
21. El mar y las costas de Cataluña ya notan el efecto...
19
Introducción A las incertidumbres que generalmente se asocian a los efectos del cambio climático en los ecosistemas, en el mar se añaden otras relacionadas con las características de los océanos respecto a las masas continentales. En efecto, el impacto del cambio climático en el medio marino es difícil de determinar porque, para comenzar, el ambiente marino todavía nos es mucho menos conocido que el terrestre; en lo tocante a las series temporales de temperaturas, por ejemplo, rara vez se remontan a más de tres décadas y en general están limitadas a aguas litorales. A ello se añade que las características físicas, las respuestas biológicas y los equilibrios químicos de los océanos están sujetos a dinámicas que se extienden por buena parte del océano mundial (recuérdese la «cinta transportadora oceánica»; Broecker, 1991), desde la superficie hasta el fondo y por un medio que tiene una inercia (térmica, por ejemplo) notable. A ello se añade que apenas comenzamos a conocer la dinámica de fenómenos a macroescala oceánica, como el caso de El Niño, o el funcionamiento de áreas que hasta ahora no habían estado suficientemente estudiadas, como el océano Austral y la Antártida. Y por si todo esto no fuera poco, se suman a esta sinergia, los efectos de otros impactos antrópicos, que pueden enmascarar o potenciar algunos de estos efectos, desde la contaminación hasta la sobrepesca. En cualquier caso, ya comienzan a tenerse bastantes indicios, en algunos casos certezas (Levitus [et al.], 2000; Duarte y Tintoré, 2001; Cicerone [et al.], 2004; Harley [et al.], 2006), de que el impacto del cambio climático se notará en el mar, y que sus repercusiones serán aún más variadas, impactantes y preocupantes de lo que se preveía hace lustros (Fields [et al.], 1993). A continuación se revisarán estas repercusiones, tanto a escala oceánica global como centrada en el Mediterráneo, diferenciando las que afectan básicamente al medio físico y al paisaje de las que tienen impactos en los organismos y en las comunidades que forman; aunque como el propio cambio climático es de naturaleza física, ambos grupos de efectos se refuerzan mutuamente, o se confunden: un aumento del nivel del mar no solo provoca alteraciones en la línea de la costa, sino que también lo hace en las especies y en las comunidades litorales, y los cambios en la distribución y la abundancia relativa de determinadas especies se trasladan por toda la red trófica e inciden de manera inmediata en el aspecto del paisaje litoral y marino. Alteraciones en el medio físico y en el paisaje Temperatura A pesar de la inexistencia de series largas de temperaturas en casi todos los mares, hoy ya puede afirmarse que en el último medio siglo las aguas oceánicas superficiales (entre 0 y 300 m de profundidad) han experimentado un aumento medio de 0,31 ºC (Parrilla [et al.], 1994; Gouretski y Koltermann, 2007; MacKenzie y Schiedek, 2007). En el norte del Mediterráneo occidental, tres series de datos (de L’Estartit, de la isla del Levante, en las islas de Hyères, y en Vilafranca de Mar, las dos últimas en la costa francesa) se remontan a algo más de treinta años y son las más extensas que existen (Francour [et al.], 1994; Romano [et al.], 2000; Salat y Pascual, 2006). El resumen de lo que indican es claro: en este tercio de siglo, la temperatura de las aguas superficiales del Mediterráneo occidental septentrional ha subido en torno a 1 ºC de media; el mayor aumento se ha dado alrededor de los 20 21. El mar y las costas de Cataluña ya notan el efecto del cambio climático Joandomènec Ros, Universidad de Barcelona Impactos en los ecosistemas acuáticos El mar y las costas de Cataluña ya notan el efecto del cambio climático 259
Transcript of 21. El mar y las costas de Cataluña ya notan el efecto...

Introducción
A las incertidumbres que generalmente se asocian a los efectos del cambio climático en los ecosistemas, en el mar
se añaden otras relacionadas con las características de los océanos respecto a las masas continentales. En efecto, el
impacto del cambio climático en el medio marino es difícil de determinar porque, para comenzar, el ambiente marino
todavía nos es mucho menos conocido que el terrestre; en lo tocante a las series temporales de temperaturas, por
ejemplo, rara vez se remontan a más de tres décadas y en general están limitadas a aguas litorales.
A ello se añade que las características físicas, las respuestas biológicas y los equilibrios químicos de los océanos están
sujetos a dinámicas que se extienden por buena parte del océano mundial (recuérdese la «cinta transportadora
oceánica»; Broecker, 1991), desde la superficie hasta el fondo y por un medio que tiene una inercia (térmica, por
ejemplo) notable. A ello se añade que apenas comenzamos a conocer la dinámica de fenómenos a macroescala
oceánica, como el caso de El Niño, o el funcionamiento de áreas que hasta ahora no habían estado suficientemente
estudiadas, como el océano Austral y la Antártida. Y por si todo esto no fuera poco, se suman a esta sinergia,
los efectos de otros impactos antrópicos, que pueden enmascarar o potenciar algunos de estos efectos, desde la
contaminación hasta la sobrepesca.
En cualquier caso, ya comienzan a tenerse bastantes indicios, en algunos casos certezas (Levitus [et al.], 2000;
Duarte y Tintoré, 2001; Cicerone [et al.], 2004; Harley [et al.], 2006), de que el impacto del cambio climático
se notará en el mar, y que sus repercusiones serán aún más variadas, impactantes y preocupantes de lo que se
preveía hace lustros (Fields [et al.], 1993). A continuación se revisarán estas repercusiones, tanto a escala oceánica
global como centrada en el Mediterráneo, diferenciando las que afectan básicamente al medio físico y al paisaje
de las que tienen impactos en los organismos y en las comunidades que forman; aunque como el propio cambio
climático es de naturaleza física, ambos grupos de efectos se refuerzan mutuamente, o se confunden: un aumento
del nivel del mar no solo provoca alteraciones en la línea de la costa, sino que también lo hace en las especies y
en las comunidades litorales, y los cambios en la distribución y la abundancia relativa de determinadas especies se
trasladan por toda la red trófica e inciden de manera inmediata en el aspecto del paisaje litoral y marino.
Alteraciones en el medio físico y en el paisaje
Temperatura
A pesar de la inexistencia de series largas de temperaturas en casi todos los mares, hoy ya puede afirmarse que en
el último medio siglo las aguas oceánicas superficiales (entre 0 y 300 m de profundidad) han experimentado un
aumento medio de 0,31 ºC (Parrilla [et al.], 1994; Gouretski y Koltermann, 2007; MacKenzie y Schiedek, 2007).
En el norte del Mediterráneo occidental, tres series de datos (de L’Estartit, de la isla del Levante, en las islas de
Hyères, y en Vilafranca de Mar, las dos últimas en la costa francesa) se remontan a algo más de treinta años y son
las más extensas que existen (Francour [et al.], 1994; Romano [et al.], 2000; Salat y Pascual, 2006). El resumen
de lo que indican es claro: en este tercio de siglo, la temperatura de las aguas superficiales del Mediterráneo
occidental septentrional ha subido en torno a 1 ºC de media; el mayor aumento se ha dado alrededor de los 20
21. El mar y las costas de Cataluña ya notan el efecto del cambio climáticoJoandomènec Ros, Universidad de Barcelona
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
259

m de profundidad (1,2 ºC) e incluso en las aguas situadas a 80 m la temperatura ha subido unos 0,7 ºC (figura 1).
A causa de la gran inercia térmica del mar, en especial en aguas profundas, el efecto del calentamiento es notable
y sus impactos pueden ser muy importantes, tanto en lo relativo al aumento de volumen de la masa de agua y a
la disminución de la densidad del agua del mar (y a su papel en la circulación geostrófica) como a la fisiología, la
fenología, la ecología y la distribución de los organismos marinos (véase el apartado 2).
pH
En el mar, el pH es ligeramente alcalino y bastante constante, entre 8 y 8,3, y solo se producen desviaciones de este
rango en función de la variación de la concentración de dióxido de carbono y de oxígeno: en aguas superficiales
bien iluminadas, cuando hay poco CO2 (porque lo usan las algas como materia prima para la fotosíntesis) y
mucho O2 (producido por estas plantas), el pH puede ascender ligeramente, hasta 9, y alcalinizar el agua del mar.
En aguas profundas, donde la materia producida en superficie se va descomponiendo y, al hacerlo, consume
oxígeno, el pH puede descender a 7,6. Esta gama no es tan extensa como en las aguas continentales.
El pH depende de la cantidad de CO2 del agua, en equilibrio con la de la atmósfera. El dióxido de carbono se
disuelve bien en agua (donde su concentración es unas cien veces superior a la de la atmósfera) y forma ácido
carbónico, que se disocia rápidamente en ión bicarbonato y, si las condiciones son alcalinas, en carbonato,
liberando hidrogeniones en cada caso; el carbonato, en condiciones más ácidas, vuelve a bicarbonato; es el
sistema carbónico-carbonatos, responsable del equilibrio dinámico del carbono disuelto. Si, alcanzado un
determinado equilibrio, la cantidad de CO2 aumenta en el agua, crece la acidez (la concentración de hidrogeniones)
e inmediatamente se favorece que los hidrogeniones se combinen con los carbonatos y se pase de carbonato a
bicarbonato, y de bicarbonato a ácido carbónico. Si la cantidad de dióxido de carbono disminuye, el sistema lo
compensa descomponiendo bicarbonato en carbonato más hidrogeniones y manteniendo el equilibrio.
Figura 1. Aumento de la temperatura del agua a distintas profundidades en dos localidades del Mediterráneo noroccidental, L’Estartit (EST) y Vilafranca de Mar (VIL, Alpes Marítimos), en el transcurso de casi un tercio de siglo (Fuente: Salat y Pascual, 2006).
260
tendencia en 29 años (°C)
pro
fun
did
ad (
m)
EST
VIL

La alcalinidad o acidez de las aguas naturales reviste importancia porque de ella depende el equilibrio entre
precipitación y disolución de algunos compuestos, y la capacidad de la propia agua para mantener el equilibrio
en cuestión de disociación iónica, la capacidad tampón. Cuando el carbonato de calcio precipitado (en forma de
calcita o aragonita) en el caparazón de un molusco o de un crustáceo o en el esqueleto de una madrépora coralina
pasa a bicarbonato, la estructura se disuelve. La capacidad de mantener en las aguas estructuras esqueléticas a
base de carbonato depende, por tanto, del pH. En ciertas condiciones naturales (aguas frías, polares o profundas)
es difícil formar caparazones o esqueletos calcáreos: no hay corales polares o abisales, los moluscos no tienen
concha o la tienen delgada (o blanda), etc. La situación es exactamente la contraria para los esqueletos formados
por silicio: en estos casos, las aguas frías propician la precipitación de silicatos y las cálidas, su disolución.
Como consecuencia del incremento de la concentración de CO2 en la atmósfera por causas antrópicas, también ha
aumentado la entrada de este gas en el mar y ha comenzado a acidificarlo, entre 0,1 y 0,3 de pH (Caldeira y Wickett,
2003; Sabine [et al.], 2004; Orr [et al.], 2005; Blackford y Gilbert, 2007; Gazeau [et al.], 2007). Si no se actúa por
reducir la entrada de CO2 en los océanos, el pH del agua de mar puede reducirse en 0,5 unidades a finales del siglo
XXI (Royal Society, 2005). (El mismo calentamiento del agua de mar está ralentizando la entrada de más CO2 en los
océanos, puesto que la solubilidad de este gas y otros es función inversa de la temperatura; Lefèvre [et al.], 2004.)
La acidificación del agua marina puede comportar un trastorno enorme a muchos organismos de una gran
importancia ecológica y económica. Entre los planctónicos destacan los cocolitóforos, los foraminíferos y los
terópodos (uno de los alimentos predilectos de las ballenas; Riebesell [et al.], 2004). Muchos organismos bentónicos,
moluscos, crustáceos y equinodermos, entre otros, tienen esqueletos calcáreos (Bibby [et al.], 2007; Gazeau [et
al.], 2007; Miles [et al.], 2007). No obstante, quizá los más afectados serán los corales formadores de arrecifes,
en especial los de aguas superficiales y cálidas (Hughes [et al.], 2003; Baker [et al.], 2004; Hoegh-Guldberg
[et al.], 2007; Turley [et al.], 2007; Fine y Tchernov, 2007; NOAA, 2008). Sus esqueletos, a cuya construcción
contribuyen las algas simbiontes, encontrarán dificultades para mantenerse intactos en aguas más ácidas, y la
gran biodiversidad que hospedan los arrecifes coralinos resultará gravemente perjudicada.
Algunos de estos efectos ya han comenzado a notarse en alta mar, si bien con resultados contradictorios (últimamente
hay indicios de que las aguas más ácidas propician la proliferación de cocolitóforos; Iglesias-Rodríguez [et al.],
2008), y tanto los experimentos de laboratorio como el estudio de fuertes gradientes de pH naturales (alrededor
de surgencias de gases en fondos de regiones volcánicas del Mediterráneo; Hall-Spencer [et al.], 2008) indican un
impacto evidente en las algas calcáreas, los madreporarios y los equinodermos, entre otros.
Determinadas comunidades mediterráneas dependientes de organismos calcificados (el trottoir mediolitoral de
Lithophyllum byssoides y, en especial, la comunidad circalitoral de coralígeno; Ros [et al.], 1985; Ballesteros,
2006) pueden resultar profundamente afectadas. Se prevé que la capacidad tampón de los océanos pueda
quedar afectada a medio plazo.
Nutrientes
El cambio climático en sí no implica variaciones en la cantidad de nutrientes del mar y el litoral, si bien algunos
procesos generados o favorecidos por dicho cambio pueden comportar modificaciones en el régimen habitual de
aportación de nutrientes y, por tanto, en la producción primaria; y el descenso del pH puede movilizar determinadas
formas iónicas raras en la alcalinidad normal del océano. En los escenarios de aumento del nivel del mar (véase el
apartado 1.6) se calcula que bastantes comunidades litorales, tanto sumergidas (praderas de fanerógamas, etc.)
como emergidas (vegetación de marismas, etc.), sufrirán una reducción de la radiación incidente (en el límite inferior
las primeras) y/o una inmersión continua (las segundas), cosa que propiciará la muerte de las plantas constituyentes
y la descomposición de la materia orgánica, con la consiguiente liberación de nutrientes.
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
261

No obstante, todo ello sucederá a un ritmo bastante lento para que no comporte ningún impacto notable, y lo mismo
puede afirmarse de los episodios de lluvias torrenciales, que podrán implicar avenidas más frecuentes y aportación
de sedimentos y nutrientes al litoral. Otro aspecto, aún poco estudiado, es el papel que pueden desempeñar los
organismos fijadores de nitrógeno atmosférico en un océano más rico en dióxido de carbono; los primeros indicios
apuntan hacia un aumento de la entrada de nitrógeno en el mar (Barcelos e Ramos [et al.], 2007; Levitan [et al.],
2007). Tal como se ha podido establecer en comunidades terrestres, para el bacterioplancton y el fitoplancton marinos
la subida de temperatura implica un mayor consumo de dióxido de carbono y de oxígeno (Vázquez-Domínguez [et
al.], 2007), de manera que no está claro que la producción primaria de los océanos en general y del mar Mediterráneo
en concreto se pueda beneficiar de este incremento de CO2, de nitrógeno fijado (ni si es cierto que aumentarán los
episodios de vientos del sur, con carga de polvo sahariano; véase el apartado 1.4), de fósforo y de hierro.
Lo que sí es seguro es que un aumento del ritmo de deshielo de los casquetes polares y una mayor frecuencia de
desprendimiento de los icebergs conllevará un aumento de la entrada de sedimentos y nutrientes en los mares de latitudes
altas (Smith [et al.], 2007), pero ignoramos qué impacto puede tener ello allende los mares regionales afectados.
Regímenes de vientos
Se ha sugerido que una de las alteraciones asociadas al cambio climático global será el aumento de episodios
catastróficos de circulación atmosférica, desde los ciclones y los huracanes tropicales hasta los vientos regionales
asociados al tiempo meteorológico. Muchas incertidumbres rodean aún este punto, pero cabe recordar al menos
tres aspectos de la biología marina en los cuales los vientos desempeñan un papel destacado.
En primer lugar, los afloramientos inducidos o reforzados por vientos litorales (como los del golfo de León, por indicar
el más cercano) pueden verse afectados en cierta medida por cambios eventuales en el régimen de vientos (y por el
calentamiento concomitante del agua). En segundo lugar, y con independencia de cuál sea la causa de la sobreabundancia
de medusas y sifonóforos que últimamente se registra cerca del litoral (Mills, 2001; Attrill [et al.], 2007; Ros, 2007), los
organismos del macroplancton y en especial del neuston dependen enormemente de las corrientes y de los vientos para
sus movimientos poblacionales pasivos. Cuando soplan vientos de levante, el litoral catalán recibe un mayor número
de estos visitantes de alta mar que en otros regímenes de vientos, con los problemas asociados conocidos (Gili y Pagès,
2005). Por último, los vientos procedentes del norte de África aportan al Mediterráneo (y a otras áreas estudiadas,
como las Canarias) polvo sahariano, con una carga notable de materiales susceptibles de actuar como nutrientes para
los productores planctónicos (fósforo, hierro) o, alternativamente, de gérmenes (véase el apartado 2.4).
Regímenes de corrientes
Más preocupante que el posible efecto en el régimen de vientos resulta el impacto del cambio climático en las
corrientes marinas. Estas corrientes son más el resultado del equilibrio dinámico entre masas de agua según su
densidad (circulación geostrófica) que el efecto de los vientos en el océano, y está demostrado que el calentamiento
global provoca que la densidad del agua marina cambie regionalmente, en unas áreas con una mayor aportación de
agua dulce (por fusión del hielo terrestre; Holland [et al.], 2007; Howatt [et al.], 2007) y en otras por una reducción
de estas entradas (aumento de la evaporación y descenso de la precipitación). Ello ha provocado que el énfasis del
impacto más notable del cambio climático en el mar se haya puesto en la posible alteración del sistema de corrientes
tal como lo conocemos ahora (Broecker, 1991; Alley, 2007), con consecuencias bruscas y catastróficas.
Efectivamente, el escenario que plantea de manera esquemática la película Pasado mañana (The day after
tomorrow) (2004), sobre una alteración repentina del sistema de circulación superficial y profunda del Atlántico
Norte, con efectos inmediatos en el clima regional y la desviación de corrientes (entre ellas la del Golfo, que
262

calienta Europa), no parece imposible, y menos a tenor de los episodios de cambios climáticos acaecidos en
alguna época histórica (la Pequeña Edad de Hielo). Estas y otras modificaciones (por ejemplo, las de las corrientes
del Mediterráneo) pueden modelarse, pero el margen de incertidumbre es demasiado grande para hacer
afirmaciones concretas. Muchas especies aprovechan las corrientes para sus movimientos migratorios, que, en
consecuencia, pueden alterarse.
Subida del nivel del mar
Seguramente uno de los efectos más conocidos del cambio climático sea la subida del nivel del mar como
consecuencia de, como mínimo, dos efectos: la dilatación de la masa de agua por efecto del calentamiento (veáse
el apartado 1.1) y la aportación diferencial del agua continental por fusión incrementada del hielo de los glaciares,
los casquetes polares y el hielo terrestre en general (la del hielo marino no hace variar el nivel del mar). Según las
distintas proyecciones y su revisión constante, en los escenarios más negativos la subida estimada del nivel del mar
se sitúa en torno al metro a lo largo del siglo XXI, equivalente al total de lo que ha ocurrido en las costas europeas y
norteamericanas, donde los registros son fiables, durante los últimos dos o tres siglos (Rahmstorf, 2007).
Aunque este aumento pueda parecer moderado, muchas regiones de costas bajas (áreas de deltas y estuarios,
islas oceánicas coralinas, etc.) están amenazadas tanto por la subida del nivel del mar como por la incidencia de
episodios de tormentas y otras adversidades cuyo aumento también se ha pronosticado. En Cataluña, los deltas
(sobre todo el del Ebro) y las zonas de humedales y marismas, además de las urbanizadas cercanas a la línea
litoral, peligran en caso de que se materialice o agrave este escenario. Pese a que se han preparado simulaciones
bastante realistas del efecto que esta subida del nivel del mar puede tener en las áreas litorales más pobladas
y urbanizadas (como la que hizo Greenpeace en el 2007, que puede llevar a esta ONG ante los tribunales por,
supuestamente, atentar contra el futuro económico de la región de Murcia), simulaciones que destacan los daños
a los bienes urbanos (McGranahan [et al.], 2007; Bosello [et al.], 2007; etc.; véase el apartado 2.9), seguramente
el mayor impacto sea ecológico.
Efectivamente, la subida del nivel del mar inundará áreas naturales que hoy en día (aunque dañadas en parte
por las actividades humanas) proporcionan servicios ecosistémicos notables: la filtración de las aguas fluviales
contaminadas que realizan los humedales; la elevada producción primaria de las poblaciones de algas fotófilas
y de las praderas de fanerógamas marinas (especialmente, en el Mediterráneo, de Posidonia oceanica), y el
papel de áreas de cría y alimentación para aves, peces e invertebrados en general de estas comunidades, que, si
quedan cubiertas por las aguas en ascenso, difícilmente se podrán restablecer en áreas ahora ya muy alteradas
o urbanizadas.
Cambios en la biota y en las comunidades
De un modo casi imperceptible hemos pasado de las repercusiones para el entorno físico a las repercusiones
para las especies de organismos y las comunidades que forman. A continuación se analizan de una manera
más concreta estos efectos más claramente ecológicos y las consecuencias para la biodiversidad y para nuestra
especie.
Cambios en las pautas de distribución geográfica y batimétrica
El primer efecto que detectamos a la hora de plantear el futuro de las poblaciones bacterianas, vegetales y animales
en un mar más cálido es que algunas especies podrán adaptarse mejor o peor a los cambios (básicamente, al
aumento de la temperatura de las aguas) y mantendrán o expandirán su distribución actual, mientras que otras
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
263

no lo harán y reducirán esta distribución e incluso desaparecerán, a ritmos que solo se han podido establecer
para contadas especies (Bames [et al.], 2001; Sabatés [et al.], 2006; Mackas [et al.], 2007; Lima [et al.], 2007;
Mackenzie [et al.], 2007; Mieszkowska [et al.], 2007; Jagtap y Nagle, 2007; Simmonds y Isaac, 2007; etc.).
En términos generales (Hughes, 2000), las especies monotípicas y adaptadas estrictamente a determinadas condiciones
térmicas serán las más afectadas, mientras que las monotípicas con más facilidad de adaptación y, en especial, las
politípicas, divididas en diferentes poblaciones adaptadas a regímenes térmicos también diferentes, serán las que se
podrán adaptar con menos dificultades al calentamiento generalizado de su hábitat, gracias a la resistencia y a la
expansión de sus poblaciones más termófilas, en detrimento de las que lo son menos. Los sistemas de dispersión de
huevos, larvas y propágulos en general, muy ligados al sistema oceánico de corrientes, de alcance mundial, deberá
favorecer más que en los ecosistemas terrestres esta redistribución según los nuevos escenarios térmicos.
Actualmente, algunas especies de amplia distribución, que en mares más fríos (como los marginales del océano
Atlántico) son propias de aguas superficiales, se suelen encontrar en el Mediterráneo en aguas profundas; puesto que
el Mediterráneo tiene temperaturas más altas tanto a lo largo del ciclo anual como siguiendo un perfil batimétrico
(la temperatura profunda de este mar no baja nunca de los 12 ºC, en clara contraposición con las aguas oceánicas,
que a partir de algunos centenares de metros ya alcanzan los 4 ºC), estas especies buscan su óptimo térmico, que
puede diferir mucho en poblaciones de la misma especie a un lado y otro del estrecho de Gibraltar, por ejemplo. Es
evidente que este «hundimiento» en busca de temperaturas más propicias no podrá producirse mucho más allá si la
masa de agua en general, y no solo las aguas superficiales, se calienta. Además, será necesario que las especies de
las cuales dependen las que modifiquen su hábitat también lo hagan; en caso contrario se pueden dar situaciones
de falta total o parcial de correspondencia entre depredador y presa, o pueden resultar alteradas otras relaciones
(Durant [et al.], 2007; MacLeod [et al.], 2007; Freitas [et al.], 2007; etc.).
Asimismo, históricamente en el Mediterráneo ha habido un claro gradiente latitudinal: las especies más termófilas,
tanto las pelágicas como las bentónicas, se encontraban limitadas a las costas norteafricanas y levantinas
(Mediterráneo oriental), mientras que las más capaces de tolerar temperaturas bajas eran más propias de las
costas europeas, en especial del Mediterráneo occidental. Era posible encontrar adecuadamente distribuidas
en función de los gradientes térmicos mencionados a las especies que habían entrado en el Mediterráneo,
una vez restablecida la conexión de este mar con el Atlántico hace unos cinco millones de años, en las épocas
glaciales (especies de afinidades templadas y boreales, más propias de las aguas septentrionales y profundas del
Mediterráneo) y las que habían entrado en épocas interglaciales (especies de afinidades tropicales y subtropicales,
más propias de las aguas meridionales y superficiales del Mediterráneo).
Últimamente, algunas de estas especies mediterráneas de afinidades tropicales o termófilas han ampliado
su distribución: el raor (Xyrichtys novacula), el pez verde (Thalassoma pavo) y el mero moreno (Epinephelus
marginatus), por ejemplo, entre los peces, son ahora comunes en aguas catalanas, mientras que hace pocas
décadas eran raros y únicamente se encontraban en abundancia en áreas más meridionales (por ejemplo, en
las Islas Baleares). Parablennius pilicornis (un blénido), Scorpaena maderensis (el rascacio) y Pomadasys incisus
(pez roncador), entre otros, se han detectado recientemente en las costas catalanas y francesas, mucho más
septentrionales que su área de distribución habitual. Es muy probable que esta situación sea más general, pero
hay pocas especies tan fáciles de observar y tan conocidas como los peces.
Asimismo, en las aguas más cálidas de las Baleares, en concreto, se han encontrado en los últimos tiempos especies
ajenas procedentes de la migración lessepsiana (Por, 1978) o de otros orígenes (véase el apartado 2.6). Así ocurre
con algunas especies de peces (como Siganus rivulatus) y con muchas especies de algas (Asparagopsis taxiformis,
Womersleyella setacea, Lophocladia lallemandii, Caulerpa racemosa, C. taxifolia, Acrothamnion preissii, etc.),
ausentes desde hace dos décadas y que hoy predominan en el paisaje submarino de algunos lugares de las Islas
(Ballesteros, com. pers.; Patzner, 1998; Ballesteros [et al.], 2007; etc.).
264

Uno de los pocos estudios que demuestran claramente el cambio en la distribución de una especie es el de
Sabatés [et al.] (2006) sobre la alacha (Sardinella aurita). El trabajo analiza los cambios en la abundancia y en la
distribución de esta especie termófila en el transcurso de las últimas décadas y establece una correlación positiva
entre las anomalías térmicas y las capturas: las abundancias de un año dependen de las temperaturas registradas
en la época de maduración del anterior. Simultáneamente, el límite septentrional de los huevos y las larvas de la
alacha ha pasado de la latitud aproximada del delta del Ebro de hace veinte años a la latitud de Blanes de hoy en
día: el calentamiento progresivo de las aguas superficiales ha permitido la extensión hacia el norte de la especie,
y no debe de ser la única (Brunel y Boucher, 2007).
Sea como fuere, si las especies de afinidades tropicales podrán ampliar su área de distribución en un escenario
de calentamiento del Mediterráneo, ¿qué ocurrirá con las especies de afinidades boreales, constituyentes de
las llamadas «faunas frías» (Pérès y Picard, 1964), muchas de las cuales se encuentran arrinconadas en aguas
profundas o localidades muy septentrionales de este mar, donde forman reductos o relictos? No es difícil
pronosticar que el ritmo de reducción de su área de distribución continuará, un proceso que seguramente se
inició con la propia introducción en aguas mediterráneas, hace ahora varios millones de años, desde las frías
aguas del Atlántico durante los períodos glaciales.
Es lo que parece ocurrirles, entre otras especies, a los moluscos Cyprina (Arctica) islandica y Buccinum undatum y
a los peces Platichthys flesus y Raja clavata. Algunos descensos importantes en las poblaciones de determinadas
especies (por ejemplo, de fucales; Thibaut [et al.], 2005) podrían deberse, entre otros impactos, al calentamiento
creciente de nuestras costas.
Cambios en la fenología
El cambio térmico no solo afecta a la distribución: algunas especies ven alteradas pautas biológicas y ecológicas
de toda clase, que van desde la variación en la disponibilidad del alimento hasta la proporción entre los sexos,
que en los peces depende mucho de la temperatura (Mora y Ospina, 2001; Navarro [et al.], 2005). El efecto
demográfico a medio plazo de estos cambios puede ser harto importante. Asimismo, tal como se ha establecido
en los ecosistemas terrestres, en los marinos puede darse un decalaje en las épocas de freza y reproducción en las
especies con una fuerte dependencia de la temperatura, en lugar de seguir una pauta más ligada al calendario
(Høye [et al.], 2007; Greenwood, 2007; Hawkes [et al.], 2007; etc.).
Uno de los cambios más espectaculares, además de los relacionados con la variación de la proporción sexual en
algunas especies de peces, corresponde a la parada nupcial y el desove del mero moreno (Epinephelus marginatus),
observados por vez primera en aguas catalanas (de las islas Medas) hace una década (figura 2; Zabala [et al.], 1997).
La reproducción de esta especie termófila no se había registrado nunca en una latitud tan alta, donde solo llegaban
alevines salidos del huevo en aguas meridionales del Mediterráneo. Y seguro que este no es el único caso.
Estrés fisiológico
Los organismos no responden como máquinas de todo o nada al cambio climático: la temperatura me conviene y
me quedo o no me conviene y me voy. Las nuevas condiciones pueden comportar modificaciones en la fisiología,
desde una menor cantidad de oxígeno disponible para las actividades normales (Morán [et al.], 2006; Portner y
Knust, 2007; Michaelidis [et al.], 2007; Anestis [et al.], 2007; etc.) hasta una menor capacidad competitiva con
relación a las otras especies en las nuevas condiciones, que se añaden a las limitaciones en las disponibilidades
alimentarias ya mencionadas. La adaptación a las nuevas condiciones, por tanto, comporta cambios notables en
la biología de las especies y en la ecología de las comunidades.
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
265

Enfermedades
Algunos de estos cambios caen dentro del apartado de las enfermedades: el estrés fisiológico debilita a unas
especies y aumenta su morbididad, y las nuevas condiciones facilitan la proliferación de otras causantes de
enfermedades. Buenas parte de los casos de enfermedades y mortalidades masivas (véase el apartado 2.6) están
relacionados con altas temperaturas episódicas o duraderas (Harvell [et al.], 1999; Lafferty [et al.], 2004; Bruno
[et al.], 2007; Bally y Garrabou, 2007; etc.). Cabe interpretar «enfermedades» en un sentido muy amplio, que
engloba desde la proliferación de gérmenes (a causa del aumento de la temperatura o de otros efectos sinérgicos,
como, por ejemplo, el incremento de nutrientes; véase el apartado 1.3) hasta la sobreabundancia de especies
Figura 2. Representación esquemática de un episodio de freza del mero moreno, según lo observaron y describieron por vez primera en las islas Medas Zabala [et al.] (1997). No todas las consecuencias del cambio climático en el Mediterráneo son tan interesantes.
266
termoclina

tóxicas, básicamente dinoflageladas y otras algas causantes de proliferaciones nocivas (Gómez y Claustre, 2001;
Vila y Masó, 2005). Las características ambientales que facilitan esta proliferación (aguas tranquilas, cálidas y ricas
en nutrientes) parecen ser cada vez más frecuentes en áreas abrigadas del litoral mediterráneo.
Huelga decir que el uso de antibióticos, una práctica común en la acuicultura extensiva, ha comportado la
generación de resistencia a muchos patógenos, cosa que se suma a los efectos del calentamiento citados
anteriormente, en un claro ejemplo de sinergia entre diferentes impactos antrópicos (véase el apartado 2.6).
Episodios de mortalidades en masa y a gran escala
De todos los efectos catastróficos relacionados con el cambio climático, el que hasta el momento parece mejor
establecido y ha producido unos resultados más espectaculares (por negativos) y catastróficos (por su alcance)
es la serie de condiciones oceanográficas y de otro tipo que provocan episodios de mortalidad en masa a gran
escala. Estos episodios no son desconocidos en el medio marino (Korringa, 1973) y últimamente han proliferado
como consecuencia de las fuertes presiones sobre los arrecifes de coral y otros ecosistemas oceánicos (Hughes [et
al.], 2003; McClanahan [et al.], 2007; Hoegh-Guldberg [et al.], 2007; etc.), pero en el Mediterráneo noroccidental
nunca se habían documentado tan bien como en el transcurso de la última década.
En las postrimerías del verano del 1999 se observó un episodio de mortalidad en masa sin precedentes a lo
largo de centenares de kilómetros de las costas de Provenza y Liguria (Perez [et al.], 2000; Cerrano [et al.], 2000;
Romano [et al.], 2000; Garrabou [et al.], 2001; etc.). Las especies afectadas fueron sobre todo invertebrados
sésiles y suspensívoros de la comunidad del coralígeno (Ros [et al.], 1985; Ballesteros, 2006), situados entre los 10
y los 40 m de profundidad: esponjas (Hippospongia communis y Spongia officinalis, etc.), cnidarios (en especial
los antozoos Corallium rubrum, Paramuricea clavata, Eunicella cavolinii, E. singularis y Cladocora caespitosa),
moluscos bivalvos, briozoos y ascidias. El sentido de propagación de la mortalidad fue de este a oeste. El resultado
fue una pérdida de biomasa estimada en el 50 % de la propia de esta comunidad (entre 150 y 1200 g (p. s.) m–2),
debida a la necrosis de los tejidos de las colonias, con posterior desprendimiento u ocupación de las colonias por
parte de las especies epibiontes.
El año 2003 se repitió este episodio, con una notable expansión hacia el sur de las áreas afectadas: casi toda la
costa italiana desde el mar Tirreno hasta el golfo de Nápoles, Córcega, el estrecho de Bonifacio, Cataluña, Menorca
y los Columbretes (figura 3). En el 2006 se registró un nuevo episodio de mortalidad masiva de suspensívoros,
aparentemente con un impacto menor que el de tres años atrás. El episodio del 1999 pudo detectarse y evaluarse
porque en algunas áreas protegidas de la zona se estaban llevando a cabo estudios de seguimiento de las
comunidades, y en los años posteriores se procedió a realizar el seguimiento del impacto y de la recuperación
posterior de las poblaciones de las especies de suspensívoros afectadas. Puesto que las mortalidades se han
repetido en años sucesivos, se ha puesto en funcionamiento un programa internacional de detección precoz,
evaluación e identificación de las causas (Medchange).
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
267
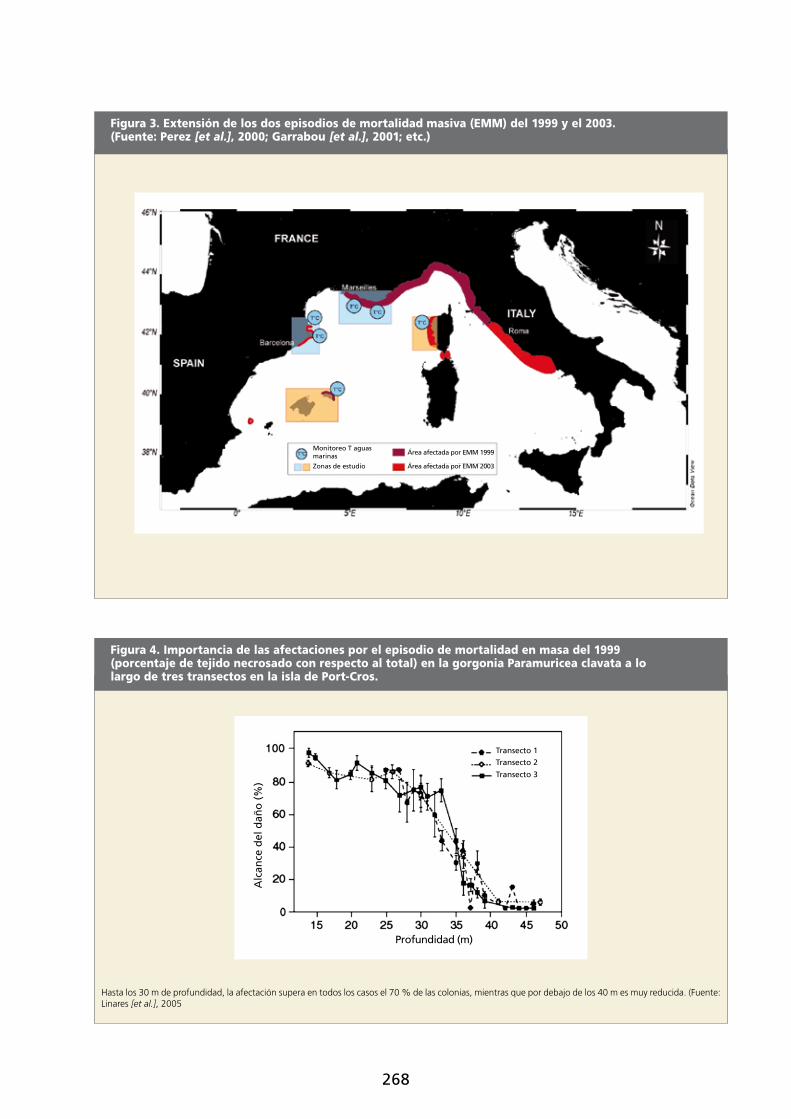
Figura 3. Extensión de los dos episodios de mortalidad masiva (EMM) del 1999 y el 2003. (Fuente: Perez [et al.], 2000; Garrabou [et al.], 2001; etc.)
Figura 4. Importancia de las afectaciones por el episodio de mortalidad en masa del 1999 (porcentaje de tejido necrosado con respecto al total) en la gorgonia Paramuricea clavata a lo largo de tres transectos en la isla de Port-Cros.
Hasta los 30 m de profundidad, la afectación supera en todos los casos el 70 % de las colonias, mientras que por debajo de los 40 m es muy reducida. (Fuente: Linares [et al.], 2005
268
Profundidad (m)
Transecto 2
Transecto 3
Transecto 1
Alc
ance
del
dañ
o (
%)
Monitoreo T aguas marinas
Zonas de estudio Área afectada por EMM 2003
Área afectada por EMM 1999

El impacto de estos episodios ha sido enorme, tanto por alcance geográfico como por el número de especies
afectadas, por la incidencia en las poblaciones (en total se calcula que la pérdida ha representado el 50 % de
la biomasa de estas especies, con un claro gradiente batimétrico, ya que las poblaciones más superficiales han
sido las más afectadas: figura 4; Perez [et al.], 2000; Cerrano [et al.], 2000; Romano [et al.], 2000; Garrabou
[et al.], 2001; Linares [et al.], 2005), por la dificultad de recuperación (algunas de estas especies son de biología
parsimoniosa: la gorgonia Paramuricea clavata, por ejemplo, crece cerca de 1 cm anual, de manera que recuperar
la complejidad estructural de las comunidades de coralígeno afectadas, aunque sea solo parcialmente, puede
llevar décadas), por el impacto en la biodiversidad asociada a los «bosques» de suspensívoros afectados, etc.
El efecto no ha sido el mismo en todas las áreas geográficas ni en todas las poblaciones, ni en los diferentes años
en que se ha detectado el fenómeno (figura 5). La causa principal ha sido la prolongación, más allá de lo habitual,
del período de mantenimiento de la termoclina estival, que separa las aguas cálidas superficiales de las frías
profundas. Efectivamente, durante los estudios de los años mencionados, esta termoclina, en lugar de romperse y
permitir la mezcla de las dos masas de agua superficial y profunda al final del verano, como acostumbra a ocurrir,
se mantuvo aproximadamente otro mes, con lo cual las aguas superficiales del Mediterráneo noroccidental
disfrutaron de una temperatura excepcionalmente elevada y estable (23-24 °C) a lo largo de toda la columna de
agua, hasta los 40 m de profundidad.
El efecto sobre las especies de suspensívoros puede haber sido directo (por una inadaptabilidad a las condiciones
térmicas estivales prolongadas, quizá mediante un mecanismo fisiológico, véase el apartado 2.3) o indirecto (por
la falta de alimento [plancton] que la estratificación y, por tanto, el retraso en la mezcla otoñal impide proliferar);
por el efecto de gérmenes patógenos favorecidos por las temperaturas elevadas (Martin [et al.], 2002; Bally y
Garrabou, 2007); por el efecto sinérgico de tóxicos en el agua procedentes de la contaminación, etc.
Casi todas estas hipótesis tienen sus defensores y habrá que esperar a resultados más concluyentes para aceptarlas
o descartarlas, pero la repercusión de estos episodios es innegable y catastrófica. Como sea que los diferentes
escenarios de cambio climático plantean un aumento de la temperatura a lo largo del siglo XXI, los episodios de
calentamiento de las aguas superficiales de Mediterráneo pueden dejar de ser fenómenos excepcionales y volverse
habituales, con las implicaciones de pérdida regional de biodiversidad previsibles (Coma [et al.], 2006; etc.).
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
269
Figura 5. Frecuencia media de colonias afectadas de las cuatro especies de gorgonias afectadas por el episodio de mortalidad en masa del 2003.
Zona 1: Baleares; zona 2: Cataluña y Cataluña Norte; zona 3: Alpes Marítimos; zona 4: Liguria; zona 5: Córcega y norte de Cerdeña; zona 6: golfos de Gaeta y de Nápoles. (Fuente: Garrabou [et al.], 2001; Coma [et al.], 2006)
Frec
uen
cia
(%)
Zona 1 Zona 2 Zona 3 Zona 4 Zona 5 Zona 6

Sinergia con otros impactos (contaminación térmica, especies alóctonas, etc.)
Ya se han mencionado las dudas en torno a las causas últimas de las mortalidades catastróficas inducidas por
el calentamiento prolongado del Mediterráneo noroccidental: pueden ser diversas. Lo mismo puede afirmarse
de otras situaciones en las cuales el calentamiento global propicia y magnifica otros impactos; a continuación
citaremos un par de ellas. Las áreas litorales cerradas (puertos, lagunas, etc.), con aguas más cálidas, son reservas
de especies exóticas, entre las cuales figuran las de afinidades tropicales, aportadas en el pasado o en la actualidad
mediante el transporte en el agua de lastre de los barcos o en el fouling de sus quillas. La subida de la temperatura
de las aguas del Mediterráneo puede comportar la «liberación» de algunas de estas especies arrinconadas en
las citadas áreas restringidas, que podrán ocupar áreas más extensas. Así ha ocurrido recientemente con el
coral Oculina patagonica, hasta hace poco restringido al puerto de Alicante y ahora extendido por los fondos
superficiales de muchas localidades del Mediterráneo occidental. Una situación similar se produce con las entradas
en el mar de agua sobrecalentada procedente de los sistemas de refrigeración de centrales térmicas y nucleares,
que son también núcleos potenciales de dispersión de especies termófilas, aunque la inercia térmica del mar es
muy grande y el alcance de este impacto, reducido.
Se ha aludido ya a la entrada en el Mediterráneo y la posterior expansión de especies exóticas lessepsianas o
de otros orígenes, una expansión que está favoreciendo el aumento de temperatura y puede hacerlo más en el
futuro inmediato (Occhipinti-Ambrogi, 2007).
También cabe hacer referencia a la situación de erosión de la biodiversidad, de agotamiento de las reservas
pesqueras, y de alteración de las aguas y de los fondos por la contaminación y otras actividades de origen
antrópico, que se añaden sinérgicamente a los impactos derivados del cambio climático. El resultado es una
fuerte repercusión en los hábitats y en las especies.
Impactos en la biodiversidad, en los servicios ecosistémicos y en los recursos apropiados para el ser
humano
Todos estos impactos, entre otros, se traducen en una pérdida notable de la biodiversidad, si bien algunos
fenómenos (como la incorporación de especies alóctonas) inducirían a pensar lo contrario. La notable biodiversidad
marina del Mediterráneo (Pérès y Picard, 1964; Ros [et al.], 1985; Giaccone y Geraci, 1989; Bianchi y Morri,
2000; Boudouresque, 2004; Boudouresque [et al.], 2004; etc.) experimenta en los últimos tiempos alteraciones
importantes como consecuencia del cambio climático y otras actividades antrópicas; algunas son de alcance
mundial y se producen a un ritmo conmensurado, mientras que otras son de alcance regional y muy rápidas.
Tales alteraciones poseen dos características comunes: a) la eliminación masiva de las especies más frágiles y
longevas, de biología parsimoniosa y más adaptadas, muchas de las cuales son endémicas y/o relictas, y la
persistencia y la expansión de especies banales, pioneras y en gran medida alóctonas; b) la «tropicalización» de
la biota: no solo las especies autóctonas favorecidas son las más termófilas, sino que tanto las que entran de
manera natural a través de Gibraltar como las migradoras lessepsianas, y buena parte de las que penetran gracias
al transporte humano (en el fouling o en el lastre de los barcos), son de afinidad subtropical o tropical.
Resulta difícil evaluar cuáles pueden ser los efectos de estos cambios en la biodiversidad a la escala de todo el
Mediterráneo, pero como se trata de un mar muy diverso, es seguro que el empobrecimiento de unos actores
y la expansión de los otros mediante impactos directos o inducidos, a través de efectos (Sala [et al.], 1998;
etc.) y de inversiones en los equilibrios poblacionales y ecosistémicos «naturales» (hasta donde sea posible
esta denominación; Jackson y Sala, 2001; Jackson [et al.], 2001), provocarán alteraciones importantes en la
biodiversidad y en la ecodiversidad (sensu Margalef, 1997).
270

Así está sucediendo en otros mares regionales (Jackson [et al.], 2001; Hughes [et al.], 2003; Sala y Knowlton,
2006; Clarke [et al.], 2007; Aronson [et al.], 2007; Jagtap y Nagle, 2007; etc.). El resultado en todos estos mares
es una simplificación de las comunidades marinas mediante diversos mecanismos: reducción acelerada de la
biodiversidad marina; hundimiento de poblaciones y extinciones de especies; homogeneización y banalización de
las comunidades; reducción de la estructura tridimensional biogénica; simplificación de las redes tróficas; dominio
de los controles desde abajo (bottom up) de las redes tróficas con relación a los controles desde arriba (top-
down); incremento de la inestabilidad y disminución de la capacidad de recuperación; reducción de la biomasa
total; aumento del cociente P/B; reducción de la producción total y de la disponible para los seres humanos.
Todo ello tendrá (ya está teniendo) consecuencias a escala ecosistémica, pero también en la provisión de servicios
ecosistémicos (Worm [et al.], 2007), algunos de los cuales son fundamentales, como la obtención de proteína en
forma de pescado y marisco (Cury [et al.], 2008).
Quizá el mar Mediterráneo no dejará de ser oligotrófico (Margalef, 1985) ni biodiverso, quizá no acabará
«tropicalizándose» (Bianchi, 2007), quizá las algas frondosas no serán sustituidas por corales (Ballesteros y Zabala,
1989), pero seguro que será muy distinto a como lo describieron los investigadores marinos de los siglos XIX y
XX y los poetas de todos los tiempos.
Referencias bibliográficas
Alley, R. B. Predictive ability of the North Atlantic “conveyor belt” hypothesis for abrupt climate change. Annual
Review of Earth and Planetary Sciences, (35): págs. 241-272, 2007.
Anestis, A.; lAzou, A.; Pörtner, H. O.; MichAelidis, B. Behavioral, metabolic, and molecular stress responses of
marine bivalve Mytilus galloprovincialis during long-term acclimation at increasing ambient temperature. American
Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 293(2): 2007, R911-R921.
Aronson, R. B.; thAtje, S.; clArke, A.; Peck, L. S.; BlAke, D. B.; WilgA, C. D.; seiBel, B. A. Climate change and invasibility
of the Antarctic benthos. Annual Review of Ecology, Evolution, and Systematics, (38): págs. 129-154, 2007.
Attrill, M. J.; Wright, J.; edWArds, M. Climate-related increases in jellyfish frequency suggest a more gelatinous
future for the North Sea. Limnology and Oceanography, 52(1): págs. 480-485, 2007.
BAker, A. C.; stArger, C. J.; McclAnAhAn, T. R.; glynn, P. W. Corals’ adaptive response to climate change. Nature,
(430): págs. 741, 2004.
BAllesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. Oceanography and
Marine Biology: An Annual Review, (44): págs. 123-195, 2006.
BAllesteros, E.; ceBrián, E.; Alcoverro, T. Mortality of shoots of Posidonia oceanica following meadow invasion
by the red alga Lophocladia lallemandii. Bot. Mar., (50): págs. 8-13, 2007.
BAlly, M.; gArrABou, J. Thermodependent bacterial pathogens and mass mortalities in temperate benthic
communities: a new case of emerging disease linked to climate change. Global Change Biology, 13(10): págs.
2078-2088, 2007.
BAMes, D. K. A.; crook, A.; o’Mohoney, M.; steicie, S.; MAguire, D. Sea temperature variability and Paracentrotus
lividus population fluctuations. Journal of the Marine Biological Association of the United Kingdom, (81): págs.
359-360, 2001.
BArcelos e rAMos, J.; BisWAs, H.; schulz, K. G.; lAroche, J.; rieBesell, U. Effect of rising atmospheric carbon dioxide
on the marine nitrogen fixer Trichodesmium. Global Biogeochemical Cycles, 21(20): art. GB2028, 2007.
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
271

BiAnchi, C. N. Biodiversity issues for the forthcoming tropical Mediterranean Sea. Hydrobiologia, 580(1): págs.
7-21, 2007.
BiAnchi, C. N.; Morri, C. Marine Biodiversity of the Mediterranean Sea: Situation, Problems and Prospects for
Future Research. Marine Pollution Bulletin, 40(5): págs. 367-376, 2000.
BiBBy, R.; cleAll-hArding, P.; rundle, S.; WiddicoMBe, S.; sPicer, J. Ocean acidification disrupts induced defences in
the intertidal gastropod Littorina littorea. Biology Letters, 3(6): págs. 699-701, 2007.
BlAckford, J. C.; gilBert, F. J. pH variability and CO2 induced acidification in the North Sea. Journal of Marine
Systems, 64(1-4): págs. 229-241, 2007.
Bosello, F.; roson, R.; tol, R. S. J. Economy-wide estimates of the implications of climate change: Sea level rise.
Environmental and Resource Economics, 37(3): págs. 549-571, 2007.
Boudouresque, C. F. Marine biodiversity in the Mediterranean: status of species, populations and communities.
Scientific Reports of Port-Cros National Park, (20): págs. 97-146, 2004.
Boudouresque, C. F.; Boury-esnAult, N.; hArMelin-vivien, M. «La biodiversitat marina: tendències, models y
amenaces». En: VILÀ, M.; RODÀ, F.; ROS, J. D. (ed.). Jornades sobre Biodiversitat y Conservació Biològica. Seminar
on Biodiversity and Biological Conservation, Barcelona: Institut d’Estudis Catalans, 2004, págs. 189-209 y 391-
410.
Broecker, W. S. The Great Ocean Conveyor. Oceanography, (4): págs. 79-89, 1991.
Brunel, T.; Boucher, J. Long-term trends in fish recruitment in the north-east Atlantic related to climate change.
Fisheries Oceanography, 16(4): págs. 336-349, 2007.
Bruno, J. F.; selig, E. R.; cAsey, K. S.; PAge, C. A.; Willis, B. L.; hArvell, C. D.; sWeAtMAn, H.; Melendy, A. M.
Thermal stress and coral cover as drivers of coral disease outbreaks. PLoS Biology, 5(5): art. e124, 2007.
cAldeirA, K.; Wickett, M. E. Anthropogenic Carbon and Ocean pH. Nature, (425): págs. 365, 2003.
cerrAno, C.; BAvestrello, G.; BiAnchi, C. N.; cAttAneo-vietti, R.; BAvA, S.; MorgAnti, C.; Morri, C.; Picco, P.; sArà,
G.; schiAPArelli, S.; siccArdi, A.; sPongA, F. A Catastrophic Mass-mortality Episode of Gorgonians and Other
Organisms in the Ligurian Sea (North-western Mediterranean), Summer 1999. Ecology Letters, (3): págs. 284-
293, 2000.
cicerone, R. et al. The ocean in a high CO2 world». Earth and Ocean Science, 85(37): págs. 351-353, 2004.
clArke, A.; MurPhy, E. J.; Meredith, M. P.; king, J. C.; Peck, L. S.; BArnes, D. K. A.; sMith, R. C. Climate change
and the marine ecosystem of the western Antarctic Peninsula. Philosophical Transactions of the Royal Society of
London [B], 362(1477): págs. 149-166, 2007.
coMA, R.; linAres, C.; riBes, M.; díAz, D.; gArrABou, J.; BAllesteros, E. Consequences of a mass mortality in
populations of Eunicella singularis (Cnidaria: Octocorallia) in Menorca (NW Mediterranean). Marine ecology.
Progress series, (327): págs. 51-60, 2006.
cury, P. M.; shin, Y.-J.; PlAnque, B.; durAnt, J. M.; froMentin, J.-M.; krAMer-schAdt, S.; stenseth, N. C.; trAvers,
M.; griMM, V. Ecosystem oceanography for global change in fisheries. Trends in Ecology and Evolution, 23(6):
págs. 338-346, 2008.
duArte, C. M.; tintoré, J. «El cambio climático y sus consecuencias para los ecosistemas costeros de las islas
Baleares». En: El cambio climático: passat, present y futur. Consejería de Medio Ambiente. Gobierno de las Islas
Baleares, 2001.
272

durAnt, J. M.; hjerMAnn, D. O.; ottersen, G.; stenseth, N. C. Climate and the match or mismatch between
predator requirements and resource availability. Climate Research, 33(3): págs. 271-283, 2007.
fields, P. A.; grAhAM, J. B.; rosenBlAtt, R. H.; soMero, G. N. Effects of expected global climate change on marine
faunas. Trends in Ecology and Evolution, 8(10): págs. 361-367, 1993.
fine, M.; tchernov, D. Scleractinian Coral Species Survive and Recover from Decalcification. Science, (315): págs.
1811, 2007.
frAncour, P.; Boudouresque, c. f.; hArMelin, j. g.; hArMelin-vivien, M. l.; quignArd, j. P. Are the Mediterranean
waters becoming warmer? Information from biological indicators. Mar. Pollut. Bull., 28(9): págs. 523-526,
1994.
freitAs, V.; cAMPos, J.; fonds, M.; vAn der veer, H. W. Potential impact of temperature change on epibenthic predator-
bivalve prey interactions in temperate estuaries. Journal of Thermal Biology, 32(6): págs. 328-340, 2007.
gArrABou, j.; Perez, t.; sArtoretto, s.; hArMelin, j. g. Mass mortality event in red coral (Corallium rubrum,
Cnidaria, Anthozoa, Octocorallia) population in the Provence region (France, NW Mediterranean). Mar. Ecol.
Prog. Ser., (217): págs. 263-272, 2001.
gAzeAu, F.; quiBlier, C.; jAnsen, J. M.; gAttuso, J.-P.; MiddelBurg, J. J.; heiP, C. H. R. Impact of elevated CO2 on
shellfish calcification. Geophysical Research Letters, 34(7): art. L07603, 2007.
giAccone, G.; gerAci, R. M. Biogeografia delle Alghe del Mediterraneo. Anales Jard. Bot. Madrid, 46(I): págs. 27-
33, 1989.
gili, J. M.; PAgès, F. Les proliferacions de meduses. Butlletí de la Societat d’Història Natural de les Balears, (48):
págs. 1-22, 2005.
góMez, F.; clAustre, H. Spreading of Gymnodinium catenatum Graham in the Western Mediterranean Sea.
Harmful Algae News, (22): 2001.
gouretski, V.; kolterMAnn, K. P. How much is the ocean really warming? Geophysical Research Letters, 34(1): art.
L01610, 2007.
greenWood, J. G. Earlier laying by Black Guillemots Cepphus grylle in Northern Ireland in response to increasing
sea-surface temperature. Bird Study, (54): págs. 378-379, 2007.
hAll-sPencer, J. M.; rodolfo-MetAlPA, R.; MArtin, S.; rAnsoMe, E.; fine, M.; turner, S. M. et al. Volcanic carbon dioxide
vents show ecosystem effects of ocean acidification. Nature: (avance publicación online 8 de junio 2008), 2008.
hArley, C. D. et al. (2006).The impacts of climate change in coastal marine systems. Ecology Letters, (9): págs.
228-241, 2006.
hArvell, c. d.; kiM, k.; Burkholder, j. M.; colWell, r. r.; ePstein, P. r.; griMes, d. j.; hofMAnn, e. e.; liPP, e. k.;
osterhAus, A. d. M. e.; overstreet, r. M.; Porter, j. W.; sMith, g. W.; vAstA, g. r. Emerging marine diseases-
Climate links and anthropogenic factors. Science, (285): págs. 1505-1510, 1999.
hAWkes, L. A.; Broderick, A. C.; godfrey, M. H.; godley, B. J. Investigating the potential impacts of climate
change on a marine turtle population. Global Change Biology, 13(5): págs. 923-932, 2007.
hoegh-guldBerg, O.; MuMBy, P. J.; hooten, A. J.; steneck, R. S.; greenfield, P.; góMez, E.; hArvell, C. D.; sAle, P. F.;
edWArds, A. J.; cAldeirA, K.; knoWlton, N.; eAkin, C. M.; iglesiAs-Prieto, R.; MuthigA, N.; BrAdBury, R. H.; duBi,
A.; hAtziolos, M. E. . Coral reefs under rapid climate change and ocean acidification. Science, 318(5857): págs.
1737-1742, 2007.
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
273

hollAnd, M. M.; finnis, J.; BArrett, A. P.; serreze, M. C. Projected changes in Arctic Ocean freshwater budgets.
Journal of Geophysical Research–Biogeosciences, 112(G4): art. G04S55, 2007.
hoWAt, I. M.; joughin, I.; scAMBos, T. A. Rapid changes in ice discharge from Greenland outlet glaciers. Science,
315(5818): págs. 1559-1561, 2007.
høye, T. T.; Post, E.; Meltofte, H.; schMidt, N. M.; forchhAMMer, M. C. Rapid advancement of spring in the High
Arctic. Current Biology, 17(12): R449-R451, 2007.
hughes, L. Biological consequences of global warming: is the signal already apparent?. Trends in Ecology and
Evolution, (15): págs. 56-61, 2000.
hughes, t. P.; BAird, A. h.; BellWood, d. r.; cArd, M.; connolly, s. r.; folke, c.; grosBerg, r.; hoegh-guldBerg,
o.; jAckson, j. B. c.; kleyPAs, j.; lough, j. M.; MArshAll, P.; nyströM, M.; PAluMBi, s. r.; PAndolfi, j. M.; rosen,
B.; roughgArden, j. Climate Change, Human Impacts, and the Resilience of Coral Reefs. Science, (301): págs.
929-933, 2003.
iglesiAs-rodríguez, M. D.; hAllorAn, P. R.; rickABy, R. E. M.; hAll, I. R.; colMenero-hidAlgo, E.; gittins, J. R. et al.
Phytoplankton Calcification in a High-CO2 World. Science, (320): págs. 336-340, 2008.
jAckson, J. B.; sAlA, E. «Unnatural Oceans». En: GILI, J. M.; PRETUS, J. L.; PACKARD, T. T. (ed.). A Marine Science
Odyssey into the 21st Century. Scientia Marina, (65, supl. 2): págs. 273-281, 2001.
jAckson, J. B. C.; kirBy, M. X.; Berger, W. H.; BjorndAl, K. A.; Botsford, L. W.; Bourque, B. J.; BrAdBury, R.
H.; cooke, R.; erlAndson, J.; estes, J. A.; hughes, T. P.; kidWell, S.; lAnge, C. B.; lenihAn, H. S.; PAndolfi, J. M.;
Peterson, C. H.; steneck, R. S.; tegner, M. J.; WArner, R. R. «Historical overfishing and the recent collapse of
coastal ecosystems. Science, (293): págs. 629-638, 2001.
jAgtAP, T. G.; nAgle, V. L. Response and adaptability of mangrove habitats from the Indian subcontinent to
changing climate. Ambio, 36(4): págs. 328-334, 2007.
korringA, C. «The Ocean as Final Recipient of the End Products of the Continent’s Metabolism. Pollution of the
Oceans: Situation, Consequences, and Outlooks for the Future». En: SIOLI, H. (ed.). Ökologie und Lebensschutz
in internationaler Sicht. Ecology and Bioprotection, International Conclusions. Friburgo de Brisgovia: Rombach,
1973, págs. 91-140 (traducción al castellano: Ecología y protección de la naturaleza. Conclusiones internacionales.
Barcelona: Blume, 1982).
lAfferty, K. D.; Porter, J. W.; ford, S. E. Are diseases increasing in the ocean?. Annual Review of Ecology,
Evolution, and Systematics, (35): págs. 31-54, 2004.
lefèvre, N.; WAtson, A. J.; olsen, A.; ríos, A. F.; Pérez, F. F.; johAnnessen, T. A decrease in the sink for atmospheric
CO2 in the North Atlantic. Geophys. Res. Lett., (31): 2004, L07306. DOI: 10.1029/2003GL018957.
levitAn, O.; rosenBerg, G.; setlik, I.; setlikovA, E.; grigel, J.; klePetAr, J.; PrAsil, O.; BerMAn-frAnk, I. Elevated CO2
enhances nitrogen fixation and growth in the marine cyanobacterium Trichodesmium. Global Change Biology,
13(2): págs. 531-538, 2007.
levitus, S.; Antonov, J. L.; Boyer, T. P.; stePhens, C. Warming of the World Ocean. Science, 287: págs. 2225-2229, 2000.
liMA, F. P.; riBeiro, P. A.; queiroz, N.; hAWkins, S. J.; sAntos, A. M. Do distributional shifts of northern and southern
species of algae match the warming pattern?. Global Change Biology, 13(12): págs. 2592-2604, 2007.
linAres, C.; coMA, R.; díAz, D.; zABAlA, M.; hereu, B.; dAntArt, L. Immediate and delayed effects of mass mortality
event on gorgonian population dynamics and benthic community structure in the NW Mediterranean. Marine
Ecology Progress Series, (305): págs. 127-137, 2005.
274

MAckAs, D. L.; BAtten, S.; trudel, M. Effects on zooplankton of a warmer ocean: Recent evidence from the
Northeast Pacific. Progress in Oceanography, 75(2): págs. 223-252, 2007.
MAckenzie, B. R.; schiedek, D. Daily ocean monitoring since the 1860s shows record warming of northern European
seas. Global Change Biology, 13(7): págs. 1335-1347, 2007.
MAckenzie, B. R.; gislAson, H.; MollMAnn, C.; koster, F. W. Impact of 21st century climate change on the Baltic
Sea fish community and fisheries. Global Change Biology, 13(7): págs. 1348-1367, 2007.
MAcleod, C. D.; sAntos, M. B. A.; reid, R. J.; scott, B. E.; Pierce, G. J. Linking sand eel consumption and the
likelihood of starvation in harbour porpoises in the Scottish North Sea: could climate change mean more starving
porpoises?. Biology Letters, 3(2): págs. 185-188, 2007.
MArgAlef, R. (ed.). Western Mediterranean. Oxford: Pergamon Press, 1985 (versió castellana: El Mediterráneo
Occidental. Barcelona: Omega, 1989).
MArgAlef, R. Our biosphere. Excellence in Ecology, 10. Oldendorf (Luhe): Ecology Institute, 1997.
MArtin, Y.; Bonnefont, J. L.; chAncerelle, L. Gorgonians mass mortality during the 1999 late summer in French
Mediterranean coastal waters: the bacterial hypothesis. Water Research, 36(3): págs. 779-782, 2002.
McclAnAhAn, T. R.; AteWeBerhAn, M.; MuhAndo, C. A.; MAinA, J.; MohAMMed, M. S. Effects of climate and
seawater temperature variation on coral bleaching and mortality. Ecological Monographs, 77(4): págs. 503-
525, 2007.
McgrAnAhAn, G.; BAlk, D.; Anderson, B. The rising tide: assessing the risks of climate change and human
settlements in low elevation coastal zones. Environment & Urbanization, 19(1): págs. 17-37, 2007.
MichAelidis, B.; sPring, A.; Portner, H. O. Effects of long-term acclimation to environmental hypercapnia on
extracellular acid-base status and metabolic capacity in Mediterranean fish Sparus aurata. Marine Biology, 150(6):
págs. 1417-1429, 2007.
MieszkoWskA, N.; hAWkins, S. J.; BurroWs, M. T.; kendAll, M. A. Long-term changes in the geographic distribution
and population structures of Osilinus lineatus (Gastropoda: Trochidae) in Britain and Ireland. Journal of the Marine
Biological Association of the United Kingdom, 87(2): págs. 537-545, 2007.
Miles, H.; WiddicoMBe, S.; sPicer, J. I.; hAll-sPencer, J. Effects of anthropogenic seawater acidification on acid-base
balance in the sea urchin Psammechinus miliaris. Marine Pollution Bulletin, 54(1): págs. 89-96, 2007.
Mills, C. E. Jellyfish blooms: are populations increasing globally in response to changing ocean conditions?
Hydrobiologia, 451: págs. 55-68, 2001.
MorA, C.; osPinA, A. F. Tolerance to high temperatures and potential impact of sea warming on reef fishes of
Gorgona Island (tropical eastern Pacific). Marine Biology, 139: págs. 765-769, 2001.
Morán, X. A. G.; seBAstián, M.; Pedrós-Alió, C.; estrAdA, M. Response of Southern Ocean phytoplankton and
bacterioplankton production to short-term experimental warming. Limnology and oceanography, 51(4): págs.
1791-1800, 2006.
nAvArro, L.; Blázquez, M.; Piferrer, F. «Efecto de la temperatura sobre la proporción de sexos de la lubina
(Dicentrarchus labrax)». En: CASTAÑO, J. P.; MALAGÓN, M. M.; GARCÍA NAVARRO, S. (ed.). Avances en
endocrinología comparada (2). Universidad de Còrdova: 2005, págs. 135-138.
NOAA. The State of the Coral Reef Ecosystems of the United States and Pacific Freely Associated States: 2008.
Washington: National Oceanic and Atmospheric Administration, 2008.
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
275

occhiPinti-AMBrogi, A. Anthropogenic Ocean Acidification over the Twenty-First Century and its Impact on
Calcifying Organisms. Nature, (437): págs. 681-686, 2005.
orr, J. C. et al. Anthropogenic Ocean Acidification over the Twenty-First Century and its Impact on Calcifying
Organisms. Nature, (437): págs. 681-686, 2005.
PArrillA, G.; lAvín, A.; Bryden, H.; gArcíA, M.; MillArd, R. Rising temperatures in the Subtropical N. Atlantic over
the past 35 years. Nature, (369): págs. 48-51, 1994.
PAtzner, R. The invasion of Lophocladia (Rhodomelaceae, Lophotaliae) at the northern coast of Ibiza (Western
Mediterranean Sea). Boll. Soc. Hist. Nat. Balears, 41: págs. 75-80, 1998.
Pérès, J.-M.; PicArd, J. Nouveau manuel de bionomie benthique de la mer Méditerranée. Rec. Trav. Sta. Mar.
Endoume, 31(47): págs. 5-137, 1964.
Perez, t.; gArrABou, j.; sArtoretto, s.; hArMelin, j. g.; frAncour, P.; vAcelet, j. Mortalité massive d’invertébrés
marins: un événement sans précédent en Méditerranée nord-occidentale – Mass mortality of marine
invertebrates: an unprecedented event in the NW Mediterranean. C. R. Acad. Sci. Paris, III, (323): págs. 853-
865, 2000.
Por, F. D. Lessepsian migration. Berlín; Nueva York: Springer, 1978.
Portner, H. O.; knust, R. Climate change affects marine fishes through the oxygen limitation of thermal tolerance.
Science, 315(5808): págs. 95-97, 2007.
rAhMstorf, S. . A semi-empirical approach to projecting future sea-level rise. Science, 315(5810): págs. 368-370,
2007.
rieBesell, U.; zondervAn, I.; rost, B.; tortell, P. D.; zeeBe, R. E.; Morel, F. M. M. Reduced calcification of marine
plankton in response to increased atmospheric CO2. Nature, (407): págs. 364-367, 2004.
roMAno, j. c.; BensoussAn, n.; younes, W. A. n.; ArlhAc, d. Anomalies thermiques dans les eaux du golfe de
Marseille durant l’été 1999. Une explication partielle de la mortalité d’invertébrés fixés. C. R. Acad. Sci. Paris, III,
(323): págs. 415-427, 2000.
ros, j. d. . Medusas. Cultura/s-La Vanguardia, (268): págs. 2-3, 2007.
ros, J. D.; roMero, J.; BAllesteros, E.; gili, J. M. Diving in blue water: the benthos. En: MARGALEF, R. (ed.).
Western Mediterranean. Oxford: Pergamon, 1985, págs. 233-295.
royAl society. Ocean Acidification Due to Increasing Atmospheric Carbon Dioxide. Londres: The Royal Society,
2005. Disponible en: http://www.royalsoc.ac.uk/displaypagedoc.asp?id=13314.
sABAtés, A.; MArtín, P.; lloret, J.; rAyA, V. Sea warming and fish distribution: the case of the small pelagic fish,
Sardinella aurita, in the Western Mediterranean. Global Change Biology, (12): págs. 2209-2219, 2006.
sABine, C. L.; feely, R. A.; gruBer, N.; key, R. M.; lee, K.; Bullister, J. L.; WAnninkhof, L.; Wong, C. S.; WAllAce, D.
W. R.; tilBrook, B.; Millero, F. J.; Peng, T. H.; kozyr, A.; ono, T.; ríos, A. F. The oceanic sink for anthropogenic
CO2. Science, (305): págs. 367-371, 2004.
sAlA, E.; knoWlton, N. . Global Marine Biodiversity Trends. Annual Review of Environment and Resources, (31):
págs. 93-122, 2006.
sAlA, E.; Boudouresque, C. F.; hArMelin-vivien, M. Fishing, trophic cascades and the structure of algal assemblages:
evaluation of an old but untested paradigm. Oikos (82): págs. 425-439, 1998.
276

sAlAt, J.; PAscuAl, J. Principales tendencias climatológicas en el Mediterráneo Noroccidental, a partir de más de
30 años de observaciones oceanográficas y meteorológicas en la costa catalana. En: Clima, sociedad y medio
ambiente. Zaragoza: Publicaciones de la Asociación Española de Climatología, 2006, págs. 283-290.
siMMonds, M. P.; isAAc, S. J. The impacts of climate change on marine mammals: early signs of significant problems.
Oryx, 41(1): págs. 19-26, 2007.
sMith, K. L.; roBison, B. H.; helly, J. J.; kAufMAnn, R. S.; ruhl, H. A.; shAW, T. J.; tWining, B. S.; vernet, M. Free-
drifting icebergs: Hot spots of chemical and biological enrichment in the Weddell Sea. Science, 317(5837): págs.
478-482, 2007.
thiBAut, T.; Pinedo, S.; torrAs, X.; BAllesteros, E. Long-term decline of the populations of Fucales (Cystoseira
spp. and Sargassum spp.) in the Albères coast (France, North-western Mediterranean). Marine Pollution Bulletin,
50(12): págs. 1472-1489, 2005.
turley, C. M.; roBerts, J. M.; guinotte, J. M. Corals in deep-water: will the unseen hand of ocean acidification
destroy cold-water ecosystems?. Coral Reefs, 26(3): págs. 445-448, 2007.
vázquez-doMínguez, E.; vAqué, D.; gAsol, A. M. Ocean warming enhances respiration and carbon demand of
coastal microbial plankton. Global Change Biology, 13(7): págs. 1327-1334, 2007.
vilA, M.; MAsó, M. Phytoplankton functional groups and harmful algae species in anthropogenically impacted
waters of the NW Mediterranean Sea. Scientia Marina, 69(1): págs. 31-45, 2005.
WorM, B.; BArBier, E. B.; BeAuMont, N.; duffy, J. E.; folke, C.; hAlPern, B. S.; jAckson, J. B. C.; lotze, H. K.; Micheli,
F.; PAluMBi, S. R.; sAlA, E.; selkoe, K. A.; stAchoWicz, J. J.; WAtson, R. Impacts of Biodiversity Loss on Ocean
Ecosystem Services. Science, (314): págs. 787-790, 1989.
zABAlA, M.; BAllesteros, E. Surface dependent strategies and energy flux in benthic communities or, why corals
do not exist in the Mediterranean. Scientia Marina, 53(1): págs. 3-17, 1989.
zABAlA, M.; gArcíA-ruBies, A.; louisy, P.; sAlA, E. Spawning behaviour of the Mediterranean dusky grouper
Epinephelus marginatus (Lowe, 1834) (Pisces, Serranidae) in the Medes Islands Marine Reserve (NW Mediterranean,
Spain). Scientia Marina, 61(1): págs. 65-77, 1997.
Impactos en los ecosistemas acuáticos
El mar y las costas de Cataluña ya notan el efecto del cambio climático
277


















