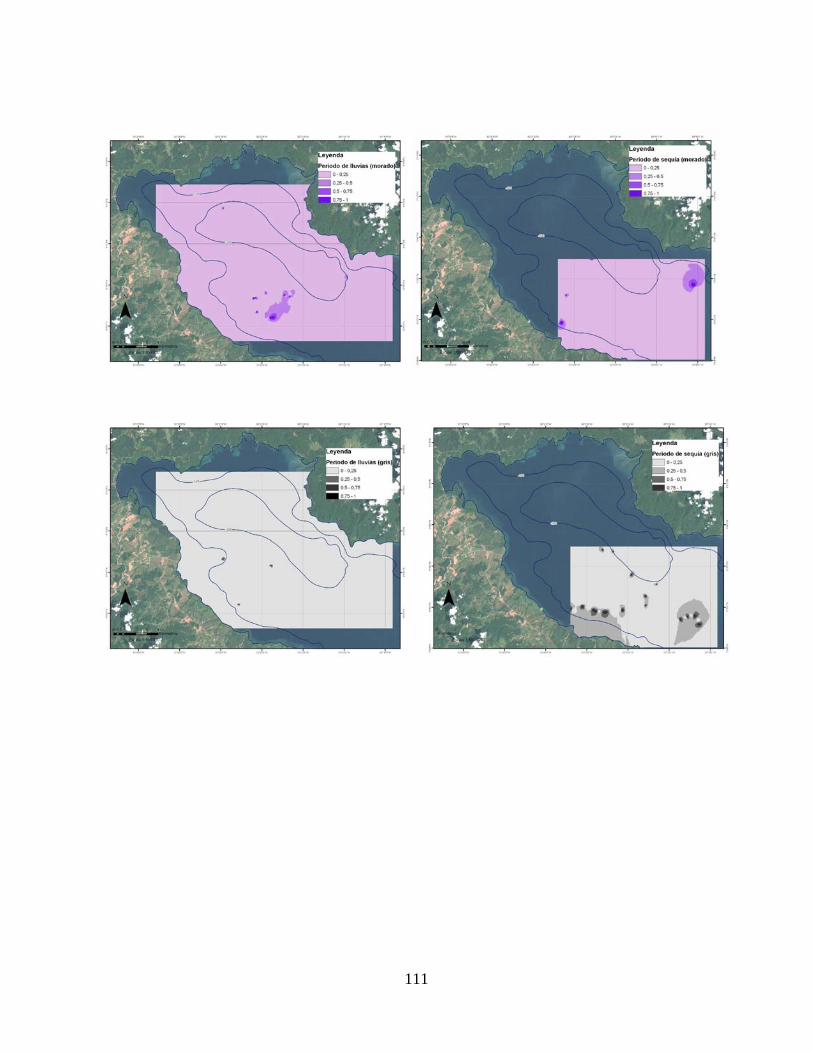
111
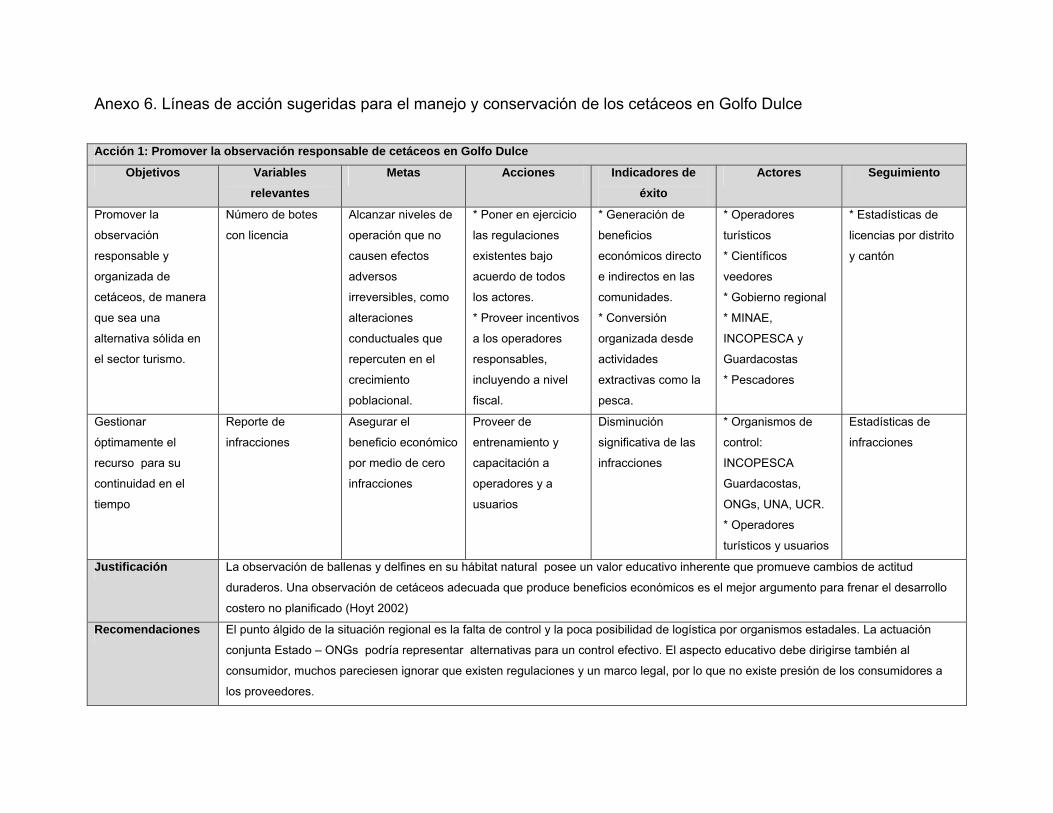
Anexo 6. Líneas de acción sugeridas para el manejo y conservación de los cetáceos en Golfo Dulce Acción 1: Promover la observación responsable de cetáceos en Golfo Dulce
Objetivos Variables relevantes
Metas Acciones Indicadores de éxito
Actores Seguimiento
Promover la
observación
responsable y
organizada de
cetáceos, de manera
que sea una
alternativa sólida en
el sector turismo.
Número de botes
con licencia
Alcanzar niveles de
operación que no
causen efectos
adversos
irreversibles, como
alteraciones
conductuales que
repercuten en el
crecimiento
poblacional.
* Poner en ejercicio
las regulaciones
existentes bajo
acuerdo de todos
los actores.
* Proveer incentivos
a los operadores
responsables,
incluyendo a nivel
fiscal.
* Generación de
beneficios
económicos directo
e indirectos en las
comunidades.
* Conversión
organizada desde
actividades
extractivas como la
pesca.
* Operadores
turísticos
* Científicos
veedores
* Gobierno regional
* MINAE,
INCOPESCA y
Guardacostas
* Pescadores
* Estadísticas de
licencias por distrito
y cantón
Gestionar
óptimamente el
recurso para su
continuidad en el
tiempo
Reporte de
infracciones
Asegurar el
beneficio económico
por medio de cero
infracciones
Proveer de
entrenamiento y
capacitación a
operadores y a
usuarios
Disminución
significativa de las
infracciones
* Organismos de
control:
INCOPESCA
Guardacostas,
ONGs, UNA, UCR.
* Operadores
turísticos y usuarios
Estadísticas de
infracciones
Justificación La observación de ballenas y delfines en su hábitat natural posee un valor educativo inherente que promueve cambios de actitud
duraderos. Una observación de cetáceos adecuada que produce beneficios económicos es el mejor argumento para frenar el desarrollo
costero no planificado (Hoyt 2002)
Recomendaciones El punto álgido de la situación regional es la falta de control y la poca posibilidad de logística por organismos estadales. La actuación
conjunta Estado – ONGs podría representar alternativas para un control efectivo. El aspecto educativo debe dirigirse también al
consumidor, muchos pareciesen ignorar que existen regulaciones y un marco legal, por lo que no existe presión de los consumidores a
los proveedores.
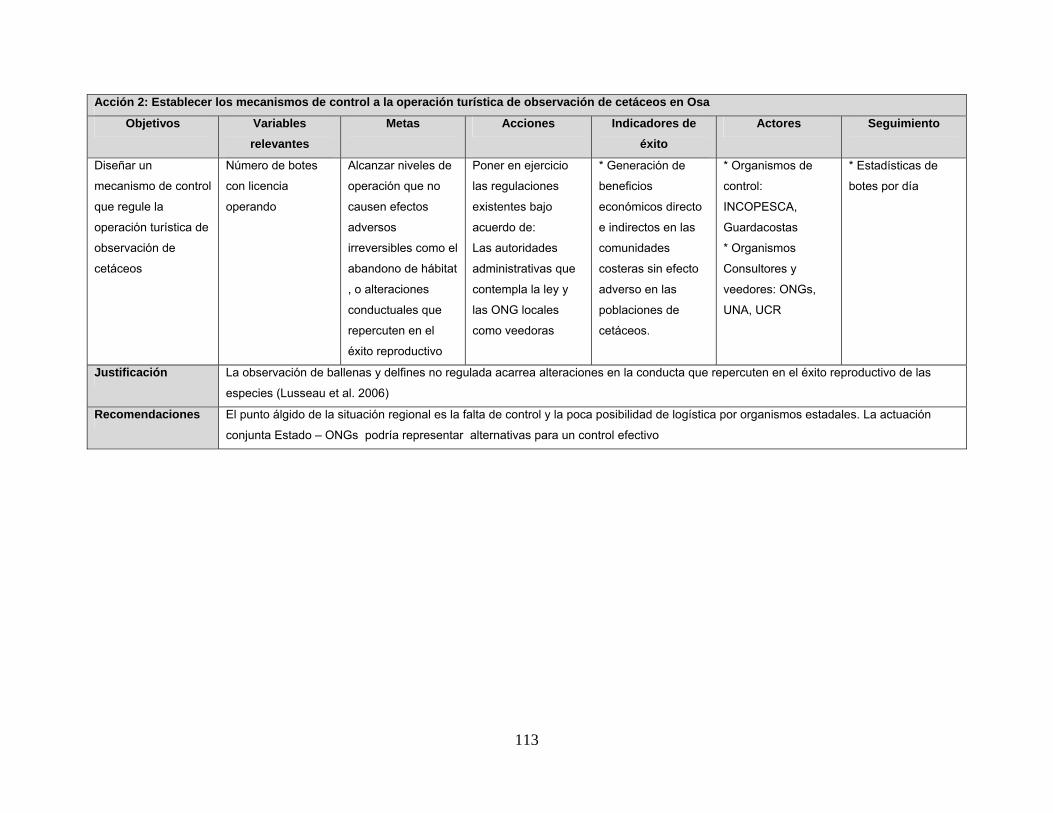
113
Acción 2: Establecer los mecanismos de control a la operación turística de observación de cetáceos en Osa
Objetivos Variables relevantes
Metas Acciones Indicadores de éxito
Actores Seguimiento
Diseñar un
mecanismo de control
que regule la
operación turística de
observación de
cetáceos
Número de botes
con licencia
operando
Alcanzar niveles de
operación que no
causen efectos
adversos
irreversibles como el
abandono de hábitat
, o alteraciones
conductuales que
repercuten en el
éxito reproductivo
Poner en ejercicio
las regulaciones
existentes bajo
acuerdo de:
Las autoridades
administrativas que
contempla la ley y
las ONG locales
como veedoras
* Generación de
beneficios
económicos directo
e indirectos en las
comunidades
costeras sin efecto
adverso en las
poblaciones de
cetáceos.
* Organismos de
control:
INCOPESCA,
Guardacostas
* Organismos
Consultores y
veedores: ONGs,
UNA, UCR
* Estadísticas de
botes por día
Justificación La observación de ballenas y delfines no regulada acarrea alteraciones en la conducta que repercuten en el éxito reproductivo de las
especies (Lusseau et al. 2006)
Recomendaciones El punto álgido de la situación regional es la falta de control y la poca posibilidad de logística por organismos estadales. La actuación
conjunta Estado – ONGs podría representar alternativas para un control efectivo
114
Acción 3: Establecer una red de Áreas Marinas Protegidas (AMPs) en Golfo Dulce
Objetivos Variables relevantes
Metas Acciones Indicadores de éxito
Actores Seguimiento
Proteger los hábitat
críticos para las
especies de cetáceos
involucradas
Áreas de
alimentación
Extensión de limites
de protección
marinos existentes
Crear un área de
protección integral
que garantice la
continuidad del éxito
reproductivo de los
cetáceos presentes
en aguas de Golfo
Dulce
Zonificar el área de
importancia en
conservación de
acuerdo al hábitat
crítico de las
especies presentes.
Asegurar que los
límites de las AMPs
a promoverse,
cubran los hábitat
claves zonificados
Establecer consulta
publica en las
comunidades
locales
* Incremento o
equilibrio en la taza
de natalidad de
cetáceos dentro de
los limites marinos
de Golfo Dulce
* Generación de
beneficios
económicos directo
e indirectos en las
comunidades
costeras por uso
racional de las
áreas de protección
* Pescadores y
comunidades
costeras
* Científicos
veedores y
consultores afiliados
a ONGs y academia
* Gobierno regional
* MINAE,
INCOPESCA y
Guardacostas
* Sector Turismo
* Índices
demográficos de las
especies presentes
* Índices que refleje
el efecto de
protección sombrilla
Justificación Golfo Dulce como ecosistema marino sostiene las poblaciones de dos especies simpátricas de delfines (T. truncatus + S. attenuata). La
cuenca interna es un hábitat crítico de alimentación particularmente las áreas de influencia de los rios
Recomendaciones Incrementar la línea base sobre la dinámica ecológica del Golfo, particularmente la influencia e importancia de los ríos no ha sido
suficientemente estudiada










