







Idiomas
Páginas
Jurídico


METABOLISMO
Metabolismo de Glúcidos
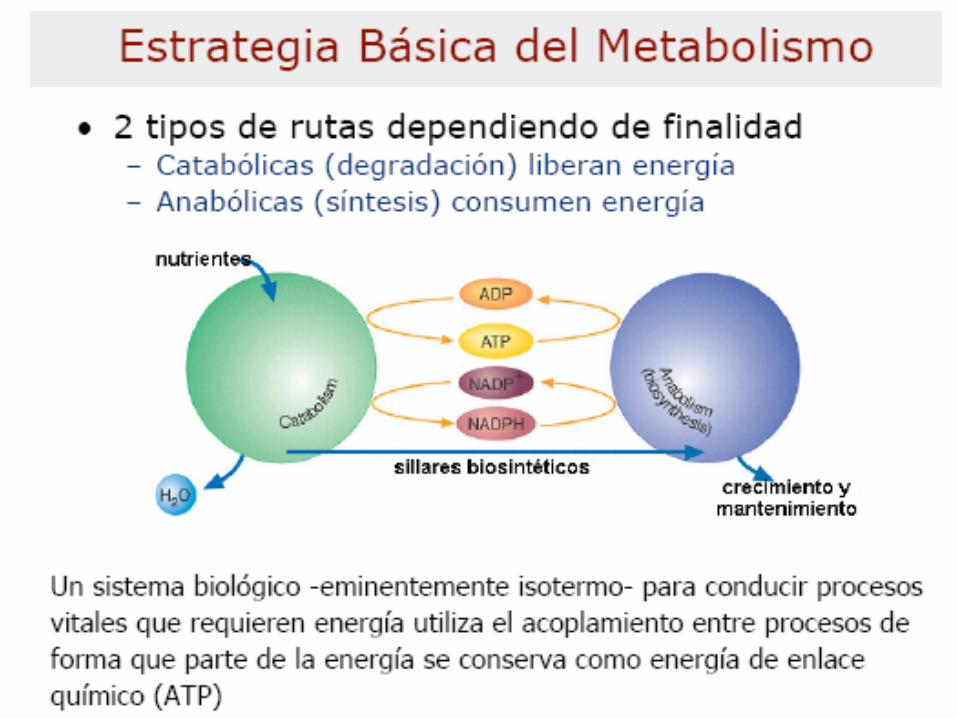
POLÍMEROS
Proteínas
Ács. Nucleicos
Polisacáridos
Lípidos
MONÓMEROS
Aminoácidos
Nucleótidos
Azúcares
ÁcidosGrasos y Glicerol
INTERMEDIARIOS
METABÓLICOS
Piruvato
Acetil-CoA
Intermediarios de Krebs
Moléculas
pequeñas
sencillas
H2O, CO2,
NH3
EnergíaEnergía
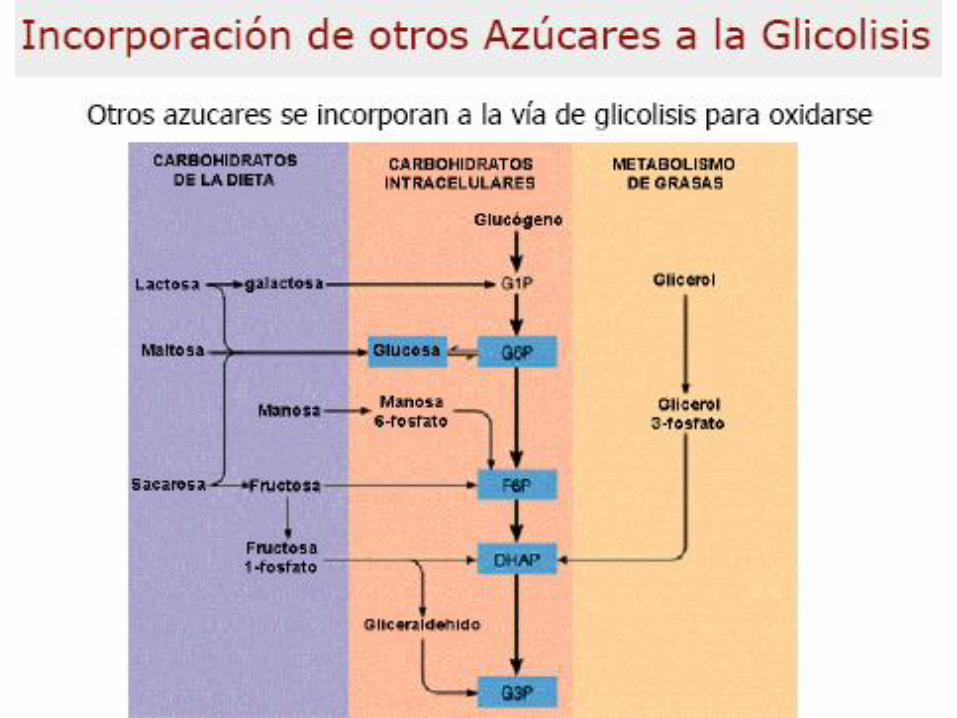
Incorporación
Neta
Producción
Neta
CATABOLISMO ANABOLISMO
Nivel
1
2
3
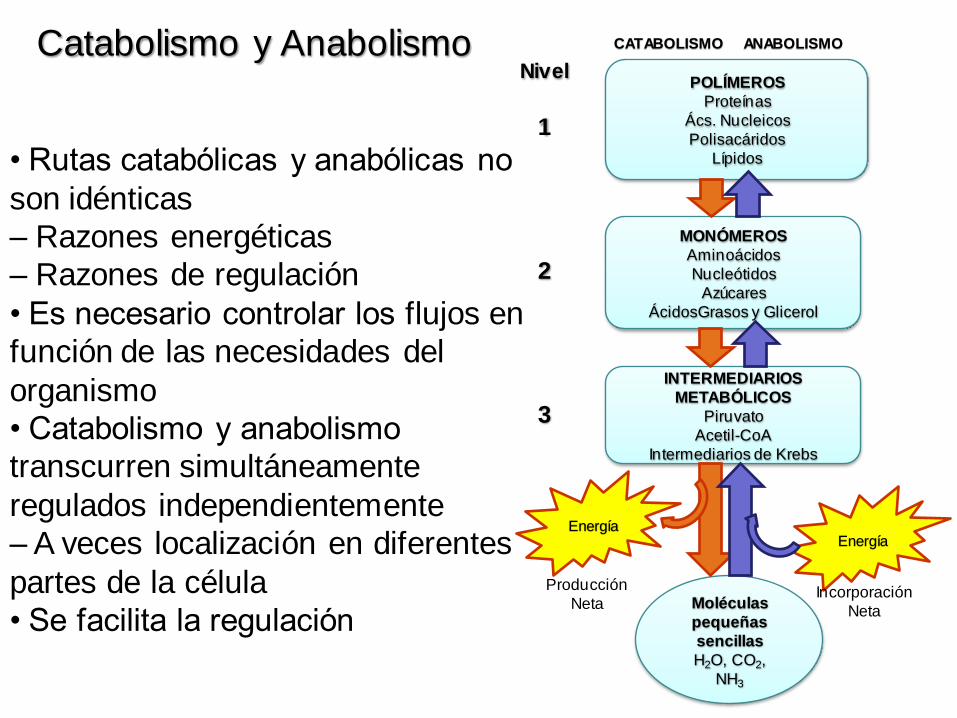
• Rutas catabólicas y anabólicas no
son idénticas
– Razones energéticas
– Razones de regulación
• Es necesario controlar los flujos en
función de las necesidades del
organismo
• Catabolismo y anabolismo
transcurren simultáneamente
regulados independientemente
– A veces localización en diferentes
partes de la célula
• Se facilita la regulación
Catabolismo y Anabolismo

• Principales rutas del
metabolismo de los hidratos de
carbono
• Glicolisis
– Fases I y II de la glicolisis
– Incorporación de otros
azúcares a la glicolisis
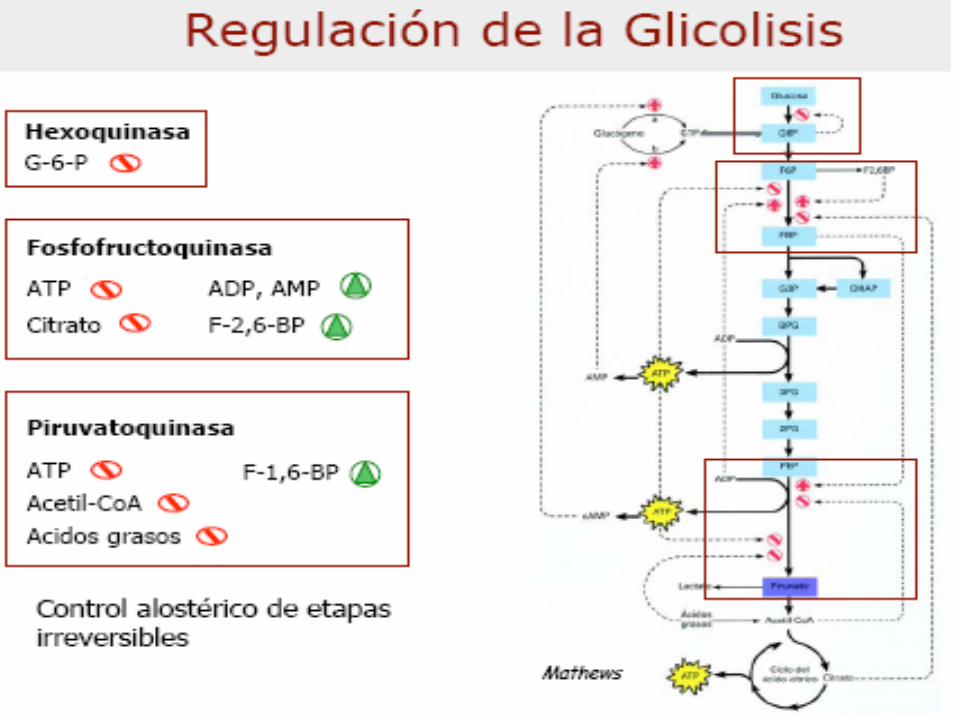
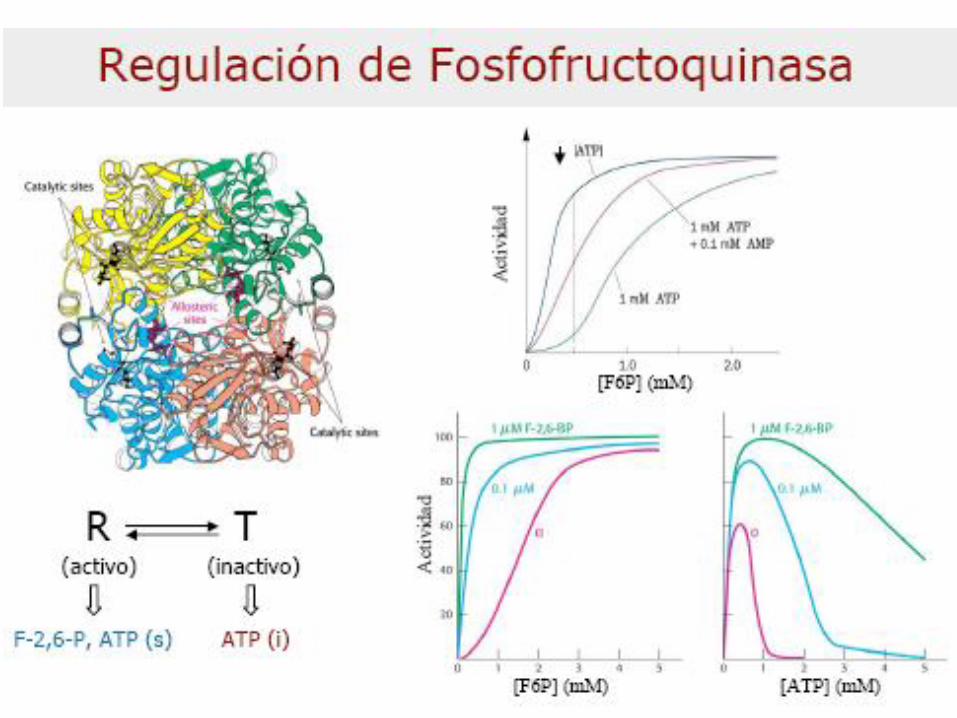
– Regulación
• Destinos del piruvato
– Fermentaciones
– Descarboxilación oxidativa
• Rendimiento de la oxidación
aerobia de la glucosa
• Lanzaderas del NADH
• Gluconeogénesis
– Glicolisis vs
gluconeogénesis
– Etapas diferenciales de la
gluconeogénesis
– Regulación
glicolisis/gluconeogénesis
– Ciclo de Cori
• Metabolismo del glucógeno
y su regulación
• Efectos del glucagón y de
la insulina sobre el
metabolismo de los
hidratos de carbono
• Ruta de los fosfatos de
pentosa
Metabolismo de Hidratos de Carbono
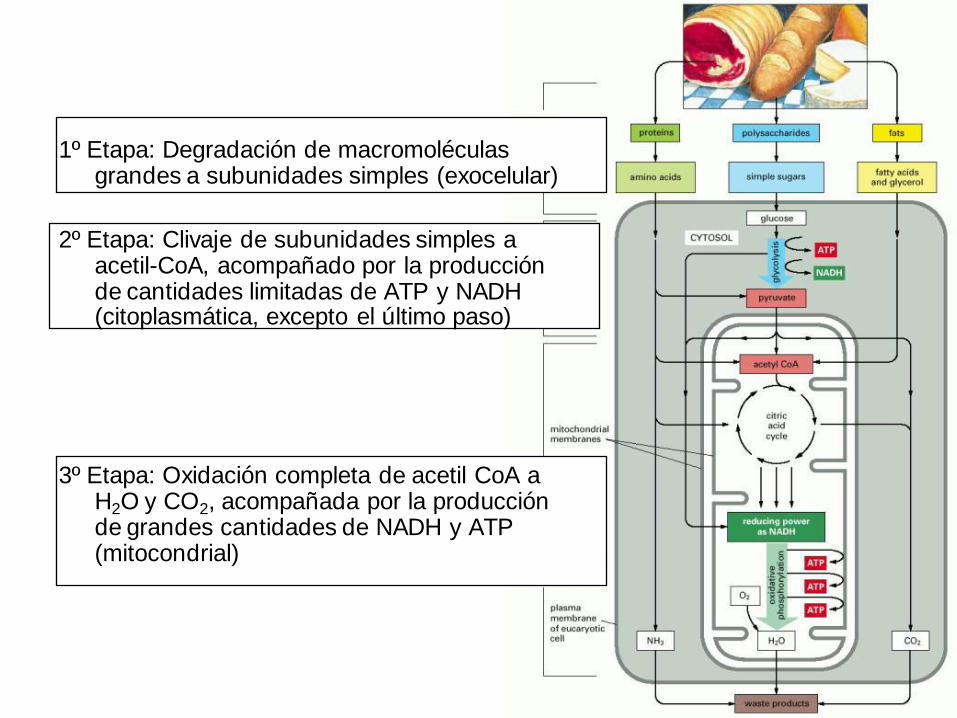
1º Etapa: Degradación de macromoléculas grandes a subunidades simples (exocelular)
2º Etapa: Clivaje de subunidades simples a acetil-CoA, acompañado por la producción de cantidades limitadas de ATP y NADH (citoplasmática, excepto el último paso)
3º Etapa: Oxidación completa de acetil CoA a H2O y CO2, acompañada por la producción de grandes cantidades de NADH y ATP (mitocondrial)

Enzimas Digestivas
Poli. -AMI 1-4 de polisac.
Oligo . Glucosidasas AMI Pancr h. 1-2 restos de ramif
Disac. Oligosac.: Maltotriosa, Maltosa,
Dextrinas, Glc.
Oligosacaridasas (rib. en cepillo), pH 6.0,
Exo 1-4- -D-Glucosidasa (C4 no reductor)
*Dextrinasa, Isomaltasa, Oligo 1-6-glucosidasa 1-
6 y 1-4
*Sacarasa o β-Fructofuranosidasa ( 1-2 y 1-6)
Lactasa o β-Galactosidasa
Trehalasa 1-1 (insectos – hongos)
HC no digeribles en monogástricos: Celulosa, inulina, agar, heteropolisacáridos
vegetales (fibra)
Lactosa (en algunos adultos H° = orientales y negros americanos)

Absorción de monosacáridos
Ribete en cepillo y
Membrana contraluminal
/ capilares
• D-Glc = 100%
• D-Gal = 110 %
• D-Fru = 43 %
• D-Man = 19 %
• D-Xil = 15 %
• D-Arab = 9 %

Glucostato hepático
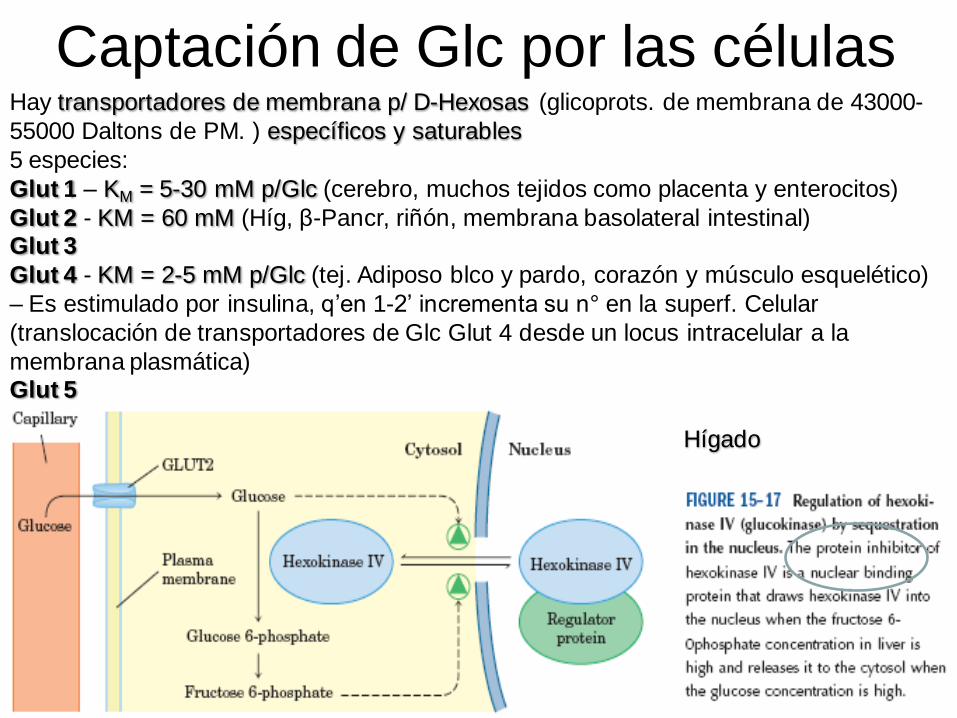
Captación de Glc por las célulasHay transportadores de membrana p/ D-Hexosas (glicoprots. de membrana de 43000-
55000 Daltons de PM. ) específicos y saturables
5 especies:
Glut 1 – KM = 5-30 mM p/Glc (cerebro, muchos tejidos como placenta y enterocitos)
Glut 2 - KM = 60 mM (Híg, β-Pancr, riñón, membrana basolateral intestinal)Glut 3
Glut 4 - KM = 2-5 mM p/Glc (tej. Adiposo blco y pardo, corazón y músculo esquelético)
– Es estimulado por insulina, q’en 1-2’ incrementa su n° en la superf. Celular
(translocación de transportadores de Glc Glut 4 desde un locus intracelular a la
membrana plasmática)Glut 5
Hígado
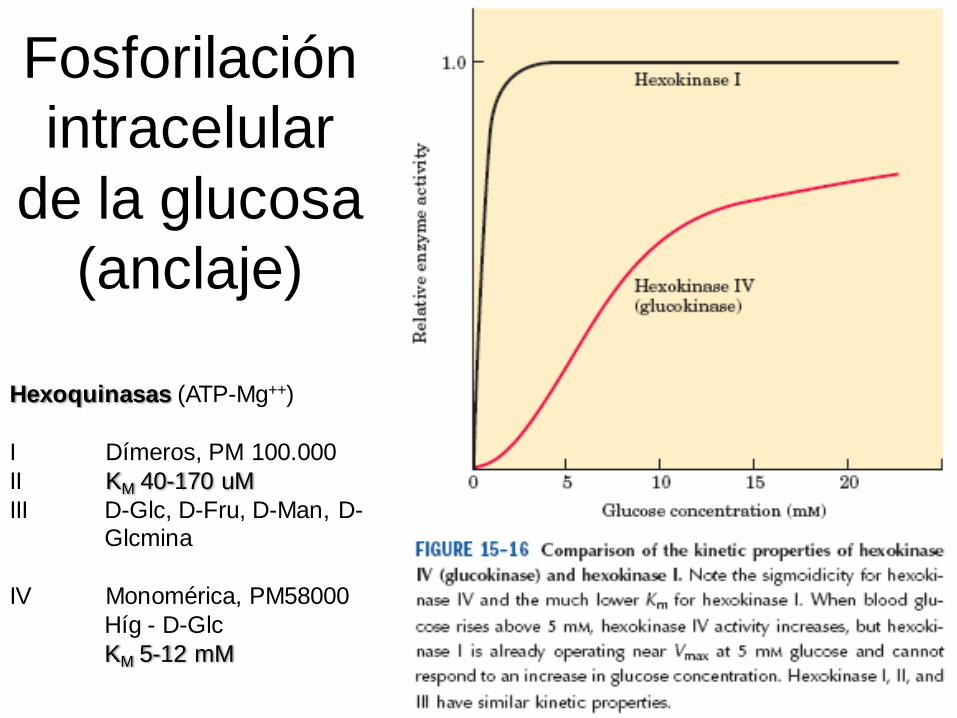
Fosforilación
intracelular
de la glucosa
(anclaje)
Hexoquinasas (ATP-Mg++)
I Dímeros, PM 100.000
II KM 40-170 uM
III D-Glc, D-Fru, D-Man, D-Glcmina
IV Monomérica, PM58000
Híg - D-Glc
KM 5-12 mM

Destino de la glucosa dentro de las
células procariotas y eucariotas.
• Vocabulario
Ej. GLUT, hexoquinasas, glucólisis, gluconeogénesis, glucogenogénesis,
glucogenolisis, etc.
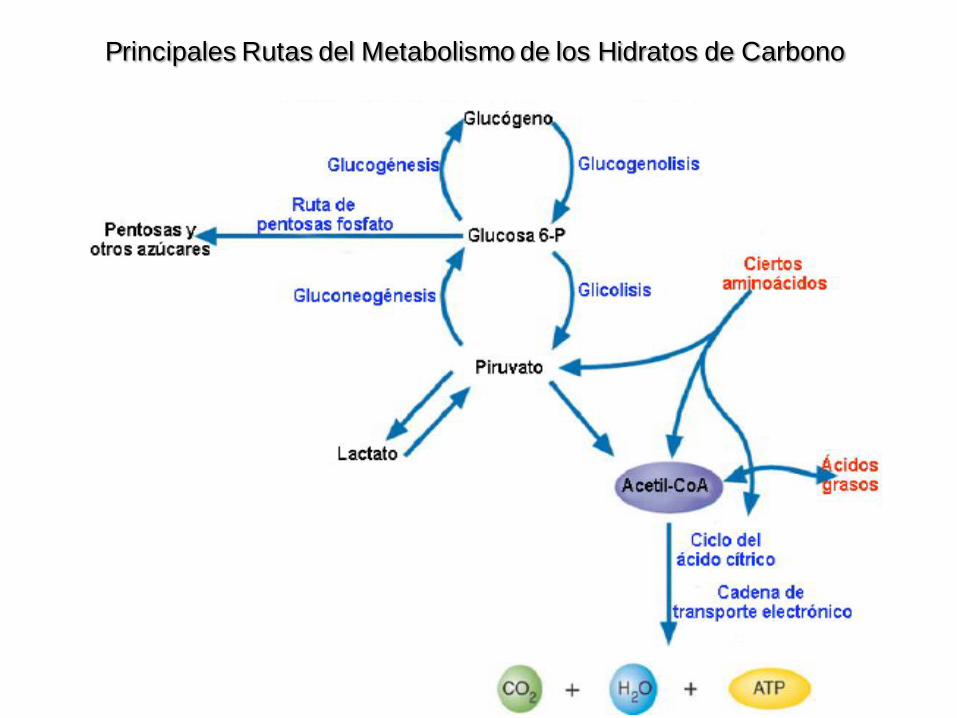
Principales Rutas del Metabolismo de los Hidratos de Carbono
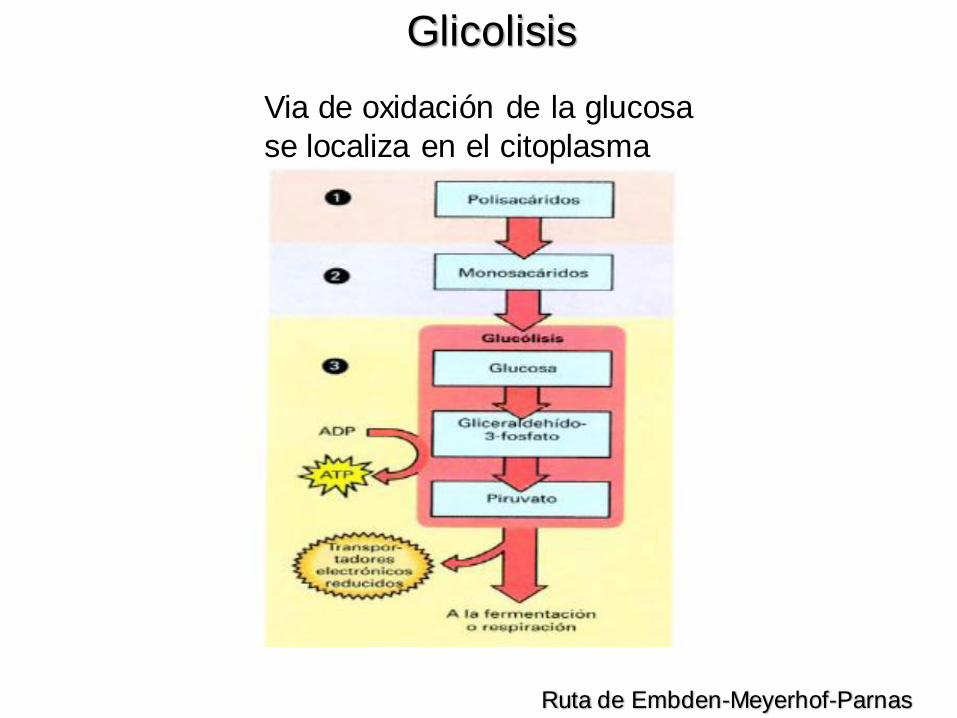
Via de oxidación de la glucosa
se localiza en el citoplasma
Ruta de Embden-Meyerhof-Parnas
Glicolisis
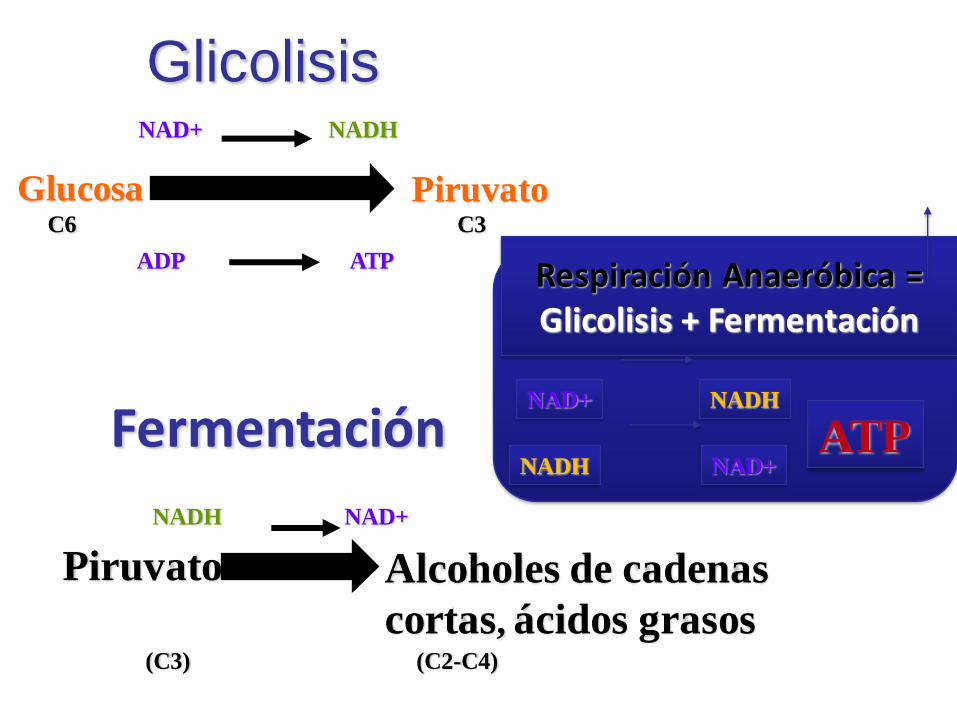
GlicolisisNAD+ NADH
Glucosa PiruvatoC6 C3
ADP ATP
Fermentación
Piruvato
(C3)
NADH NAD+
Alcoholes de cadenas
cortas, ácidos grasos(C2-C4)
Respiración Anaeróbica = Glicolisis + Fermentación
NAD+ NADH
NADH NAD+ATP

Anaerobiosis
• El oxígeno no es el aceptor final sino otra
molécula que se reduce:
Levaduras usan acetaldehido -> etanol
Músculo usa piruvato -> lactato
Estos productos son sustancias orgánicas y
el proceso se denomina fermentación (ej.
Microorganismos ruminales) y hay gran
pérdida de energía.
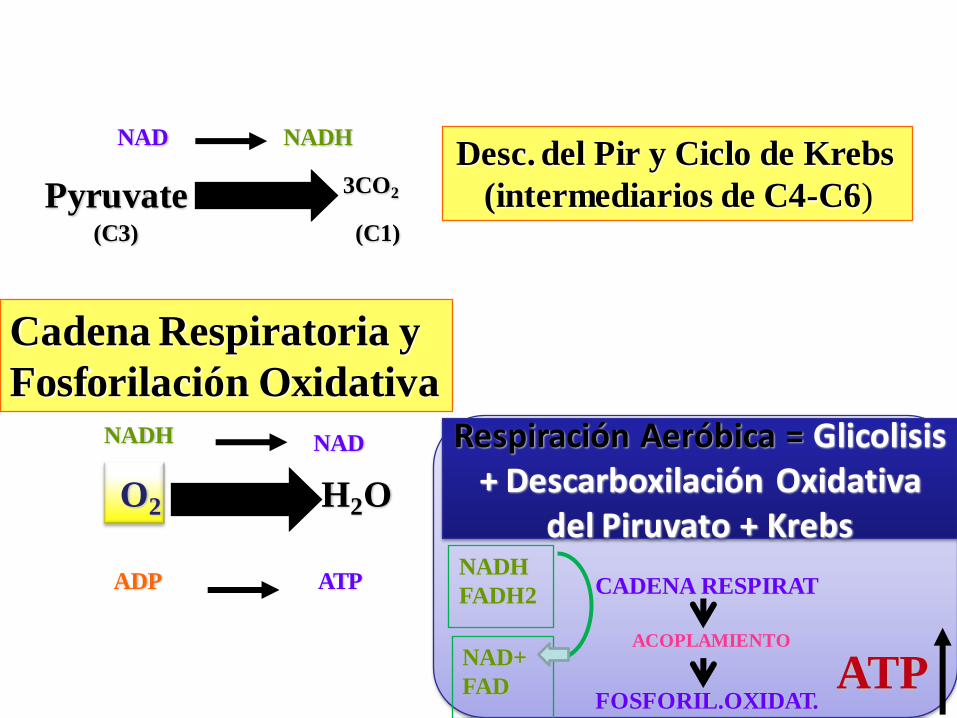
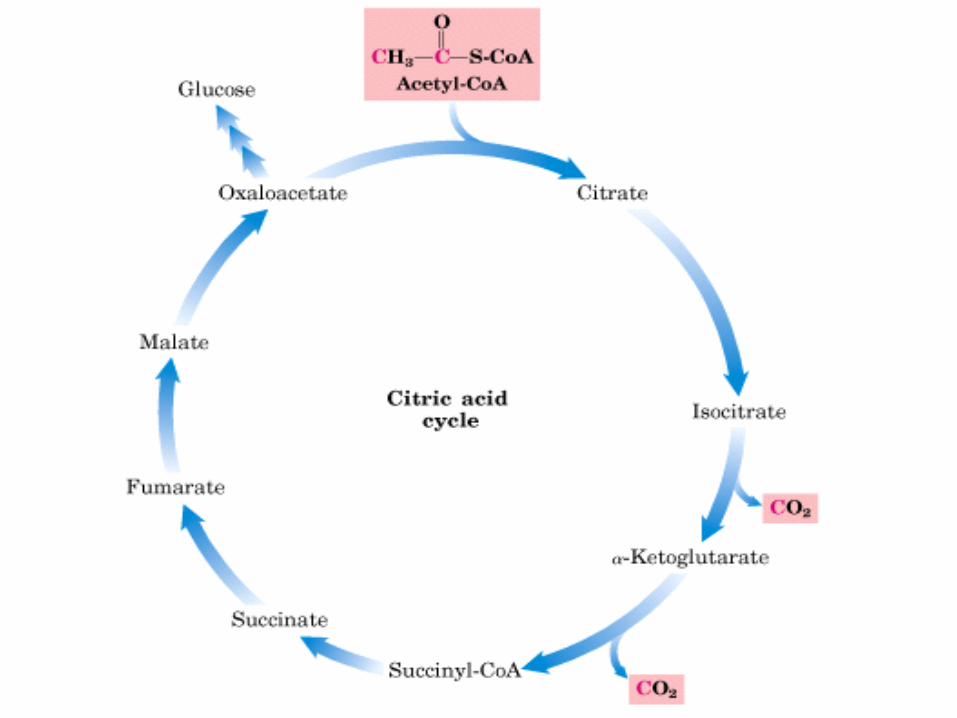
Desc. del Pir y Ciclo de Krebs
(intermediarios de C4-C6)
Cadena Respiratoria y
Fosforilación OxidativaNADH NAD
O2 H2O
ADP ATP
Pyruvate 3CO2
(C3)
NAD NADH
(C1)
Respiración Aeróbica = Glicolisis + Descarboxilación Oxidativa
del Piruvato + Krebs
CADENA RESPIRAT
ACOPLAMIENTO
FOSFORIL.OXIDAT.
NADH
FADH2
NAD+
FAD ATP
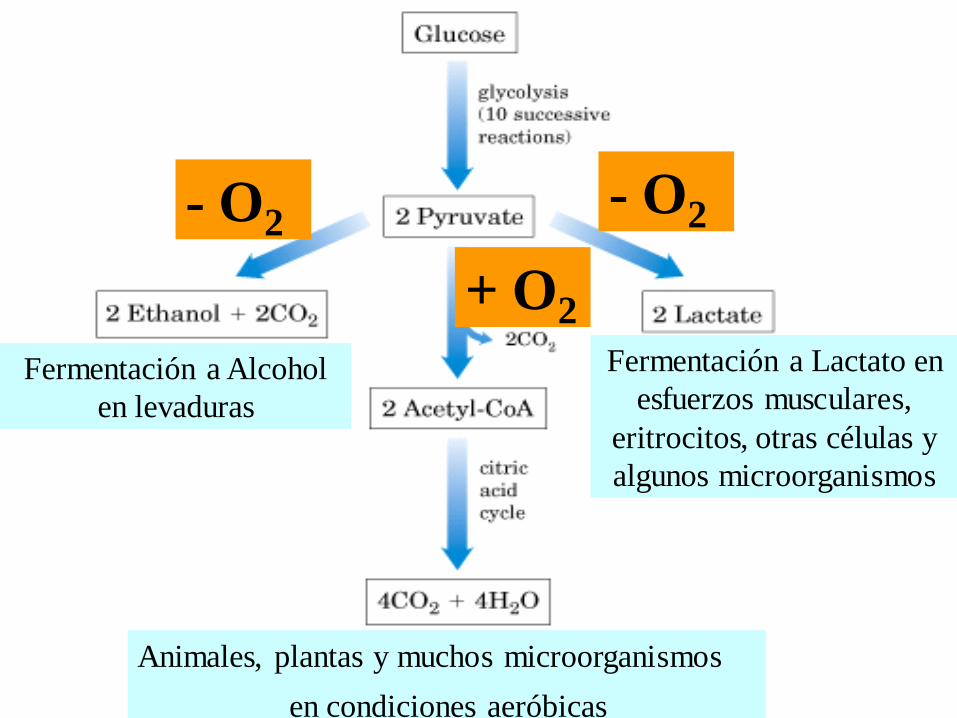
Animales, plantas y muchos microorganismos
en condiciones aeróbicas
Fermentación a Lactato en
esfuerzos musculares,
eritrocitos, otras células y
algunos microorganismos
Fermentación a Alcohol
en levaduras
- O2- O2
+ O2
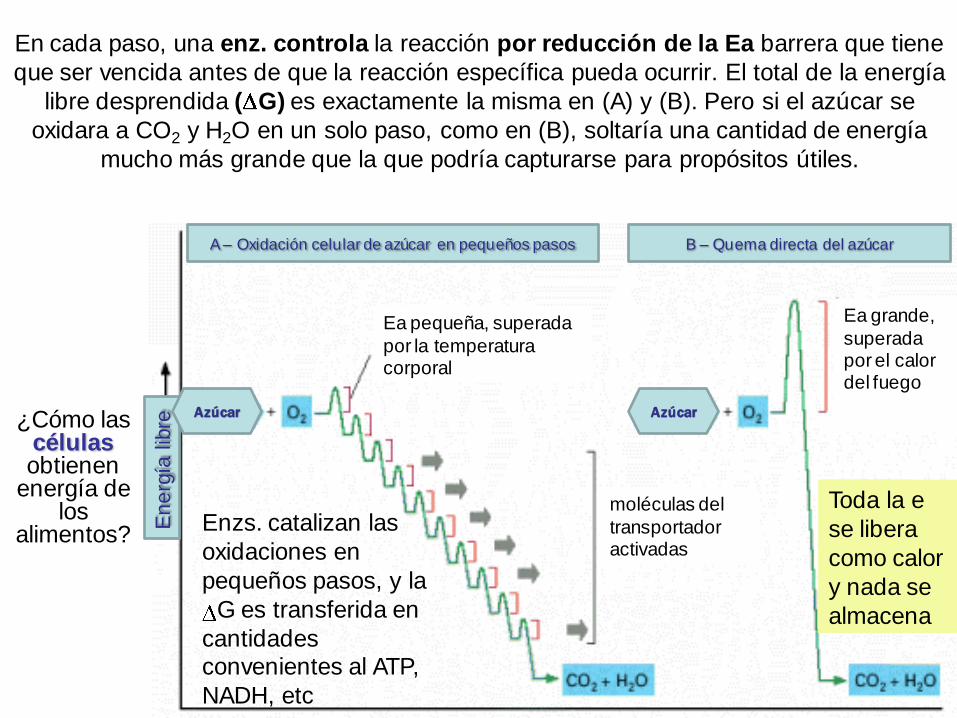
En cada paso, una enz. controla la reacción por reducción de la Ea barrera que tiene
que ser vencida antes de que la reacción específica pueda ocurrir. El total de la energía
libre desprendida ( G) es exactamente la misma en (A) y (B). Pero si el azúcar se
oxidara a CO2 y H2O en un solo paso, como en (B), soltaría una cantidad de energía
mucho más grande que la que podría capturarse para propósitos útiles.
¿Cómo las célulasobtienen
energía de los
alimentos?Enzs. catalizan las
oxidaciones en
pequeños pasos, y la
G es transferida en
cantidades convenientes al ATP,
NADH, etc
Toda la e
se libera
como calor
y nada se
almacena
Ea pequeña, superada
por la temperatura corporal
Ea grande,
superada por el calor del fuego
moléculas del
transportador activadas
B – Quema directa del azúcarA – Oxidación celular de azúcar en pequeños pasos
En
erg
ía lib
re
Azúcar Azúcar

GLUCÓLISIS
• Citoplasmática.
• 10 reacciones (2 etapas/fases), c/u
c/reaccs cataliz x enzimas.
• Transformar una molécula de glucosa (C6)
en dos moléculas de ácido pirúvico (C3).
• Ganancia neta de 2 ATP y 2 NADH + H+

La glucosa es un combustible muy
importante para la mayoría de
organismos
Fase I
Se consumen 2 ATP
Fase II
Se producen 4 ATP
Rendimiento
2 moles ATP/mol glucosa
Fases de la Glicolisis

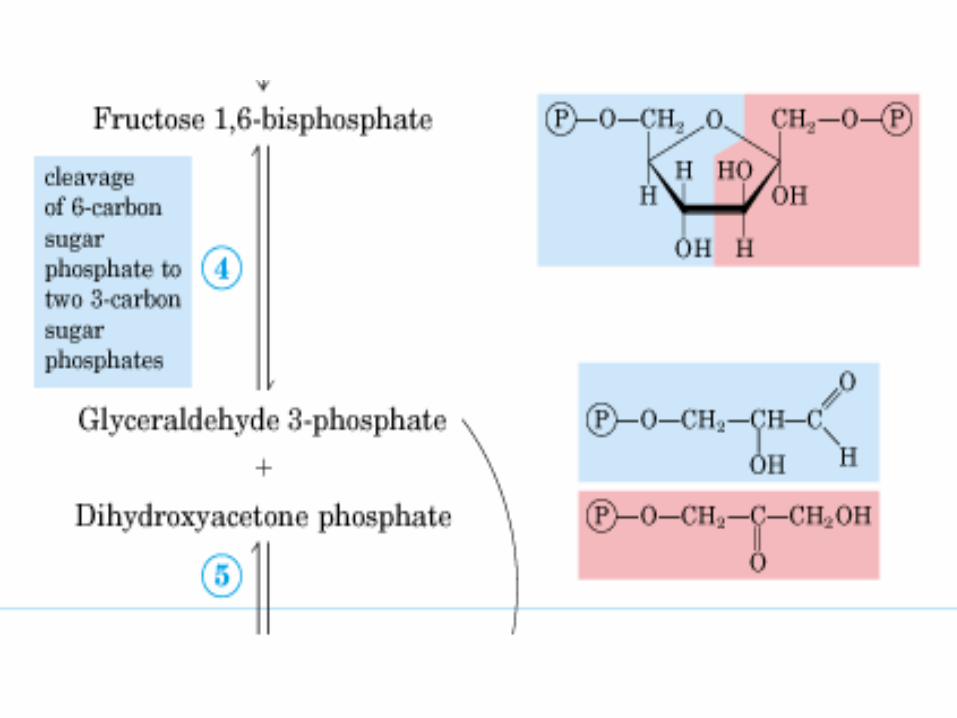
Primera parte
• c/gasto de 2 ATP, para fosforilar
la glucosa y la fructosa.
• Al final se obtienen, dos moléculas
de P-GALD, ya que la molécula de
DHA-P (dihidroxiacetona-fosfato),
se puede transformar en P-GALD.
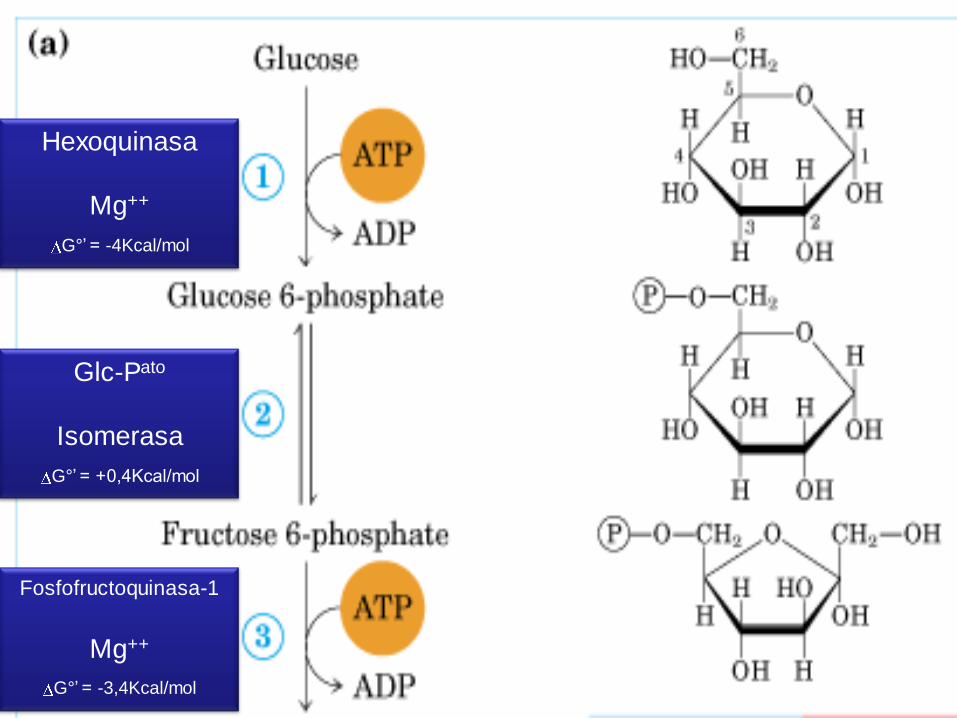
Hexoquinasa
Mg++
G°’ = -4Kcal/mol
Glc-Pato
Isomerasa
G°’ = +0,4Kcal/mol
Fosfofructoquinasa-1
Mg++
G°’ = -3,4Kcal/mol
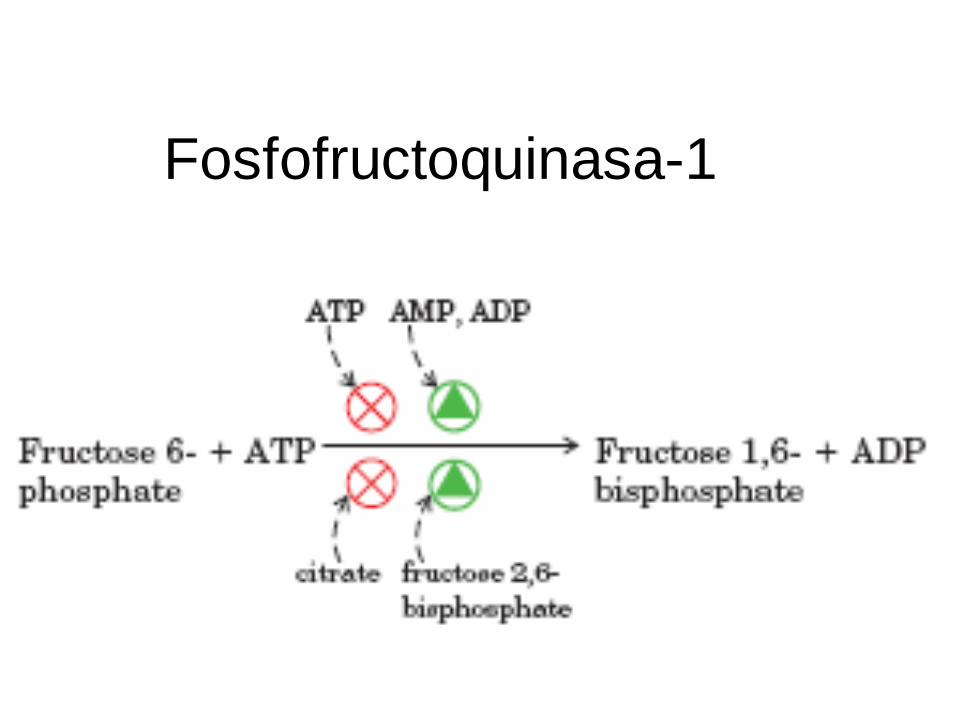
Fosfofructoquinasa-1
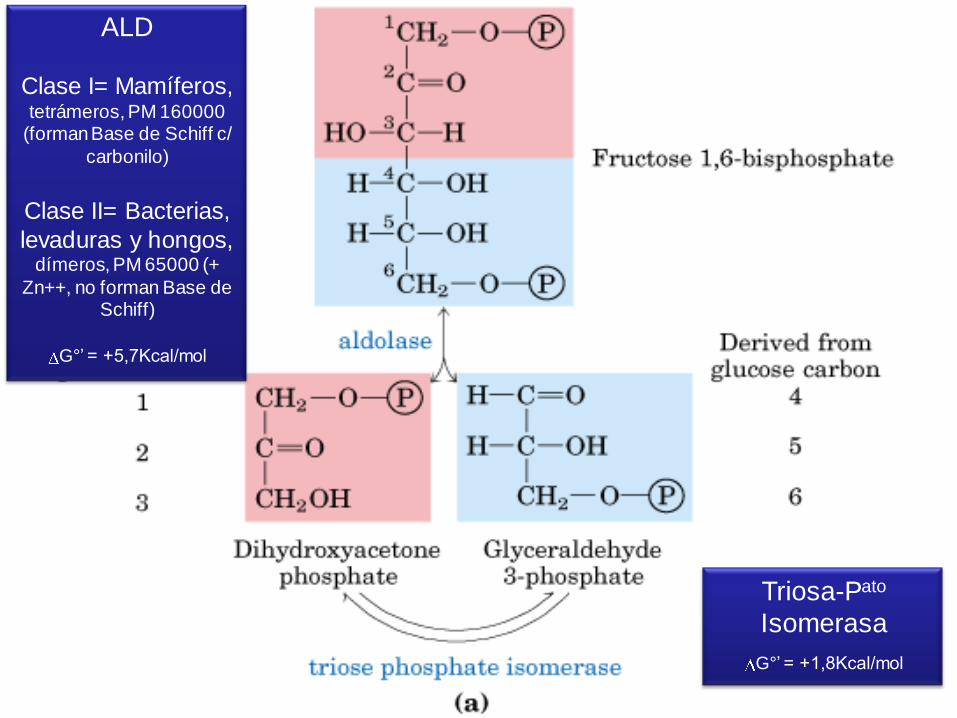
ALD
Clase I= Mamíferos, tetrámeros, PM 160000
(forman Base de Schiff c/
carbonilo)
Clase II= Bacterias,
levaduras y hongos, dímeros, PM 65000 (+
Zn++, no forman Base de Schiff)
G°’ = +5,7Kcal/mol
Triosa-Pato
Isomerasa
G°’ = +1,8Kcal/mol
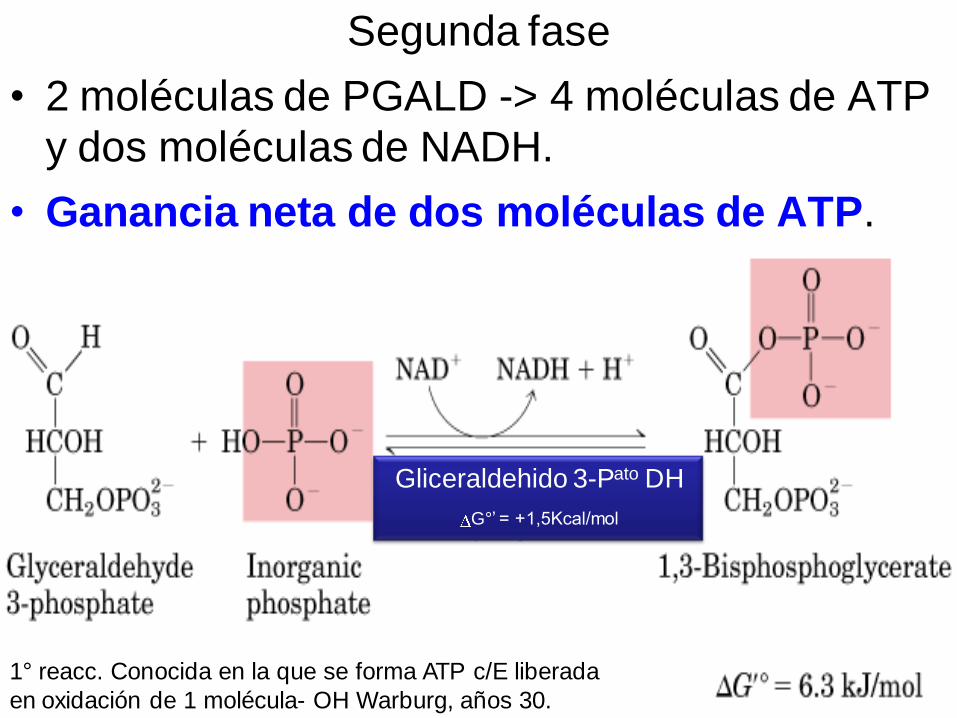
Segunda fase
• 2 moléculas de PGALD -> 4 moléculas de ATP
y dos moléculas de NADH.
• Ganancia neta de dos moléculas de ATP.
Gliceraldehido 3-Pato DH
G°’ = +1,5Kcal/mol
1° reacc. Conocida en la que se forma ATP c/E liberada
en oxidación de 1 molécula- OH Warburg, años 30.

Entre el Gliceraldehido-3-P (Sustrato) y el grupo –SH de una cisteína de la
Enz (Gliceraldehido-3-P-DH) se forma un enlace covalente.
La Enz también se une No Covalentemente al NAD+
La oxidación del Gliceraldehido-3-P ocurre cuando 2 electrones + 1 átomo de
H son transferidos al NAD+
Parte de la E liberada por la oxidación del aldehido es así almacenada en el
NADH y parte queda en el enlace entre la Enz y su sustrato (unión tioéster de
alta E)
Una molécula de Fosfato inorgánico desplaza el enlace de alta E de la Enz
p/crear un enlace acil-anhidrido de alta E.
El enlace P es transferido al ADP p/formar ATP
Mucha de la E de oxidación ha sido almacenada en transportadores activos
ATP y NADH.
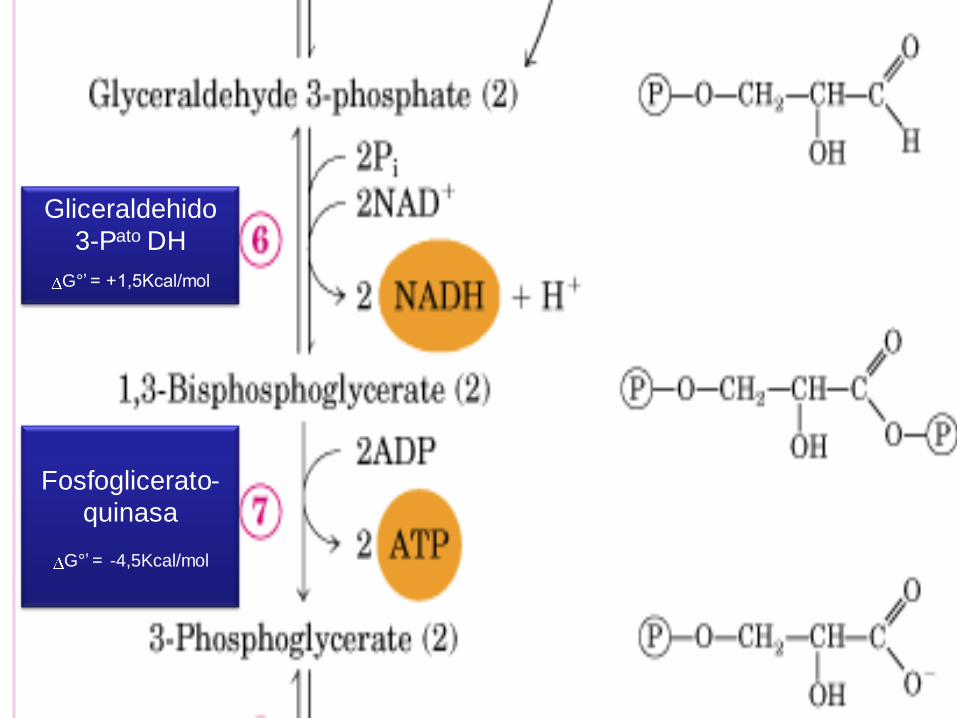
Fosfoglicerato-
quinasa
G°’ = -4,5Kcal/mol
Gliceraldehido
3-Pato DH
G°’ = +1,5Kcal/mol
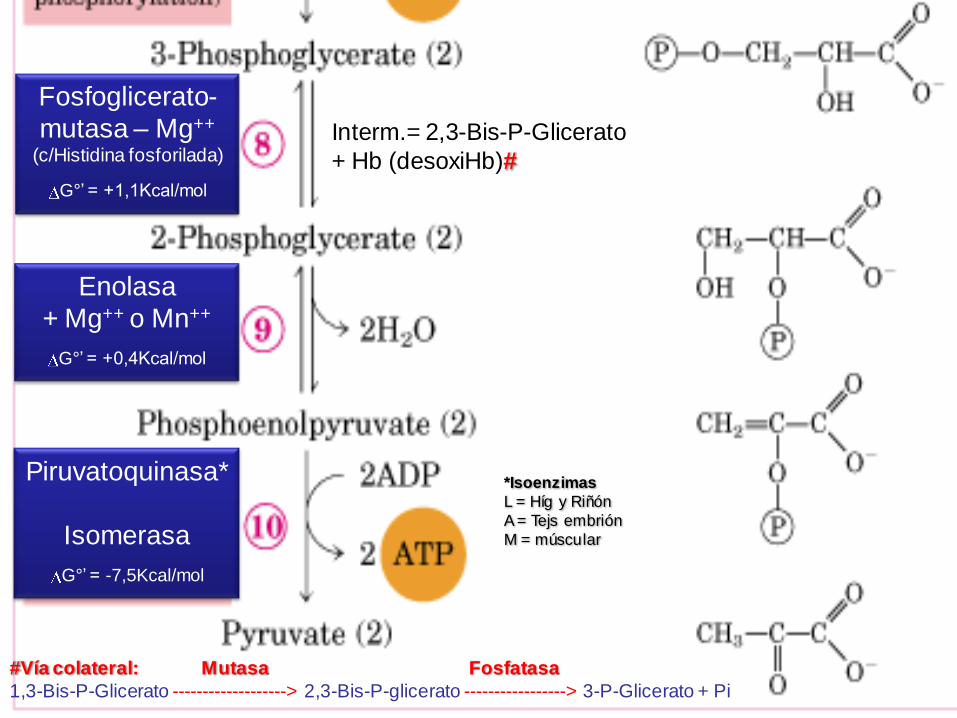
Piruvatoquinasa*
Isomerasa
G°’ = -7,5Kcal/mol
Fosfoglicerato-
mutasa – Mg++
(c/Histidina fosforilada)
G°’ = +1,1Kcal/mol
Interm.= 2,3-Bis-P-Glicerato
+ Hb (desoxiHb)#
#Vía colateral: Mutasa Fosfatasa
1,3-Bis-P-Glicerato -------------------> 2,3-Bis-P-glicerato -----------------> 3-P-Glicerato + Pi
Enolasa
+ Mg++ o Mn++
G°’ = +0,4Kcal/mol
*Isoenzimas
L = Híg y Riñón
A = Tejs embrión
M = múscular
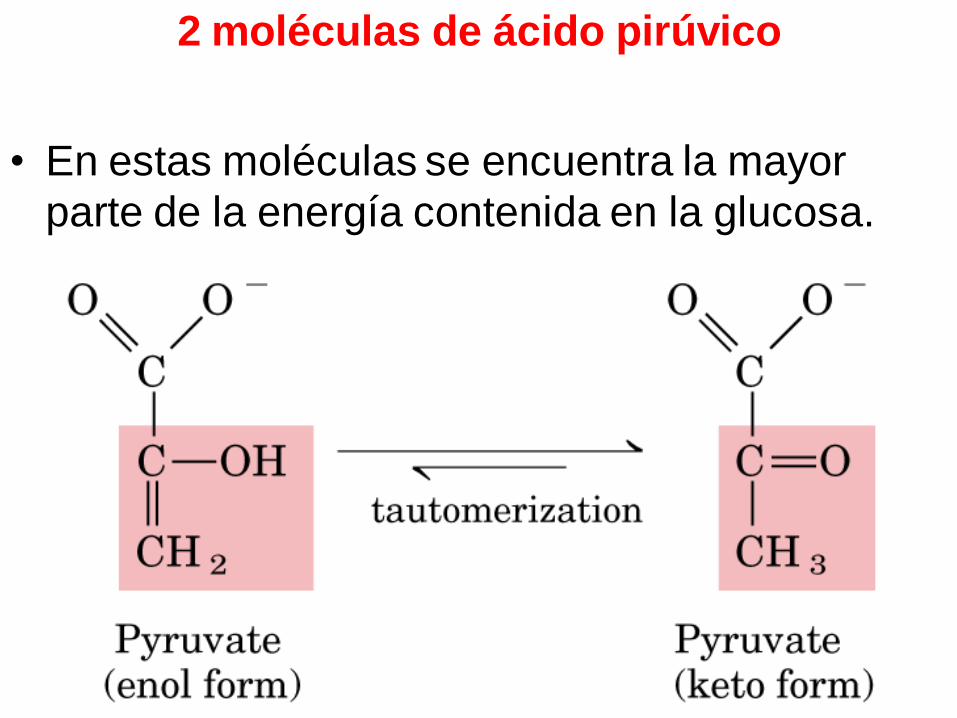
2 moléculas de ácido pirúvico
• En estas moléculas se encuentra la mayor
parte de la energía contenida en la glucosa.
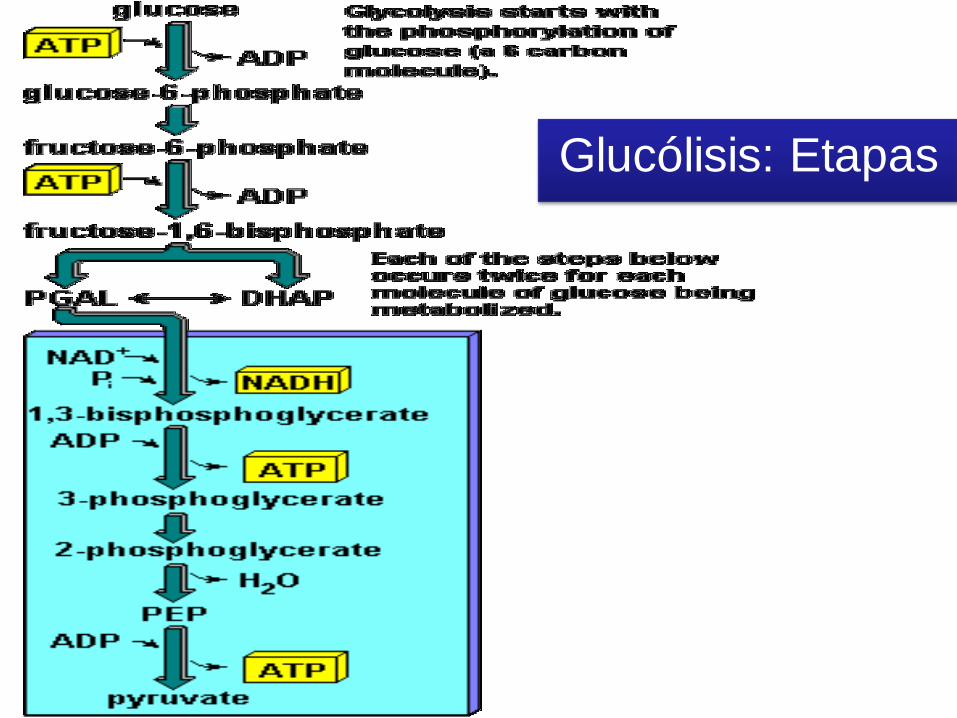
Glucólisis: Etapas
Enlace
enol Pato
Enlace
anhidrido a
1 COO-
Enlace
de Pato en
creatina
Pato
Enlace
anhidrido
al Pato
(enlace
fosfoanhidrido)
Enlace
fosfoéster
PEP
1,3-
Bisfosfoglicerato
Creatina-Pato
(carrier activado
q´almacena E en
músculos
ATP cuando se
hidroliza a ADP
Glc-6-Pato
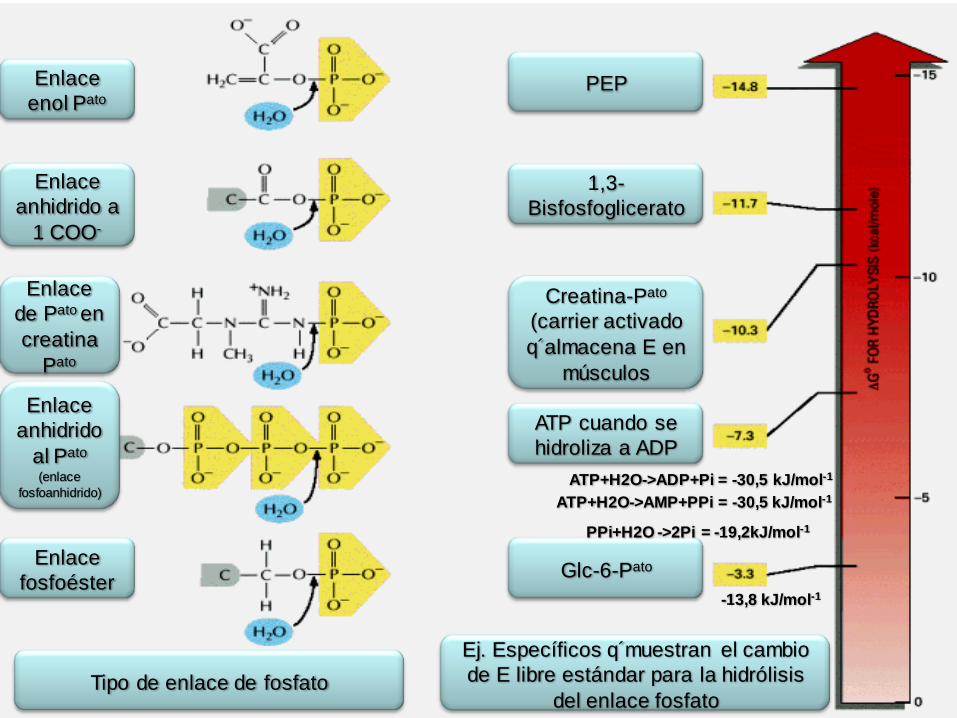
Tipo de enlace de fosfato
Ej. Específicos q´muestran el cambio
de E libre estándar para la hidrólisis
del enlace fosfato
PPi+H2O ->2Pi = -19,2kJ/mol-1
ATP+H2O->ADP+Pi = -30,5 kJ/mol-1
-13,8 kJ/mol-1
ATP+H2O->AMP+PPi = -30,5 kJ/mol-1

Destinos del Piruvato
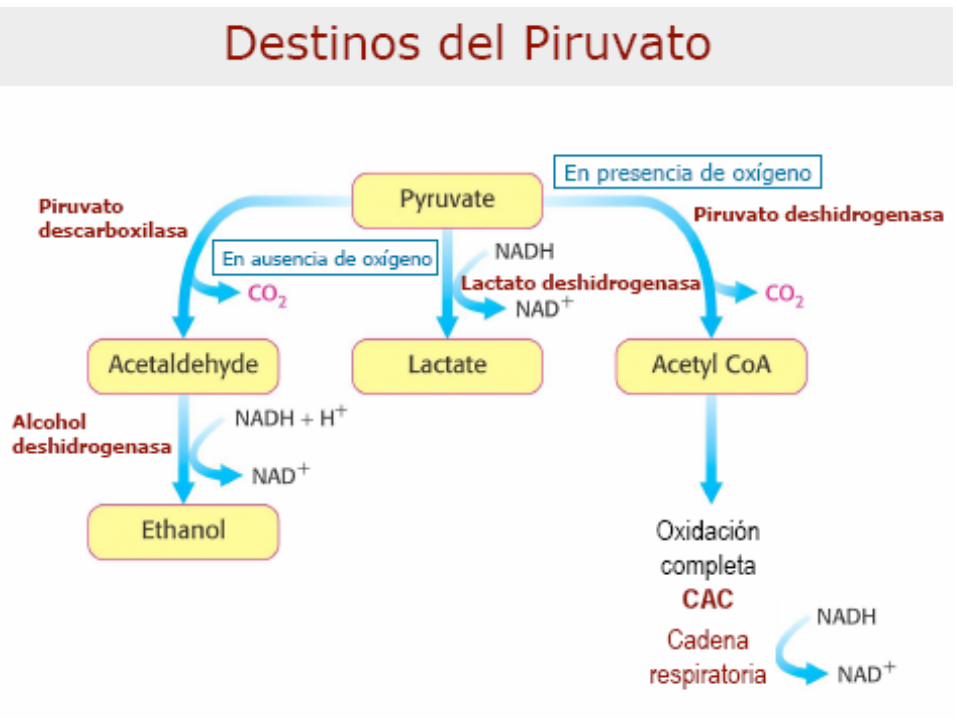

Animales, plantas y muchos microorganismos
en condiciones aeróbicas
Fermentación a Lactato en
esfuerzos musculares,
eritrocitos, otras células y
algunos microorganismos
Fermentación a Alcohol en
levaduras
- O2- O2
+O2
En general:
•Síntesis de Lactato (mamíf,
hongos, protozoos, bacterias)
•Fermentación alcohólica (algas,
levaduras, vegetales)•Otros tipos de fermentaciones
(bacterias)
•Otros destinos del Piruvato
(precursor en vías anabólicas)
Mamíferos: Según estado del metab. Celular y
disponibilidad de O2
•Producción de Lactato
LDH =tetrámero, 140000 PM, 5
isoenz en animales superiores
M4 y M3H KM bajos p/Piruvato
MH3 y H4 KM altos p/Piruvato
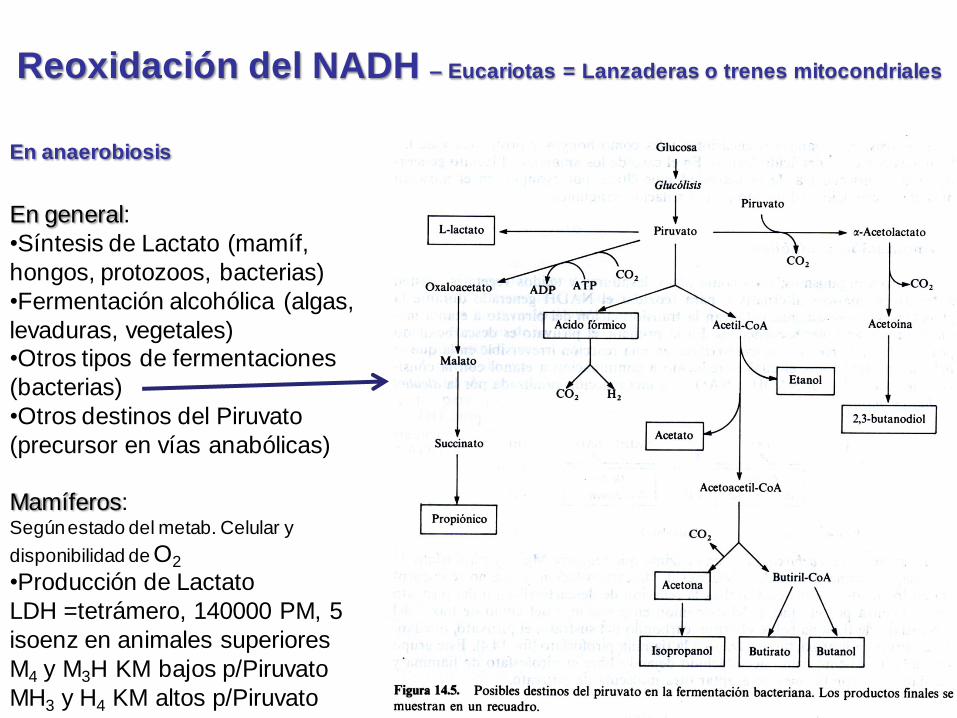
Reoxidación del NADH – Eucariotas = Lanzaderas o trenes mitocondriales
En anaerobiosis

Destinos del
Piruvato (anaerob.)Dos alternativas p/degradar piruvato en
condiciones anaeróbicas son:
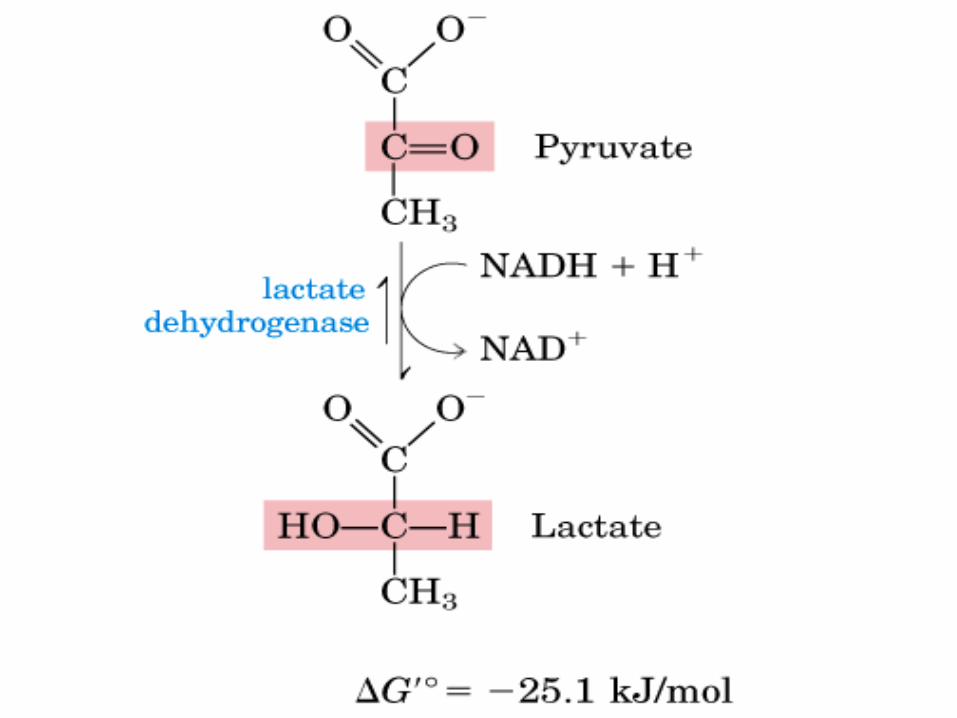
(1)A baja pO2, por ejemplo, en una célula muscular en ejercicio vigoroso, el piruvato producido por glucólisis se convierte en lactato. Esta reacción regenera el NAD+ (oxidado) que es necesario p/ser consumido en paso 6 de la glucólisis (rinde mucho menos E global que la oxidación completa).
(2)En algunos organismos que pueden crecer anaerobicamente, como las levaduras, el piruvato se convierte vía acetaldehido en anhídrido carbónico y etanol. De nuevo, esta senda regenera NAD+ de NADH (necesario p/ permitir continuar la glucólisis).
Ambos, (1) y (2), son ejemplos de fermentaciones.
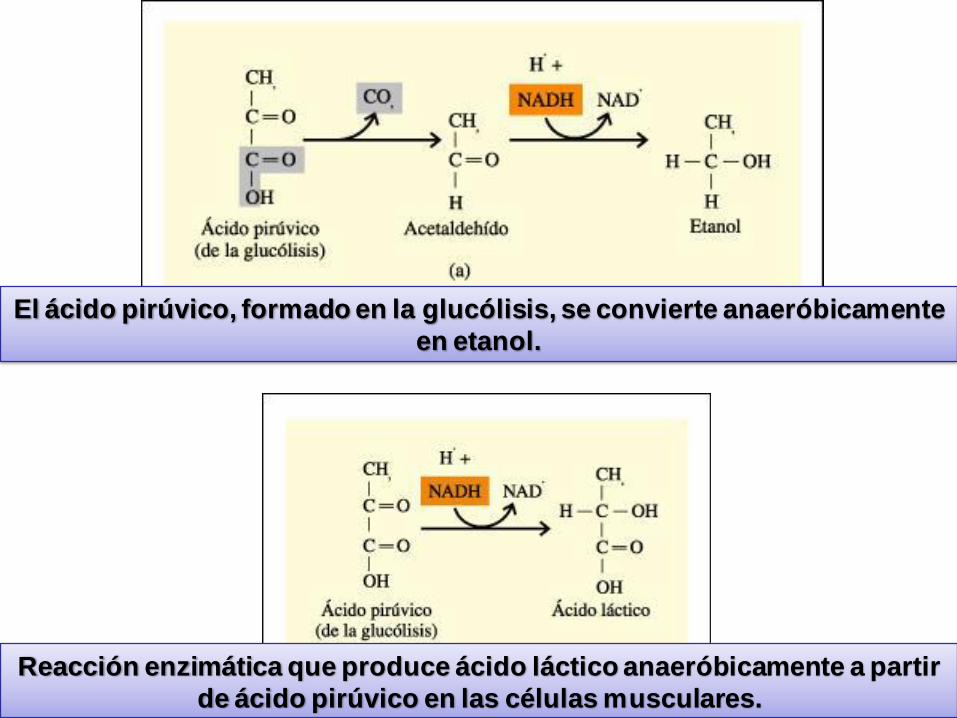
El ácido pirúvico, formado en la glucólisis, se convierte anaeróbicamente
en etanol.
Reacción enzimática que produce ácido láctico anaeróbicamente a partir
de ácido pirúvico en las células musculares.

Matriz mitocondrial, Hay 1 ADN circular y pequeños ribosomas que sintetizan un pequeño número de proteínas. Enzimas de la
oxidación de AG, aminoácidos, ácido pirúvico que se
descarboxila y se encuentran las enzimas del ciclo de
Krebs ( Hans Krebs postuló esta vía metabólica en 1937 y
luego recibió el premio Nobel) o de los ácidos tricarboxílicos, o
ciclo del ácido cítrico.
Membrana Interna: Los sistemas Red-0x del transporte de
electrones se encuentran adosados a las crestas mitocondriales,
el sistema de la fosforilación oxidativa (ocurre tanto en bacterias
aeróbicas como en mitocondrias de células eucarióticas).
Destinos aeróbicos del Piruvato en la
MITOCONDRIA

Descarboxilación Oxidativa del Piruvato
1) Descarboxilación exergónica
2) Formación de sulfoester de alta E con Lipoato
3) Transtiolación isoexergónica del acetilo desde el Lipoato al HS-CoA
Enzima Cosustrato prostético Sustrato soluble
Piruvato deshidrogenasa ó Pir. descarboxilasa
E1 TPP = Tiamina pirofosfato
Dihidrolipoil transacetilasa E2 Lipoamida Coenzima A
Dihidrolipoil deshidrogenasa
E3 FAD NAD+
Otras enzimas con igual mecanismo:
a)Alfa-cetoglutarato DH,
b)Alfa-cetobutirato DH,
c)cadena ramificada cetoácido DH

Resumen del máximo rendimiento energético a
partir de la oxidación de una molécula de glucosa
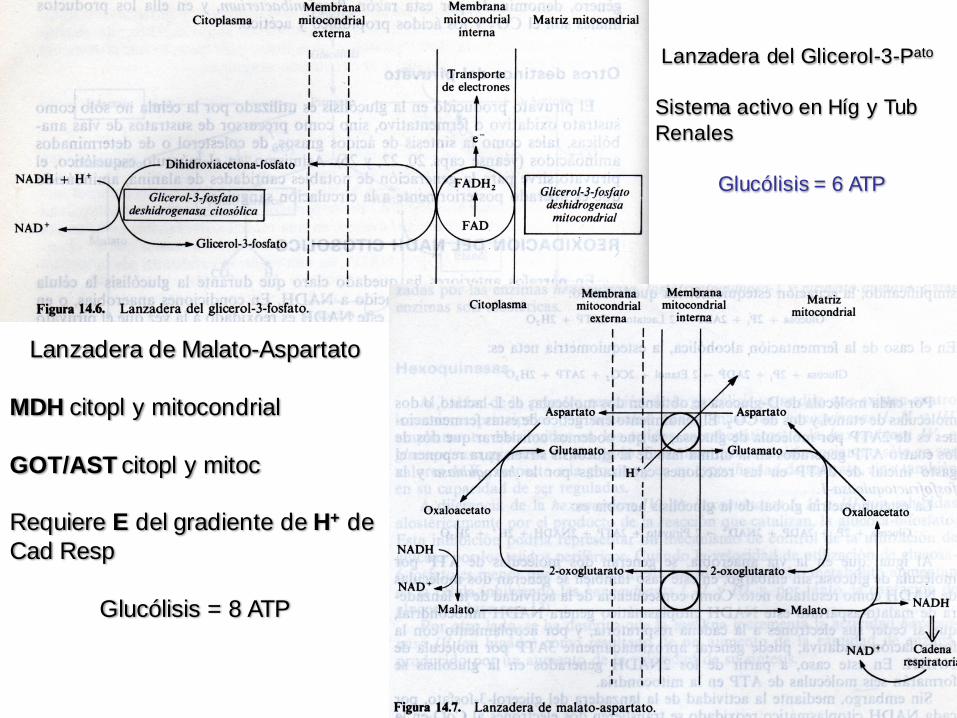
Lanzadera del Glicerol-3-Pato
Sistema activo en Híg y Tub
Renales
Glucólisis = 6 ATP
Lanzadera de Malato-Aspartato
MDH citopl y mitocondrial
GOT/AST citopl y mitoc
Requiere E del gradiente de H+ de
Cad Resp
Glucólisis = 8 ATP

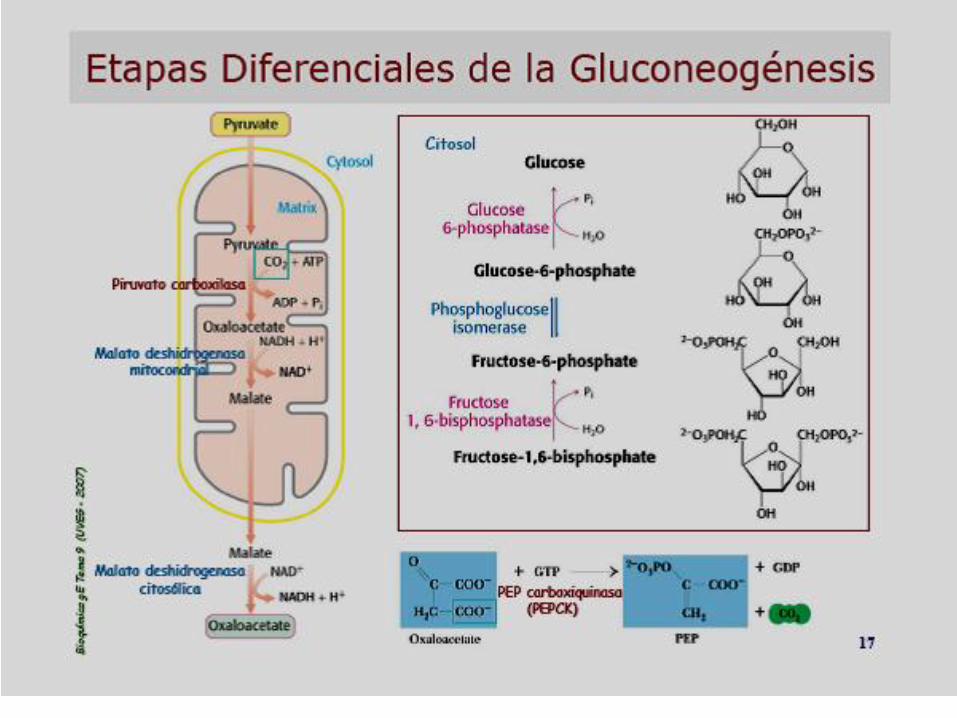
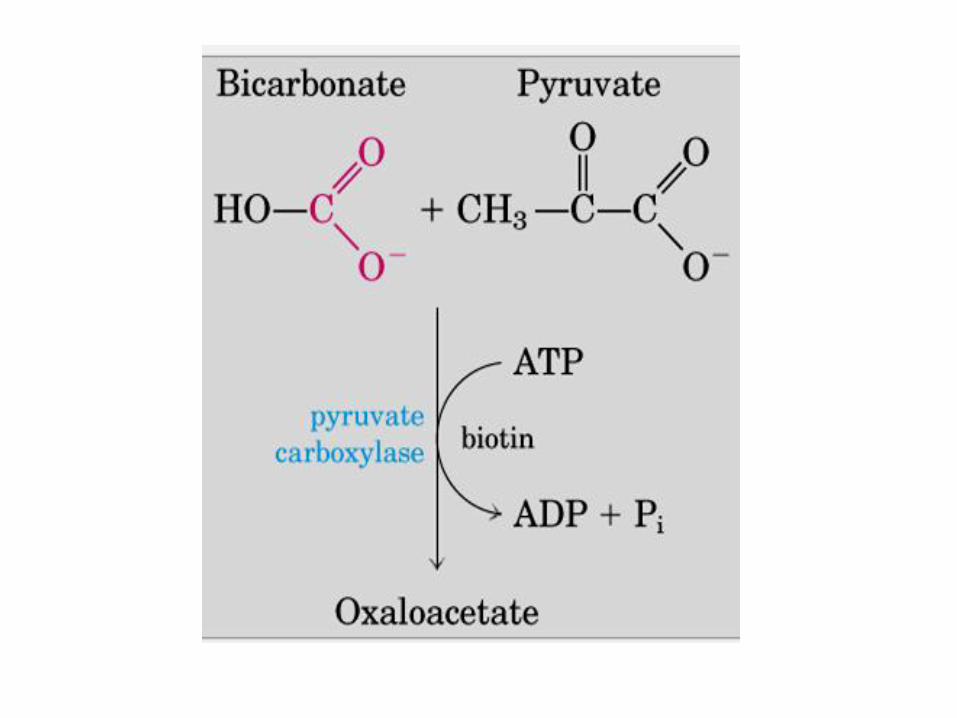
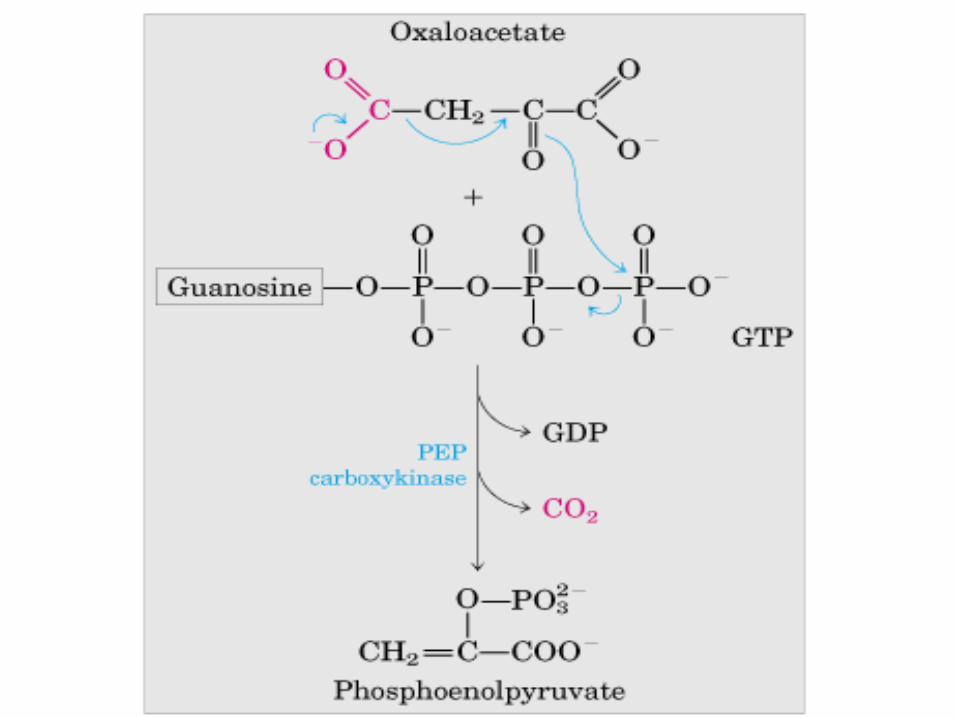
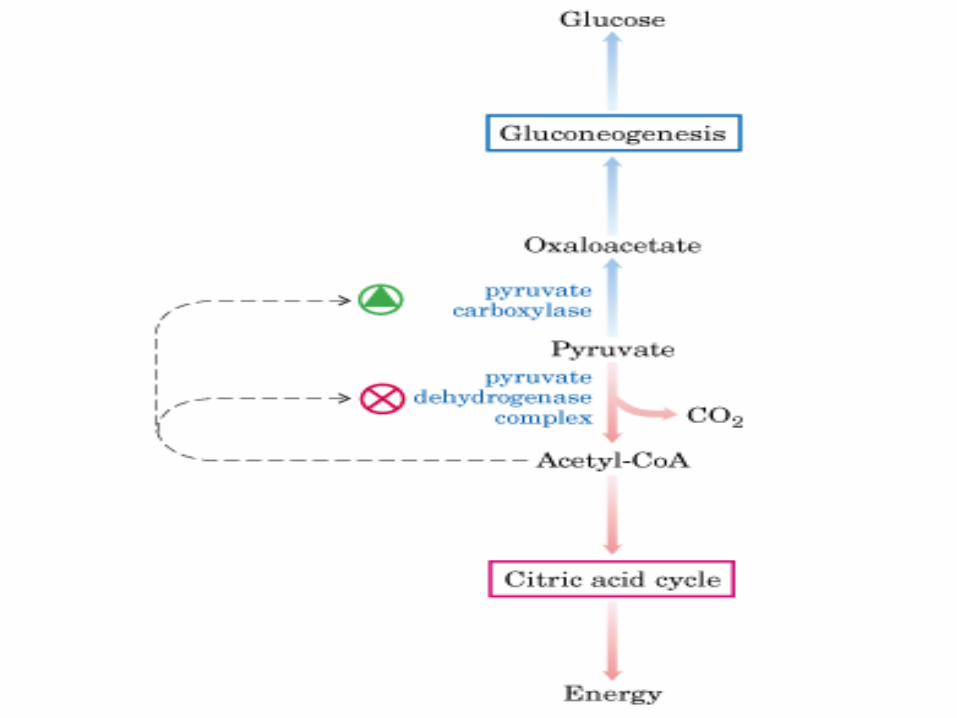
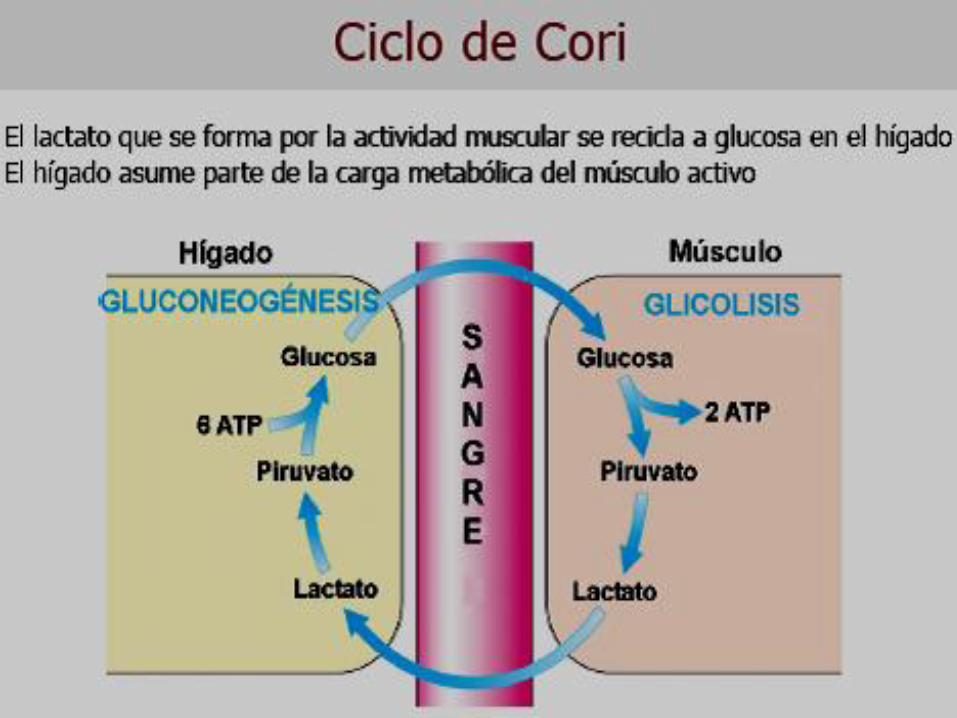
GLUCONEOGÉNESIS

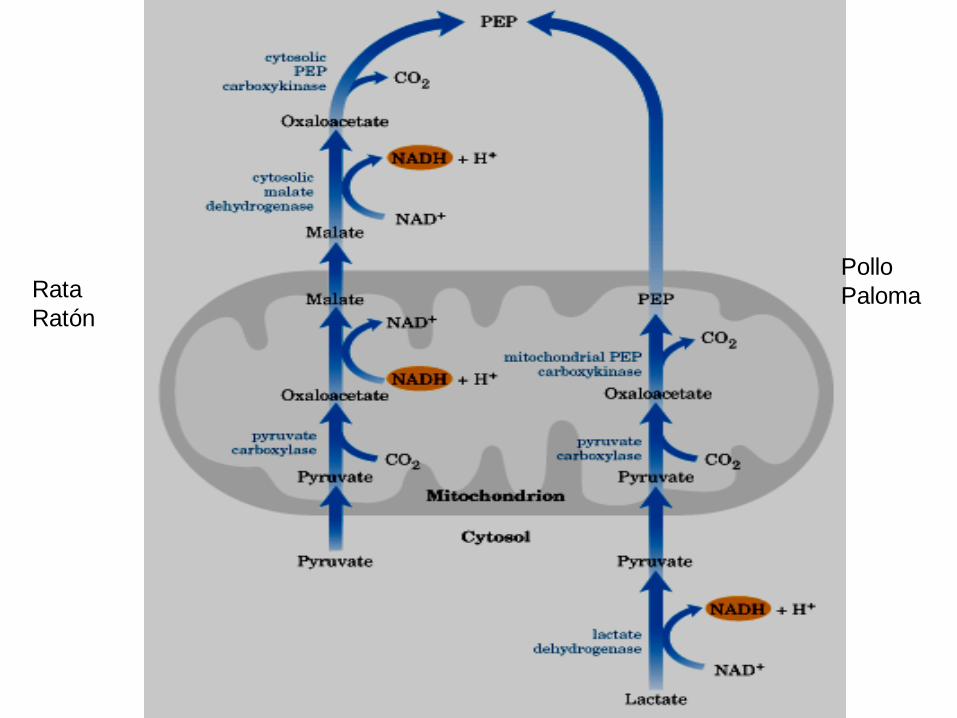
Pollo
PalomaRata
Ratón
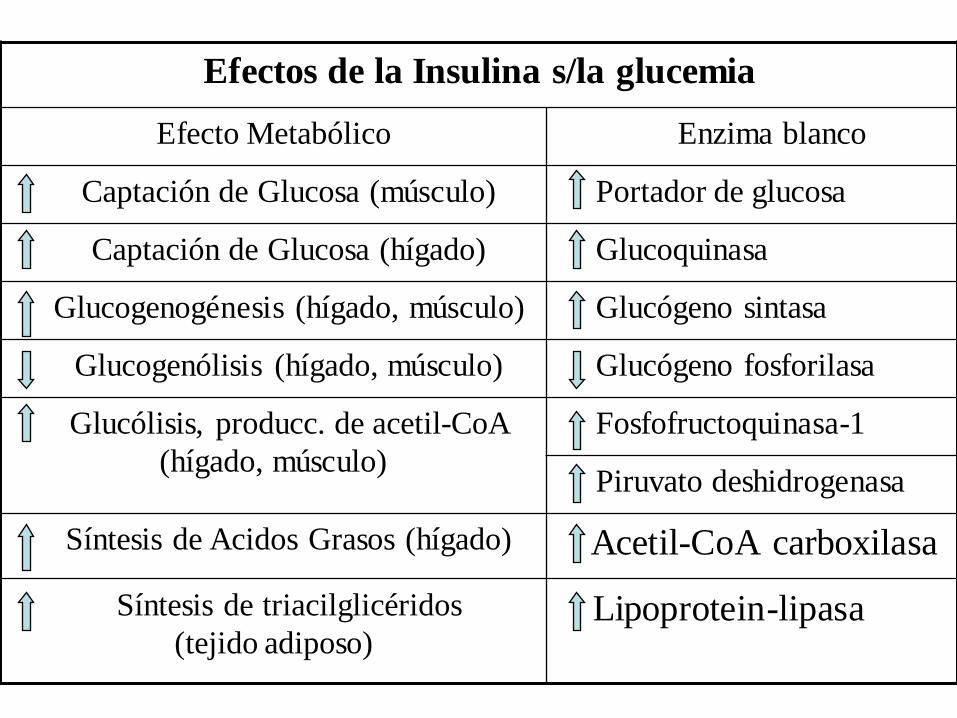
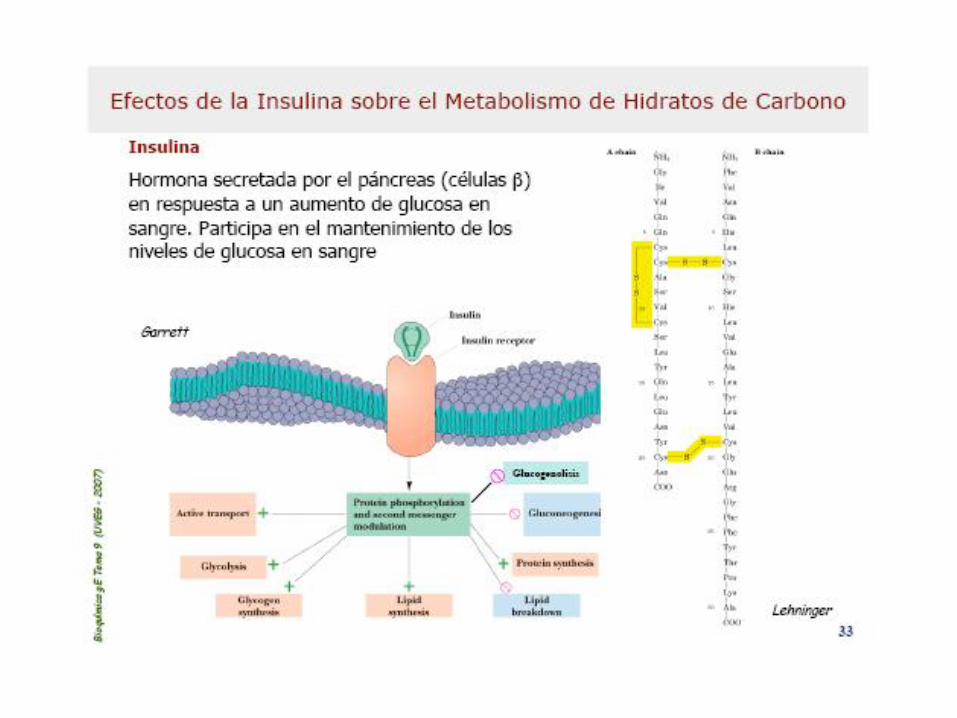
Efectos de la Insulina s/la glucemia
Efecto Metabólico Enzima blanco
Captación de Glucosa (músculo) Portador de glucosa
Captación de Glucosa (hígado) Glucoquinasa
Glucogenogénesis (hígado, músculo) Glucógeno sintasa
Glucogenólisis (hígado, músculo) Glucógeno fosforilasa
Glucólisis, producc. de acetil-CoA
(hígado, músculo)
Fosfofructoquinasa-1
Piruvato deshidrogenasa
Síntesis de Acidos Grasos (hígado) Acetil-CoA carboxilasa
Síntesis de triacilglicéridos
(tejido adiposo)Lipoprotein-lipasa

Derivacion de la glucolisis anaerobia
Ciclo de Rapaport-Luebering
1,3 DPG -> 2,3 DPG
2,3 DPG -> 3-PGlicerato
Catalizadas por una misma enzima, con doble funcion: MUTASA y
FOSFATASA
Importancia:
• 2,3 DPG regula la función hemoglobínica,
• y reservorio energético frente a situaciones como una disminución de pH intraeritrocitario c/merma en la glucólisis.

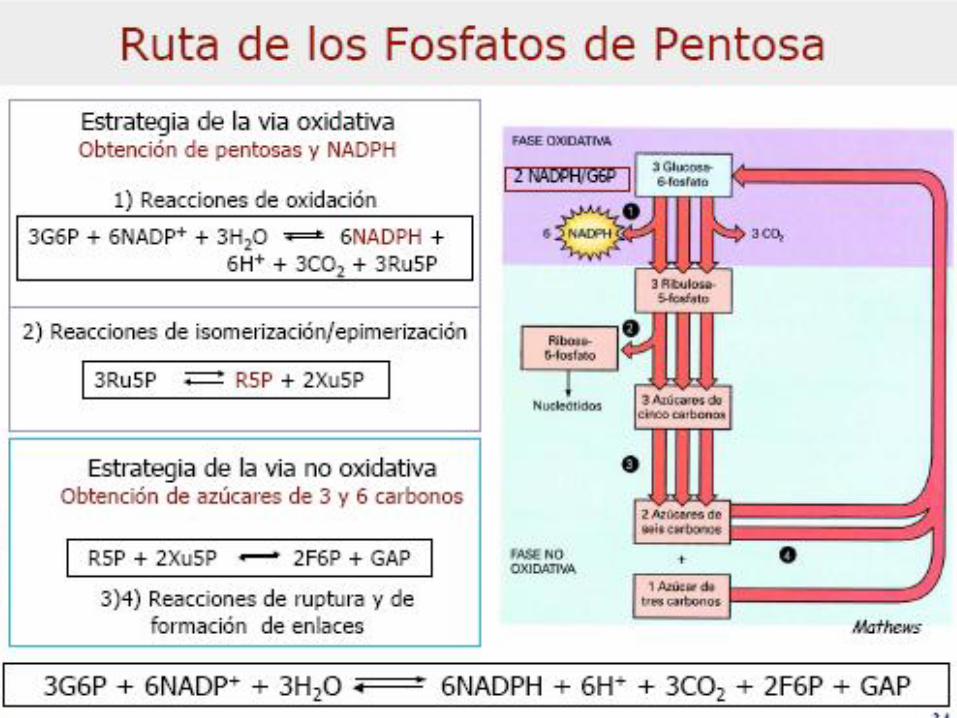
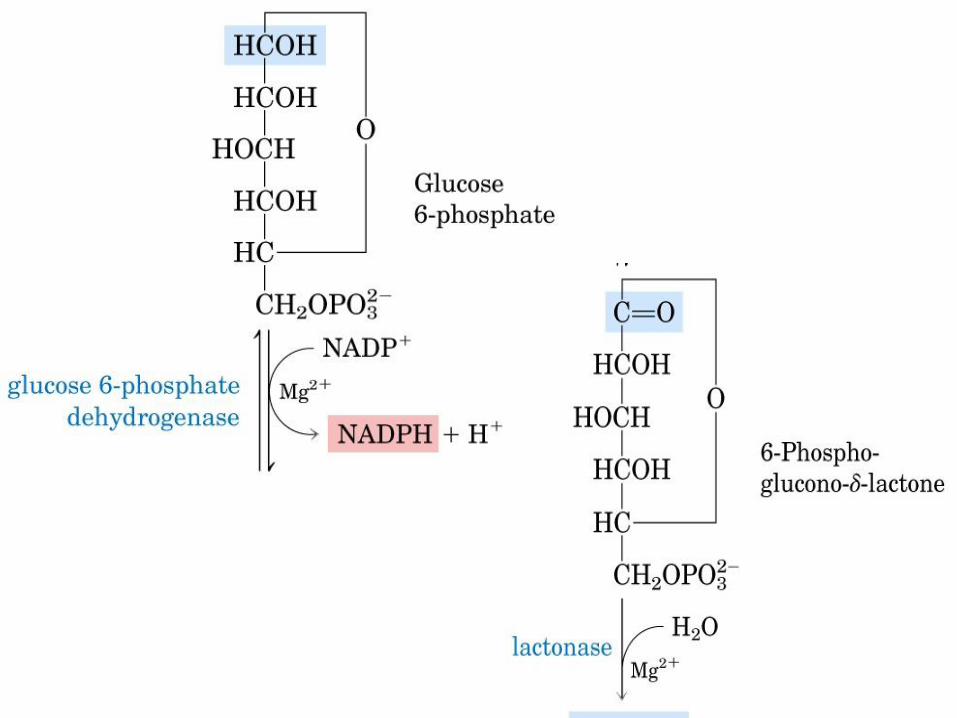
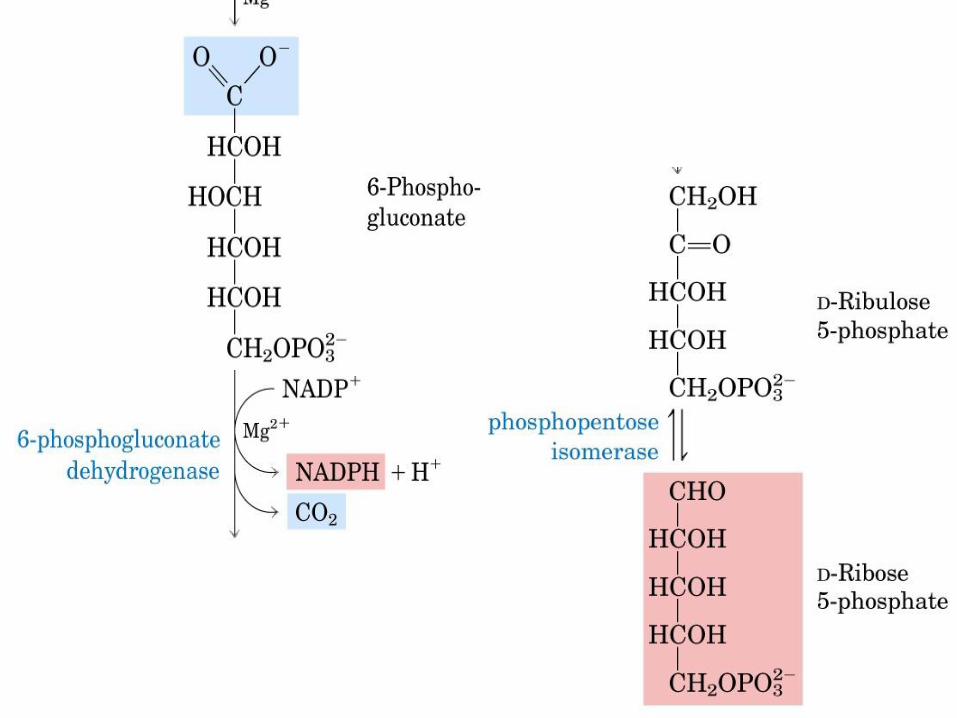
VIA DE LAS PENTOSAS
FOSFATO
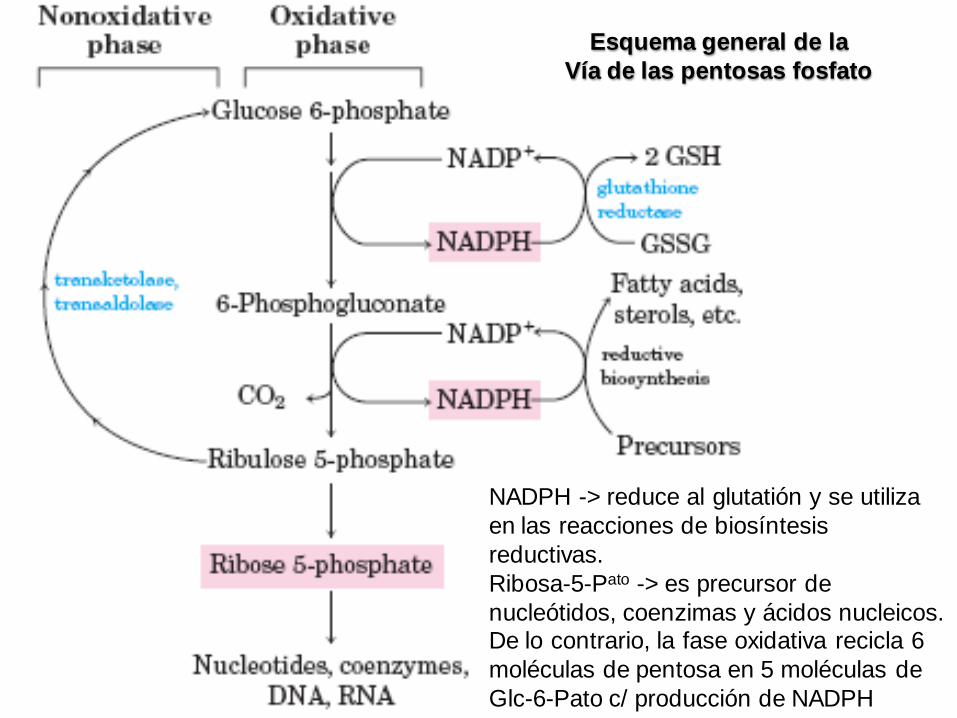
NADPH -> reduce al glutatión y se utiliza
en las reacciones de biosíntesis
reductivas.
Ribosa-5-Pato -> es precursor de
nucleótidos, coenzimas y ácidos nucleicos. De lo contrario, la fase oxidativa recicla 6
moléculas de pentosa en 5 moléculas de
Glc-6-Pato c/ producción de NADPH
Esquema general de la
Vía de las pentosas fosfato
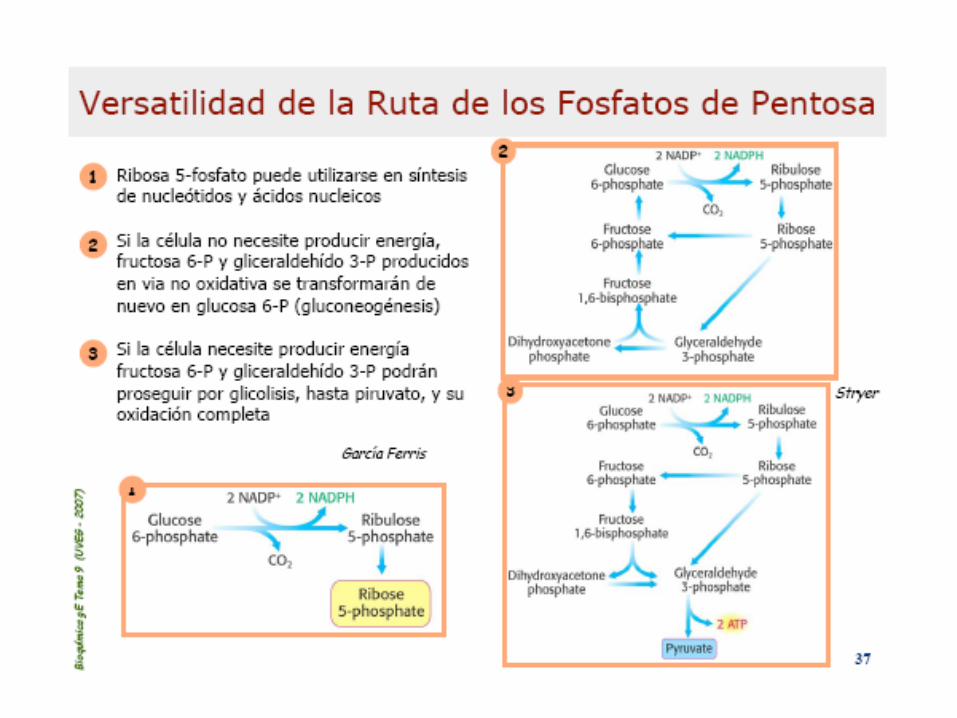
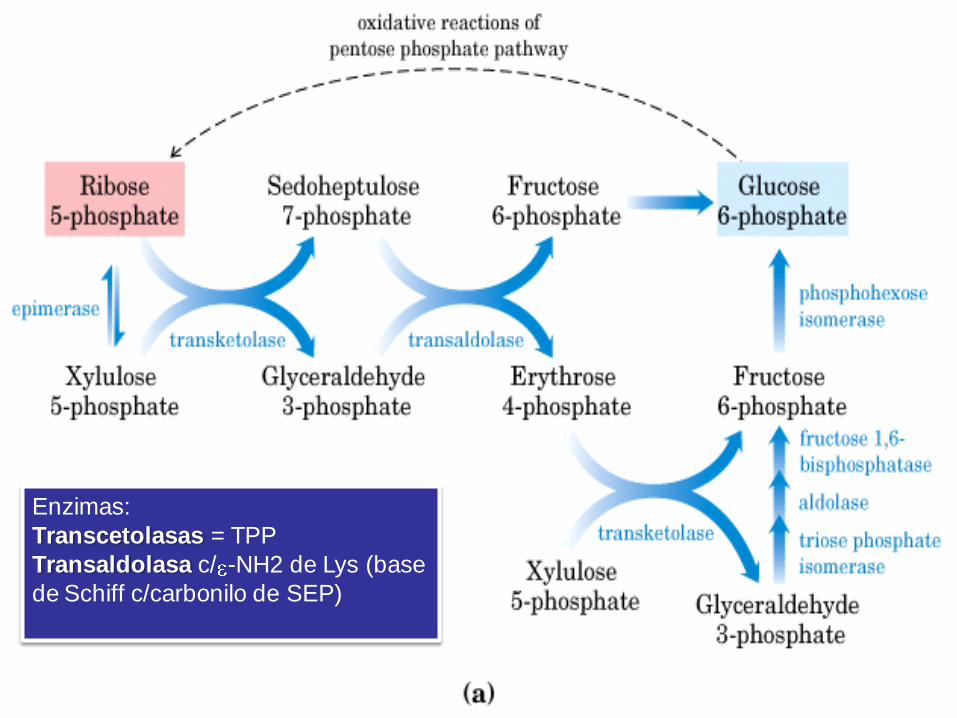
Enzimas:
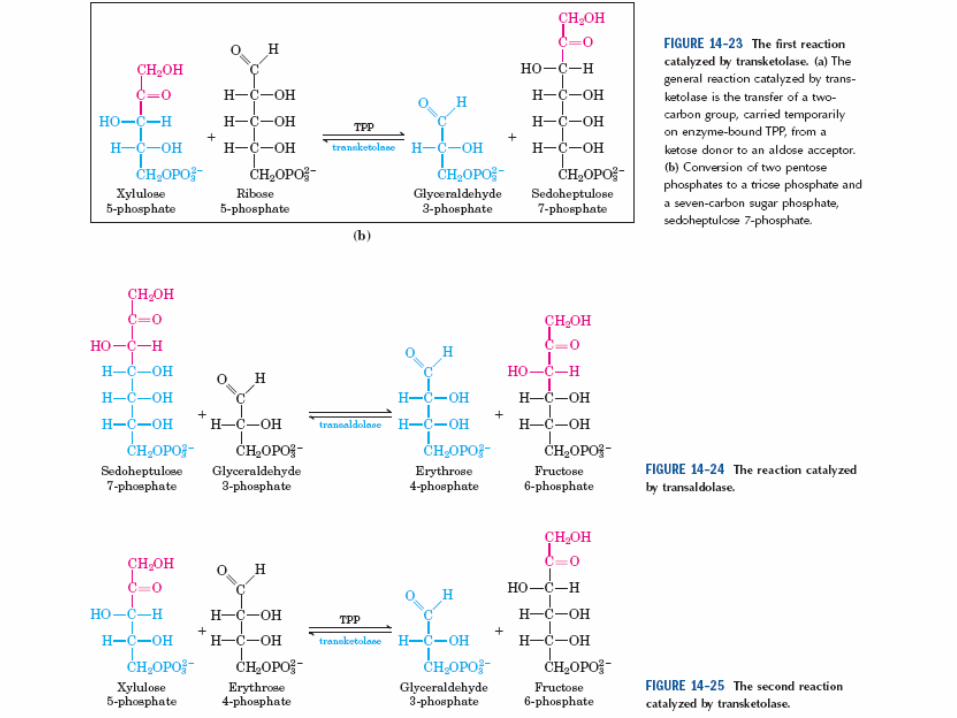
Transcetolasas = TPP
Transaldolasa c/ -NH2 de Lys (base
de Schiff c/carbonilo de SEP)
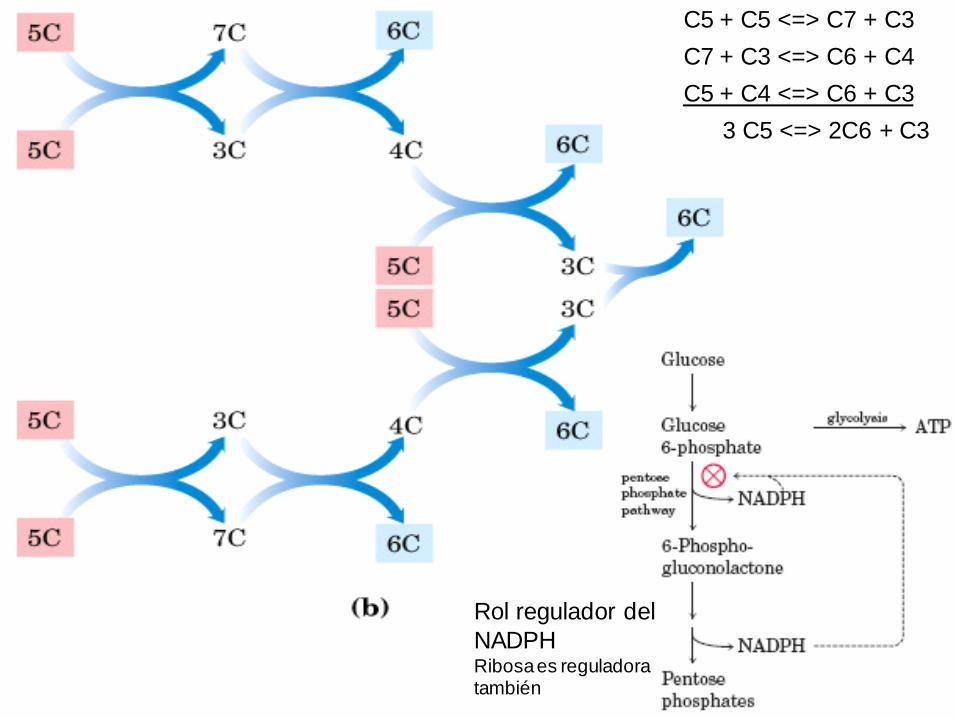
Rol regulador del
NADPHRibosa es reguladora también
C5 + C5 <=> C7 + C3
C7 + C3 <=> C6 + C4
C5 + C4 <=> C6 + C3
3 C5 <=> 2C6 + C3

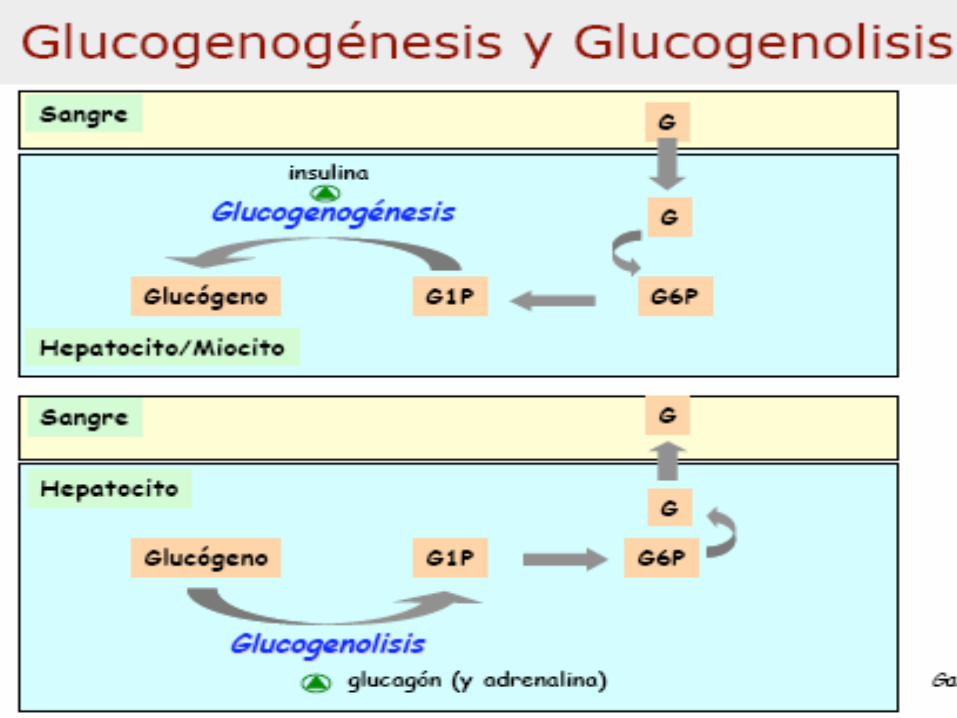
Metabolismo del Glucógeno

GLUCOGENOGÉNESIS
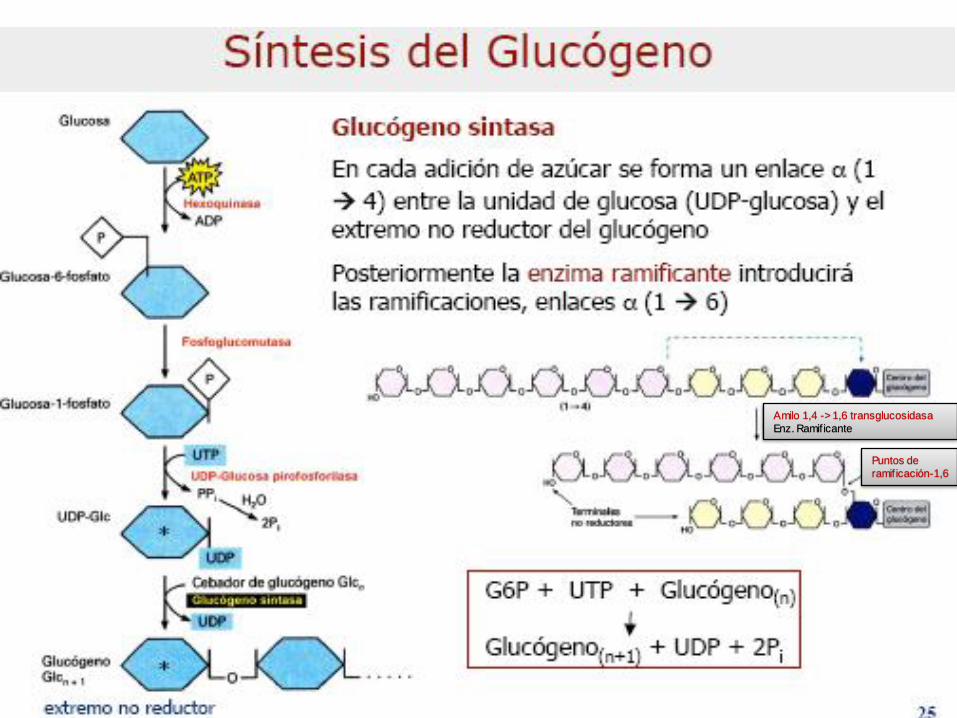
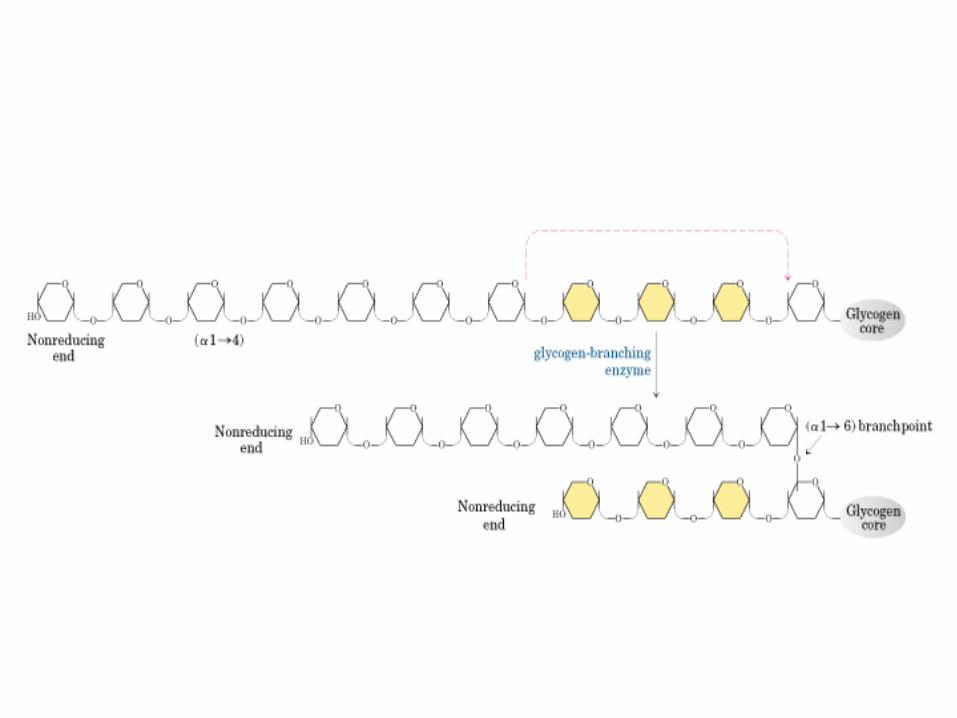
Amilo 1,4 -> 1,6 transglucosidasa
Enz. Ramificante
Puntos de
ramificación-1,6
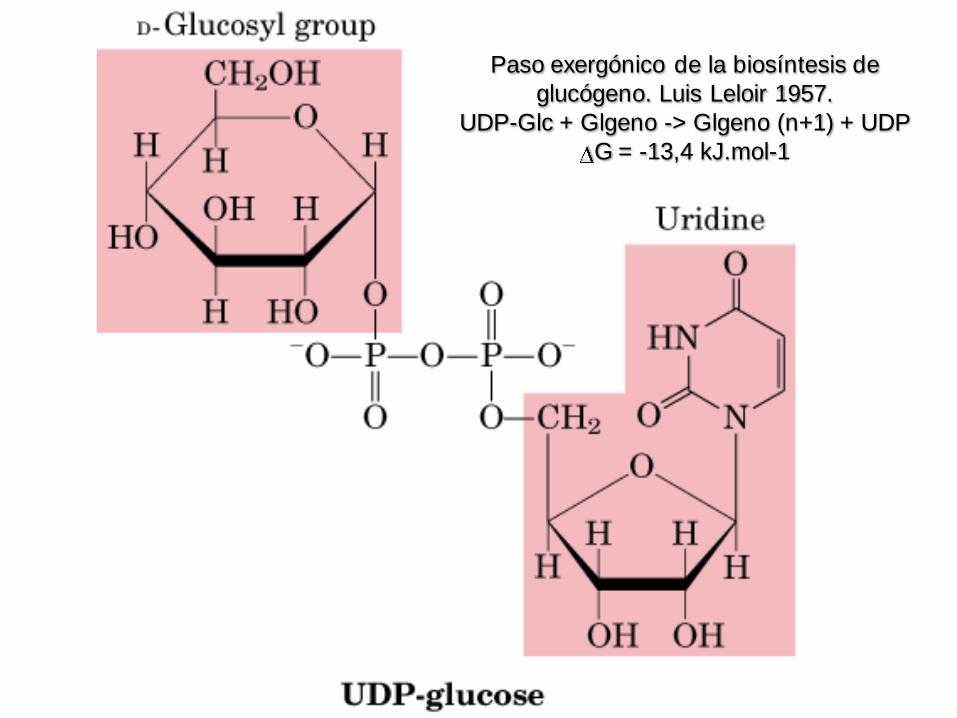
Paso exergónico de la biosíntesis de
glucógeno. Luis Leloir 1957.
UDP-Glc + Glgeno -> Glgeno (n+1) + UDP
G = -13,4 kJ.mol-1

Paso exergónico de la biosíntesis
de glucógeno. Luis Leloir 1957.
Donador de glucosilo (c/E de Pirofosforilasa inorgánica)
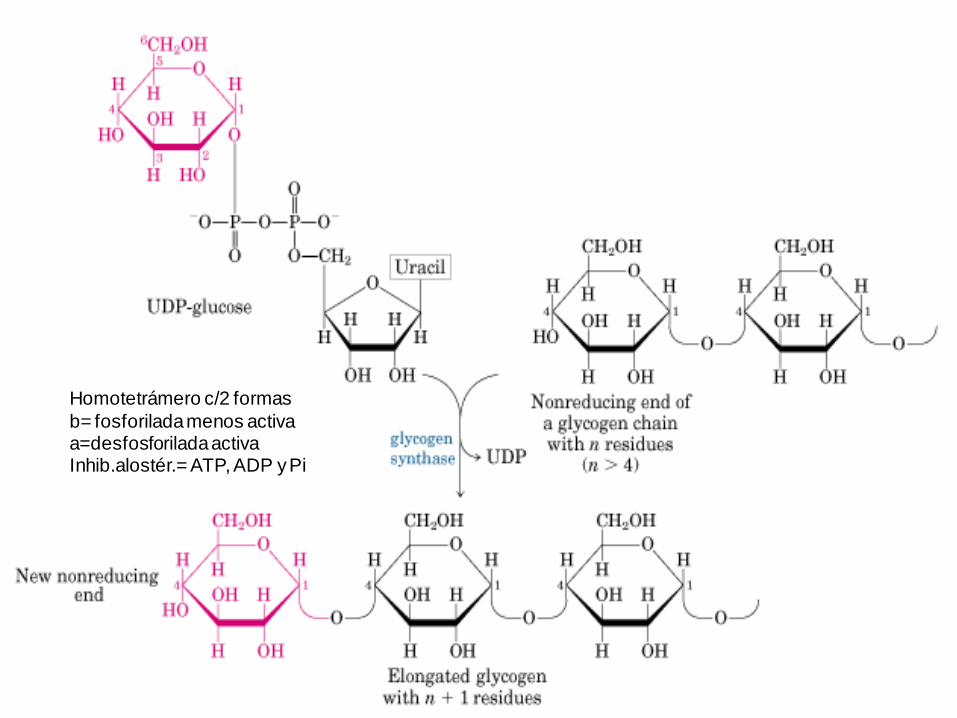
Homotetrámero c/2 formas
b= fosforilada menos activaa=desfosforilada activa Inhib.alostér.= ATP, ADP y Pi
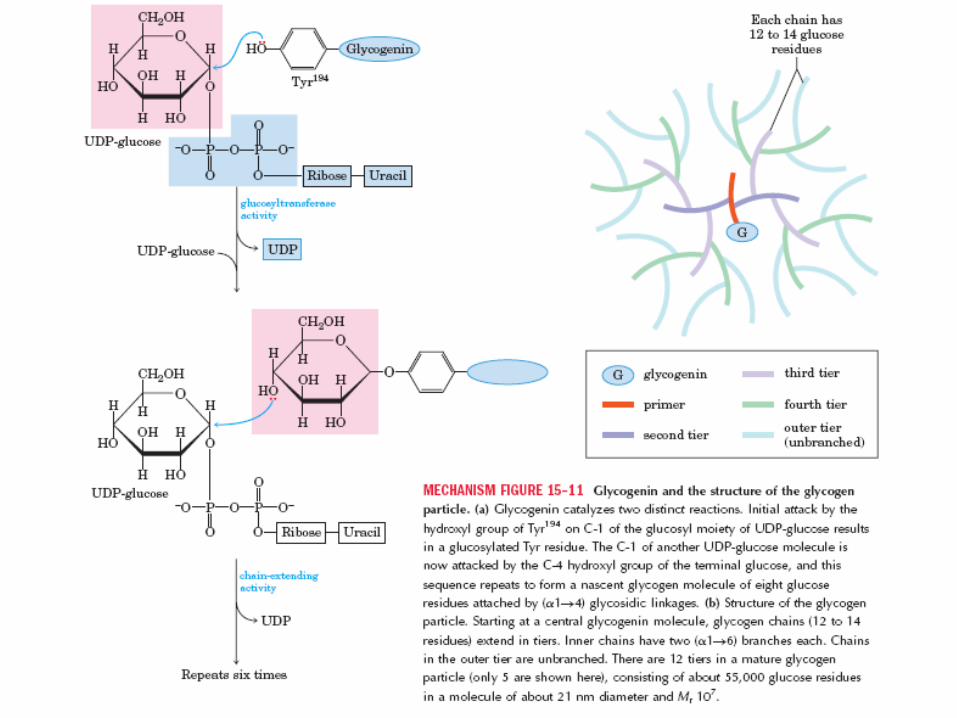

Glicogenina q’ cataliza 2 diferentes reacciones:
a)ataque inicial por el grupo –OH de la Tyr194 s/ C1 de
UDP-glucosa q’ resulta en un residuo Tyr glicosilado.
b)El C1 de otra UDP-Glc es atacado por el –OH del C4
de la Glc terminal. Secuencia que se repite 8 veces
formando enlaces 1-4 glicosídicos (moléc. de
glucógeno naciente)
Estructura de la partícula de glucógeno: comienza
por una molécula de glicogenina central, se extienden
cadenas de 12-14 residuos en etapas. De c/cadena
salen 2 ramas 1-6 glicosídicas. Hay 12 etapas de
formación en una partícula de glucógeno maduro,
alrededor de 55.000 residuos de Glc en 1 moléc. De
21 nm de diám.
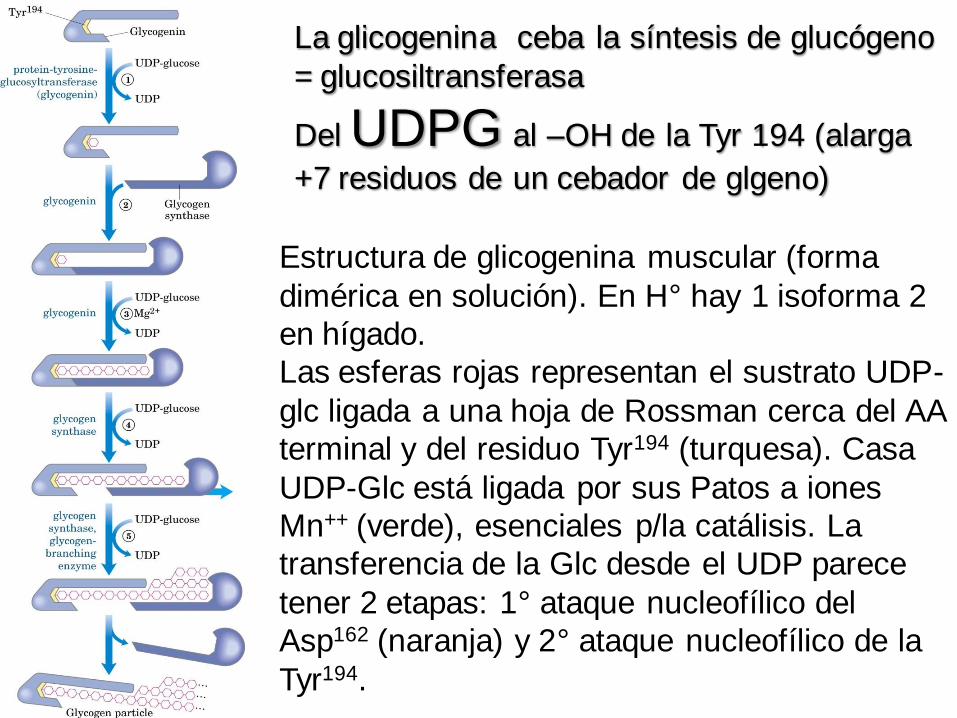
Estructura de glicogenina muscular (forma
dimérica en solución). En H° hay 1 isoforma 2
en hígado.
Las esferas rojas representan el sustrato UDP-
glc ligada a una hoja de Rossman cerca del AA
terminal y del residuo Tyr194 (turquesa). Casa
UDP-Glc está ligada por sus Patos a iones
Mn++ (verde), esenciales p/la catálisis. La
transferencia de la Glc desde el UDP parece
tener 2 etapas: 1° ataque nucleofílico del
Asp162 (naranja) y 2° ataque nucleofílico de la
Tyr194.
La glicogenina ceba la síntesis de glucógeno
= glucosiltransferasa
Del UDPG al –OH de la Tyr 194 (alarga
+7 residuos de un cebador de glgeno)
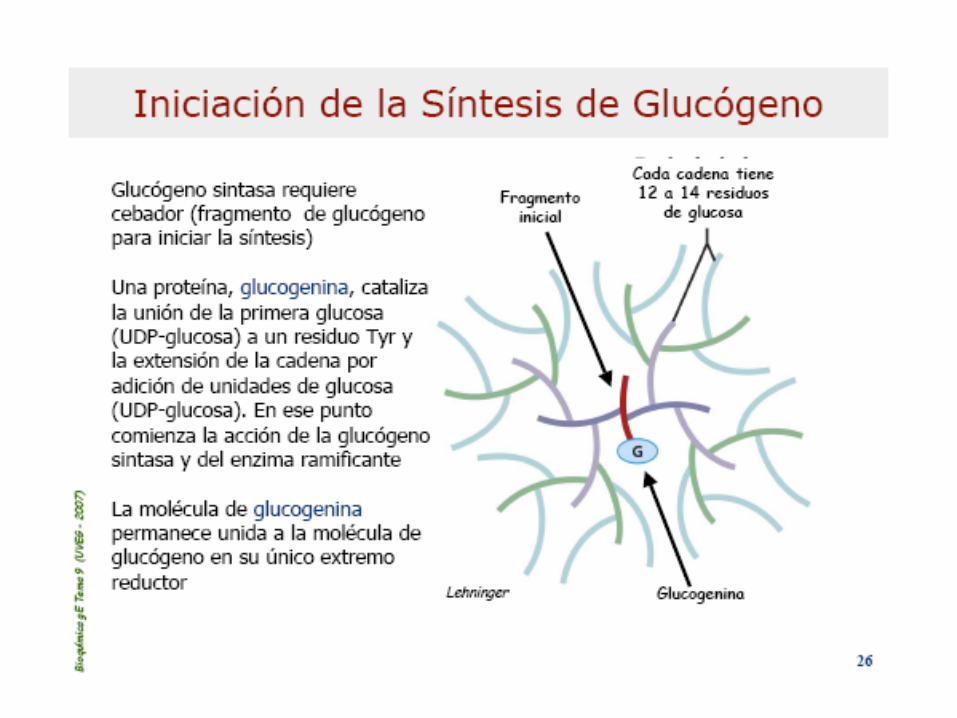

GLUCOGENOLISIS
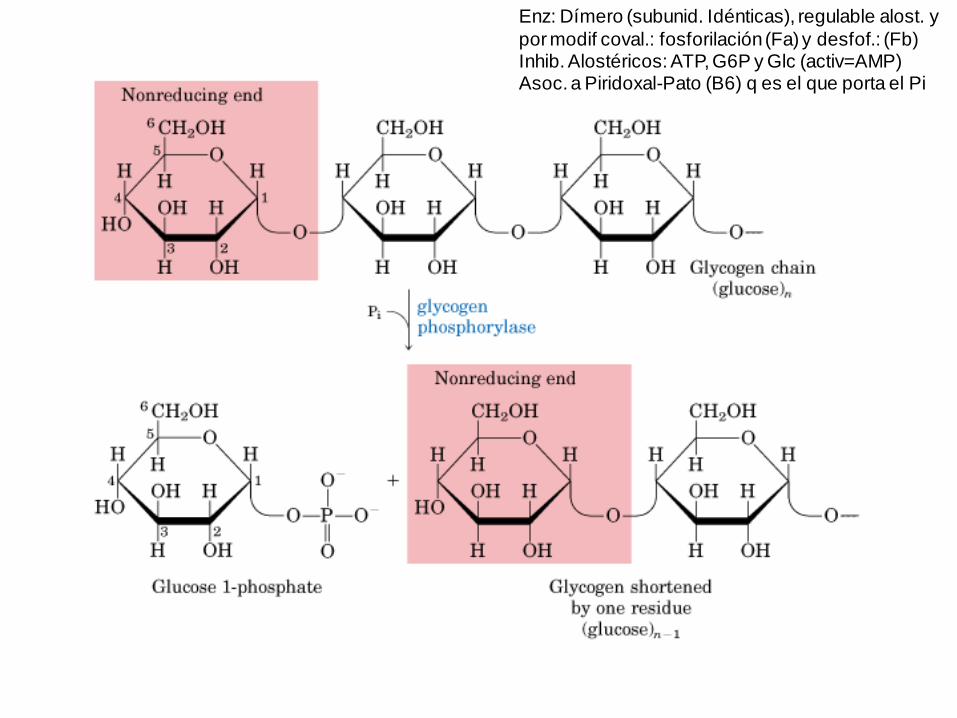
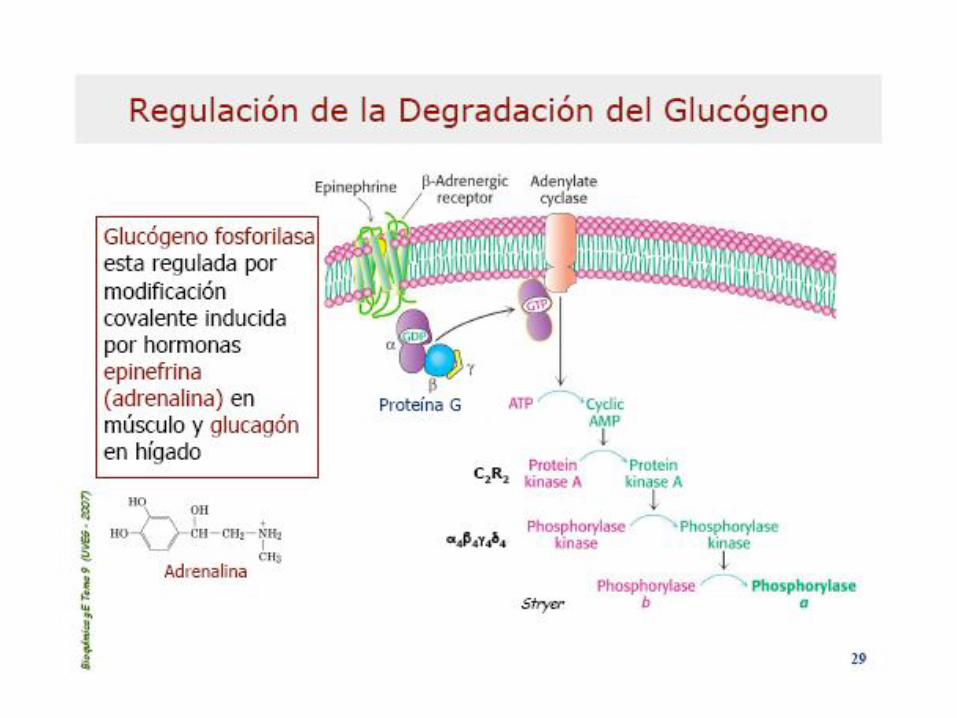
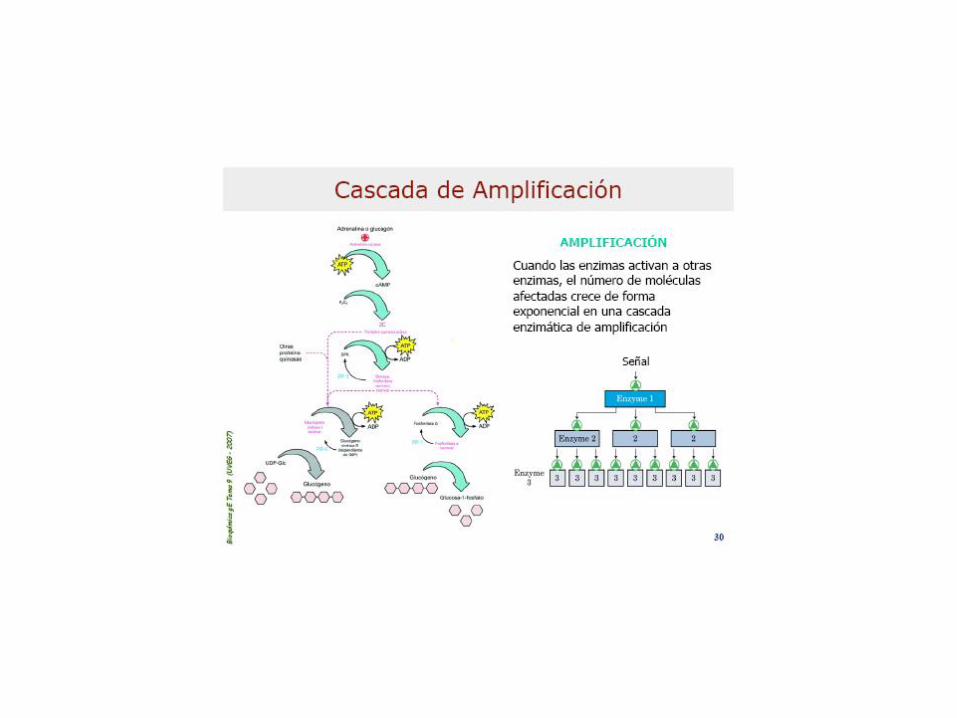
Enz: Dímero (subunid. Idénticas), regulable alost. y
por modif coval.: fosforilación (Fa) y desfof.: (Fb)Inhib. Alostéricos: ATP, G6P y Glc (activ=AMP)Asoc. a Piridoxal-Pato (B6) q es el que porta el Pi
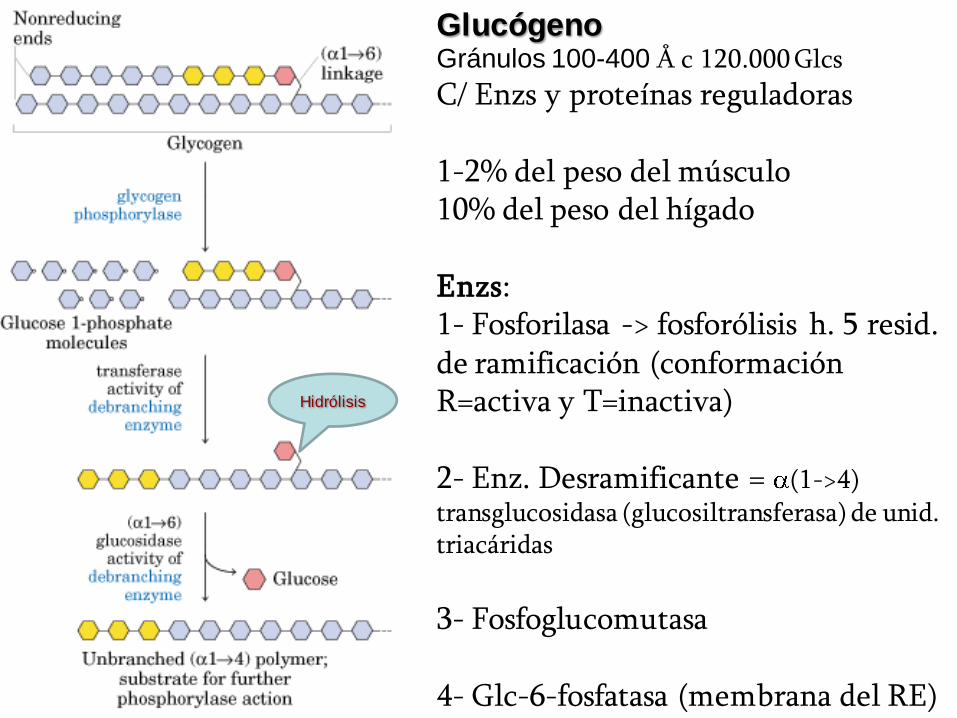
GlucógenoGránulos 100-400 Å c 120.000 Glcs
C/ Enzs y proteínas reguladoras
1-2% del peso del músculo10% del peso del hígado
Enzs:1- Fosforilasa -> fosforólisis h. 5 resid. de ramificación (conformación R=activa y T=inactiva)
2- Enz. Desramificante = (1->4) transglucosidasa (glucosiltransferasa) de unid. triacáridas
3- Fosfoglucomutasa
4- Glc-6-fosfatasa (membrana del RE)
Hidrólisis
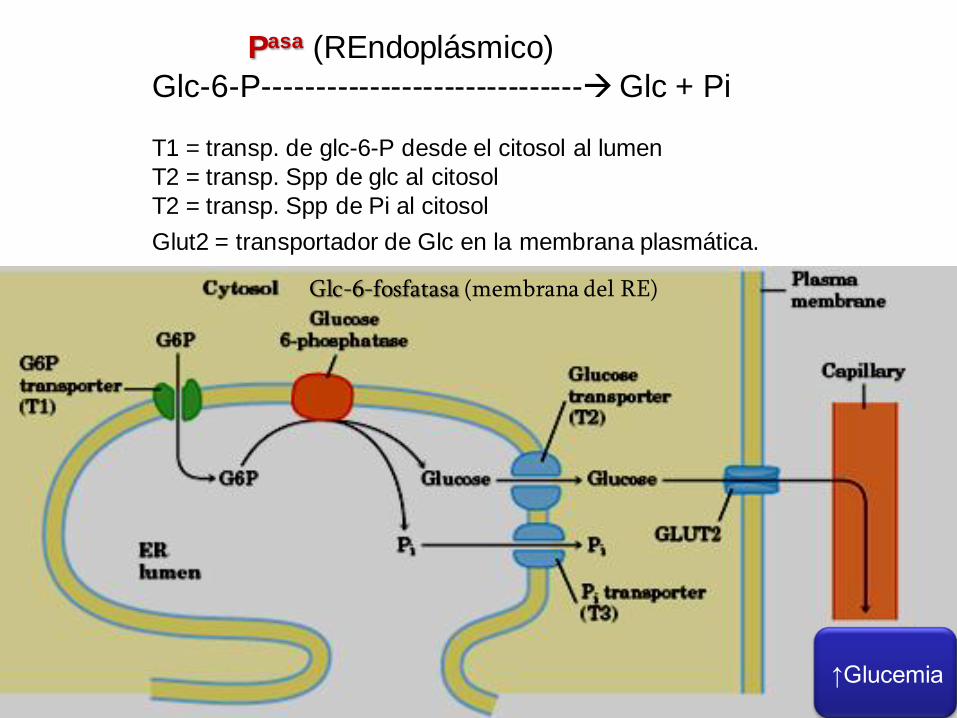
Pasa (REndoplásmico)
Glc-6-P------------------------------Glc + Pi
T1 = transp. de glc-6-P desde el citosol al lumen
T2 = transp. Spp de glc al citosol
T2 = transp. Spp de Pi al citosol
Glut2 = transportador de Glc en la membrana plasmática.
↑Glucemia
Glc-6-fosfatasa (membrana del RE)
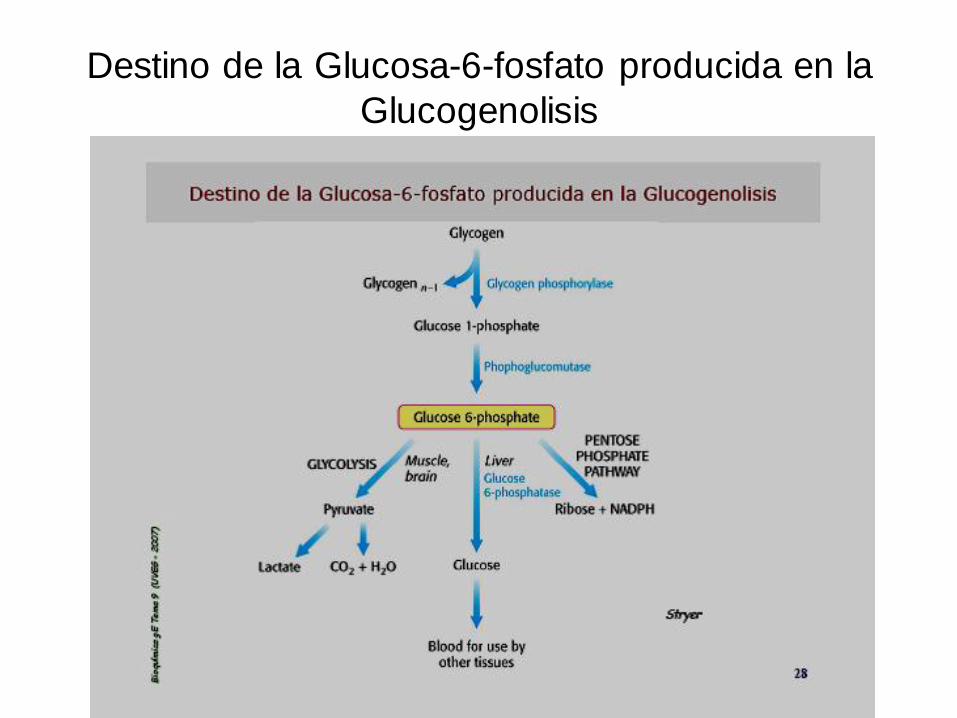
Destino de la Glucosa-6-fosfato producida en la
Glucogenolisis
Top Related