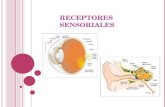
Receptores
14
Receptores Dr. Fornaguera Hoy vamos a hablar de una palabra clave: la comunicación; pero para que se comunica la célula? Para integrarse como un todo, para enterarse de que es lo que esta pasando en todo lado y en todo momento, todo esto es lo que permite que respondamos al ambiente. Hay varios tipos de comunicación entre las células: la paracrina, la autocrina y la endocrina. 1. Paracrinas: se da entre 2 células muy cercanas entre sí. Es muy rápido, no necesita pasar al torrente sanguíneo. 2. Autocrinas: se comunica consigo misma. Existen las internas, que es cuando no salen sustancias de sí misma, y las externas, que es cuando debe de liberar las sustancias para poder reconocerlas. 3. Endocrina: fue el primer tipo conocido, se descubrió con hormonas, porque las hormonas van a componer lo que se conoce como un sistema endocrino en el sistema de comunicación interna. Necesita del torrente sanguíneo y se da a largas distancias. Requiere de mayor tiempo de acción. “El concepto de receptores es importante porque cuando se libera la sustancia para que se den estos tipos de comunicación no se van a afectar a todas las células sino solo a aquellas que tengan el receptor necesario” Hay dos grandes familias de receptores que son: -Receptores intracelulares o nucleares: son receptores que están en el citoplasma o adentro del núcleo pero la acción siempre va a ser dentro del núcleo. -Receptores de membrana: Asociados a membranas celulares o las de las organelas, es decir algunos también están dentro de la célula, pero no son intracelulares o nucleares porque están asociados a una membrana siendo entonces receptores de membrana. Todos los receptores de membrana son proteínas integrales y todos ellos son transmembrana.
-
Upload
api-3696656 -
Category
Documents
-
view
3.634 -
download
0
Transcript of Receptores

Receptores Dr. Fornaguera
Hoy vamos a hablar de una palabra clave: la comunicación; pero para que se comunica la célula? Para integrarse como un todo, para enterarse de que es lo que esta pasando en todo lado y en todo momento, todo esto es lo que permite que respondamos al ambiente. Hay varios tipos de comunicación entre las células: la paracrina, la autocrina y la endocrina.
1. Paracrinas: se da entre 2 células muy cercanas entre sí. Es muy rápido, no necesita pasar al torrente sanguíneo.
2. Autocrinas: se comunica consigo misma. Existen las internas, que es cuando no salen sustancias de sí misma, y las externas, que es cuando debe de liberar las sustancias para poder reconocerlas.
3. Endocrina: fue el primer tipo conocido, se descubrió con hormonas, porque las hormonas van a componer lo que se conoce como un sistema endocrino en el sistema de comunicación interna. Necesita del torrente sanguíneo y se da a largas distancias. Requiere de mayor tiempo de acción.
“El concepto de receptores es importante porque cuando se libera la sustancia para que se den estos tipos de comunicación no se van a afectar a todas las células sino solo a aquellas que tengan el receptor
necesario”
Hay dos grandes familias de receptores que son: -Receptores intracelulares o nucleares: son receptores que están en el citoplasma o adentro del núcleo pero la acción siempre va a ser dentro del núcleo.-Receptores de membrana: Asociados a membranas celulares o las de las organelas, es decir algunos también están dentro de la célula, pero no son intracelulares o nucleares porque están asociados a una membrana siendo entonces receptores de membrana. Todos los receptores de membrana son proteínas integrales y todos ellos son transmembrana.
El primer receptor de membrana fue el receptor nicotínico de acetilcolina y se llamo así porque es muy parecido al receptor de nicotina, ya que los receptores son llamados de acuerdo al ligando con el que fueron descubiertos. Este receptor nicotínico de acetilcolina pertenece a la familia de cuatro dominios transmembrana, los receptores de esta familia se caracterizan porque son canales iónicos.
“Todos los receptores sin importar si son de membrana o intracelulares cumplen dos características: son específicos y tienen que ser bastante afines a su ligando”
-Especificidad: un receptor es selectivo para un determinado tipo de ligando, pero la especificidad no es del 100%, sino que también puede interactuar con sustancias parecidas, por lo que no hay exclusividad.
-Afinidad: la concentración necesaria de esa sustancia para llegar al tope de unión, siendo la constante de afinidad, entre mayor sea la constante de afinidad menos afín será por esa sustancia.

-Mecanismos de señalización: todos los receptores están asociados de una a otra manera a mecanismos de señalización que llamaremos de transducción, es decir cuando una sustancia se une al receptor sucede algo, estos son los segundos mensajeros ( sustancia que se genera por la estimulación del receptor por un primer mensajero)
“Todos los segundos mensajeros son intracelulares”
-Redundancia: un mismo ligando se puede unir a varios recetores y producir la cascada de amplificación, redundancia porque sobre los receptores actúa un mismo ligando, siendo redundante el mismo ligando. O también puede ser redundante que varios receptores generen la misma respuesta ( redundante la misma respuesta) -Diversidad: un ligando puede actuar sobre varios receptores y producir la cascada de amplificación, aquí se aplica el concepto de diversidad porque el ligando puede actuar sobre varios receptores, es decir tiene diversidad para escoger sobre que receptor va a actuar.
“Todos los ligandos requieren de tiempo para poder generar una respuesta en el cuerpo”
Receptores de membrana
-Todos los receptores de membrana son proteínas integrales y tienen dominios transmembrana y citosolicos-Hay tres familias de estos receptores de membrana según la cantidad de dominios transmembrana:
1 dominio transmembrana: El receptor tiene capacidad enzimática, pero la acción enzimatica no es sobre su ligando porque el ligando se une extracelularmente mientras la función enzimatica es intracelular.
4 dominios transmembrana: Son canales iónicos.
7 dominios transmembrana: Receptores serpentina, se asocian siempre a proteínas G (dependen de nucleótidos de guanina).
“Hay ligandos que actúan sobre dos tipos de receptores, por ejemplo el glutamato que actúan sobre
receptores tipo IV y tipo VII”
Todos los receptores tienen lo que se llama cascada de amplificación, un numero pequeño de ligandos son capaces de producir grandes respuestas.

Receptores Tipo Canal
-Tienen cuatro dominios transmembrana-Fueron los primeros en descubrirse-Son receptores ionotropicos-Son canales iónicos, para iones como Cl-, Na+, Ca+2, K+ …
1. Canal nicotínico de acetilcolina: esta en mayor cantidad en el músculo esquelético tiene 5 subunidades, cada una con 4 dominios
transmembrana A través de este canal pasa Na+, cuando pasa la
membrana se despolariza produciendo liberación de calcio y con esto la contracción del músculo esquelético.
Fueron descubiertos en el pez torpedo Inhibidores de este canal: Tubocurina, cobroxina y
bungarotoxina. Los inhibidores lo que hacen es que bloquean el canal
2. Receptor para el glutamato: El glutamato tiene por lo menos 5 canales
ionotropicos El canal mas estudiado para el glutamato es el de N–
metil-D-aspartato (NMDA). Tiene 5 subunidades de 4 dominios transmembrana; canal para entrada del Ca. El NMDA es el único que tiene que ser coactivado ya
que necesita además de Glu a la Gly (coactivador) para que se pueda dar la entrada del Ca, pero aun así se encuentra un tapón de de Mg para esto se
encuentra asociado a otro receptor de glutamato que se llama AMPA, el cual mete Na el cual despolariza la membrana de provocando que se retire el tapón de Mg y se pueda meter el Ca.
El receptor de NMDA es un receptor muy regulado, por ejemplo es inhibido por espermita que es una amina, es estimulado por aspartato.
Es un receptor muy importante, porque esta relacionado con el proceso del aprendizaje.
3. Receptor GABAA: Receptor ácido gama amino butírico. Es GABAA porque hay un GABAB que es de 7 dominios
transmembrana Es de 5 subunidades con 4 dominios cada una. Es el principal receptor inhibitorio del sistema nervioso
disminuyendo loa velocidad de disparo de las neuronas, esto porque el cloruro entra a favor de gradiente de concentración e hiperpolariza la membrana, por lo que será muy difícil activar un impulso nervioso
Este canal es muy importante en anestésicos Puede unirse a barbitúricos, esteroides, picrotoxina y
benzodiazepinas ( todos estos favorecen la apertura del canal es decir favorecen la hiperpolarizacion de la membrana.

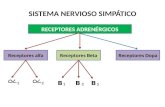
Receptores asociados a proteínas G
-7 dominios transmembrana. -Son segundos mensajeros. -receptores serpentina-son metabotropicos-Son los más abundantes en la naturaleza-están asociados con proteínas G
-Son muchos los ligandos que pueden activar proteínas G, algunos son:
Características de las Proteínas G:
-Son proteínas integrales de membrana pero no transmembrana, -se encuentran en la cara interna de la membrana, están constituidas por 3 subunidades (α,β y γ). -Son triméricas o de alto peso molecular. -Se llaman proteínas G porque su activación e inactivacion depende nucleótidos de guanina. GTP (GTP y GDP). -La subunidad α tiene actividad GTPasica, y es aquí donde se unen los nucleótidos de guanina. -Todas las proteínas G se activan cuando se les une el GTP( ocurre un cambio donde sale el GDP y llega GTP) y se desactivan cuando el GTP se cambia por GDP (aquí si ocurre una fosforilación), todo esto en la subunidad α - La subunidad α necesita de una proteína para realizar su actividad GTPasica (cambiar GTP por GDP), que son las GAP( proteínas activadoras de la GTPasica)-β y γ también pueden activar, la primera vez que se descubrió fue en el corazón donde activaban canales de potasio

Tipos de proteínas G:
- Gs: estimula la enzima adenilato ciclasa, adenilciclasa o adenilil ciclasa que actúa en adenosín monofosfato cíclico; va actúa en el sustrato ATP para producir AMPc.
- Gi: inhibe a la adenilato ciclasa. - Gq: activa la fosfolipasa C.
Nota: las proteínas G12, no es una proteína especifica sino que es un conjunto de proteínas a las cuales no se les ha definido bien.
Acción de la Adenilato Ciclasa
-Por cada ATP que entre a la adenilato ciclasa sale un AMPc-Lo que pasa es que se rompe el pirofosfato y se forma ciclo formado por un solo fosfato establecido por un enlace fosfodiester entre el carbono 3 y el carbono 5 del azúcar
Esto que tienen aquí es una adenilato ciclasa típica, hay mas de un tipo, pero todas ellas tienen 12 dominios transmembrana y eso que se les llama C1y C2 son los dominios citoplasmáticos de acción. Tenemos la posibilidad de regular estas enzimas, el C2 es el sitio de mayor regulación, este está muy cerca del carboxilo Terminal. Tenemos hoy en día por lo menos 6 tipos de adenilato ciclasa; en la adenilato ciclasa II y IV, β y γ son coactivadores de la enzima, es decir, además de α, β y γ pueden favorecer a la acción de la adenilato ciclasa, mientras que la clase I, β y γ son inhibidores de la enzima; el tipo III no es afectada por β y γ. La calmodulina también afecta ala adenilato ciclasa tipo I mientras que a la tipo II no la afecta.
Protein Kinasa A
-Tiene 4 subunidades, dos catalíticas y dos reguladoras- Una sola subunidad catalítica puede fosforilar más de una proteína y no solo a más de una proteína sino a más de una proteína diferente-Pertenece a la familia de las serin-treonin kinasa, esto quiere decir que solo fosforilan en residuos de serina y treonina -Se activa con 4 AMPc, dos en cada subunidad reguladora, esto porque cuando llega el AMPc a las subunidades reguladoras estas se separan de las catalíticas dejándolas libres para que realicen su función.

Un mismo ligando por ejemplo la epinefrina actúa a través del sistema de AMPc, en dos diferentes tejidos, en el tejido adiposo y en el hígado por ejemplo, el ligando es el mismo, pero el tejido es diferente por lo que se pueden obtener diferentes respuestas, o sea, actuando juntas la protein kinasa A y el AMPc producen efectos distintos en diferentes tejidos.
Estas son algunas de las enzimas que son activadas por la protein kinasa A, estos son pequeño ejemplos de las acciones que pueden tener las protein kinasas A, dependiendo del tejido en el que estén.
Mecanismos para detener el sistema del AMPc
1. Inhibiendo a las proteínas G: esto se logra activando la vía inhibitorio a través de las proteínas Gi
2. Activar la actividad GTPasica: se favorece la acción de las GAP, para pasar el GTP a GDP
3. Por medio de la fosfodiesterasa: se rompen los enlaces fosfodiester, rompiendo entonces el AMPc y pasándolo a 5’ AMP. Un efecto importante asociado con el mecanismo de las fosfodiesterasa es el de los estimulantes, que se llaman las metilxantinas, estas lo que hacen es meter grupos metilo, dentro de estas metilxantinas hay tres sustancias muy conocidas que son: la cafeína, teobromina y la teofilina. La cafeína se encuentra sobre todo en café, la teobromina se encuentra en el chocolate, fundamentalmente en el cacao y la teofilina en el te y también en el café. Todos estos tienen un efecto

que se llama estimulante que empieza por la inhibición de la fosfodiesterasa, lo cual produce que el AMPc se mantenga elevado, esto provoca que la proteín Kinasa A fosforile todas las proteínas que ella fosforila, en el sistema reticular activante es donde se mantiene el proceso de alerta del organismo debido a este mecanismo.
4. Por fosfatasas: para desfosforilar lo que las subunidades catalíticas de la protein kinasa A fosforilaron, al ser activadas por la unión del AMPc a sus subunidades reguladoras. Las fosforilaciones se darán fundamentalmente en tres clases de proteínas: tirosina, serina o treonina. Existen en varias clases de fosfatasas:
Serin / treoninfosfatasa : desfosforilan residuos de serina o de treonina. Esta familia de fosfatasas son
las más abundantes y las más estudiadas. Hay varios tipos: PP-1, PP-2A, PP-2B y PP-2C, el nombre
depende de la estructura y de la acción. Tirosinfosfatasas : desfosforilan proteínas que fueron fosforiladas en tirosina, en otras palabras
desfosforilan residuos de tirosina.
5. internalizando a los receptores: este mecanismo es un poco mas complicado sobre todo porque requiere de otro proceso de fosforilación, este proceso ocurre cuando hay demasiados ligandos y la subunidad ha permanecido mucho tiempo activa, cuando esto ocurre actúa, se establece una señal para que actúa una protein kinasa que no es la protein kinasa A, sino que es la kinasa asociada a receptores asociados a proteínas G (GRK), cuando estas kinasas son activadas, y vean que interesante las que las activan son β y γ, es decir que estas Kinasas solo van a ser activadas cuando halla mucha activación de proteínas G. Cuando la GRK esta activa se fosforila el receptor en sus residuos de treonina o serina, y al ser fosforilados ya no los pueden reconocer las subunidades de las proteínas G no pudiendo ser activadas aunque halla mucho ligando.Estas fosforilaciones además de no permitir la actividad de las proteínas G, estas fosforilaciones son reconocidas por las Arrestinas, que son proteínas que reconocen esos lugares del receptor y lo jalan hacia la porción intracelular, lo quitan de la membrana; entonces no importa que halla mucho ligando ya que los receptores no están afuera y no se da la acción. Estos receptores internalizados pueden ser en algún momento reciclados.
Ejemplo:
La protein kinasa A es activada por el AMPc y resulta que esta protein kinasa A fosforilo a I-PIP(inhibidor de la protein fosfatasa I) activándolo por lo que la protein fosfatasa I estará inactiva. ¿Como estará la actividad de la protein fosfatasa I si los niveles de AMPc son bajos?
R/ La protein Fosfatasa I estará activa porque los bajos niveles de AMPc no permiten la activación de

la protein kinasa A, entonces no se dará la activación del I-PIP, el cual es el inhibidor de la PIP (protein fosfatasa I)¿Qué le hace una metilxantina a la actividad del inhibidor de la protein fosfatasa? R/ Aumenta la actividad ya que mantiene elevada la concentración de AMPc, activando a las protein kinasas A, las cuales van a ir a fosforilar a este receptor activándolo.
Hace varios años se creía que los receptores de membrana actuaban a través de mecanismos rápidos ycitoplásmicos, pero se descubrió que algunos ligandos que se unen a receptores de membrana pueden afectar procesos intranucleares igual que como actúan los receptores intranucleares solo que con receptores de membrana, ahora se sabe que los receptores de membrana también pueden producir efectos lentos y de larga duración y uno de los ejemplos clásicos es el del CREB. El CREB (unión del elemento de respuesta del AMPc) es una proteína que tiene que ver con la trascripción genética, es una proteína citoplasmática que es fosforilada por la PKA y una vez fosforilada pasa al núcleo para unirse al ADN en un lugar que se llama el CRE (elemento de respuesta del AMPc), este es el lugar especifico de unión al CREB, este lo que hace es favorecer la trascripción de ciertas proteínas.
Gracias a toxinas extracelulares se ha descubierto la posibilidad de activar o desactivar proteínas G a través de la que se conoce como ribosilacion, esto se descubrió gracias a la toxina del cólera, esta toxina del cólera lo que hace es unirse a las proteínas GS pero a las que están activas, entonces lo que hace esta toxina es producir una ribosilacion en la subunidad α de la proteína G, inhibiendo la actividad GTPasica aumentando los niveles de AMPc, y estos niveles de AMPc aumentados a nivel intestinal producen la activación de canales iónicos en el epitelio intestinal, provocando la salida de iones al lumen intestinal jalando agua produciendo as grandes diarreas características de la toxina del cólera.
Hay otra toxina que es la de la tosferina, que es la de la bacteria Bordetella pertussis, esta bacteria actúa principalmente en el epitelio respiratorio produciendo un exceso de mucus el cual es un caldo de cultivo para otras bacterias, esta toxina actúa también sobre proteínas G ribosilandolas, pero a diferencia de la toxina del cólera esta ribosila a las Gi, ribosila al trímero impidiendo la activación de la proteinaGi, esta proteína es importante para detener la producción de AMPc la acción va a ser similar que en la de la toxina del cólera pero esta vez liberando mucus
Mecanismo de transducción del Fosfatidil Inositol difosfato PIP2
Este mecanismo de transducción se asocia con receptores de siete dominios de membrana, por lo cual se encuentra asociada a proteínas G, pero no son Gs ni Gi son del tipo Gq. Su mecanismo es muy similar al del AMPc, se activa la proteína Gq, aquí no se activa la adenilato ciclasa sino otra enzima la fosfolipasa C (PLC), fosfolipasa C β específicamente, el PLC parte el PIP2 en IP3 (inositol 3 fosfato)y en DAG (diacil glicerol). El IP3 y el DAG son segundos mensajeros.

Isoforma: proteínas que tienen la misma función pero diferente estructura primaria
Estos segundos mensajeros tienen la característica de que el DAG es liposoluble por lo que se va a la membrana y el IP3 es hidrosoluble y se va hasta el retículo endoplasmatico, aquí produce la apertura de canales de Ca+ hacía el citoplasma, el IP3 produce respuesta celular pero el DAG también. El Ca2+ al salir al citoplasma activa a las proteínas kinasas C (también puede activar a las protein kinasa calcio calmodulina dependiente (PKC/CaM)), estas proteín kinasas van a fosforilar en residuos de serina y treonina (son serin/treonin Kinasas al igual que las proteín kinasas A).
Con la liberación del calcio las PKC migran hacia la membrana para activarse, en la membrana la activa la DAG, entonces el calcio favorece la migración y la DAG la activación. La activación de la protein kinasa C va a permitir que se fosforilen otras proteínas.
Protein Kinasa C-Tiene 8 isoformas-Algo importante es que la PKC (proteín kinasa c) tiene la habilidad de activar la cascada de otras kinasas que se llaman MAP-kinasas, las cuales son secuencias de kinasas que se fosforilan así mismas y forman cascadas de kinasas. La PKC fosforila a la MAP-Kinasa Kinasa Kinasa (Raf), la cual a su vez fosforila a la MAP-Kinasa Kinasa (Mek), la que a su vez fosforilará a la MAP-Kinasa (Erk). MAP significa proteínas activadoras de mitosis, tiene que ver con procesos de división celular, la ERK puede una vez que este fosforilada migrar al núcleo y al llegar ahí activar genes que tienen que ver con la división y
diferenciación celular, esto se observa en el cáncer. -Este es otro proceso en que un receptor de membrana afecta procesos intranucleares.
Protein Kinasas calcio calmodulina dependientes (PKC/CaM)
Esto que tienen aquí es la calmodulina, ustedes la conocen de la protein kinasa calcio calmodulina dependiente que se activan con la unión de cuatro Ca2+. La Protein Kinasa calcio calmodulina dependiente II es una de las mas interesantes.

Existen 12 subunidades, cuando se habla de esto es por la capacidad de combinación que tienen. La PKC/CaM II es interesante por sus procesos de regulación, es una enzima que puede ser regulado tanto por calcio como por fosforilación.
Vamos a hacer de cuenta que la primera imagen de la esquina derecha es la protein kinasa calcio calmodulina dependiente II que esta inactiva, la segunda de la derecha es la calmodulina que se activa con cuatro calcios, una vez que el calcio se une con la calmodulina se produce un cambio conformacional que va a hacer que se una a la protein kinasa, al unirse la calmodulina la PKC/CaM se activa, una vez que esta enzima esta así puede autofosforilarse (es kinasa de otras proteínas pero además de si misma) y al autofosforilarse obtiene mayor actividad que cuando únicamente tenia al calcio con la calmodulina, teniendo aquí el 100% de la actividad.Cuando el calcio se va, se suelta la calmodulina pero la PKC/CaM continua activa porque todavía esta fosforilada y tiene un 50-80% de su actividad total, se inactiva totalmente con una fosfatasa que le quita la fosforilación convirtiéndola en una enzima totalmente inactiva.
Aquí tenemos un grafico que resume los dos mecanismos de transducción que vimos hoy, el del AMPc y el del fosfatidil inositol. Tenemos una proteína, la DARPP-32, lo importante es que esta proteína fosforilada es activa, la fosforila la protein kinasa A (PKA), al estar fosforilada y por lo tanto activa esta proteína inhibe a la protein fosfatasa I (PP-I), no permitiendo que esta valla a desfosforilar.La bomba de sodio/potasio fosforilada es inactiva, entonces ¿que pasara cuando aumente el AMPc con la bomba de NA/K?
R/ La bomba estaría totalmente inactiva, porque si aumentan los niveles de AMPc estará muy activa la
PKA fosforilando a la DARPP-32 manteniéndola activa y por lo tanto inhibida a la PP-I. La PP-I es la que desfosforila a la bomba de NA/K para que esté activa, pero no puede ser desfosforilada porque la PP-I esta inhibida, por lo tanto la bomba estará inactiva.
Ahora resulta que llego un activador del sistema fosfatidil inositol difosfato y por lo tanto aumento el calcio, al aumentar el calcio se activa una fosfatasa calcio calmodulina dependiente (también hay fosfatasas calcio calmodulina dependientes), y esta desfosforila a la DARPP-32, ahora ¿Cómo estará la activación de la bomba de potasio si aumenta la concentración de calcio?
R/ La bomba de Na/K estará activa, porque si aumenta el calcio se activara a la fosfatasa calcio calmodulina dependiente que desfosforila a la DARPP-32, la DARPP-32 desfosforilada esta inactiva por lo que no podrá inhibir a la PP-I, entonces esta PP-I podrá desfosforilar a la bomba para que este activa