







Idiomas
Páginas
Jurídico


1
AGROCURSO CONFINAMENTO DE GADO
DE CORTE
AULA 6 – VITAMINAS, MINERAIS E
ADITIVOS
Juliano Ricardo Resende – Zootecnista, professor de nutrição de
ruminantes nas Faculdades Associadas de Uberaba

2
SUMÁRIO
1 - VITAMINAS PARA BOVINOS.......................................................................3
1.1 – Vitamina A................................................................................................4
1.2 – Vitamina D................................................................................................6
1.3 – Vitamina E................................................................................................7
1.4 – Niacina......................................................................................................8
2 - SUPLEMENTAÇÃO MINERAL PARA BOVINOS....................................11
2.1 - Importância dos nutrientes minerais....................................................11
2.2 - Sintomas da deficiência mineral ..........................................................12
2.3 - Adição de vermífugos e outros suplementos.....................................14
2.4 - Fornecimento de minerais ao gado......................................................14
3 – ADITIVOS.................................................................................................15
3.1 - Ionóforos.................................................................................................16
3.2 - Monensina sódica..................................................................................19
3.3 – Lasalocida sódica..................................................................................23
3.4 – Antibióticos não ionóforos...................................................................29
3.4.1 – Virginiamicina.....................................................................................31
3.5 - Promotores de Eficiência Alimentar Biológicos..................................35
3.5.1 - As leveduras ativas...........................................................................35
3.5.2 - Os probióticos bacterianos...............................................................37
4 – REFERENCIAS BIBLIOGRÁFICAS...........................................................41

3
1 - VITAMINAS PARA BOVINOS
As vitaminas são compostos orgânicos sintetizados por plantas
superiores e microrganismos, e fundamentais ao metabolismo celular. Os
animais requerem a presença das vitaminas para a síntese de muitas
coenzimas, que por sua vez são indispensáveis à ação das enzimas, sem as
quais não existiria a vida. Uma reação química não se processa a contento
sem a ação destes catalisadores biológicos. Os avanços em genética e
nutrição nos últimos anos têm permitido melhorias significativas na
performance e na saúde dos bovinos, e isto tem levado os nutricionistas a
reconsiderar a função de algumas vitaminas, tendo como princípio básico sua
função coenzimática.
As vitaminas são classificadas em dois grandes grupos a saber:
a - Lipossolúveis (A, D, E e K);
b - Hidrossolúveis (B1, B2, B6, B12, ácido pantotenico, fólico, biotina, colina e
niacina).
O Quadro Q apresenta as vitaminas de interesse zootécnico e seus sintomas
carênciais.
QUADRO 1 - Vitaminas, suas coenzimas, metabolismo e sintomas carenciais.
Vitaminas Coenzimas Metabolismo Sintomas carenciais
A Retinol Visão Epitélio
Cegueira, queratinização epitelial, reprodução e
saúde.
D Ergocalciferol(D2) Colecalciferol(D3)
Calcificação da matriz óssea
Raquitismo e osteomalácia.
E Tocoferol Antioxidante celular Reprodução, saúde.
Niacina NAD, NADP
Oxireduções biológicas
e transporte de hidrogênio.
Metabolismo protéico e energético.
NAD- Flavina adenina dinucleotídeo e NADP- Nicotinamida-adenina
dinucleotídeo fosfato
Fonte: BACILA (1980)

4
Ao iniciar uma discussão sobre vitaminas, deve-se considerar a
existência de muitos fatores que interferem em seu requerimento e seus
sintomas de deficiência a saber:
- doenças respiratórias e infecciosas, estresse;
- transtornos metabólicos que diminuem síntese microbiana;
- processamento dos alimentos (calor, fermentação);
- fertilização das pastagens (excesso de nitrogênio, oxálico,nitratos);
- estado fisiológico dos animais.
1.1 - Vitamina A
Esta vitamina é denominada de "vitamina do epitélio", tendo em vista
sua função primordial para a formação e manutenção da integridade da
mucosa.
A vitamina A é, desta forma, essencial para a maior resistência às
doenças e fertilidade dos ruminantes. A vitamina A não ocorre em sua forma
natural e sim em seus precursores, os carotenos. Durante muito tempo,
acreditou-se que todo pigmento carotenóide do alimento era transformado em
vitamina A. Na verdade, reconhece-se o valor do B-caroteno como potencial
transformador principal, e que o bovino tem a capacidade de sua conversão de
24% (1mg de beta-caroteno equivale a 400 UI de vitamina A). Estes compostos
com atividade vitamínica A são muito sensíveis a oxidação, onde a presença
da luz, temperatura, unidade e íons metálicos aceleram sua decomposição.
Nas forrageiras, as maiores perdas ocorrem durante o processo de
conservação (ensilagem e fenação). No período da seca, as forragens são
bastante deficientes em vitamina A.
Os sintomas carenciais da vitamina A podem incluir a redução do
apetite, menor ritmo de crescimento, pelagem áspera, lacrimejação, baixa taxa
de concepção, natimortos, abortos, retenção de placenta, cistos ovarianos,
anormalidades do sêmen, a libido reduzida e diarréia.
A maior necessidade de vitamina A ocorre durante a época da cobertura
e do parto. Bezerros nascidos de vacas deficientes (final de gestação
coincidente com longo período de forragem seca), estão sujeitos à infeções

5
diversas (quando não natimortos), sendo comum a diarréia com
lacrimejamento, descarga nasal e algumas vezes incoordenação motora. A
causa é a deficiência da mãe, bem como a baixa ingestão de colostro e de
leite. Outro fator condicionante é a deficiência de zinco e de proteínas, onde
são necessários para a mobilização da vitamina A do fígado (órgão de
armazenamento), de acordo com Mc. Dowell (1989). Os Quadros 2 e 3 ilustram
o efeito da suplementação da vitamina A em bovinos.
QUADRO 2 - Efeito da administração de vitamina A sobre a taxa de
natalidade (%) de bovinos.
Categoria 1- Controle 2- Suplementado
Vacas 70,10 84,50
Primíparas 64,50 79,30
Fonte: BRADFIELD e BEHRENS (1968)
Os mesmos autores relataram melhoria na taxa de natalidade, com
suplementações do complexo ADE, entre 14 e 30 dias anteriores, à estação de
monta de vacas, cujos dados são apresentados no quadro 3.
QUADRO 3 - Efeito da suplementação de ADE sobre a taxa de natalidade
(%).
Ano Controle ADE
1964 77,40 88,00
1965 80,00 92,20
1966 54,80 74,40
1967 71,20 82,20
Média 70,80 84,20
Fonte: BRADFIELD e BEHRENS (1968)
Outro trabalho de Mechan et al. (1970), ilustra o efeito da vitamina A
sobre a taxa de mortalidade de bezerros, apresentado no Quadro 4
.

6
QUADRO 4 - Efeito da suplementação de vitamina A sobre a taxa de
mortalidade em bezerros.
Tratamento Natimortos(%) Mortalidade pós-natal(%)
Controle 8,80 16,50
Vitamina A 6,60 8,10
Alguns trabalhos demonstram o efeito da própria suplementação do
betacaroteno sobre parâmetros reprodutivos em vacas leiteiras (incidência de
cios silenciosos, atraso na ovulação, cistos ovarianos). Ainda que o
requerimento não esteja definido, os dados sugerem suplementação de 150 a
300 mg/dia de betacaroteno, no período compreendido entre 15 dias antes do
parto e a confirmação da gestação, para rebanhos leiteiros de alta produção,
onde o volumoso seja baseado em silagens.
De acordo com os estudos sobre a vitamina A, pode-se sugerir a sua
suplementação para ruminantes em algumas situações:
a - Gado de corte ao iniciar o período da seca (maio-junho), principalmente
para animais jovens, novilhas a serem entouradas (outono ou primavera),
novilhas e vacas em gestação e touros.
b - Gado de corte em confinamento.
c - Rebanho leiteiro, para qual o volumoso é constituído por silagens.
1.2 - Vitamina D
Esta vitamina é denominada "anti-raquítica", tendo em vista que sua
função principal seja ligada ao metabolismo do cálcio (Ca) e do fósforo (P) -
transporte e mobilização - e quando em deficiência causa redução drástica no
ritmo de crescimento dos animais novos. O requerimento em vitamina D
depende de uma boa relação Ca/P, na qual relações estreitas ou muito largas
irão interferir na sua exigência. Talvez este seja o motivo pelo efeito positivo do
uso de altas dosagens de vitamina D na prevenção da febre do leite. Trabalhos
mostram a vantagem de sua utilização cerca de cinco dias antes do parto e um
dia após o mesmo, em vacas predisponentes a este distúrbio metabólico
(HIBBS e CONRAD, 1966; VINET et al, 1985), no qual a dosagem é de
500.000 UI diariamente. Honett et al (1992) mostram redução de incidência da

7
febre do leite (38 x 8%) em vacas secas (15 dias antes do parto), em dietas de
silagens de alfafa, feno e concentrado.A vitamina D é encontrada na forma de
ergocalciferol (D2) de origem vegetal e na forma de colecalciferol (D3) de
origem animal. A ação da luz solar (raios ultravioleta) sobre a vitamina D (7-
dehidrocolesterol) encontrado na pele dos animais, produz a vitamina D3.
Diante deste efeito, normalmente os técnicos não recomendam a
suplementação desta vitamina para animais em pastejo, em regiões tropicais.
No entanto, algumas considerações são necessárias:
- o efeito da irradiação deve acontecer nas partes do corpo desprovidas, ou
com pouco pêlo;
- os ruminantes dotados de pelame espesso recebem pouca irradiação através
da pele;
- uma irradiação muito intensa resulta em substâncias sem atividade vitamínica
(supra-esteróis) e substâncias tóxicas ao organismo;
- a vitamina D2 somente é encontrada em fenos curados ao sol, desde que o
tempo de cura seja relativamente curto (inferior a 36 horas);
- forragens verdes e silagens possuem pouca quantidade de vitamina D;
- o organismo animal tem baixa capacidade de armazenamento.
Independentes destas considerações, os conhecimentos atuais mostram
ser incomum a deficiência de vitamina D em bovinos em pastejo.
1.3 - Vitamina E
Essa vitamina tem diferentes funções no organismo celular, ou seja, atua
na prevenção da oxidação dos ácidos graxos insaturados dentro da célula,
protegendo assim a ruptura da membrana. Se ocorre a oxidação dos lipídios
(formação de peróxidos) há destruição da integridade da membrana celular,
com sérios danos metabólicos. A distrofia nutricional que acomete bezerros e
cordeiros é um exemplo deste dano causado pela deficiência do selênio, mas
influenciada pelo status vitamínico E. Assim, em dietas deficientes em selênio,
a distrofia não é prevenida pela vitamina E, ou é prevenida pelo selênio, se
houver vitamina E, mostrando a interação dos dois nutrientes (ISLABÃO.
1978).

8
O papel da vitamina E na reprodução não está perfeitamente elucidado.
No entanto, sabe-se que as glândulas endócrinas possuem alto nível desta
vitamina, particularmente a hipófise. Trabalhos mostram que a vitamina E
promove a liberação do FSH (hormônio folículo estimulante), do ACTH
(hormônio adrenocorcotrófico) e do LH (hormônio luteinizante). Problema muito
comum em rebanhos leiteiros, a retenção de placenta, está relacionado à
deficiência de vitamina E. No entanto, deve-se considerar sua relação também
com deficiência de selênio, do fósforo da vitamina A, bem como de agentes
infecciosos. Jullien et al (1976), relatam a redução da retenção de placenta em
gado leiteiro com a suplementação de 680 mg/dia de vitamina E e de 60 mg/dia
de selenito de sódio, durante 21 dias antes do parto. Outro problema verificado
em rebanhos leiteiros, a mastite, mostra-se ser auxiliada em seu controle pela
suplementação em vitamina E. No entanto, os trabalhos científicos sobre o
assunto não são conclusivos.
Quanto aos bovinos de corte, principalmente em regime de
confinamento, a vitamina E está relacionada à qualidade da carne, mas
lamentavelmente o aspecto econômico ainda é um entrave, não compensando
a suplementação. Pesquisas da Universidade de Wisconsin (WILLIANS, 1993),
ilustram efeito positivo da suplementação sobre a qualidade da carne, onde o
grupo tratado recebeu, diariamente, 350 UI, durante um período de 30 dias. O
trabalho demonstrou que após 12 dias da exposição da carne ao varejo, esta
conservou sua cor "vermelho vivo". Além disto, houve retardamento na
formação da metamioglobina (cor escura), em relação ao grupo não
suplementado.
Como conclusão, ainda não é vantajosa economicamente sua
suplementação como melhoradora da qualidade da carne.
1.4 – Niacina
Esta é uma vitamina do complexo B, bastante pesquisada nos últimos
anos. Considerava-se que a síntese de niacina no rúmen era adequada para o
requerimento do ruminante. No entanto, face aos progressos obtidos na
genética animal e sua correspondente melhoria na performance, estudos
recentes tem mostrado resposta positiva com a suplementação, para animais

9
de alta produção, como é o caso de vacas leiteiras estabuladas e garrotes
precoces, quer seja oriundos de cruzamentos industriais ou mesmo de
zebuinos puros, mas de alta performance.
A niacina participa como componente da coenzima (NAD e NADP), de
vários sistemas enzimáticos, de grande importância no metabolismo do
ruminante, entre os quais pode-se citar:
- metabolismo dos carboidratos;
- metabolismo dos lipídeos;
- metabolismo dos aminoácidos.
O efeito da niacina no metabolismo está no aumento da síntese da
proteína microbiana, na digestibilidade da celulose e dos ácidos graxos
voláteis, principalmente o ácido propiônico. No caso dos bovinos de corte em
confinamento, sugere-se a suplementação quando a taxa de ganho for superior
a 1,50 Kg/dia, principalmente na fase de adaptação do gado particularmente
nos primeiros 21 dias do encerramento desses animais.
QUADRO 5 - Efeitos da niacina sobre a performance de bovinos de corte
em confinamento (adaptação) , em dietas com uréia.
ITENS CONTROLE NIACINA (70 mg/dia)
Peso inicial (Kg) 198,60 193,30
Peso final (Kg) 216,00 219,00
Ganho diário (g/dia) 600 690
Conversão (Kg MS/Kg ganho) 8.98 6,41
Fonte: BIERS (1981)
Concluindo, sugere-se os seguintes níveis de suplementação de
vitaminas para os bovinos de corte:

10
TABELA 1 - Níveis vitamínicos sugeridos para ruminantes.
Animal Vit A Vit D
UI/Kg MS Vit E
Niacina mg/Kg MS
1-Bovinos de corte
a-Bezerros 2200 275 30 -
b-Novilhas/vacas gestação 2800 275 30 -
c-Lactação e touros 3900 275 60 -
d-Confinamento 2200 275 60 60
Fonte: N.R.C ( 1996)

11
2 - SUPLEMENTAÇÃO MINERAL PARA BOVINOS
2.1 - Importância dos nutrientes minerais
Uma das mais importantes limitações nutricionais do gado bovino nas
regiões tropicais é a deficiência de minerais, uma vez que as forrageiras,
geralmente, não atendem as exigências dos animais. O conteúdo de mineral da
forragem depende de vários fatores, como solo, clima e espécie forrageira e
sua maturidade.
A maioria dos solos brasileiros é de média a baixa fertilidade, com
elevada quantidade de alumínio (Al) e de ferro (Fe), favorecendo a formação de
compostos insolúveis para a planta e exacerbando a deficiência do P. A
reposição dos nutrientes exportados pelos produtos animais ao solo, por
intermédio da adubação é pouco comum na maioria das regiões, o que
ocasiona um decréscimo gradativo do conteúdo de minerais na pastagem.
A suplementação mineral na pequena e média produção é
extremamente precária, principalmente por falta de informação (VEIGA et al.,
1996). A correção das deficiências minerais, pela suplementação no cocho, à
vontade, é bastante eficiente.
Constatou-se em estudos que o custo da mistura mineral no mercado
influencia a escolha do produto a ser utilizado. Além disso, a maioria dos
criadores desconhece os fundamentos básicos da nutrição mineral,
especialmente relacionados à suplementação do cálcio, elemento crítico na
alimentação de vacas leiteiras (MANESCHY, 2002).
Embora compondo apenas cerca de 5% do corpo de um animal, os
nutrientes minerais contribuem com grande parte do esqueleto (80% a 85%) e
compõem a estrutura dos músculos, sendo indispensáveis ao bom
funcionamento do organismo (MCDOWELL, 1992). Os desequilíbrios dos
minerais na dieta animal podem ocorrer tanto pela deficiência como pelo
excesso.

12
2.2 - Sintomas da deficiência mineral
Como se trata de um grande número de elementos que desempenham
as mais variadas e complexas funções no organismo, os sintomas causados
pelos desequilíbrios minerais da dieta não são específicos. Esses sintomas
podem ser confundidos com aqueles causados por deficiência de energia e
proteína (alimentação deficiente qualitativa e quantitativamente) ou por
problemas de saúde (parasitismo, doenças infecciosas ou ingestão de plantas
tóxicas).
Os principais sintomas gerais que indicam a ocorrência de deficiências
minerais no rebanho são, conforme Veiga et al. (1996) e Veiga e Lau (1998):
Apetite depravado - Os animais comem terra, pano e plástico; roem e ingerem
ossos, madeira e casca de árvores; lambem uns aos outros; apresentam
avidez por sal de cozinha.
Redução do apetite - Mesmo em pastagens com plena disponibilidade de
forragem e de boa qualidade, os animais apresentam baixo consumo,
mostrando o ventre sempre vazio (afundado).
Aspecto fraco ou doentio - Os animais ficam magros, com dorso arqueado,
pêlos arrepiados e sem brilho, lesões na pele e dificuldade de locomoção.
Anomalias dos ossos - Os ossos longos se tornam curvos e as extremidades
dilatadas.
Fraturas espontâneas - Freqüentemente, ocorrem quebraduras ósseas,
sobretudo quando os animais são manejados, evidenciando fraqueza do
esqueleto.
Anomalias da pele - Despigmentação e perda de pêlo, e desordem da pele,
como ressecamento e descamação.
13
Baixo crescimento e produtividade - O crescimento dos animais jovens é
retardado, o ganho de peso é baixo ou negativo (perda de peso) e a produção
leiteira é prejudicada.
Baixa fertilidade - Rebanhos com carência mineral apresentam uma reduzida
fertilidade das vacas, em face da ocorrência de cios irregulares ou ausentes,
abortamento e retenção placentária, resultando em baixa produção de
bezerros.
Baixa resistência a doenças - Animais deficientes em minerais são menos
resistentes (mais susceptíveis) a doenças e se ressentem mais dos ataques de
parasitas internos (vermes).
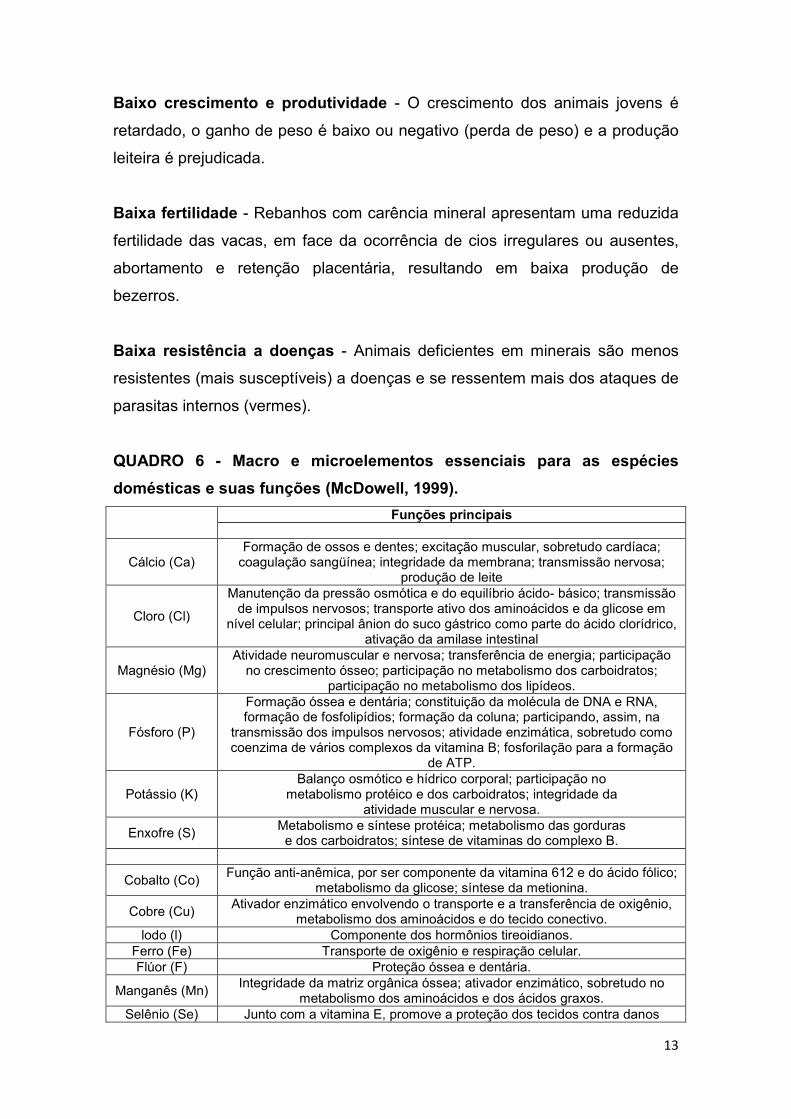
QUADRO 6 - Macro e microelementos essenciais para as espécies
domésticas e suas funções (McDowell, 1999).
Minerais Funções principais MACROELEMENTOS
Cálcio (Ca) Formação de ossos e dentes; excitação muscular, sobretudo cardíaca; coagulação sangüínea; integridade da membrana; transmissão nervosa;
produção de leite
Cloro (Cl)
Manutenção da pressão osmótica e do equilíbrio ácido- básico; transmissão de impulsos nervosos; transporte ativo dos aminoácidos e da glicose em
nível celular; principal ânion do suco gástrico como parte do ácido clorídrico, ativação da amilase intestinal
Magnésio (Mg) Atividade neuromuscular e nervosa; transferência de energia; participação
no crescimento ósseo; participação no metabolismo dos carboidratos; participação no metabolismo dos lipídeos.
Fósforo (P)
Formação óssea e dentária; constituição da molécula de DNA e RNA, formação de fosfolipídios; formação da coluna; participando, assim, na
transmissão dos impulsos nervosos; atividade enzimática, sobretudo como coenzima de vários complexos da vitamina B; fosforilação para a formação
de ATP.
Potássio (K) Balanço osmótico e hídrico corporal; participação no
metabolismo protéico e dos carboidratos; integridade da atividade muscular e nervosa.
Enxofre (S) Metabolismo e síntese protéica; metabolismo das gorduras e dos carboidratos; síntese de vitaminas do complexo B.
MICROELEMENTOS
Cobalto (Co) Função anti-anêmica, por ser componente da vitamina 612 e do ácido fólico;
metabolismo da glicose; síntese da metionina.
Cobre (Cu) Ativador enzimático envolvendo o transporte e a transferência de oxigênio,
metabolismo dos aminoácidos e do tecido conectivo. lodo (l) Componente dos hormônios tireoidianos.
Ferro (Fe) Transporte de oxigênio e respiração celular. Flúor (F) Proteção óssea e dentária.
Manganês (Mn) Integridade da matriz orgânica óssea; ativador enzimático, sobretudo no
metabolismo dos aminoácidos e dos ácidos graxos. Selênio (Se) Junto com a vitamina E, promove a proteção dos tecidos contra danos

14
oxidativos; componente da enzima glutationa peroxidase; metabolismo dos aminoácidos sulfurados.
Zinco (Zn)
Ativador enzimático, principalmente nos processos de formação óssea, do metabolismo dos ácidos nucléicos, do processo da visão, do sistema imunológico e do sistema
reprodutivo.
Fonte: MCDOWELL, 1999
2.3 - Adição de vermífugos e outros suplementos
De modo geral, não é aconselhável utilizar a mistura mineral como
veículo para administração de remédios e aditivos alimentares, por várias
razões. Por exemplo, os vermífugos necessitam ser aplicados em épocas
definidas (início e fim da estação chuvosa e terço final da estação seca),
enquanto a mistura mineral é fornecida de maneira contínua. Além disso, os
vermes são combatidos com doses específicas, conforme o peso dos animais e
não em dose qualquer.
A adição de uréia ao sal mineral poderia ser admitida em condições
bastante restritas, onde fosse possível um cuidadoso controle do consumo,
para evitar risco de intoxicação do gado, inclusive obedecendo a um período
bastante rígido de adaptação. Em face desses problemas, não se aconselha
adicionar uréia ao sal mineral.
No mercado, existem alguns concentrados minerais enriquecidos com as
vitaminas A, D e E, vendidos a preços bastante elevados. Do ponto de vista
nutricional, o complemento dessas vitaminas, nas condições regionais de
forragem verde e luz solar, disponíveis durante o ano inteiro, não parece se
justificar na prática.
2.4 - Fornecimento de minerais ao gado
As formulações minerais são calculadas visando ao suprimento diário
das exigências minerais, geralmente por meio de uma mistura única e
completa. Por isso, há necessidade dos animais terem acesso diário, à
vontade, à mistura.

15
3. ADITIVOS
Serão abordados neste capítulo todos os aspectos relacionados à
utilização de aditivos alimentares na nutrição de ruminantes, tanto de natureza
química, quanto biológica. Também serão discutidos seus possíveis efeitos
nocivos e benéficos nos sistemas produtivos, assim como suas conseqüências
futuras para a criação e o ser humano.
Um aditivo alimentar é toda substância química ou não, que somada à
dieta dos animais, é capaz de causar efeitos benéficos aos diversos aspectos
que envolvem o sistema produtivo. Em ruminantes, estes compostos
geralmente provocam interferências nos padrões de fermentação do rúmen-
retículo, melhorando a eficiência de utilização das dietas. Além disso, podem
participar na manutenção da saúde intestinal, ajudando na absorção dos
nutrientes ingeridos e/ou produzidos do rúmen.
Os aditivos são normalmente utilizados na nutrição de ruminantes
quando se desejam melhorar os índices de desempenho de sistemas
produtivos de caráter intensivo, porém, bons resultados são observados
quando os mesmos são acrescidos em suplementos para animais em pastejo.
Isto ocorre, pois o aditivo muita das vezes aumenta a digestibilidade da
forragem ingerida, tornando este efeito mais expressivo quando a qualidade da
mesma estiver comprometida (época da seca).
Todavia, segundo Compêndio Brasileiro de Alimentação Animal, citado
por Medeiros e Lanna (1999) observa-se falta de definição adequada ao termo
“aditivo”. Neste caso, é proposto um termo mais abrangente, chamado pró-
nutriente, com a seguinte definição: microingrediente de alimentação utilizados
oralmente em pequenas doses que promovem os valores intrínsecos de uma
mistura de nutrientes em uma dieta animal.
Segundo de Goes (2004) os pró-nutrientes são divididos em: ionóforos,
antibióticos, leveduras ativas, probióticos e prebióticos. Os hormônios também
são considerados aditivos, porém não serão abordados neste capítulo.

16
3.1 – Ionóforos
Nota: Apesar do curso ser específico para confinamento, faremos uma revisão
envolvendo animais em pasto e vacas leiteiras acreditando melhor embasar o
tema e transmitir conhecimentos sobre o assunto ao publico.
Os ionóforos são classificados como antibióticos poliéteres carboxílicos
que transportam íons através da membrana e parede celular de bactérias do
grupo Gram-positivas, diminuindo ou inibindo seu crescimento. São produzidos
por espécies de estreptomicetos, constituindo um conjunto de mais de 70
compostos diferentes, sendo os mais comuns a monensina sódica (produzida
pelo Streptomyces cinnamonensis) e a lasalocida sódica (Streptomyces
lasaliensis). Como dito, existem outros ionóforos que também são utilizados na
alimentação de ruminantes, porém alguns destes são menos utilizados no
Brasil: Salinomicina, Laidlomicina, Lisocelina, Narasina e Tetronasina
(NAGARAJA, et al., 1997; MEDEIROS e LANNA, 1999; ONOFRI e GRAÇA,
2005).
A Tab. 2 apresenta uma relação destes ionóforos, os microorganismos
que as produzem, peso molecular e sua seletividade de cátions por ordem de
afinidade.

17
Tabela 2 - Ionóforos já utilizados ou sob investigação para uso em dietas de
ruminantes
Neste capítulo serão apresentadas características e aplicações dos três
ionóforos mais utilizados na alimentação de ruminantes no Brasil: monensina
sódica, lasalocida sódica e salinomicina. Entretanto, os modos de ação destas
moléculas são bastante semelhantes entre si.
Segundo De Goes (2004) os ionóforos são capazes de modificar a
fermentação ruminal, alterando a produção e proporção de AGV’s (Ácidos
Graxos Voláteis) e o metabolismo de proteínas no rúmen.
Essa alteração ocorre, pois estes aditivos são capazes de interferir, em
nível celular, a atividade de bactérias Gram-positivas presentes no rúmen,
modificando os padrões ruminais.
Segundo Peres e Simas (2006) a estrutura dos ionóforos determina sua
ação, porque ela torna possível o transporte de certos cátions pelas
membranas lipídicas da células bacterianas. O termo “polieter” refere-se a uma
estrutura molecular capaz de complexar substâncias lipo-solúveis com cátions
e mediar seu transporte pela barreira lipídica da célula bacteriana (NAGARAJA,
et al., 1997).

18
Esta afinidade por cátions é específica de cada tipo de ionóforo (TAB. 5).
A monensina sódica possui cerca de 31 vezes mais atração por sódio (Na+)
que a lasalocida, enquanto que esta apresenta 10 mil vezes mais afinidade por
cálcio (Ca2+) que a monensina (PERES e SIMAS, 2006).
Em trabalho realizado por Russell (1987) utilizando cepas da bactéria
Streptococcus bovis foi possível observar essa diferença de passagem de íons
entre um sistema sem monensina sódica e outro com este ionóforo. Os
resultados estão apresentados na Tab. 3.
Tabela 3 - Efeitos da monensina sódica na passagem de íons da
membrana plasmática de células da bactéria Streptococcus bovis
Segundo Peres e Simas (2006) como o cátion mais abundante dentro da
célula é o K+ que excede muito ao Na
+, a exposição da monensina provoca um
aumento de H+ intracelular reduzindo o pH. Na tentativa de equilibrar o
processo, o K+ é expulso da célula. Esse desequilíbrio osmoelétrio (pH)
provoca uma série de reações de homeostase celular que gastam muito ATP
(energia), reduzindo consideravelmente o crescimento das bactérias Gram-
positivas.
Como conseqüência desta seleção microbiológica provocada pelos
ionóforos tem-se uma mudança nos padrões ruminais com maior eficiência no
aproveitamento energético. Segundo Oscar et al. (1987) ocorre diminuição na
relação acetato:propionato com incrementos na produção de ácido propiônico e
redução do ácido acético. Além disso, as proporções de metano no rúmen são

19
reduzidas, caracterizando menores perdas de energia, que segundo Nagaraja
et al. (1997) podem chegar a 10% em um sistema normal de alimentação, sem
uso de aditivos melhoradores de eficiência nutricional. O conjunto destas
mudanças resulta em melhor retenção de energia pelo ruminante com reflexos
positivos à sua performance produtiva.
Os ionóforos também provocam uma diminuição na catálise de
peptídeos no rúmen, aumentando o escape destes para o intestino delgado
(DE GOES, 2004) com redução do N amoniacal (OSCAR et al., 1987). Esta
resultante implica em vantagens no aporte de aminoácidos pelo animal
hospedeiro, podendo contribuir para o desempenho animal.
Outro benefício causado pelo uso de estes aditivos, principalmente em
sistemas intensivos de produção, esta relacionado ao aparecimento de
distúrbios alimentares. Segundo De Goes (2004) ocorre redução na incidência
de acidose ruminal e timpanismo, devido a menor concentração de ácido lático
e produção de mucopolissacarídeos, que dão estabilidade à espuma do líquido
ruminal. Estes mucopolissacarídeos são produzidos por bactérias
metanogênicas, seres altamente afetados pela suplementação de ionóforos.
Portanto, o uso destes aditivos auxilia muito no aumento da eficiência alimentar
refletindo então no desempenho animal. A seguir serão discutidos os efeitos
práticos da utilização de cada ionóforo sobre os índices produtivos.
3.2 - Monensina sódica
Segundo Peres e Simas (2006) de maneira geral, há uma similaridade
entre o modo de ação dos ionóforos. Todavia, a monensina foi utilizada na
grande maioria dos trabalhos que encontraram efeitos destes aditivos na
fermentação ruminal. É possível gerar hipóteses sobre a atividade dos outros
ionóforos, mas é necessário considerar que as diferenças químicas, físicas e
biológicas, bem como a especificidade por íons, pKa e propriedades
farmacocinéticas, conferem diferentes efeitos e magnitudes dos efeitos das
diferentes moléculas. Por isso, ao se comparar a ação de diferentes ionóforos
deve-se atentar não só ao efeito produzido pela molécula, mas também a
situação de uso da mesma que pode proporcionar bons resultados em graus
diferentes.
20
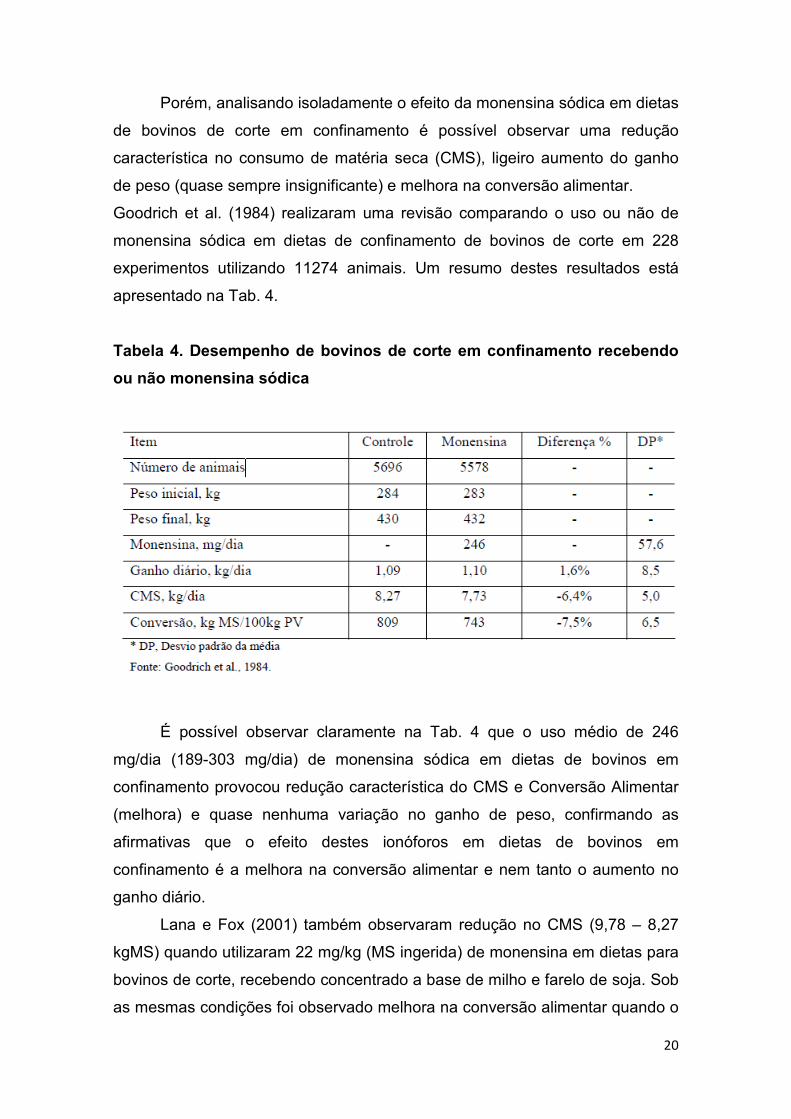
Porém, analisando isoladamente o efeito da monensina sódica em dietas
de bovinos de corte em confinamento é possível observar uma redução
característica no consumo de matéria seca (CMS), ligeiro aumento do ganho
de peso (quase sempre insignificante) e melhora na conversão alimentar.
Goodrich et al. (1984) realizaram uma revisão comparando o uso ou não de
monensina sódica em dietas de confinamento de bovinos de corte em 228
experimentos utilizando 11274 animais. Um resumo destes resultados está
apresentado na Tab. 4.
Tabela 4. Desempenho de bovinos de corte em confinamento recebendo
ou não monensina sódica
É possível observar claramente na Tab. 4 que o uso médio de 246
mg/dia (189-303 mg/dia) de monensina sódica em dietas de bovinos em
confinamento provocou redução característica do CMS e Conversão Alimentar
(melhora) e quase nenhuma variação no ganho de peso, confirmando as
afirmativas que o efeito destes ionóforos em dietas de bovinos em
confinamento é a melhora na conversão alimentar e nem tanto o aumento no
ganho diário.
Lana e Fox (2001) também observaram redução no CMS (9,78 – 8,27
kgMS) quando utilizaram 22 mg/kg (MS ingerida) de monensina em dietas para
bovinos de corte, recebendo concentrado a base de milho e farelo de soja. Sob
as mesmas condições foi observado melhora na conversão alimentar quando o

21
ionóforo foi adicionado à dieta. Ambos os resultados estão de acordo com os
obtidos nos experimentos revisados por Goodrich et al. (1984).
Todavia, neste mesmo experimento realizado por Lana e Fox (2001)
pôde-se observar uma piora na conversão alimentar quando a monensina foi
utilizada juntamente com dietas contendo 4% de gorduras insaturadas.
Segundo os autores isto ocorreu porque a microbiota ruminal foi altamente
afetada pela associação de dois depressores do crescimento bacteriano.
É importante verificar que esta redução no consumo de matéria seca
provocada pela monensina deve ser minuciosamente acompanhada durante a
alimentação dos bovinos, pois existe um limite que se ultrapassado, pode
depreciar o ganho de peso dos animais, piorando a conversão alimentar.
Resumidamente isso pode acontecer de duas formas: se ocorrer ingestão
excessiva de monensina pelos animais (>300 mg/animal/dia); ou se estes
aditivo ionóforo for utilizado juntamente com outros depressores de consumo
(gorduras insaturadas livres).
Segundo ALPHARMA (2002) há aprovação de uso da monensina sódica
em doses que variam de 50 a 360 mg/animal/dia, sendo que na prática
normalmente se utiliza 200-300 mg/animal/dia para bovinos em confinamento;
e 100 a 200 mg para bovinos suplementados a pasto.
Segundo De Goes (2004) a utilização 200 mg de monensina sódica por
dia para bovinos criados a pasto resulta em ótima resposta sobre o
desempenho dos animais, pois melhora o aproveitamento energético de dietas
ricas em fibras. A ingestão de monensina sódica via suplemento geralmente
não influencia o consumo de matéria e eleva o ganho de peso dos animais. Isto
ocorre porque, neste caso, o suplemento participa em uma pequena parcela do
alimento ingerido, e portanto, interfere pouco na matéria seca total ingerida. O
principal efeito deste ionóforo esta no aumento da energia metabolizável
fornecida pelo capim.
Na revisão realizada por Goodrich et al. (1984) verificou-se efeito
positivo no ganho de peso quando se utilizou 154 mg de monensina/cab./dia
em suplementos para animais a pasto. Em média, os ganhos aumentaram de
609 para 690g (P<0,01). Nesta avaliação foram utilizados 914 novilhos de 24
experimentos. Neste caso os autores citam que as dosagens mais
22
recomendadas nos trabalhos revisados, variam de 150 a 200 mg de
monensina/animal/dia.
Em experimentos realizados por Fieser et al. (2007) utilizando 253
novilhos mestiços em um primeiro ano (Peso Inicial - 255 kg PV) e 116 animais
no segundo ano (287 kg PVinicial), pastejando em áreas de forragem de
inverno (trigo), observou-se elevação nos ganhos de peso para ambos os anos,
quando se incluiu monensina nos suplementos dos animais.
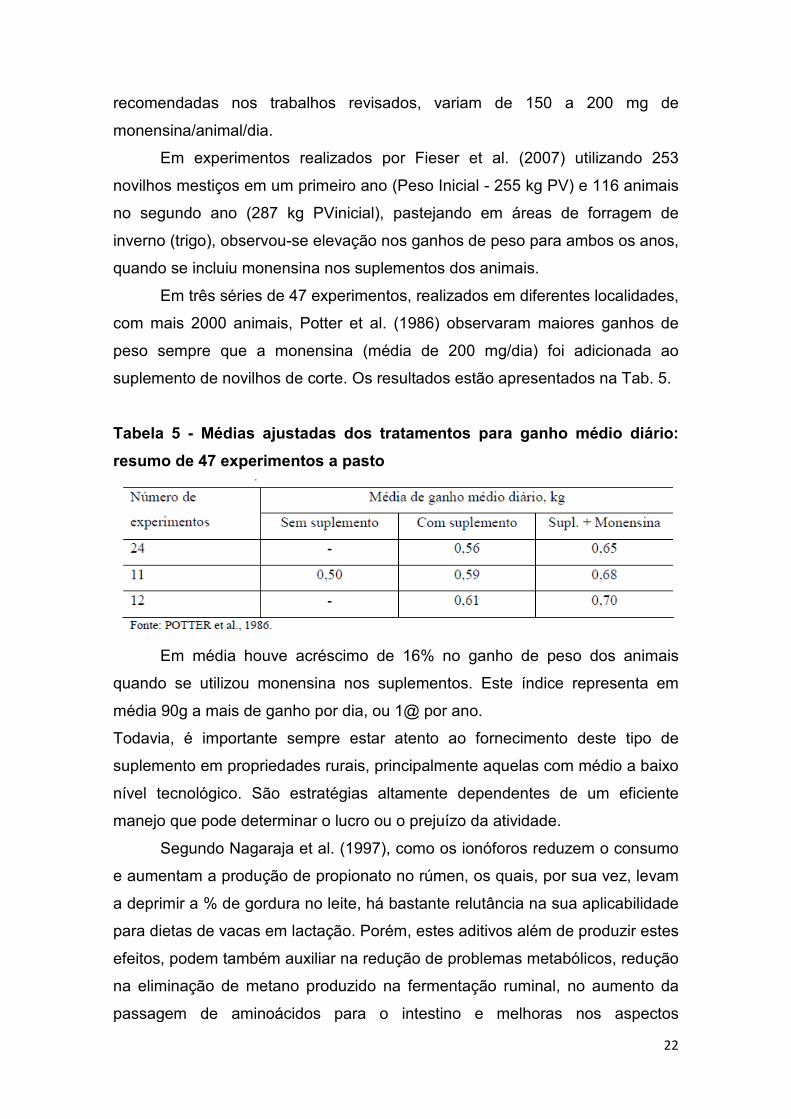
Em três séries de 47 experimentos, realizados em diferentes localidades,
com mais 2000 animais, Potter et al. (1986) observaram maiores ganhos de
peso sempre que a monensina (média de 200 mg/dia) foi adicionada ao
suplemento de novilhos de corte. Os resultados estão apresentados na Tab. 5.
Tabela 5 - Médias ajustadas dos tratamentos para ganho médio diário:
resumo de 47 experimentos a pasto
Em média houve acréscimo de 16% no ganho de peso dos animais
quando se utilizou monensina nos suplementos. Este índice representa em
média 90g a mais de ganho por dia, ou 1@ por ano.
Todavia, é importante sempre estar atento ao fornecimento deste tipo de
suplemento em propriedades rurais, principalmente aquelas com médio a baixo
nível tecnológico. São estratégias altamente dependentes de um eficiente
manejo que pode determinar o lucro ou o prejuízo da atividade.
Segundo Nagaraja et al. (1997), como os ionóforos reduzem o consumo
e aumentam a produção de propionato no rúmen, os quais, por sua vez, levam
a deprimir a % de gordura no leite, há bastante relutância na sua aplicabilidade
para dietas de vacas em lactação. Porém, estes aditivos além de produzir estes
efeitos, podem também auxiliar na redução de problemas metabólicos, redução
na eliminação de metano produzido na fermentação ruminal, no aumento da
passagem de aminoácidos para o intestino e melhoras nos aspectos

23
reprodutivos. A principal conseqüência final deste balanço entre aspectos
positivos e negativos provoca um saldo que normalmente resultado em
incrementos na produção diária de leite.
Em uma reunião de experimentos feita por McGuffey et al. (2001) é
possível verificar claramente este efeito positivo na atividade produtiva dos
animais (Tab. 6).
Tabela 6 - Efeitos da monensina sódica no desempenho de vacas em
lactação
Pode-se verificar na Tab. 6 um aumento significativo na produção leiteira
(PL em kg/dia) de 27,5 para 28,8 (+1,3). Todavia houve decréscimo na % de
gordura no leite de 3,98 para 3,78, porém sem prejudicar a produção deste
nutriente diariamente. A pequena redução na % de proteína foi compensada
pela PL resultando em efeito inverso sobre a quantidade diária produzida
(0,846 – 0,872, respectivamente para Controle e Monensina). O CMS também
não foi prejudicado, mostrando o efeito positivo que a monensina causa sobre
a fermentação ruminal e conseqüentemente sobre o desempenho.
Mutsvangwa, et al. (2002) realizaram um experimento avaliando a
utilização da monensina sódica (22 mg/kg de MS ingerida) em situação
induzida de acidose sub-clínica por 10 dias (22-32 dias de lactação) e período
de recuperação posterior (32-42 dias). O trabalho foi conduzido em “crossover”
utilizando-se 6 vacas holandesas multíparas com média 667kg PV. Os
resultados estão apresentados nas Tab. 7 e 8.
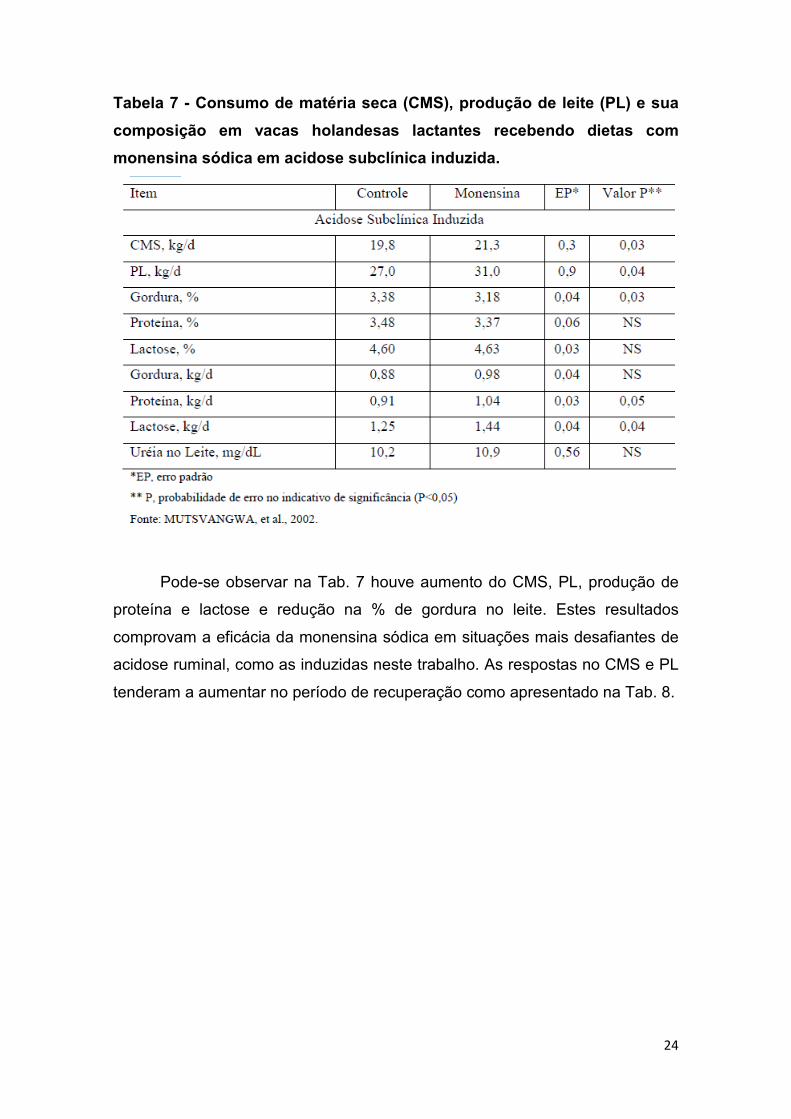
24
Tabela 7 - Consumo de matéria seca (CMS), produção de leite (PL) e sua
composição em vacas holandesas lactantes recebendo dietas com
monensina sódica em acidose subclínica induzida.
Pode-se observar na Tab. 7 houve aumento do CMS, PL, produção de
proteína e lactose e redução na % de gordura no leite. Estes resultados
comprovam a eficácia da monensina sódica em situações mais desafiantes de
acidose ruminal, como as induzidas neste trabalho. As respostas no CMS e PL
tenderam a aumentar no período de recuperação como apresentado na Tab. 8.
25
Tabela 8 - Consumo de matéria seca (CMS), produção de leite (PL) e sua
composição em vacas holandesas lactantes recebendo dietas com
monensina sódica no período de recuperação após indução de acidose
subclínica
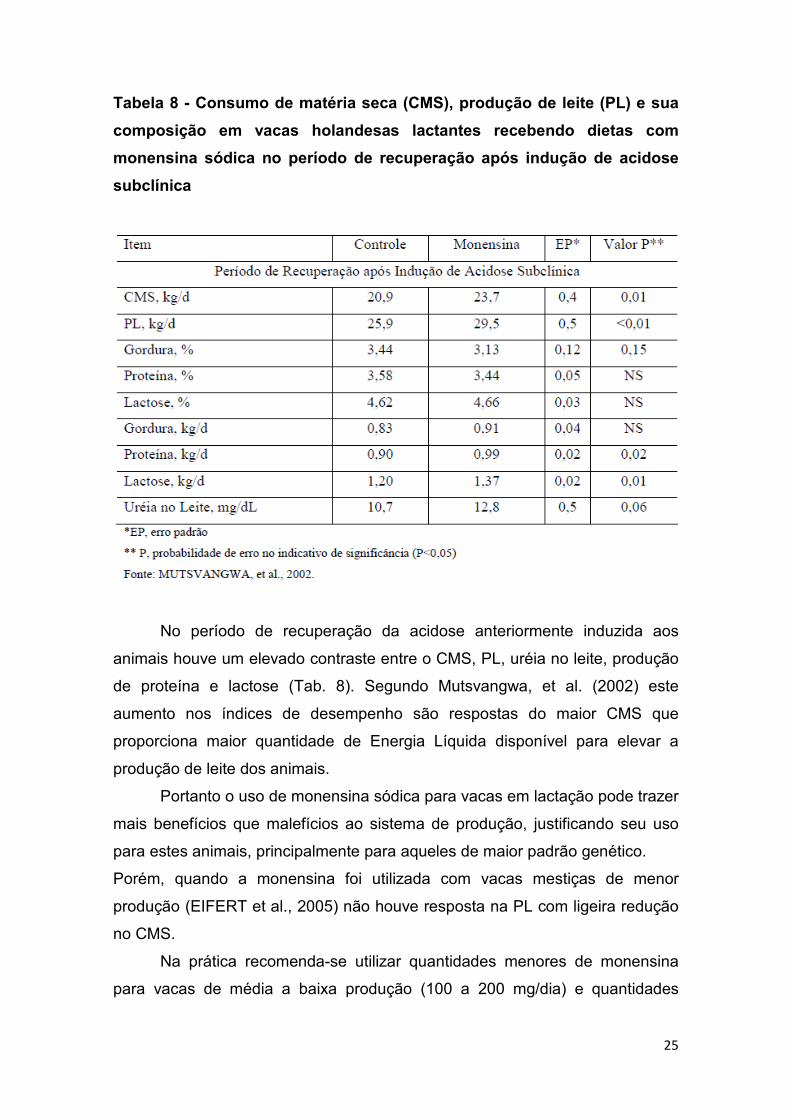
No período de recuperação da acidose anteriormente induzida aos
animais houve um elevado contraste entre o CMS, PL, uréia no leite, produção
de proteína e lactose (Tab. 8). Segundo Mutsvangwa, et al. (2002) este
aumento nos índices de desempenho são respostas do maior CMS que
proporciona maior quantidade de Energia Líquida disponível para elevar a
produção de leite dos animais.
Portanto o uso de monensina sódica para vacas em lactação pode trazer
mais benefícios que malefícios ao sistema de produção, justificando seu uso
para estes animais, principalmente para aqueles de maior padrão genético.
Porém, quando a monensina foi utilizada com vacas mestiças de menor
produção (EIFERT et al., 2005) não houve resposta na PL com ligeira redução
no CMS.
Na prática recomenda-se utilizar quantidades menores de monensina
para vacas de média a baixa produção (100 a 200 mg/dia) e quantidades

26
maiores para alta lactação (200 a 300 mg/dia). Em geral, no Brasil, não se
recomendam quantidades superiores a 300 mg/dia de monensina por evitar
interferências no CMS que possam deprimir a PL.
3.3 - Lasalocida sódica
A lasalocida sódica é um ionóforo que assim como a monensina, age
reduzindo o crescimento de bactérias Gram-positivas através de um gasto
energético (ATP) excessivo provocado pela troca constante de íons na
membrana celular. Todavia, a monensina possui grande afinidade com o Na+,
K+ e H
+, enquanto que a lasalocida apresenta mais afinidade com Ca
++ e Mg
++
(ONOFRI e GRAÇA, 2005).
Existem várias evidências que a lasalocida apresenta melhor
aceitabilidade aos animais comparada com a monensina (ZINN, 1987;
ERICKSON, et al. 2004).
ALPHARMA (2002) apresenta uma reunião de estudos mostrando os
níveis de toxicidade da monensina sódica e lasalocida para três espécies
animais. Os resultados estão na Tab. 9.
Tabela 9. Níveis de toxicidade de monensina sódica e lasalocida em três
diferentes espécies animais
Pode-se observar na Tab. 9 que em geral os níveis máximos de
fornecimento da monensina são mais estreitos que os da lasalocida. Merece
destaque a quantidade potencialmente tóxica da monensina para eqüinos,
indicando que este ionóforo, mesmo em pequenas quantidades é
extremamente tóxico para esta espécie animal, fato que não ocorre com a
lasalocida.

27
Outro fator que diferencia a lasalocida da monensina é o tempo
necessário para adaptação. ALPHARMA (2002) cita que a monensina
necessita de adapatação enquanto que a lasalocida não necessita.
Esta característica ficou evidenciada no trabalho realizado por Delfino et
al. (1988), onde foram utilizados 20 animais para cada um dos 5 tratamentos:
Controle – sem ionóforo; Monensina – 33 mg/kg MS; Lasalocida em 3 níveis
(24,36,54 mg/kg MS, respectivamente para Las1, Las2 e Las 3). Os resultados
para Ganho de Peso Diário (GPD) estão apresentados no Gráf. 1.
Pode-se observar no Gráf. 1 que o Las 3 (54 mg/kg MS) apresentou uma
acentuada elevação nos ganhos de peso logo nos primeiro 28 dias de
avaliação, caracterizando um aditivo que possui ação rápida no desempenho
sem necessidade de adaptação. Enquanto isso, a monensina apresentou uma
elevação nos ganhos (1,32kg/dia) em relação ao tratamento controle, mas
ainda menor que Las 3 (1,46kg/dia).
No experimento de Delfino et al. (1988) a conversão alimentar para
lasalocida (6,4; 6,2 e 6,2; respectivamente para Las1, 2 e 3) foi semelhante em
relação à monensina sódica (6,5 kgMS/kgPV), porém menor que o controle (6,9
kgMS/kgPV).

28
A melhor adaptação dos animais à lasalocida pode influenciar no
sistema produtivo como um todo, acelerando o crescimento dos mesmos com o
intuito de reduzir a idade ao abate.
Os resultados para animais em confinamento são inconstantes, existindo
experimentos que demonstram menores ganhos de peso para lasalocida em
relação a monensina (Clary, et al., 1993). Estes experimentos devem ser
analisados com cuidado, pois as situações em que foram realizados podem
explicar, quase sempre, as variações dos resultados. No caso de Clary, et al.
(1993), o maior ganho para monensina pode ser explicado pela composição do
concentrado e sua proporção na dieta total (90%).

29
Tabela 10 - Desempenho de animais cruzados Zebu X Chianina, em
Brachiaria decumbens, durante 112 dias, com ou sem adição de
lasalocida
Observa-se na Tab. 10 que houve maior GPD para o suplemento que
utilizou lasalocida como aditivo, mostrando que mesmo em condições de
pastejo a lasalocida possui efeito benéfico ao desempenho dos animais.
Para bovinos leiteiros em lactação, McGuffey et al. (2001) encontrou menor
resposta da lasalocida em relação ao tratamento controle, sobre o desempenho
dos animais. Uma reunião de 6 experimentos analisados por estes autores
mostraram produções de leite inalteradas (28,7 e 28,3 kg/dia, respectivamente
para Controle e Lasalocida) e consumos de matéria seca semelhantes (19,4 e
18,6 kg/dia).
Na prática a lasalocida sódica é recomendada em níveis semelhantes as
da monensina sódica, variando de 100-300mg/cab./dia.
3.4 - Antibióticos não ionóforos
Os antibióticos que não apresentam característica ionófora também
podem ser utilizados como promotores de eficiência alimentar. Entretanto, seus
princípios de ação podem variar de acordo com suas características
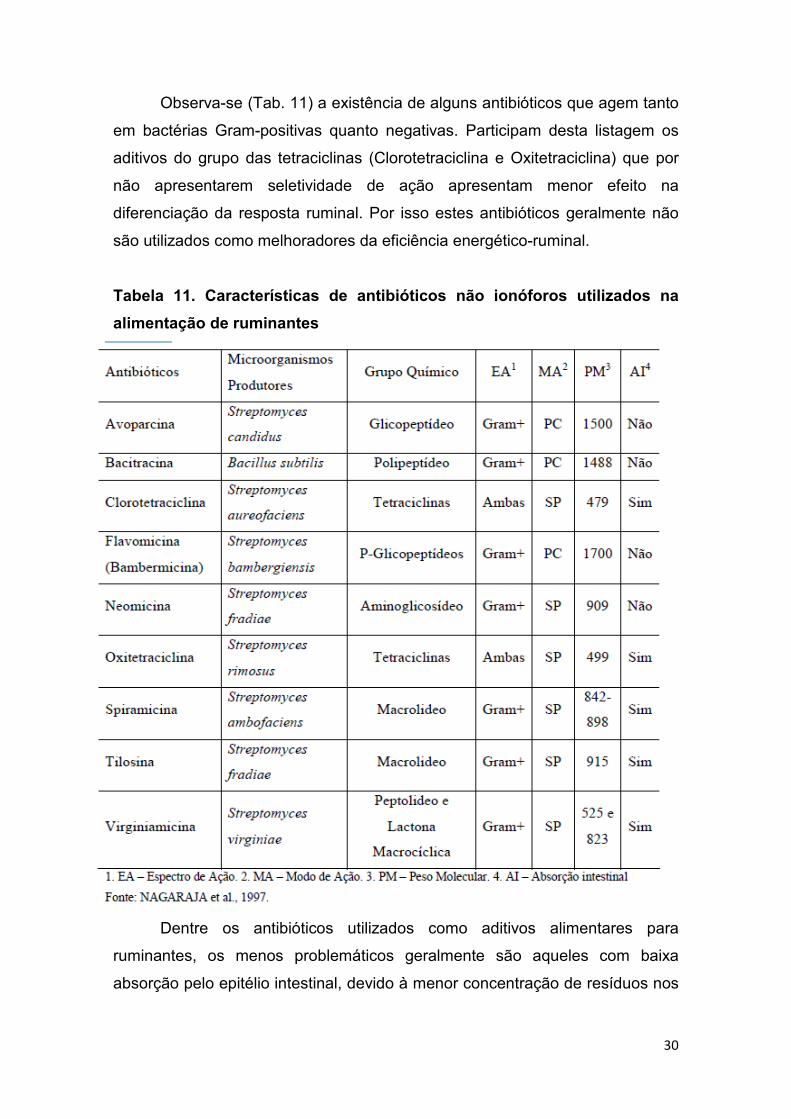
bioquímicas. A Tab. 11 apresenta uma relação de vários antibióticos
comumente utilizados na alimentação animal e suas respectivas características
intrínsecas.
30
Observa-se (Tab. 11) a existência de alguns antibióticos que agem tanto
em bactérias Gram-positivas quanto negativas. Participam desta listagem os
aditivos do grupo das tetraciclinas (Clorotetraciclina e Oxitetraciclina) que por
não apresentarem seletividade de ação apresentam menor efeito na
diferenciação da resposta ruminal. Por isso estes antibióticos geralmente não
são utilizados como melhoradores da eficiência energético-ruminal.
Tabela 11. Características de antibióticos não ionóforos utilizados na
alimentação de ruminantes
Dentre os antibióticos utilizados como aditivos alimentares para
ruminantes, os menos problemáticos geralmente são aqueles com baixa
absorção pelo epitélio intestinal, devido à menor concentração de resíduos nos

31
alimentos (carne e leite) e ao menor período de carência para o abate
(MANTOVANI, et al., 2006).
Todavia, apesar de apresentar absorção intestinal, atualmente no Brasil,
a Virginiamicina é o antibiótico não ionóforo mais utilizado como promotor de
eficiência alimentar para ruminantes devido a sua eficácia no controle dos
padrões ruminais.
3.4.1 - Virginiamicina
A Virginiamicina, um produto da fermentação da Streptomyces virginiae,
é formada pela mistura de dois subfatores (M e S) que possuem ação anti-
microbiana sinérgica. É um antibiótico que age seletivamente em bactérias
Gram-positivas, penetra na parede celular da célula, alcança as regiões onde
os ribossomos estão realizando a síntese protéica, liga-se à sub-unidade 50S
desta organela e bloqueia a formação das proteínas, inibindo a ligação dos
peptídeos. O fator S potencializa a atividade do fator M e a junção dos dois
desfaz o processo metabólico dentro da célula, resultando no efeito bactericida
(ROGERS et al., 1995; NAGARAJA, 1997).
Segundo Rogers et al. (1995) a Virginiamicina é um aditivo alimentar que
ajuda a estabilizar a fermentação ruminal diminuindo as variações de ingestão
de MS. Portanto, este antibiótico auxilia muito na adaptação dos animais às
dietas de alto grão, não permitindo que estes desacelerem o ganho de peso
durante o período inicial do confinamento.
O efeito sobre o padrão ruminal é semelhante aos ionóforos reduzindo a
produção de metano e ácido lático, elevando o propionato e diminuindo a
degradação protéica. Sendo assim, auxilia na prevenção de acidoses, ruminites
e formação de abscessos hepáticos. Este último ocorre por possuir ação sobre
a Fusobacterium necrophorum, agente causador desta doença (Coe et al.,
1999).
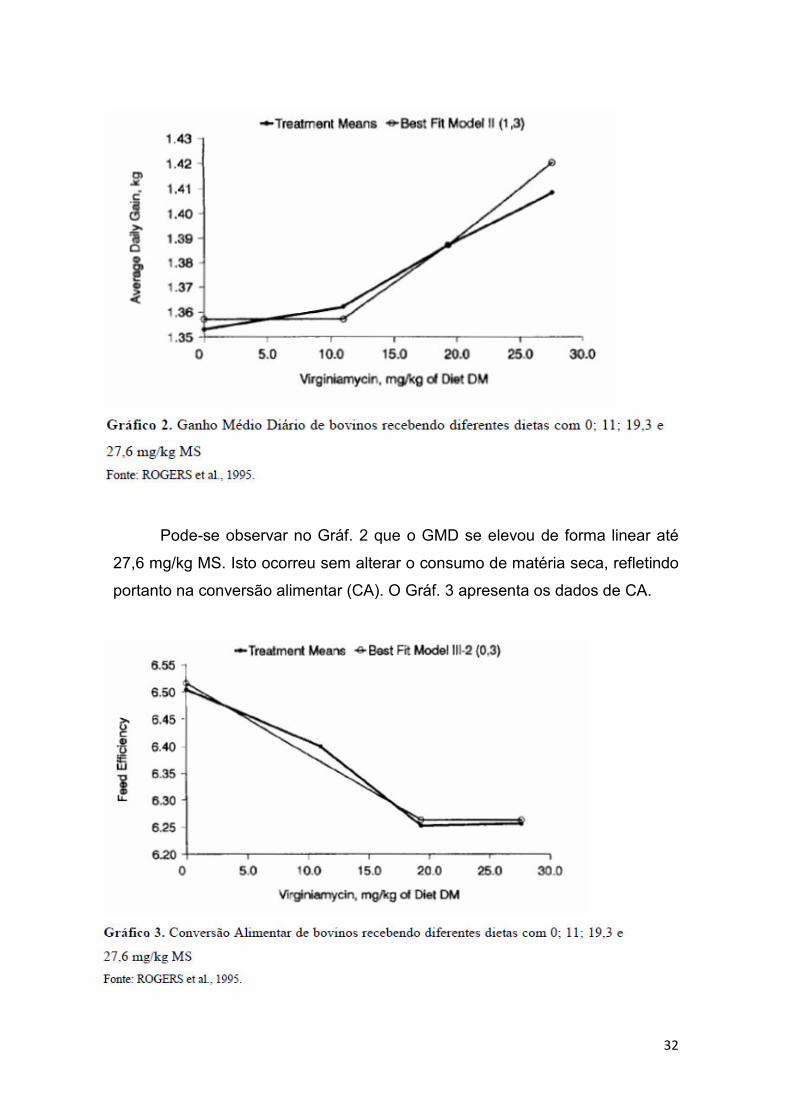
Rogers et al. (1995) realizaram um estudo de 7 experimentos com
aproximadamente 3100 novilhos e novilhas, em confinamento, avaliando a
inclusão de diferentes doses de Virginiamicina. O CMS não sofreu alteração
entre Controle e os demais tratamentos. O GMD está apresentado no Gráf. 2.
32
Pode-se observar no Gráf. 2 que o GMD se elevou de forma linear até
27,6 mg/kg MS. Isto ocorreu sem alterar o consumo de matéria seca, refletindo
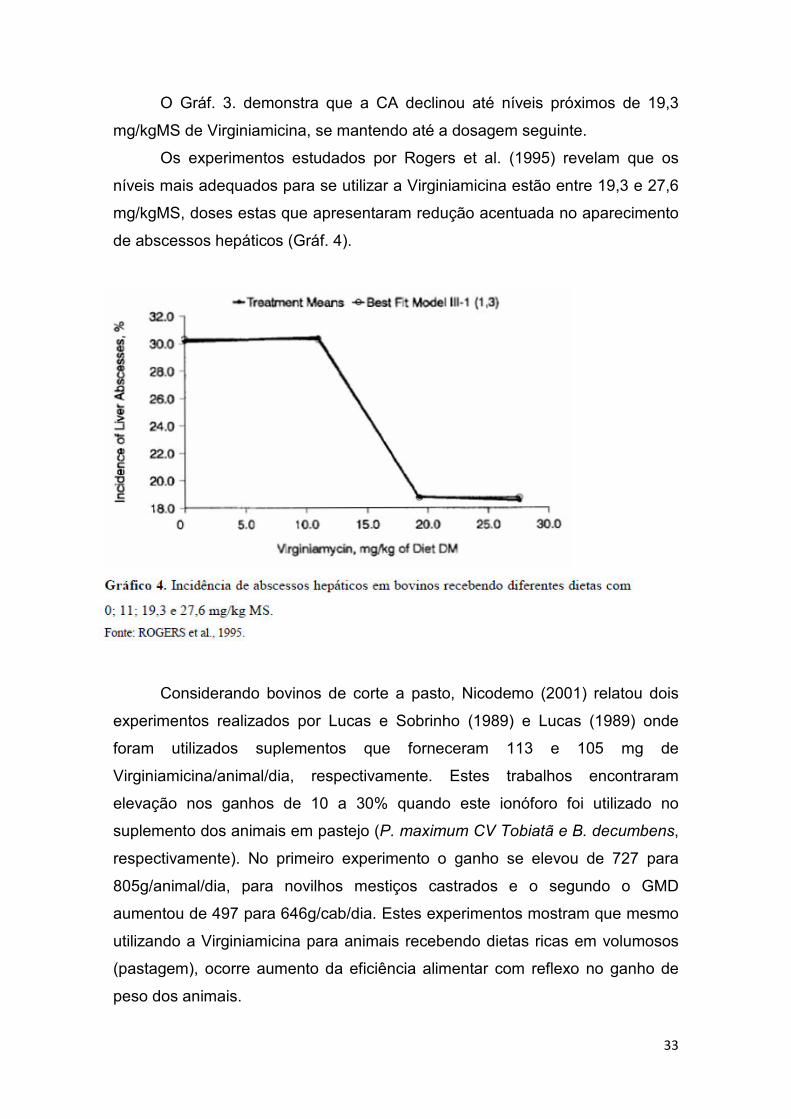
portanto na conversão alimentar (CA). O Gráf. 3 apresenta os dados de CA.
33
O Gráf. 3. demonstra que a CA declinou até níveis próximos de 19,3
mg/kgMS de Virginiamicina, se mantendo até a dosagem seguinte.
Os experimentos estudados por Rogers et al. (1995) revelam que os
níveis mais adequados para se utilizar a Virginiamicina estão entre 19,3 e 27,6
mg/kgMS, doses estas que apresentaram redução acentuada no aparecimento
de abscessos hepáticos (Gráf. 4).
Considerando bovinos de corte a pasto, Nicodemo (2001) relatou dois
experimentos realizados por Lucas e Sobrinho (1989) e Lucas (1989) onde
foram utilizados suplementos que forneceram 113 e 105 mg de
Virginiamicina/animal/dia, respectivamente. Estes trabalhos encontraram
elevação nos ganhos de 10 a 30% quando este ionóforo foi utilizado no
suplemento dos animais em pastejo (P. maximum CV Tobiatã e B. decumbens,
respectivamente). No primeiro experimento o ganho se elevou de 727 para
805g/animal/dia, para novilhos mestiços castrados e o segundo o GMD
aumentou de 497 para 646g/cab/dia. Estes experimentos mostram que mesmo
utilizando a Virginiamicina para animais recebendo dietas ricas em volumosos
(pastagem), ocorre aumento da eficiência alimentar com reflexo no ganho de
peso dos animais.

34
Atualmente no Brasil a Virginiamicina não é muito utilizado como aditivo
para bovinos leiteiros em lactação, porém Clayton et al. (1999) estudaram sua
utilização (30 mg/kgMS) para 71 vacas lactantes em pastejo. Estes autores não
encontraram diferenças na produção de gordura, proteína, lactose e consumo
de matéria seca. Todavia, houve tendência de aumento na produção (P=0,089)
em relação ao tratamento controle. Estas variações podem ser observadas no
Gráf. 6. que apresenta a lactação das vacas estudadas.
É possível observar que o uso da Virginiamicina aparentemente afetou a
lactação da vacas estudadas em certos momentos das lactações. Houve maior
produção para os tratamentos com este ionóforo, principalmente no ápice da
curva de lactação, onde essas vacas receberam maiores quantidades de
concentrado. Porém, quando as médias de produções decaíram com o passar
do tempo, este efeito diferencial não ficou tão evidenciado.
Na prática recomenda-se o uso de 150 a 300 mg/cab./dia de Virginiamicina
para bovinos de corte em confinamento, 100 mg para gado de corte a pasto e

35
250 a 400 mg para bovinos de leite em lactação, dependendo da produção de
leite.
3.5 - Promotores de Eficiência Alimentar Biológicos
A fim de aumentar a eficiência no aproveitamento energético das dietas
para ruminantes, técnicos têm recomendado a utilização dos ionóforos
supracitados que funcionam como “selecionadores” de microorganismos
ruminais tornando o meio mais eficiente em produzir energia. Porém, tais
aditivos vêem sendo abolidos das rações de animais que servem ao mercado
internacional, segundo exigências impostas pelos mesmos, apesar do Brasil e
outros países (EUA) ainda não atenderem tal exigência em sua legislação
corrente. A justificativa dos países compradores (UE) de carne são que o
mercado consumidor destas localidades tendem a rejeitar a carne que possa
conter resíduos de aditivos químicos, fazendo valer a chamada segurança
alimentar.
Para contornar estas limitações cresce o interesse científico mundial em
pesquisar alternativas que produzam os mesmos resultados sem deixar
resíduos químicos. As formas naturais de provocar estes efeitos nos padrões
ruminais incluem adições de cepas de leveduras ativas (vivas) e bactérias
benéficas ao ecossistema do rúmen.
Quanto às leveduras (Saccharomyces cerevisae) utilizam-se atualmente
cepas especialmente selecionadas, desidratadas que apresentem o maior
efeito ruminal possível. Um grande exemplo é a cepa 1026 que efetivamente
reduzem as perdas energéticas no rúmen e, conseqüentemente, melhoram o
ganho de peso dos animais.
3.5.1 - As leveduras ativas
As Saccharomyces cerevisae cepa 1026 são leveduras viáveis as quais
compõe o produto Yea-Sacc®1026
. Estes microorganismos não são
normalmente encontrados no ambiente ruminal, mas estudos mostram que eles
podem proporcionar efeito altamente benéfico a tal ecossistema se

36
suplementados junto à dieta diariamente. Segundo Ingledew e Jones (1982)
esta espécie de levedura é metabolicamente ativa no fluido ruminal por mais de
6 horas.
Enquanto ativas, elas favorecem indiretamente o crescimento das
bactérias que digerem fibras (bactérias celulolíticas). Um aumento na
população destas bactérias poderia provocar melhoria no aproveitamento de
alimentos fibrosos (volumoso) e conseqüentemente, um aumento nos índices
produtivos (NAGARAJA et. al., 1997). Isto ocorre principalmente devido ao
efeito que estas leveduras causam no pH ruminal, fazendo com que este se
mantenha em padrões adequados ao crescimento das bactérias celulolíticas (≥
6,0), mesmo quando alimentos altamente energéticos são adicionados à dieta.
A Saccharomyces cerevisae é capaz de estimular o crescimento de bactérias
(Selenomonas ruminantium) que consomem o ácido lático produzido pelas
bactérias amilolíticas (que utilizam o amido) convertendo-o em AGV´s, fontes
de energia (MARTIN e STREETER, 1995).
Um aumento no crescimento das bactérias celulolíticas proporcionaria não
só elevação no aproveitamento da fibra, mas como também, maximização das
taxas de produção de ácidos graxos voláteis no rúmen, disponibilizando mais
energia ao ruminante hospedeiro (NAGARAJA et. al., 1997).
Outra conseqüência do crescimento das bactérias celulolíticas seria no
consumo de amônia livre no rúmen, utilizando-a no processo de síntese
microbiana, considerando que essas bactérias são altamente dependentes
desta molécula à sua multiplicação (GÜRTLER, 1987).
Smink e Fitie (2007) realizaram um experimento in-vitro onde avaliaram a
inclusão ou não de leveduras ativas em substratos de líquido ruminal + dieta
(50% Volumoso – 50% Concentrado). Foi obtida redução significativa do
metano, além de elevação nas quantidades de AGV’s como um todo. Estes
resultados mostram a capacidade destas leveduras em modificar os padrões
ruminais em benefício do ganho energético obtido no meio.
Outro efeito causado pelas leveduras seria o consumo de oxigênio
eventualmente presente no rúmen. As bactérias ruminais são, em sua maioria,
anaeróbicas estritas e portanto não toleram a presença de oxigênio. Newbold
et al. (1993) mediram a ingestão de oxigênio e obtiveram índice entre 60 e 100
nmol/min/mL, indicando existência permanente, mas não constante, de

37
oxigênio dissolvido no conteúdo ruminal. Segundo Arcuri et al. (2006) leveduras
também são anaeróbicas facultativas, com elevada capacidade respiratória
(200 a 300 nmol/min/g). No experimento de Newbold et al. (1993) foi
encontrado redução de 46 a 89% do oxigênio ruminal quando havia adição de
levedura. Este aspecto leva crer num aumento das populações de bactérias
anaeróbicas estritas, beneficiando o ambiente ruminal.
Gurita (2007) desenvolveu um trabalho utilizando bovinos de corte
alimentados com dois tipos de dietas (52% Volumoso/48% Concentrado – P1;
e posteriormente, 25% Volumoso/75% Concentrado – P2) com ou sem
leveduras. Em ambos os experimentos foram encontrados maiores ganhos de
peso para o tratamento com levedura ativa, sendo que em P1 o ganho se
elevou de 1,195 para 1,380 e em P2 de 1,100 para 1,448 kg/animal/dia.
Segundo o autor essas expressivas diferenças de ganho ocorreram devido às
maiores produções de AGV’s obtidas, assim como melhorias nos padrões
ruminais que definem a funcionalidade dos microorganismos, aumentando o
consumo de MS e conseqüentemente o ganho de peso.
Em bovinos leiteiros Steingass et al. (2007) também observaram
melhorias na produção quando utilizaram leveduras vivas nas dietas de 45
vacas Holstein Friesian. Os índices se elevaram de 34,6 para 36 kg/vaca/dia.
Portanto, os mesmos efeitos causados em bovinos de corte podem afetar
bovinos de leite melhorando o desempenho.
A recomendação de inclusão da levedura irá depender da concentração
da mesma assim como da cepa utilizada. É importante salientar que este último
fator é de grande importância e é ele que determina a funcionalidade da
levedura. A cepa 1026 é a mais utilizada em todo o mundo para este fim e por
isso imprime confiança em quem a usa. Atualmente existem muitas empresas
que comercializam leveduras, porém nem todas apresentam a idoneidade
necessária para garantir resultados.
3.5.2 - Os probióticos bacterianos
Os probióticos (do grego ‘pró-vida’) são culturas vivas de
microorganismos (geralmente bactérias láticas) que favorecem o equilíbrio
ecológico da microbiota ruminal e intestinal, promovendo efeitos benéficos à

38
saúde e crescimento animal (MANTOVANI, 2006). Segundo Arcuri et al. (2006)
a definição mais aceita descreve o probiótico como “um suplemento alimentar à
base de micróbios vivos que beneficia o animal hospedeiro pela melhoria do
seu balanço microbiano do meio em questão”.
Os benefícios diretos e indiretos obtidos pela adição de probióticos nas
dietas dos bovinos podem apresentar-se como: promoção do crescimento
animal; melhoria na conversão alimentar; maior absorção de nutrientes pelo
controle da diferenciação e proliferação das células epiteliais; neutralização de
fatores antinutricionais; melhoria no metabolismo de carboidratos, cálcio e
síntese de vitaminas; produção de enzimas microbianas para compensar
atividades deficientes de enzimas do hospedeiro; eliminação ou controle de
microorganismos patogênicos causadores de doenças subclínicas ou clínicas;
e estímulo de imunidade específica ou não-específica no intestino (ACURI et
al., 2006).
De um modo geral o princípio de ação de um probiótico bacteriano está
relacionado ao princípio da competição por alimentos em um determinado
meio. É possível portanto controlar o meio ruminal por adição de populações
específicas de bactérias que são benéficas aos padrões ruminais.
Bactérias como a Lactobacillus acidophilus reduzem a proteólise no
rúmen e produzem ácido lático no intestino, diminuindo populações de
bactérias patogênicas. A Megasphaera elsdenii é um microorganismo
consumidor de ácido lático e, portanto, sua adição ajuda a reduzir o acúmulo
de composto no rúmen, controlando o pH do meio. (NAGARAJA, 1997; IMEVE,
2003).
IMEVE (2003) relaciona 6 espécies bacterianas essências tanto à saúde
do rúmen quanto intestinos e que possuem grande funcionalidade quando
adicionadas aos alimentos. Suas características estão apresentadas na Tab.
16.
Observa-se na Tab. 12 que todas as bactérias destinadas ao rúmen são
Gram-, pois estas melhoram o aproveitamento energético do meio, como
descrito anteriormente. Quando ocorre aumento da população destas
bactérias, elas tendem a dominar sobre as outras, restringindo o crescimento
de bactérias Gram+ (menor aproveitamento energético). Com o aumento da
população de bactérias Gram-, o meio ruminal produz maiores quantidades de
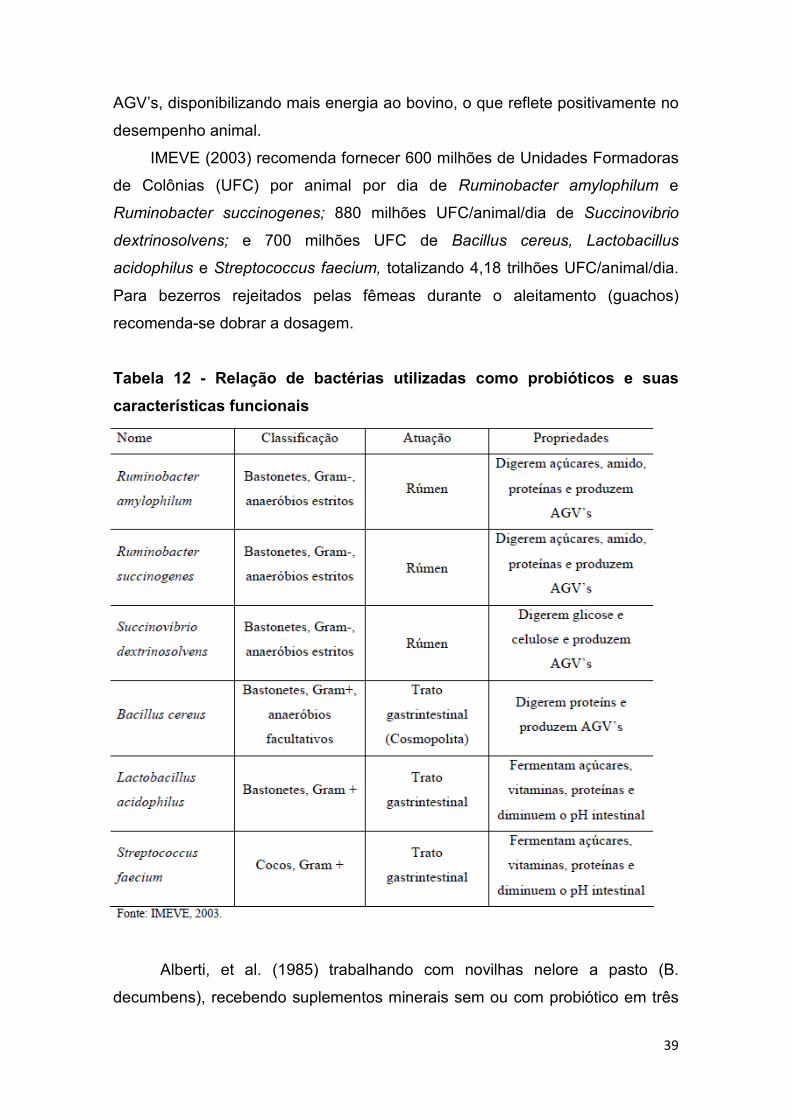
39
AGV’s, disponibilizando mais energia ao bovino, o que reflete positivamente no
desempenho animal.
IMEVE (2003) recomenda fornecer 600 milhões de Unidades Formadoras
de Colônias (UFC) por animal por dia de Ruminobacter amylophilum e
Ruminobacter succinogenes; 880 milhões UFC/animal/dia de Succinovibrio
dextrinosolvens; e 700 milhões UFC de Bacillus cereus, Lactobacillus
acidophilus e Streptococcus faecium, totalizando 4,18 trilhões UFC/animal/dia.
Para bezerros rejeitados pelas fêmeas durante o aleitamento (guachos)
recomenda-se dobrar a dosagem.
Tabela 12 - Relação de bactérias utilizadas como probióticos e suas
características funcionais
Alberti, et al. (1985) trabalhando com novilhas nelore a pasto (B.
decumbens), recebendo suplementos minerais sem ou com probiótico em três

40
níveis, registrou aumento significativo no ganho de peso (0,303-0,692
kg/animal/dia).
Almeida, et al. (1985) avaliando misturas minerais com ou sem
probiótico fornecidas a novilhos cruzados observaram aumento nos ganhos de
peso diários de 0,429 para 0,552 kg/animal/dia.
Portanto, o uso de probióticos melhora os ganhos de peso em animais
de corte, por aumentar o aproveitamento dos alimentos ingeridos e digeridos
no rúmen.
Na prática pode-se observar que a associação de aditivos como
probióticos e leveduras ativas causa maiores benefícios ao sistema, pois além
de melhorar a saúde do sistema gastrointestinal, potencializa ainda mais o
aproveitamento dos alimentos ingeridos.

41
4 – REFERENCIAS
ROGERS, J. A. et al. Effects of Dietary Virginiamycin on Performance and Liver
Abscess Incidence in Feedlot Cattle. Journal Animal Science, v. 73, p. 9-20,
1995.
RUSSELL, J. B. A proposed mechanism of monensin action in inhibitin ruminal
bacterial growth: effects on ion flux and protonmotive force. Journal Animal
Science. v. 64, p. 1519-1525, 1987.
SAMPAIO, A.A.M.; EZEQUIEL, J.M.B.; CAMPOS, F.P. de. Utilização da cama
de frangos e da soja-grão na alimentação de bovinos confinados. Revista
Brasileira de Zootecnia, v.24, n.2, p. 252, 1995.
SICILIANO-JONES, J. Garantindo a segurança e lucratividade com Optigen®
1200: um sistema complete de fornecimento de nitrogênio. In: SIMPÓSIO
BRASILEIRO DA ALLTECH, 2005, Curitiba. Anais.... Curitiba: Alltech, 2005. p.
111-118.
SILVA et al. Digestão total e parcial de alguns componentes de dietas contendo
diferentes níveis de casca de soja e fontes de nitrogênio, em bovinos. Revista
Brasileira de Zootecnia, v.31, n.3, p. 1258-1268, 2002.
SILVA, J.F.C. da. Uréia como aditivo para alimentos volumosos. In: SIMPÓSIO
SOBRE NUTRIÇÃO DE BOVINOS, 2., 1984, Piracicaba. Anais... Piracicaba:
FEALQ, 1984. p. 80-118.
ANDRIGHETTO,J.M., Nutrição Animal. 2 vols., Livraria Nobel, 1986.
MAYNARD, L. A. et al. Nutrição animal. 3. ed. Rio de Janeiro: Ed. Freitas
Bastos, 1984.
PEIXOTO, A.M.; MOURA, J.C.; FARIA, V.P., Ed. ; Minerais para ruminantes, 3º
Simpósio sobre nutrição de bovinos, Anais..., Piracicaba, FEALQ, 1985

42
SILVA, J.F.C.; Fundamentos de nutrição dos ruminantes, Piracicaba, FEALQ,
1979
TEIXEIRA, J.C.; Nutrição de ruminantes, Lavras, 1992
Top Related