
ANÁLISIS BIOECONÓMICO DE LA PESQUERÍA DEL CAMARÓN …Figura 7 – Captura de camarón para el...
266
INSTITUTO POLITECNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS ANÁLISIS BIOECONÓMICO DE LA PESQUERÍA DEL CAMARÓN EN EL GOLFO DE CALIFORNIA TESIS QUE PARA OBTENER EL GRADO DE DOCTORADO EN CIENCIAS EN BIOECONOMÍA PESQUERA Y ACUÍCOLA PRESENTA FERNANDO ARANCETA GARZA LA PAZ, B.C.S., OCTUBRE DE 2016
Transcript of ANÁLISIS BIOECONÓMICO DE LA PESQUERÍA DEL CAMARÓN …Figura 7 – Captura de camarón para el...

INSTITUTO POLITECNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
ANÁLISIS BIOECONÓMICO DE LA PESQUERÍA
DEL CAMARÓN EN EL GOLFO DE CALIFORNIA
TESIS
QUE PARA OBTENER EL GRADO DE
DOCTORADO EN CIENCIAS EN BIOECONOMÍA PESQUERA Y
ACUÍCOLA
PRESENTA
FERNANDO ARANCETA GARZA
LA PAZ, B.C.S., OCTUBRE DE 2016



Agradecimientos
Se agradece a CONACyT por la beca, a los apoyos recibidos por el IPN como
son la beca BEIFI y COFAA.
Las siguientes personas que quiero agradecer la paciencia y dedicación son mis
co-directores el Dr. Francisco Arreguín-Sánchez y el Dr. Juan Carlos Seijo. Que en
verdad para mí ha sido un honor y la experiencia más interesante y grata el trabajar y
tener como directores dos personas de reconocimientos mundiales y muy sencillos de
trato, siempre amables y con una gran disposición.
Le agradezco a mi comité revisor el tiempo, comentarios y sugerencias para la
tesis y mi trabajo doctoral en general.
Agradezco a CONAPESCA y de manera personal Raul Villaseñor Talavera por
el apoyo brindado en la tesis.
También agradezco mucho a todas las personas que me acompañaron desde
el inicio hasta el final de esta experiencia de vida, enriqueciéndola y ayudando a no
volverse más loco, con énfasis en la personita más bella y linda que me pude encontrar
y me apoyo mucho al final de la tesis, Carolina.
Dedicatoria
Esta tesis va dirigida a mi familia y otra persona muy especial. Esta persona por
desgracia ya no está con nosotros que fue quien me impulsó y motivo a tomar el reto
“de la pesquería más importante y problemática de todo México, para diseñar el mejor
modelo posible con los mejores datos posibles, ¿cómo ve? ¿Acepta el reto?” Dr. Daniel
Lluch Belda
Mi familia que es lo más importante y valioso que alguien puede tener y que
siempre me apoyaron para seguir adelante no importando las presiones internas y
externas en el transcurso de este viaje de cuatro años, los amo mamá, papá, ale y ro.

I
Índice
Índice de figuras .......................................................................................................... V
Índice de tablas ......................................................................................................... XII
Resumen ................................................................................................................. XVII
Abstract .................................................................................................................. XVIII
2. Antecedentes .......................................................................................................... 5
2.1 Biológicos ........................................................................................................... 5
2.1.1. Biología general .......................................................................................... 5
2.1.2. Parámetros de edad y crecimiento ........................................................... 14
2.1.3. Mortalidad natural ..................................................................................... 16
2.1.4. Variabilidad ambiental y reclutamiento ..................................................... 21
2.2 Pesqueros: La pesquería en el sur del Golfo de California .............................. 27
2.3. Pesqueros: variables pesqueras ..................................................................... 38
2.3.1. Esfuerzo pesquero en Sinaloa sur ............................................................ 38
2.3.2. Dinámica del esfuerzo de alta mar ............................................................ 40
2.3.3. Selectividad .............................................................................................. 40
2.3.4. Capturabilidad ........................................................................................... 42
2.3.5. Mortalidad por pesca, F ............................................................................ 50
2.3. Normatividad ................................................................................................... 53
2.4. Ordenación Pesquera ..................................................................................... 57
2.4.1. Ordenación y manejo pesquero a través de modelos pesqueros en camarón
............................................................................................................................ 57
2.4.2. Estrategias de manejo pesquero globales ................................................ 59
2.4.3. Estrategias de manejo aplicadas en la pesquería de camarón ................. 62

II
2.4.4. Manejo pesquero del camarón en México ................................................ 64
2.5. Modelos bioeconómicos empleados en las pesquerías de camarón .............. 69
3. Justificación ....................................................................................................... 75
4. Objetivos ............................................................................................................... 77
4.1. Objetivo general .............................................................................................. 77
4.2. Objetivo particular ........................................................................................... 77
5. Materiales y métodos ............................................................................................ 78
5.1. Parámetros de la dinámica poblacional .......................................................... 78
5.1.1. Crecimiento para las especies de camarón Litopenaeus stylirostris, L.
vannamei y Farfantepenaeus californiensis. ....................................................... 78
5.1.2. Clave edad-longitud para las especies de camarón Litopenaeus stylirostris,
L. vannamei ........................................................................................................ 79
5.1.3. Uso de intervalos gnomónicos para estimar la mortalidad natural a la edad
(Mi) ...................................................................................................................... 81
5.1.4. Variabilidad en la estimación de Mi ........................................................... 84
5.2. Parámetros del modelo pesquero ................................................................... 85
5.2.1. Estimación del esfuerzo pesquero por tipo flota ....................................... 85
5.2.2. Bases de datos de captura por flotas ....................................................... 86
5.2.3. Obtención de la estructura de edades de la captura................................. 87
5.2.4. Modelo de capturabilidad a la edad (qi) .................................................... 88
5.3. Modelo bioeconómico para la pesquería multi-específica y multi-flota de
camarón ................................................................................................................. 92
5.3.1. Sub-modelo biológico ............................................................................... 92
5.3.2. Sub-modelo Tecnológico .......................................................................... 95
5.3.3 Sub-modelo económico ............................................................................. 96

III
5.3.4. Sub-modelo ambiental - Escenarios de reclutamiento en función de su
variabilidad interanual ....................................................................................... 103
5.3.5 Escenarios de manejo pesquero ............................................................. 105
6. Resultados .......................................................................................................... 110
6.1. Parámetros de la dinámica poblacional ........................................................ 110
6.1.1. Selección de ecuaciones de crecimiento para las especies de especies de
camarón Litopenaeus stylirostris, L. vannamei y Farfantepenaeus californiensis.
.......................................................................................................................... 110
6.1.2. Claves edad-longitud para el camarón blanco y azul en la zona de Sinaloa
Sur .................................................................................................................... 115
6.1.3. Vectores de mortalidad natural en camarón del Pacífico mexicano
generados por modelo gnomónico (Caddy, 1996) ............................................ 118
6.1.3. Variabilidad en M .................................................................................... 122
6.2. Parámetros del modelo pesquero ................................................................. 123
6.2.1. Estimación del esfuerzo pesquero por tipo flota ..................................... 123
6.2.2. Bases de datos de captura por flotas ..................................................... 127
6.3. Obtención de la estructura de longitudes de la captura ................................ 131
6.3.1. Estructura de longitudes de la captura para la flota de pequeña escala . 131
6.3.2. Estructura de longitudes de la captura para la flota industrial ................. 134
6.4. Modelo de capturabilidad a la edad (𝑞𝐿, 𝑡) por especie ................................. 146
6.4.1. Capturabilidad a la longitud de la flota de pequeña escala ..................... 146
6.4.2. Capturabilidad a la longitud de la flota industrial ..................................... 152
6.5. Modelo bioeconómico para la pesquería multi-específica y multi-flota de
camarón ............................................................................................................... 174
6.5.1. Salidas del modelo bioeconómico de estructura por edades para la
pesquería del camarón en Sinaloa sur temporada 2014-15 ............................. 174

IV
6.5.2. Escenarios de manejo para temporada 2014-2015 de la pesquería de
camarón en Sinaloa sur .................................................................................... 179
6.5.3. Escenarios de variabilidad interanual del reclutamiento para la temporada
2014-2015 de la pesquería de camarón en Sinaloa sur. .................................. 184
6.5.4. Escenarios de manejo para temporada 2014-2015 de la pesquería de
camarón en Sinaloa sur incluyendo la variabilidad interanual del reclutamiento.
.......................................................................................................................... 190
7. Discusión ............................................................................................................. 196
7.1. Parámetros de la dinámica poblacional ........................................................ 196
7.2. Parámetros pesqueros .................................................................................. 200
7.2.1. Sección esfuerzo y captura de la pesquería de camarón en Sinaloa sur.
.......................................................................................................................... 200
7.2.2. La capturabilidad a la longitud (𝑞(𝐿, 𝑡)) en la pesquería de camarón 2014-15
en Sinaloa sur ................................................................................................... 203
7.3. Modelo bioeconómico para la pesquería multi-específica y multi-flota de
camarón ............................................................................................................... 207
7.3.1. Manejo actual de la pesquería de camarón por parte de las autoridades
pesqueras ......................................................................................................... 207
7.3.3. Manejo de la pesquería de camarón bajo el punto de referencia de máximo
rendimiento económico (siglas en inglés MEY) con variabilidad en el reclutamiento
pesquero. .......................................................................................................... 211
7.4. Discusión global ........................................................................................... 213
8. Conclusiones ....................................................................................................... 216
9. Recomendaciones ............................................................................................... 218
10. Bibliografía ........................................................................................................ 220

V
Índice de figuras
Figura 1 – Periodos de madurez sexual en hembras de camarón para el Golfo de
California (estudio centrado en Mazatlán) Las barras grises muestran los picos de
madurez (Lluch, 1977). ............................................................................................... 9
Figura 2– Frecuencia promedio de hembras maduras o en fase III para las costas de
Sinaloa de acuerdo al INP (2013). ............................................................................ 10
Figura 3– Volúmenes de producción de camarón por origen: cultivo, esteros y bahías
y altamar a nivel nacional (a), litoral del pacífico (b) y Golfo y Caribe Mexicano (c). . 28
Figura 4– Volumen de producción pesquera de camarón 2013 para cada estado del
Golfo de California. (B.C. y B.C.S.: Baja California y Baja California Sur)
(CONAPESCA, 2013). .............................................................................................. 29
Figura 5– Proporción de especies dominantes en las capturas par la flota de alta mar
(a) y de aguas interiores (b) dentro del Golfo de California (INP, 2013; INP, 2014a,b).
.................................................................................................................................. 32
Figura 6 - Proporción de volúmenes (ton) de captura de camarón silvestre en el estado
de Sinaloa. CONAPESCA (2007). ............................................................................. 33
Figura 7 – Captura de camarón para el estado de Sinaloa reportado por el Anuario
Estadístico de Acuacultura y Pesca para los años 2014 y 2015. a) Producción de
aguas interiores y b) producción de mar abierto. ...................................................... 36
Figura 8– Porcentaje de lances de cada flota por zonas de pesca asignadas por el INP.
Donde los puertos son: Guaymas (GY); Mazatlán (MZ); Pto Chiapas (PC); Pto Madero
(PM); Pto Peñasco (PP); Salina Cruz (SZ); San Blas (SB); San Carlos (SC); San Felipe
(SF); Topolobampo (TB); y Yavaros (YV). Las zonas son: Las zonas estudiadas por
cada CRIP son: Zona 10 (Ensenada); Zona 50 (La Paz); Zona 20 (Guaymas); Zonas
30 (Sinaloa centro y norte),40 (Sinaloa sur) y 60 (Bahía de Banderas, Nay.); y Zona 90
(Salina Cruz). ............................................................................................................ 40
Figura 9– Selectividad a la talla en la pesquería de camarón del Pacífico (Mazatlán)
usando el arte de pesca autorizado por cada flota, siendo la atarraya en aguas
interiores y la red de arrastre para altamar. .............................................................. 42
Figura 10– Tácticas de manejo pesquero empleadas en el manejo de las pesquerías
en el mundo. .............................................................................................................. 61

VI
Figura 11 - Estrategias de manejo en la pesquería de camarón. .............................. 63
Figura 12– Instrumentos de manejo para la pesquería de camarón (CONAPESCA,
2004). ........................................................................................................................ 65
Figura 13- Objetivos de manejo para la pesquería de camarón en el Pacífico mexicano
(CONAPESCA, 2004). .............................................................................................. 66
Figura 14 - Puntos de referencia en manejo de la pesquería de camarón en el Pacífico
mexicano. .................................................................................................................. 67
Figura 15– Sitios de muestreo único en Sinaloa sur. 1: Huizache, 2: Chametla, 3:
Teacapán, 4: Palmilla y 5: zona de costera de Teacapán. ........................................ 79
Figura 16 – Porcentaje de hembras maduras en fase III para el camarón blanco (a),
azul (b) y café (c). .................................................................................................... 104
Figura 17– Ecuaciones de crecimiento de von Bertalanffy aplicadas al camarón blanco
en el Pacífico Mexicano. ......................................................................................... 111
Figura 18– Ecuaciones de crecimiento de Von Bertalanffy aplicadas al camarón azul
en el Pacífico Mexicano. La línea negra gruesa es la estimación seleccionada. .... 112
Figura 19– Ecuaciones de crecimiento de von Bertalanffy aplicadas al camarón café
en el Pacífico Mexicano. ......................................................................................... 113
Figura 20 – Comparación entre las ecuaciones de crecimiento de von Bertalanffy
seleccionadas para el presente trabajo en Sinaloa sur. .......................................... 114
Figura 21– Resultados del análisis de Battacharya (1967) para el camarón blanco
donde se grafica la desviación estándar observada para cada talla modal de Chávez
(1973) y Gutiérrez (1980). ....................................................................................... 115
Figura 22- Clave edad-longitud en términos de probabilidad para el camarón blanco
Litopenaeus vannamei. ........................................................................................... 116
Figura 23– Resultados del análisis de Battacharya (1967) para el camarón azul donde
se obtuvo la desviación estándar observada para cada talla modal de las frecuencias
de Castro-Ortiz y Sánchez-Rojas (1976) e informes técnicos de la pesquería de
altamar (INP, 1974; 1975). ...................................................................................... 117
Figura 24 – Clave edad-longitud en términos de probabilidad para el camarón azul
Litopenaeus stylirostris. ........................................................................................... 118

VII
Figura 25– Vectores de mortalidad natural estimados para el camarón café (a), azul
(b) y blanco (c) en círculos. La estimaciones independientes de la literatura están con
taches. ..................................................................................................................... 121
Figura 26– Frecuencia relativa de las capturas por base de datos [a: base industrial
para el puerto de Mazatlán, Sinaloa y para Sinaloa sur (satélite); b: flota de pequeña
escala). .................................................................................................................... 129
Figura 27- Captura industrial por especie para el puerto de Mazatlán (SAGARPA 2014-
15). .......................................................................................................................... 130
Figura 28 – Captura de la flota de pequeña escala para la temporada 2014-15 en
Sinaloa sur (SAGARPA 2014-15). ........................................................................... 130
Figura 29 – Frecuencias relativas de la captura en camarón blanco (Litopenaeus
vannamei) de pequeña escala por mes en la temporada 2014-15 en Sinaloa sur. . 132
Figura 30– Individuos totales en la captura de pequeña escala en camarón blanco
(Litopenaeus vannamei) por mes en la temporada 2014-15 en Sinaloa sur. .......... 133
Figura 31 – Captura por unidad de esfuerzo (lance/día) en individuos para la flota de
pequeña escala en camarón blanco (Litopenaeus vannamei) por mes en la temporada
2014-15 en Sinaloa sur. .......................................................................................... 133
Figura 32– Frecuencias relativas por longitud total de maquila obtenidas de la
maquiladora Mexican Shrimp Paradise para camarón blanco (Litopenaeus vannamei)
para la temporada 2014-15. .................................................................................... 135
Figura 33– Frecuencias relativas por longitud total de maquila obtenidas de la
maquiladora Mexican Shrimp Paradise para camarón azul (Litopenaeus stylirostris)
para la temporada 2014-15. .................................................................................... 136
Figura 34– Frecuencias relativas por longitud total de maquila obtenidas de la
maquiladora Mexican Shrimp Paradise para camarón café (Farfantepenaeus
californiensis) para la temporada 2014-15. ............................................................. 137
Figura 35– Captura de la flota industrial en individuos por clase de longitud de camarón
blanco (Litopenaeus vannamei) en Sinaloa sur para la temporada 2014-15. ......... 139
Figura 36 - Captura de la flota industrial en individuos por clase de longitud de camarón
azul (Litopenaeus stylirostrisi) en Sinaloa sur para la temporada 2014-15. ............ 140

VIII
Figura 37- Captura de la flota industrial en individuos por clase de longitud de camarón
café (Farfantepenaeus californiensis) para la flota que pesco en Sinaloa sur la
temporada 2014-15. ................................................................................................ 141
Figura 38– Captura por unidad de esfuerzo (día/barco) en individuos para la especie
de camarón blanco (Litopenaeus vannamei) en Sinaloa sur para la temporada 2014-
15. ........................................................................................................................... 143
Figura 39– Captura por unidad de esfuerzo (día/barco) en individuos para la especie
de camarón azul (Litopenaeus stylirostris) en Sinaloa sur para la temporada 2014-15.
................................................................................................................................ 144
Figura 40– Captura por unidad de esfuerzo (día/barco) en individuos para la especie
de camarón café (Farfantepenaeus californiensis) en Sinaloa sur para la temporada
2014-15. .................................................................................................................. 145
Figura 41– Función logística general de capturabilidad a la edad 𝑞(𝐿, 𝑡) en camarón
blanco para la flota de pequeña escala en Sinaloa sur temporada 2014-15. .......... 147
Figura 42 - Patrón mensual de capturabilidad por longitud de camarón blanco para la
flota de pequeña escala en Sinaloa sur temporada 2014-15 (a-d). El resumen de los
patrones de 𝑞(𝐿, 𝑡) mensual se observa en (e). Los círculos vacíos representan valores
extremos eliminados para fines del ajuste. ............................................................. 151
Figura 43– Función logística general de capturabilidad a la edad 𝑞(𝐿, 𝑡) en camarón
blanco para la flota industrial en Sinaloa sur temporada 2014-15. .......................... 153
Figura 44– Patrón mensual de capturabilidad por longitud de camarón blanco para la
flota industrial en Sinaloa sur temporada 2014-15 (a-d). El resumen de los patrones de
q(L, t) mensual se observa en (f). Los círculos vacíos representan valores extremos
eliminados para fines del ajuste. ............................................................................. 158
Figura 45– Vulnerabilidad del recurso en función de la capturabilidad por cada longitud
para la pesquería de camarón blanco en Sinaloa sur temporada 2014-15. ............ 159
Figura 46– Función logística general de capturabilidad a la edad 𝑞(𝐿, 𝑡) en camarón
azul para la flota industrial en Sinaloa sur temporada 2014-15. .............................. 160
Figura 47- Patrón mensual de capturabilidad por longitud de camarón azul para la flota
industrial en Sinaloa sur temporada 2014-15 (a-d). El resumen de los patrones de

IX
𝑞(𝐿, 𝑡) mensual se observa en (f). Los círculos vacíos representan valores extremos
eliminados para fines del ajuste. ............................................................................. 164
Figura 48– Función logística general de capturabilidad a la edad 𝑞(𝐿, 𝑡) en camarón
café para la flota industrial en Sinaloa sur temporada 2014-15............................... 166
Figura 49- Patrón mensual de capturabilidad por longitud de camarón café para la flota
industrial en Sinaloa sur temporada 2014-15 (a-d). El resumen de los patrones de
𝑞(𝐿, 𝑡) mensual se observa en (f). Los círculos vacíos representan valores extremos
eliminados para fines del ajuste. ............................................................................. 171
Figura 50 – Relación entre la capturabilidad a la talla (q(L,t) mensual promedio con la
captura por unidad de esfuerzo (CPUE) mensual para la flota de pequeña escala para
camarón blanco (a), y en la flota industrial para el camarón blanco (b), azul (c) y café
(d). ........................................................................................................................... 173
Figura 51 – Captura mensual observada y estimada por la flota de pequeña escala (a)
y la industrial (b) para Litopenaeus vannamei (Lv) en Sinaloa sur. ......................... 175
Figura 52 – Captura mensual observada y estimada por cada flota para Litopenaeus
stylirostris (Ls) en Sinaloa sur. ................................................................................ 176
Figura 53 – Captura mensual observada y estimada por cada flota para
Farfantepenaeus californiensis (Fc) en Sinaloa sur. ............................................... 176
Figura 54 – Reclutamiento estacional mensual estimado para las especies de
Litopenaeus vannamei, L. stylirostris y Farfantepenaeus californiensis (Fc) en Sinaloa
sur. .......................................................................................................................... 177
Figura 55 – Composición de la captura de la flota industrial de camarón en Sinaloa sur.
................................................................................................................................ 178
Figura 56 – Valor presente neto de la pesquería de camarón por cada escenario de
manejo en Sinaloa sur, temporada 2014-15............................................................ 180
Figura 57 – Utilidades por barco industrial, esfuerzo pesquero en días y barcos para la
flota industrial por escenario de manejo de camarón en Sinaloa sur, temporada 2014-
15. ........................................................................................................................... 180
Figura 58 – Utilidades por cayuco y esfuerzo total para la flota de pequeña escala por
escenario de manejo de camarón en Sinaloa sur, temporada 2014-15. ................. 181

X
Figura 59 – Reclutas anuales por escenario de manejo junto a las distintas
combinaciones de esfuerzo estimado. .................................................................... 181
Figura 60 – Biomasa de reproductores anuales por escenario de manejo junto a las
distintas combinaciones de esfuerzo estimado. ...................................................... 182
Figura 61 – Valor presente neto de la pesquería de camarón en Sinaloa sur,
temporada 2014-15 por cada escenario climático-reclutamiento. Status quo: no varió
el esfuerzo en la pesquería, CD: calentamiento débil, CM: calentamiento moderado,
ED: enfriamiento débil. ............................................................................................ 185
Figura 62 – Utilidades por barco industrial, esfuerzo pesquero en días y barcos para la
flota industrial del camarón en Sinaloa sur, temporada 2014-15 por cada escenario
climático-reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD:
calentamiento débil, CM: calentamiento moderado, ED: enfriamiento débil. .......... 185
Figura 63 – Utilidades por cayuco y esfuerzo total para la flota de pequeña escala del
camarón en Sinaloa sur, temporada 2014-15 por cada escenario climático-
reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD: calentamiento
débil, CM: calentamiento moderado, ED: enfriamiento débil. .................................. 186
Figura 64 – Reclutas anuales sin modificar el esfuerzo anual por cada escenario
climático-reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD:
calentamiento débil, CM: calentamiento moderado, ED: enfriamiento débil. .......... 186
Figura 65 – Reproductores anuales sin modificar el esfuerzo anual por cada escenario
climático-reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD:
calentamiento débil, CM: calentamiento moderado, ED: enfriamiento débil. .......... 187
Figura 66 – Valor presente neto bajo condiciones de máximo rendimiento económico
(MEY) de la pesquería de camarón en Sinaloa sur, temporada 2014-15 por cada
escenario climático-reclutamiento. Status quo: no varió el esfuerzo en la pesquería,
CD: calentamiento débil, CM: calentamiento moderado, ED: enfriamiento débil. ... 191
Figura 67 – Utilidades por barco industrial, esfuerzo pesquero en días y barcos para la
flota industrial del camarón bajo condiciones de máximo rendimiento económico (MEY)
en Sinaloa sur, temporada 2014-15 por cada escenario climático-reclutamiento. Status
quo: no varió el esfuerzo en la pesquería, CD: calentamiento débil, CM: calentamiento
moderado, ED: enfriamiento débil. .......................................................................... 191

XI
Figura 68 – Utilidades por cayuco y esfuerzo total para la flota de pequeña escala del
camarón bajo condiciones de máximo rendimiento económico (MEY) en Sinaloa sur,
temporada 2014-15 por cada escenario climático-reclutamiento. Status quo: no varió
el esfuerzo en la pesquería, CD: calentamiento débil, CM: calentamiento moderado,
ED: enfriamiento débil. ............................................................................................ 192
Figura 69 – Reclutas anuales bajo condiciones de máximo rendimiento económico
(MEY) por cada escenario climático-reclutamiento. Status quo: no varió el esfuerzo en
la pesquería, CD: calentamiento débil, CM: calentamiento moderado, ED: enfriamiento
débil. ........................................................................................................................ 193
Figura 70 – Biomasa de reproductores anuales bajo condiciones de máximo
rendimiento económico (MEY) por cada escenario climático-reclutamiento. Status quo:
no varió el esfuerzo en la pesquería, CD: calentamiento débil, CM: calentamiento
moderado, ED: enfriamiento débil. .......................................................................... 193

XII
Índice de tablas
Tabla 1 - Distribución y hábitat de las principales especies comerciales de camarón en
el Pacífico Mexicano (Brusca y Hendrickx, 2008). ...................................................... 5
Tabla 2– Nicho ecológico por estadio de vida de los camarones peneidos comerciales
en el Pacífico Mexicano. ............................................................................................. 6
Tabla 3– Las tallas de primera madurez por especie para los camarones del Pacífico
Mexicano. .................................................................................................................... 8
Tabla 4– Las tallas de primera madurez por especie para los camarones del Pacífico
Mexicano. .................................................................................................................... 8
Tabla 5- Valores de la fecundidad mínima, máxima y promedio para de las especies
de camarón café (Farfantepenaeus californiensis), azul (Litopenaeus stylirostris) y
blanco (L. vannamei) basado en la literatura reportada. ........................................... 10
Tabla 6- Duración del estadio de huevo reportado para de las especies de camarón
café (Farfantepenaeus californiensis), azul (Litopenaeus stylirostris) y blanco (L.
vannamei).................................................................................................................. 11
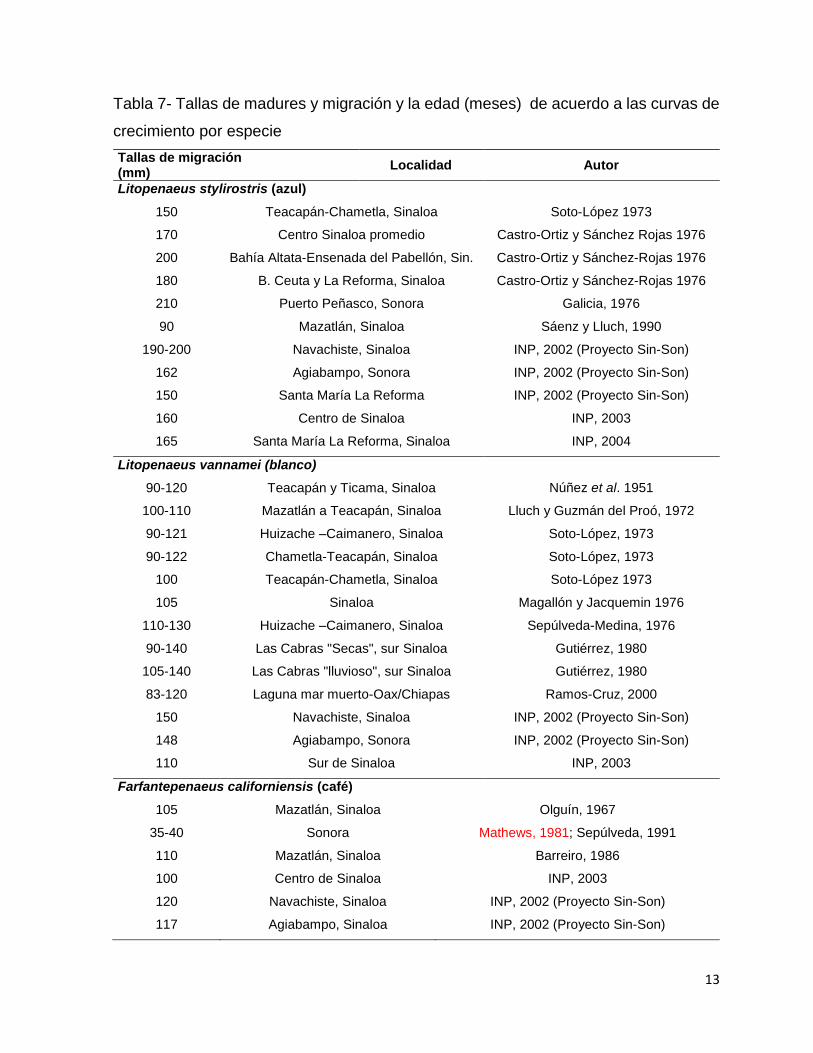
Tabla 7- Tallas de madures y migración y la edad (meses) de acuerdo a las curvas de
crecimiento por especie ............................................................................................ 13
Tabla 8 – Parámetros de edad y crecimiento estimados para cada especie de camarón.
.................................................................................................................................. 14
Tabla 9– Funciones clave edad-longitud en camarones del Pacífico Mexicano........ 16
Tabla 10– Referencias de valores estimados de mortalidad natural en las especies de
camarón blanco (Litopenaeus vannamei), azul (L. stylirostris) y café (Farfantepenaeus
californiensis) en el Pacífico mexicano. ..................................................................... 19
Tabla 11– Principales variables ambientales y su efecto sobre la abundancia y
reclutamiento de los peneidos dentro del Golfo de California. .................................. 23
Tabla 12– efecto de los periodos de calentamiento (Niño) y enfriamiento (Niña) sobre
el reclutamiento del camarón en el Pacífico Mexicano. ............................................. 26
Tabla 13 – Volumen de producción peso vivo de camarón en toneladas para el litoral
del Pacífico Mexicano de 1990-2013 incluyendo la referencia nacional (CONAPESCA,
2013). ........................................................................................................................ 30

XIII
Tabla 14- Producción pesquera del estado de Sinaloa (tons) en bahías, esteros y
altamar (2000 – 2007). .............................................................................................. 35
Tabla 15 - Peso vivo reportado para las capturas silvestres por el Anuario Estadístico
de Acuacultura y Pesca en Sinaloa para los años 2014 y 2015 para el recurso camarón.
(CONAPESCA 2014, 2015)....................................................................................... 37
Tabla 16- Embarcaciones de altamar registradas por estado para el litoral del Pacífico
mexicano (Anuarios de Pesca). ................................................................................. 39
Tabla 17 – Estimaciones independientes de capturabilidad en la literatura haciendo
énfasis en la pesquería de camarón en México. ....................................................... 47
Tabla 18 – Supuesto de capturabilidad (q) más comunes empleados en los modelos
poblacionales y de producción con sus implicaciones (Arreguín-Sánchez, 1996). ... 50
Tabla 19 – Valores de mortalidad por pesca (F) para la pesquería de camarón en el
Pacífico Mexicano. .................................................................................................... 51
Tabla 20 - Normatividad en la pesquería de camarón del Pacífico Mexicano en la NOM-
002-SAG/PESC-20131, Carta Nacional Pesquera (2012)2, y Plan de Manejo para la
Pesquería de Camarón en el Litoral del Océano Pacífico Mexicano del 20043. ....... 54
Tabla 21– Objetivos de ordenación en las pesquerías. ............................................. 57
Tabla 22– Criterios de bienestar aplicados a pesquerías marinas (Seijo et al., 1997;
Bergstrom y Randall, 2016). ...................................................................................... 60
Tabla 23 - Modelos bioeconómicos aplicados en la pesquería de camarón en el Golfo
de California. ............................................................................................................. 71
Tabla 24– Estudios empleados en la elaboración de la clave edad por especie....... 81
Tabla 25– División gnomónica del ciclo de vida de las especies de camarón café
(Farfantepenaeus californiensis), azul (Litopenaeus stylirostris) y blanco (L. vannamei)
basado en la literatura reportada. .............................................................................. 84
Tabla 26– Parámetros de crecimiento y peso empleadas para las especies de
camarones café (Farfantepenaeus californiensis), azul (Litopenaeus stylirostris) y
blanco (L. vannamei). ................................................................................................ 88
Tabla 27– Costos del primer viaje y del resto de viajes de la temporada 2014-15 por
parte de la flota Pesca Mar de Manzanillo, S.A. de C.V. en Mazatlán, Sinaloa. ....... 97

XIV
Tabla 28– Costos variables de la flota de pequeña escala para la temporada 2014-15
en Sinaloa sur. .......................................................................................................... 98
Tabla 29- Costo de pesca industrial y de pequeña escala por día (US$). ................ 98
Tabla 30- Costo de oportunidad de la flota industrial y de pequeña escala. ............ 99
Tabla 31- Precios por tonelada de la captura por especie y por flota para la temporada
2014-15. .................................................................................................................... 99
Tabla 32 – Captura incidental por barco y flota, especie y mes en toneladas al día y en
ingresos (US$). ....................................................................................................... 101
Tabla 33- Parámetros bioeconómicos de las flotas de pequeña escala e industrial de
la pesquería del sur del Golfo de California............................................................. 102
Tabla 34 – Manejo pesquero propuesto para la pesquería de camarón del sur de
Sinaloa en la temporada 2014-15. .......................................................................... 107
Tabla 35 – Escenarios de manejo para la pesquería de camarón en Sinaloa sur 2014-
15. ........................................................................................................................... 108
Tabla 36- Escenarios de reclutamiento basados en el efecto de las variables
ambientales sobre la reproducción de las especies de camarón del Pacífico. ........ 109
Tabla 37– Parámetros de la ecuación de crecimiento estimados en este estudio para
el camarón blanco de Sinaloa sur. .......................................................................... 110
Tabla 38– Vectores de mortalidad natural (𝑀𝑖) estimados con la duración promedio del
primer estadio de vida para las especies de camarón café (Farfantepenaeus
californiensis), azul (Litopenaeus stylirostris) y blanco (L. vannamei) con el modelo
gnomónico (Caddy, 1996). 𝐺 y ∝ son constantes generadas por el modelo. .......... 120
Tabla 39– Variaciones debido a diferentes duraciones en la etapa huevo para el
camarón blanco (L. vannamei), azul (L. stylirostris) y café (F. californiensis) usando el
modelo gnomónico; n, número de diferentes duraciones de la etapa huevo usadas en
la estimación; DE, desviación estándar; CV, coeficiente de la variación. ................ 122
Tabla 40– Estimación del esfuerzo pesquero para la flota industrial para la temporada
2014-15 en Sinaloa sur. .......................................................................................... 123
Tabla 41– Estimación del esfuerzo pesquero para la flota de pequeña escala para la
temporada 2014-15 en Sinaloa sur. ........................................................................ 124

XV
Tabla 42– Número de pescadores para la temporada 2014-15 reportados en Sinaloa
Sur. .......................................................................................................................... 125
Tabla 43 - Capturas reportadas por base de datos generada en toneladas de peso
vivo: a) Puerto de Mazatlán 2014-15; b) Captura estimada vía satelital 2014-15; y c)
Captura de la flota de pequeña escala 2014-15. ..................................................... 128
Tabla 44- Captura por unidad de esfuerzo de pequeña escala (CPUE) estimada en la
temporada 2014-15 en Sinaloa sur (solo Litopenaeus vannamei). ......................... 132
Tabla 45– Captura por unidad de esfuerzo industrial (CPUE) por día/barco/especie en
la temporada 2014-15 en Sinaloa sur. .................................................................... 146
Tabla 46– Valores mensuales de la capturabilidad a la longitud del modelo de
CATCHABILITY en camarón blanco estimados para la flota de pequeña escala por
lance en Sinaloa sur para la temporada 2014-15. ................................................... 148
Tabla 47– Valores mensuales estimados de la capturabilidad a la longitud/edad de
camarón blanco para la flota de pequeña escala por lance en Sinaloa sur para la
temporada 2014-15. ................................................................................................ 149
Tabla 48– Parámetros de la función logística de la capturabilidad a la longitud de
camarón blanco para la flota de pequeña escala en Sinaloa sur para la temporada
2014-15. .................................................................................................................. 149
Tabla 49 - Valores mensuales de la capturabilidad a la longitud del modelo de
CATCHABILITY en camarón blanco estimados para la flota industrial por día efectivo
de pesca/barco en Sinaloa sur para la temporada 2014-15. ................................... 154
Tabla 50- Valores mensuales estimados de la capturabilidad a la longitud/edad de
camarón blanco para la flota industrial por día efectivo/barco en Sinaloa sur para la
temporada 2014-15. ................................................................................................ 155
Tabla 51 - Parámetros de la función logística de la capturabilidad a la longitud de
camarón blanco para la flota industrial en Sinaloa sur para la temporada 2014-15. 155
Tabla 52- Valores mensuales de la capturabilidad a la longitud del modelo de
CATCHABILITY en camarón azul estimados para la flota industrial por día efectivo de
pesca/barco en Sinaloa sur para la temporada 2014-15. ........................................ 161

XVI
Tabla 53- Valores mensuales estimados de la capturabilidad a la longitud/edad de
camarón azul para la flota industrial por día efectivo/barco en Sinaloa sur para la
temporada 2014-15. ................................................................................................ 162
Tabla 54 - Parámetros de la función logística de la capturabilidad a la longitud de
camarón azul para la flota industrial en Sinaloa sur para la temporada 2014-15. ... 162
Tabla 55 - Valores mensuales de la capturabilidad a la longitud del modelo de
CATCHABILITY en camarón café estimados para la flota industrial por día efectivo de
pesca/barco en Sinaloa sur para la temporada 2014-15. ........................................ 167
Tabla 56 - Valores de la capturabilidad a la longitud de camarón café obtenidos de
individuo/días/flota para la flota industrial en Sinaloa sur para la temporada 2014-15.
................................................................................................................................ 168
Tabla 57 - Parámetros de la función logística de la capturabilidad a la longitud de
camarón café para la flota industrial en Sinaloa sur para la temporada 2014-15. ... 169
Tabla 58 – Escenarios de manejo pesquero para el camarón en Sinaloa sur para la
temporada de pesca 2014-15. ................................................................................. 183
Tabla 59– Frecuencias relativas de hembras maduras (promedio 1980-81 a 2012-13)
y porcentaje de aumento (+) o disminución (-) de hembras maduras de acuerdo a cada
escenario climático. ................................................................................................. 188
Tabla 60 - Escenarios de manejo pesquero para el camarón en Sinaloa sur con
escenarios climáticos para la temporada de pesca 2014-15. .................................. 189
Tabla 61 - Escenarios de manejo pesquero para el camarón en Sinaloa sur con
escenarios climáticos con manejo de esfuerzo a MEY para la temporada de pesca
2014-15. .................................................................................................................. 195

XVII
Resumen
La pesquería secuencial de camarón en el Golfo de California es la actividad pesquera
más importante del país. La conforma una flota industrial (Fi) que captura adultos de
camarón azul, café y blanco y otra de pequeña escala (Fpe) que captura en su mayoría
juveniles de blanco. Los camarones peneidos reflejan una alta plasticidad adaptativa
a los cambios ambientales observada en su alta variabilidad internanual reproductiva.
Bajo condiciones de calentamiento moderado la producción pesquera aumenta y bajo
clima extremos disminuye, pero desconociendo su magnitud sobre la pesquería. El
objetivo de la investigación es analizar el efecto de la variabilidad interanual del
reclutamiento sobre la pesquería de camarón para la temporada 2014-15 en Sinaloa
sur y diseñar una estrategia de manejo basado en el Máximo Rendimiento Económico
(MEY) y en el criterio de bienestar Pareto seguridad que beneficie la pesquería y
reduzca los efectos ambientales extremos. Para lo cual se construyó un modelo
bioeconómico multiflota, multiespecie, con coeficientes de mortalidad natural (Mi) y
capturabilidad (qi) dinámicos con la edad, y el segundo también con el tiempo,
empleando un modelo gamma de reclutamiento estacional por especie. Se
representaron escenarios de manejo exclusivamente para la flota industrial. Los
resultados señalan, comparando con condiciones status quo, una reducción de Fi con
MEY (-60%) aumentando las utilidades por cada unidad de esfuerzo (Fi:+868%;
Fpe:+12%). Aun conservando el número de barcos originales y disminuyendo los días
de pesca con MEY las utilidades aumentaron en Fi:+339% y Fpe:7%. Bajo escenarios
sin manejo con calentamiento moderado las utilidades aumentan a Fi:+583% y
Fpe:+279% y en condiciones de enfriamiento hay pérdidas de Fi:-52% y Fpe:-30%. En
las mismas condiciones ajustando Fi en MEY se pudo aumentar las utilidades
industriales (Fi:+364% y Fpe: -29%). Se concluye que la Fpe mostró mayor
vulnerabilidad a ambiente por solo explotar una especie y operar en acceso abierto. El
uso de MEY siempre promovió el aumentó stock desovante y reclutamiento (objetivo
de conservación) y del beneficio a la sociedad (objetivo Económico). Además el
modelo sugiere una estrategia de manejo que ayuda a reducir los efectos de las
interacciones ambientales con el sector pesquero.

XVIII
Abstract
The sequential shrimp fishery of the Gulf of California is the most important fishery of
México. It comprises an industrial fleet (Fi) which targets adults of blue, brown and
white shrimp and a small scale fleet (Fss) which targets mostly white’s juveniles. The
penaid shrimp has shown high adaptive plasticity to environmental variability observed
in their highly variable reproductive performance. Under moderate warming climatic
conditions the fishery production grows and under extreme conditions it diminishes, but
we still ignore its magnitude over the shrimp fishery. The study objective is to analyze
the interannual variability of recruitment over the shrimp fishery season 2014-15 in
south Sinaloa, and to elaborate fishery management scenarios under Maximum
Economic Yield (MEY) criteria for the industrial fleet and using a Pareto security welfare
criteria to benefit the fishery’s production and to reduce the negative effects of the
environmental variability. To achieve the later a construction of a multifleet,
multispecies, age structure bioeconomical model with dynamic coefficients of natural
mortality and capturability at age, and using a and a gamma recruitment function to
represent recruitment seasonality for each species was made. Results showed in
comparison to the fishery status quo condition an industrial effort reduction (-60%)
under MEY conditions, with utilities gains for each effort unit (Fi:+868%; Fss:+12%).
Using the original Fi boats magnitude but reducing the boats day trips to MEY the
utilities also improve (Fi:+339%;Fss: +7%) Under warming conditions without MEY
management the utilities improve to Fi+583% and Fss +279%. But in cold conditions
the utilities diminished to Fi-52% and Fss -30%. Under this condition but adjusting Fi
botas to MEY it was shown their utilities could improve to +364% not been the case for
Fss (-29%). We conclude that Fss under status quo conditions showed high
vulnerability to negative environmental variations because they only exploit one
species. The use of MEY as a fishery objective showed improvements in the
conservation status of the resource (spawning stock biomass and individual
recruitment) and in the fisheries global utilities. Furthermore it suggests a strategy to
reduce the environmental negative interactions to the shrimp fishery sector. Finally the
proposed bioeconomic model could serve as a managing tool under the supposition of
data availability and could help to explore more managing options.

1
1. Introducción
Los camarones peneidos son crustáceos decápodos bentónicos muy importantes
a nivel mundial por el valor que representa su producción. Ellos habitan los fondos
blandos, son de tamaño grande a pequeño y presentan sexos separados. Su ciclo de
vida es corto (1-2 años), muy fecundos (una hembra produce hasta 500,000 huevos),
con tasas de crecimiento rápidas (alcanzan su madures sexual de 4-5 meses) y su
ciclo de vida está dividido por etapas que ocupan distintos hábitats. Los adultos se
encuentran en el fondo marino, donde se estacionalmente se agregan para fecundar
los huevos de las hembras y luego son liberados a la columna de agua. Los huevos y
primeros estadios larvales son pelágicos y flotan en la columna de agua teniendo
distintas metamorfosis larvarias hasta penetrar las aguas salobres de los sistemas
estuarinos, donde se convierten en postlarvas y cambian a hábitos bentónicos. En
estos lugares cuneros se desarrollan hasta pre-adultos y migran con las mareas
lunares hacia altamar donde terminan su ciclo de madurez y ocurre la reproducción.
Las especies de ciclo de vida corto son muy sensibles a los cambios ambientales
y reflejan una gran plasticidad adaptativa observada en su alta variabilidad
reproductiva en función del ambiente. Las principales variables ambientales que
afectan el desempeño reproductivo en peneidos y reflejado en la producción total del
camarón por temporada son la temperatura, precipitación, salinidad y la producción
primaria (Lluch, 1974; Arosemena, 1976; Castro-Aguirre, 1976; Castro-Ortiz y
Sánchez-Rojas, 1976; Sepúlveda, 1976; Lluch, 1977; Sepúlveda, 1991; Días del
Léon, 1993; Del Valle y Martín, 1995; Cervantes-Hernández et al. 2008).
La especies de peneidos más importantes capturadas en el Pacífico Mexicano
son los camarones Farfantepenaeus californiesis, camarón café; Litopenaeus
stylirostris, camarón azul; y L. vannamei, camarón blanco. Debido a su alta
importancia comercial son especies muy estudiadas en casi todos los aspectos de su
biología, no obstante, en términos de parámetros poblacionales aún no se conocen
sus tasas de mortalidad natural específicas por cada etapa de vida, lo cual será de
mucha importancia al considerarse la supervivencia específica en los modelos
pesqueros empleados para su predicción. Se sabe que al principio en las etapas

2
huevo y larva hay una mortalidad natural masiva que después se va atenuando hacia
valores relativamente constantes en las edades adultas.
Estos organismos presentan un gran interés comercial a nivel mundial debido a
que puede alcanzar precios muy altos en el mercado internacional, lo que incentiva a
que los países desarrollen rápidamente su pesquería. La pesquería del camarón
representa la actividad pesquera más importante del país. Del total nacional
representa el primer lugar en infraestructura (i.e. embarcaciones registradas y longitud
de atraque de los puertos pesqueros nacionales, siendo Mazatlán el más importante),
en valor de la producción pesquera nacional y tercer lugar en volumen de producción
pesquera, primer lugar en valor de exportaciones de productos pesqueros y cuarto en
volumen de exportación de productos pesqueros.
Está regida por la Ley General de Pesca y Acuacultura Sustentables (2015) y
en ella se establecen las condiciones de acceso al recurso así como los derechos y
obligaciones de los usuarios y/o armadores. Otras leyes concurrentes son: a) Ley de
Metrología y Normalización (2009); b) Ley General de Sociedades Cooperativas
(2009); y c) Ley General del Equilibrio Ecológico y Protección al Ambiente (LGEEPA,
2014). Como instrumentos normativos de la pesquería de camarón se encuentran a)
la Norma Oficial Mexicana NOM-002-SAG/PESC-2013 y sus modificaciones, con el
objetivo de ordenar el aprovechamiento de las especies de camarón en aguas de
jurisdicción federal de los Estados Unidos Mexicanos; b) la Carta Nacional Pesquera
(2012) que es un instrumento jurídico normativo donde contiene el estado que guardan
las poblaciones y sus pesquerías, así como los lineamientos y estrategias de manejo
por región de las pesquerías en México; y c) el Plan de Manejo para la Pesquería de
Camarón en el Litoral del Océano Pacífico Mexicano del 2004 con énfasis en la
problemática junto con los objetivos y estrategias de manejo de la pesquería.
La pesquería es denominada de tipo secuencial, esto significa que dos o más
flotas capturan diferentes componentes de las etapas del ciclo de vida del camarón.
La pesquería de camarón del Pacífico está dividida comúnmente en dos flotas que
explotan el recurso: una flota de pequeña escala o artesanal que explota el
componente de juveniles y pre-adultos en las zonas de las lagunas costeras, esteros
y bahías someras (i.e. aguas protegidas), usando embarcaciones menores (con remos

3
o motores de bajo caballaje fuera de borda) y con artes de pesca variados como red
tipo chinchorro, red suripera, red de arrastre tipo chango y red atarraya (mayor detalle
en tabla 1). Sin embargo, la parte sur del estado de Sinaloa hasta Tehuantepec los
pescadores usan la red tipo atarraya y se apoyan de barreras artificiales llamadas
tapos, cuya función es bloquear la migración del camarón juvenil y pre-adulto a sus
zonas de reproducción fuera de las lagunas, donde los individuos se acumulan y son
capturados día y noche (CONAPESCA, 2004).
La otra flota es llamada flota industrial, flota de altura o flota de altamar, y explota
el componente de pre-adultos y adultos en aguas abiertas, emplean embarcaciones
mayores con más de 10m de eslora, con autonomía de hasta 40 días en el mar y usan
de arte de pesca las redes de arrastre que emplean sobre el fondo marino extrayendo
el camarón y otras especies no objetivo denominada fauna de acompañamiento (FAC).
El litoral del Pacifico representa el mayor volumen de captura nacional (86%) y
dentro de esta, el Golfo de California equivale al 80% del volumen total nacional
(CONAPESCA, 2013). Para el 2013 la producción total de camarón para el Golfo de
California fue de 74,000 toneladas (sin contar la producción acuícola) donde el estado
de Sinaloa represento el 56% del total de la producción seguido de Sonora con el
25% (CONAPESCA, 2013).
La proporción de especies capturadas en el Golfo de California varía por hábitat,
región y profundidad. Para la zona sur del Golfo de California (Sinaloa sur) el dominio
de aguas interiores es del camarón blanco (89%) y seguido del azul (11%); para la
zona de altamar la principal especie es el camarón café (55%), seguida del blanco
(41%) y azul (4%) [CONAPESCA, 2013]. Globalmente el camarón café domina las
capturas siendo el 70-80% de la producción (López-Martínez et al., 2010).
Las evaluaciones de los recursos pesqueros se basan en la modelación de las
pesquerías en función de la calidad de datos disponibles. Estos modelos en su mayoría
fueron creados para peces tropicales y no tropicales, y no fueron diseñados para
evaluar especies de ciclos cortos como el caso de los peneidos (García y Le Reste,
1997). De acuerdo con García y Le Reste (1997) existen modelos globales (ó de
producción), modelos analíticos (p.e. rendimiento por recluta, técnicas de simulación,

4
modelo estructurado por edades) y métodos aproximativos (p.e. estimaciones de
biomasa por comparación).
En México la gran mayoría de los modelos pesqueros para peneidos se basan
en la producción excedente, debido a la poca disponibilidad de la información biológica
– pesquera, y buscan como objetivo de manejo el máximo rendimiento sostenible
(definición: es la producción máxima constante que se puede tomar año con año,
Sissenwine, 1978) [Plan de manejo para el camarón del litoral del Pacífico Mexicano,
2004]. Por desgracia los supuestos del modelo son incompatibles con la biología,
dinámica poblacional y la pesquería secuencial de los camarones peneidos como los
de estabilidad poblacional y ambiental, la dinámica poblacional (crecimiento,
mortalidad y reclutamiento) se incluyen indirectamente en la relación esfuerzo –
captura, una capturabilidad constante para todas las edades, y no permite manipular
la mortalidad por pesca usando la captura por edades para regular la pesquería
secuencial.
Otro tipo de modelos bioeconómicos con supuestos más realistas a la dinámica
secuencial en peneidos son los modelos estructurados por edades donde se puede
seguir a una cohorte en el tiempo, con procesos de reclutamiento, crecimiento y
mortalidad (natural y por pesca). Este tipo de modelos se ha incorporado al INAPESCA
(2002) para el análisis del estado de las poblaciones de peneidos y el inicio de la veda
para las temporadas pesqueras en el Pacífico Mexicano.

5
2. Antecedentes
2.1 Biológicos
2.1.1. Biología general
Las tres especies habitan en la plataforma continental asociados a sustratos
blandos (Brusca y Hendrickx, 2008). Los adultos del camarón azul y blanco habitan
aguas oceánicas hasta los ≈72m de profundidad, con sus mayores densidades entre
0 y 18m. El camarón café habita hasta los 100m de profundidad y su mayor densidad
varía por zona y época. Se tiene reportado para el norte del Golfo de California que
sus poblaciones se localizan entre 0 y 50m de profundidad, mientras que en para la
porción sur están entre 18 - 72m, además esta especie presenta movimientos
estacionales de repliegue y alejamiento de la costa (Lluch, 1982).
El camarón azul y el blanco dependen de los sistemas estuarinos y lagunares
en fases tempranas de su ciclo de vida (Tabla 1). Este no es el caso estricto para el
camarón café, considerado mayormente de hábitos marinos y sus juveniles raramente
se encuentran dentro de las zonas lagunares (Rodríguez de la Cruz y Rosales, 1976;
Holthuis, 1980).
Tabla 1 - Distribución y hábitat de las principales especies comerciales de camarón en el Pacífico Mexicano (Brusca y Hendrickx, 2008).
a) Camarón café b) Camarón azul
c) Camarón
blanco
Registro más
norteño
Bahía de San Francisco,
California, EUA
Punta Abre Ojos,
BCS, y Punta
Willards, BC, México
Yavaros, Sonora,
México
Distribución en
el Golfo de
California
GCN, GCC, GCS, SOB, RB GCN, GCC, GCS GCC, GCS
Registro más
sureño Callao, Perú Paita, Perú
Caleta de la Cruz,
Perú

6
Profundidad
más somera 2m 5m 5m
Profundidad
más grande 180m 45m 72m
Hábitat general
Litoral bentónico, Lagunas
costeras, lagunas con
mangles
Litoral bentónico,
Lagunas costeras,
lagunas con mangles
Litoral bentónico,
Lagunas costeras,
lagunas con mangles
Sustrato Arena y lodo Arena y lodo Arena y lodo
Comunidad Plataforma continental Plataforma continental Plataforma continental
* GCN: Golfo de California Norte, GCC: Golfo de California Centro, GCS: Golfo de California Sur,
SOB: Región Sur-Oeste de Baja California Sur (de Los Cabos a Bahía Magdalena), RB – Reserva de
la Biosfera del Alto Golfo de California y Delta del Río Colorado.
La estructura trófica de los peneidos (tabla 2) se ubica en tres niveles a lo
largo de su ciclo de vida: 1) consumidor primario, 2) consumidor secundario y 3)
detritófago ú omnívoro a nivel adulto (Garduño, 1976).
Tabla 2– Nicho ecológico por estadio de vida de los camarones peneidos comerciales en el Pacífico Mexicano.
Estadío Ubicación Nivel trófico Alimentación Depredadores
Huevo Plancton NA NA
Zooplancton y
organismos
filtradores
Nauplio Plancton Consumidor
primario
Nanoplancton, fitoplancton y
detritus.
Zooplancton, peces
juveniles y filtradores
Protozoea Plancton
Consumidor
secundario
(omnívoro)
Nanoplancton, zooplancton,
fitoplancton y detritus.
Zooplancton, peces
juveniles y filtradores
Mysis Plancton
Consumidor
secundario
(omnívoro)
Zooplancton y fitoplancton. Zooplancton, peces
juveniles y filtradores
Postlarva Plancton-
Bentos
Consumidor
secundario
(omnívoro)
Detritus de plantas
vasculares y microalgas de
la columna y fondo de las
Peces, invertebrados

7
lagunas, así como de
anélido, cadáveres y
microrganismos.
Juvenil Bentónico Detritófago
(omnívoro)
Detritus, moluscos,
anélidos, cadáveres de
peces, esponjas, otros
crustáceos, algas, tallos de
plantas vasculares y el
canibalismo también es
común.
Peces,
invertebrados, aves
Preadulto Bentónico Detritófago
(omnívoro)
Detritus, moluscos,
anélidos, cadáveres de
peces, esponjas, otros
crustáceos, algas, tallos de
plantas vasculares y el
canibalismo también es
común.
Peces,
invertebrados, aves
Adulto Bentónico Detritófago
(omnívoro)
Detritus, moluscos,
anélidos, cadáveres de
peces, esponjas, otros
crustáceos, algas, tallos de
plantas vasculares.
Peces, invertebrados
La dominancia ecológica por especie y hábitat (Lluch, 1982) muestra que
el camarón azul domina en aguas protegidas con salinidades altas o superiores al
promedio marino (35 psu), pudiendo esporádicamente disminuir debido a descargas
de agua dulce continentales. Este hábitat es típico de aguas marinas ligadas a climas
desérticos y semidesérticos en el Golfo de California, como los estados de Baja
California, Sonora y norte de Sinaloa, pero se observa que su abundancia cae
drásticamente a partir del sur de Sinaloa hasta Tehuantepec, donde domina el
camarón blanco. El camarón blanco es dominante en las aguas protegidas de baja
salinidad permanente o estacional que responden al ciclo de lluvias, desde el sur de
Sinaloa hasta Guatemala (Hernández-Covarrubias et al. 2012). En las aguas
protegidas del norte y centro de Sinaloa confluyen las dos especies, dominando hacia
el norte el azul y al sur el blanco.

8
La especie marina dominante en los fondos fuera de las bocas y las lagunas
costeras es el camarón café. En el norte del Golfo de California, en sistemas con alta
salinidad, sus larvas y postlarvas llegan a penetrar las aguas protegidas y bahías
someras por un periodo corto (un mes) y después emigran al alcanzar 40mm de
longitud total (Lt) hacia las aguas marinas. Sus juveniles habitan zonas costeras
someras y únicamente habitan zonas francamente marinas a partir de Sinaloa sur
hasta Chiapas (Rodríguez de la Cruz y Rosales, 1973).
La talla de primera madurez sexual es diferente entre las tres especies de
camarón como se muestra en la tabla 4.
Tabla 3– Las tallas de primera madurez por especie para los camarones del Pacífico
Mexicano.
Especie Tallas L50 Autor
Litopenaeus vannamei 140mm Sepúlveda, 1991.
L. stylirostris 170mm García-Gómez, 1976; Aragón-Noriega y
Alcántara-Razo, 2005.
Farfantepenaeus californiensis 117mm García-Gómez, 1976.
* L50 longitud total a la cual el 50% de los individuos de la población están sexualmente maduros.
La talla de primera madurez sexual es diferente entre las tres especies de
camarón como se muestra en la tabla 4.
Tabla 4– Las tallas de primera madurez por especie para los camarones del Pacífico
Mexicano.
Especie Tallas L50 Autor
Litopenaeus vannamei 140mm Sepúlveda, 1991.
L. stylirostris 170mm García-Gómez, 1976; Aragón-Noriega y
Alcántara-Razo, 2005.
Farfantepenaeus californiensis 117mm García-Gómez, 1976.
* L50 longitud total a la cual el 50% de los individuos de la población están sexualmente
maduros.

9
La maduración sexual no depende únicamente de la edad, también de la latitud
y condiciones climáticas dominantes. En el Golfo de California se ha observado que la
maduración sexual se ha relacionado con el aumento de la temperatura, de la
productividad primaria o con el inicio de época de lluvias (i.e. sur del Golfo de
California) [Del Valle y Martín, 1995]. En la figura 1 se describen los principales
periodos de madurez gonadal reportados para el Golfo de California, donde para el
camarón café se reporta una maduración continua anual con máximos en verano y
otro menos importante en invierno. Para el camarón blanco es en verano y otro menor
en invierno, y para el camarón azul es en primavera-verano.
ene feb mar abr may jun jul ago sep oct nov dic
Fc
Lv
Ls
* Fc: Farfantepenaeus californiensis; Lv: Litopenaeus vannamei; Ls: L. stylirostris.
Figura 1 – Periodos de madurez sexual en hembras de camarón para el Golfo de
California (estudio centrado en Mazatlán) Las barras grises muestran los picos de
madurez (Lluch, 1977).
En las costas de Sinaloa la presencia de hembras maduras (INP, 2013) desde
1981 a 2013 (fig. 2) muestra un patrón de reproducción similar en las especies azul y
blanco, con sus picos más importantes en primavera y verano; de manera diferencial
el camarón café presentó una mayor presencia de hembras maduras todo el año con
picos en verano-otoño.

10
Figura 2– Frecuencia promedio de hembras maduras o en fase III para las costas de Sinaloa de acuerdo al INP (2013).
Los camarones peneidos presentan sexos separados, las hembras presentan
una estructura llamada télico y machos el pestasma, localizadas en el primer par de
pleópodos (apéndices natatorios). La reproducción ocurre cuando los individuos de
ambos sexos se agregan y el macho introduce el petasma con el espermatóforo
adherido en el télico de la hembra, de manera que son fecundados conforme los
huevos son expulsados al medio (Fischer et al.1995). La fecundidad total promedio
varía por especie y por talla (tabla 5).
Tabla 5- Valores de la fecundidad mínima, máxima y promedio para de las especies
de camarón café (Farfantepenaeus californiensis), azul (Litopenaeus stylirostris) y
blanco (L. vannamei) basado en la literatura reportada.
Especie Fecundidad Tallas (mm) Referencias
Litopenaeus
vannamei
Mínima = 80,0001
Promedio = 265,0001
Máxima = 450,0001
𝐿𝑇50 = 1402
𝐿𝑇𝑀𝐴𝑋 = 2101,3
1 Hernández-Covarrubias et al., 2012
2 Sepúlveda, 1991
3 Chávez, 1973
L. stylirostris Mínima = 60,0004 𝐿𝑇50 = 1705 4 Hernández-Covarrubias et al., 2003

11
Promedio = 350,0004
Máxima = 650,0004
𝐿𝑇𝑀𝐴𝑋 =2406 5 García-Gómez, 1976
6 Castro-Ortiz y Sánchez-Rojas, 1976
F. californiensis Mínima = 100,0005
Promedio = 550,0005
Máxima = 1,000,0005
𝐿𝑇50 = 1175
𝐿𝑇𝑀𝐴𝑋 = 2407
7Chávez y Rodríguez de la Cruz,
1971
*LT = Longitud total (mm); Fecundidad = numeró promedio de huevos
La etapa huevo en peneidos dura de 10 a 16 horas antes de eclosionar las
larvas nauplio (tabla 6). El desarrollo larvario para las especies del Pacífico se observa
en la tabla 15. En resumen son 11 estadios larvarios: 5 estadios nauplio, 3 de
protozoea y 3 de mysis que duran en promedio 20 días. El ciclo de vida total por
especie reportado en la literatura es de 411 días para el camarón azul, 565 días para
el camarón blanco y 365 para el camarón café (tabla 15).
Tabla 6- Duración del estadio de huevo reportado para de las especies de camarón
café (Farfantepenaeus californiensis), azul (Litopenaeus stylirostris) y blanco (L.
vannamei).
Duración de estadio de huevo
Especie (horas) Referencia
Litopenaues vannamei 10 Andrade-Vizcaíno (2010); Torres-Acuña (2008);
Kitani (1986a) 13
16
L. stylirostris 13
Kitani (1986b); Prahl y Gardeazábal (1977) 14
15
F. californiensis 13 Kitani y Alvarado (1982); Schafer (1971);
Rodríguez de la Cruz (1976) 14
15
La migración reproductiva de los camarones hacia aguas oceánicas varía para
cada especie (tabla 7). El camarón blanco abandona las lagunas entre 90-150mm, el

12
azul entre 150-200 mm y para el camarón café los individuos reportados al norte del
Golfo de California abandonan las lagunas costeras a los 40 mm y existen otros
reportes de arrastre en altamar donde reportan la migración de los 80 a los 120mm.
13
Tabla 7- Tallas de madures y migración y la edad (meses) de acuerdo a las curvas de
crecimiento por especie
Tallas de migración (mm)
Localidad Autor
Litopenaeus stylirostris (azul)
150 Teacapán-Chametla, Sinaloa Soto-López 1973
170 Centro Sinaloa promedio Castro-Ortiz y Sánchez Rojas 1976
200 Bahía Altata-Ensenada del Pabellón, Sin. Castro-Ortiz y Sánchez-Rojas 1976
180 B. Ceuta y La Reforma, Sinaloa Castro-Ortiz y Sánchez-Rojas 1976
210 Puerto Peñasco, Sonora Galicia, 1976
90 Mazatlán, Sinaloa Sáenz y Lluch, 1990
190-200 Navachiste, Sinaloa INP, 2002 (Proyecto Sin-Son)
162 Agiabampo, Sonora INP, 2002 (Proyecto Sin-Son)
150 Santa María La Reforma INP, 2002 (Proyecto Sin-Son)
160 Centro de Sinaloa INP, 2003
165 Santa María La Reforma, Sinaloa INP, 2004
Litopenaeus vannamei (blanco)
90-120 Teacapán y Ticama, Sinaloa Núñez et al. 1951
100-110 Mazatlán a Teacapán, Sinaloa Lluch y Guzmán del Proó, 1972
90-121 Huizache –Caimanero, Sinaloa Soto-López, 1973
90-122 Chametla-Teacapán, Sinaloa Soto-López, 1973
100 Teacapán-Chametla, Sinaloa Soto-López 1973
105 Sinaloa Magallón y Jacquemin 1976
110-130 Huizache –Caimanero, Sinaloa Sepúlveda-Medina, 1976
90-140 Las Cabras "Secas", sur Sinaloa Gutiérrez, 1980
105-140 Las Cabras "lluvioso", sur Sinaloa Gutiérrez, 1980
83-120 Laguna mar muerto-Oax/Chiapas Ramos-Cruz, 2000
150 Navachiste, Sinaloa INP, 2002 (Proyecto Sin-Son)
148 Agiabampo, Sonora INP, 2002 (Proyecto Sin-Son)
110 Sur de Sinaloa INP, 2003
Farfantepenaeus californiensis (café)
105 Mazatlán, Sinaloa Olguín, 1967
35-40 Sonora Mathews, 1981; Sepúlveda, 1991
110 Mazatlán, Sinaloa Barreiro, 1986
100 Centro de Sinaloa INP, 2003
120 Navachiste, Sinaloa INP, 2002 (Proyecto Sin-Son)
117 Agiabampo, Sinaloa INP, 2002 (Proyecto Sin-Son)

14
Resumen Migración laguna-mar
(mm)
Edad
(meses)
L. vannamei 90 - 140 2.5 - 4.8
L. stylirostris 150 - 200 4 - 9
F. californiensis *35 – 40; **100-120 1-1.5; 3 - 4
*Penetra lagunas como postlarva y las abandona a los 35mm; **capturas de arrastre con barcos en
zona costeras donde se acumulan los juveniles hasta los 30 m.
2.1.2. Parámetros de edad y crecimiento
La edad en los peneidos solo puede ser estimada por métodos indirectos debido
a la falta de estructuras duras, como otolitos en el caso de los peces, que dejen una
huella constante y medible en el organismo. De tal forma el método indirecto más
empleado es el análisis de frecuencias de tallas, donde se da un seguimiento a las
modas (cohortes) en un tiempo determinado estimando su edad y crecimiento. Dentro
de las estimaciones de las edades máximas que alcanzan los peneidos reportadas
abarcan de 1 a 2.5 años (García y Le Reste, 1987). Las especies de camarón blanco,
azul y café presentan un rango de 1-1.6 años de edad máxima.
Los parámetros de edad y crecimiento han sido obtenidos en todo el Golfo de
California incluyendo la costa occidental del estado de Baja California Sur (tabla 8).
Para el caso del camarón blanco únicamente se cuenta con el trabajo publicado de
Chávez (1973) que incluye a todas las edades del camarón; otros trabajos abarcan
únicamente el crecimiento en lagunas o en altamar.
Tabla 8 – Parámetros de edad y crecimiento estimados para cada especie de camarón.
𝑾∞ 𝑳∞ K t0 Rango de tallas
empleado Lugar Referencia
(gr) (mm) (año) (mes)
Litopenaeus vannamei (blanco)
26.6 200 3.19 0.292 58-192 Lt Mazatlán Chávez, 1973
130 2.94 0.5 40-130 Lt
Sinaloa (bahías) Sepúlveda-Medina,
1976
15 136 5.66 0.016 11-145Lt Tehuantepec
(bahías) Ramos-Cruz, 2000
230 2.83 1.42 35-235Lt
Sinaloa y Nayarit Romero-Moreno,
2009

15
L. stylirostris (azul)
117.42 229 2.57 -0.507 23-221Lt Sonora Galicia, 1976
- 245 2.14 -0.477
Sonora Rodríguez de la
Cruz, 1976
194 244 2.78 0.660 Sinaloa centro
Castro-Ortiz y
Sánchez-Rojas, 1976
191.77 242 2.53 0.569 25-200 Lt Sinaloa La Reforma
190.25 242 2.56 0.296 24-220 Lt Sinaloa Altata-
Ensenada Pabellón
- 246 2.76 0.273
Sonora García de Quevedo,
1990
- 244 2.04 -0.080 55-216 Lt Sonora 1993-94
López Martínez et
al., 2005 - 246 1.92 -0.085 55-216 Lt Sonora 1996-97
- 246 1.92 -0.085 55-216 Lt Sonora
Farfantepenaeus californiesis (café)
119 239 1.63 -0.759 23 – 210Lt Sonora Galicia, 1976
220 242 2.23 0.14 0.022 – 260Lt
Golfo de California Chávez y Rodríguez,
1971
- 227 0.95 -
Tehuantepec Sierra-Rodríguez y
Reyna Cabrera, 1993
- 239 1.88 - 52-233 Lt
Sonora (1978-1994) López-Martínez,
2000
133 245 1.89 0.76 68-238 Lt Bahía Magdalena García-Borbón, 2007
122 237 1.48 68-242 Lt Bahía Magdalena
(1995-2007)
García-Borbón,
2009
Existen muchos estudios sobre las tasas de crecimiento diario (mm/día) por
especie, pero estas solo son válidas para las tallas y los tiempos que abarca el estudio
(blanco: Menz y Bowers, 1980; café: Sepúlveda y Soto, 2001 azul: Sepúlveda, 1999).
Lluch (1972) para el área de Sinaloa sur englobo el crecimiento para peneidos
comerciales en 1 mm diario.
Para asignar edades a los organismos presentes en las capturas se elaboran
claves edad – longitud, estas representan las distribuciones de cada grupo de edad en
términos de una normal estándar con el objeto de asignar una función clave-edad que
indique las probabilidades relativas de que un individuo de longitud L pertenezca a una
o varias edades. Esto permite la asignación de un número de individuos a edades
correspondientes, permitiendo evaluar las variaciones anuales en las estructura de

16
edades de las capturas y de la población. Actualmente solo se cuenta con la clave
edad-longitud del camarón café (tabla 9).
Tabla 9– Funciones clave edad-longitud en camarones del Pacífico Mexicano.
Especie Función clave edad-longitud Lugar Referencia
F. californiensis
y = -0.0575x2 + 0.7697x + 2.8188
R² = 0.6292
y = desviación estándar de las
edades; x = edad
Sonora López-Martínez,
2000
F. californiensis
y = -0.070977 + 22.007425
R² = 0.8984
y = desviación estándar; x =
longitud total media y esta se
transforma a edad.
Bahía Magdalena,
BCS
García-Borbón,
2009
2.1.3. Mortalidad natural
La estimación del parámetro de mortalidad natural o M es crítica para la
dinámica poblacional, pesquera y los análisis de producción de reclutas y de cohortes.
Este parámetro es vital para la modelación y evaluación de los stocks pesqueros que
finalmente sirven para el desarrollo y aplicación de planes efectivos de manejo (Caddy,
1991; Ramírez-Rodríguez y Arreguín-Sánchez, 2003). Los sesgos causados por la
sobre o subestimación de M afectan los análisis de evaluación y administración de los
recursos sujetos a explotación pudiendo tener consecuencias muy graves sobre los
stocks pesqueros (Ramos-Cruz et al., 2006).
El “axioma de la M constante” es el supuesto automático que se hace en la
evaluación de stock pesqueros de peces e invertebrados, donde para la explotación
de una estructura de tallas o edades diferenciales se usa la misma probabilidad de
muerte por causas naturales (Caddy, 1991). Del mismo modo, de manera
convencional, en casi todos los modelos dinámicos de biomasa se incluye una M

17
constante; lo cual es aceptable donde la pesca incide únicamente en una edad (e.g.
generalmente adultos), y es empleado correctamente para la evaluación de stocks
(Vetter 1988; Caddy 1996). Sin embargo esto no aplica en pesquerías secuenciales
donde distintos componentes del ciclo de vida del recurso son explotadas, como
sucede en camarones peneidos.
Las estimaciones de M en peneidos en el Pacífico Mexicano se han enfocado
al componente explotado (pre-adultos y adultos), en menor medida a juveniles y casi
ninguno al componente larvario y de huevo (tabla 10). Los rangos de M año-1 en la
literatura para las tres especies abarcan de 46.8postlarva a 0.78adulto en L. vannamei;
10.08juvenil a 0.48adulto en L. stylirostris; y de 1559huevo a 0.24adultos en F. californiensis.
Estos valores cuales fueron derivados de distintos métodos indirectos e
independientes para estimar la M, los cuales de manera general están basados en su
relación con datos pesqueros y el coeficiente de capturabilidad (Silliman, 1943); con la
edad máxima, talla máxima y el coeficiente de crecimiento “K” (Beverton y Holt, 1959);
con los sobrevivientes y la mortalidad (Ricker, 1958); con la edad promedio de
madurez de la especie (Rikhter y Efanov, 1976); con parámetros de crecimiento (i.e.
von Bertalanffy) y variables de temperatura promedio ambiental (Pauly, 1980); y con
el coeficiente instantáneo de crecimiento en un tiempo determinado (Jensen, 1996).
Muchos de estos modelos se deben tomar con precaución porque la precisión de los
datos empleados para cada parámetro es desconocida (i.e. se desconocen métodos
confiables para estimar la edad en crustáceos por ausencia de estructuras duras como
en peces) y como consecuencia los valores de M pueden presentar sesgo (García y
Le Reste, 1987).
Esta grave suposición de una M constante se ha solventado por medio del
modelo de intervalos gnomónicos (Caddy, 1991, 1996; Morales-Bojórquez et al., 2003
Martínez-Aguilar et al. 2005), el cual asume que el stock está en condiciones estables,
así cada individuo reemplaza al otro, y su ciclo de vida se puede dividir en intervalos
de tiempo donde la probabilidad de muerte por causas naturales se puede asumir
constante, esto es satisfecho por la estrategia de división gnomónica del ciclo de vida
(i.e. los intervalos de tiempo desiguales incrementan su duración proporcionalmente
con la edad). El término “gnomónico” significa distancias lineares constantes movidas

18
por la sombra de gnomon que corresponden a unidades desiguales del tiempo (Caddy,
1996). El método no describe efectos denso dependientes o condiciones ambientales
atípicas pero es muy útil en condiciones ambientales y denso-dependientes promedio
(Ramírez-Rodríguez y Arreguín-Sánchez, 2003).
El método de intervalos gnomónicos ya ha sido empleado exitosamente en
especies anuales como en los calamares Loligo forbesi y L. vulgaris en el canal inglés
(Royer et al., 2002), y en Dosidicus gigas en el golfo de California (Martínez-Águilar et
al., 2010); y también en camarones peneidos (Farfantepenaeus duorarum) del golfo
de México (Ramírez-Rodríguez y Arreguín-Sánchez, 2003) y de la costa occidental de
la península de Baja California (García-Borbón, 2009). También se ha evaluado la M
a la edad en especies multianuales como la sardina del Pacífico (Sardinops caeruleus)
[Martínez-Águilar et al., 2005] y el serranido Epinephelus morio del Golfo de México
(Giménez et al., 2009).
Dentro del modelo gnomónico también se ha examinado la variabilidad en los
vectores estimados de M con relación a la fecundidad media y la duración de la etapa
huevo. Se ha observado en especies anuales una relación directa entre la fecundidad
media y los valores de M (Ramírez-Rodríguez y Arreguín-Sánchez, 2003); y se ha
observado una relación inversa entre la duración de la etapa huevo y la sobrevivencia,
así como diferencias en los últimos intervalos de M donde la principal fuente de
variabilidad se relacionó con la etapa huevo (en sardina, Martínez-Aguilar et al., 2005;
en calamar Martínez-Aguilar et al., 2010).

19
Tabla 10– Referencias de valores estimados de mortalidad natural en las especies de
camarón blanco (Litopenaeus vannamei), azul (L. stylirostris) y café (Farfantepenaeus
californiensis) en el Pacífico mexicano.
Estadio
Mortalidad
natural M
(anual)
Método Sitio Referencias
a) Litopenaeus vannamei
Huevo
Nauplio
Protozoea
Misis
Postlarva 46.8
Diferencia entre
postlarvas entrantes
y captura comercial
Laguna Huizache
Caimanero, Sinaloa Edwads, 1977
Juvenil
3.24 - 25.321
21.322
Método Ricker
(1975)1
Marcación de
juveniles2
Laguna Huizache-
Caimanero,
Sinaloa1,2
Sepúlveda, 19811
Edwads,19772
Preadulto 6.6 - 8.4 Método Ricker
(1975)
Topolobampo,
Sinaloa Sepúlveda, 1999
Adulto
1.92 - 4.983
1.924
0.78 – 2.525
Pauly (1980)3
Ricker (1959)5
Golfo de
Tehuantepec3,4
Mazatlán, Sinaloa5
Ramos-Cruz et al.,
20063
Rodríguez de la
Cruz y Chávez,
19964
Jaquemín, 19765
b) Litopenaeus stylirostris
Huevo
Nauplio
Protozoea
Misis
Postlarva
Juvenil

20
Preadulto 6.34
0.96-10.08
Método Ricker
(1975)
Sonora norte
Sistema lagunar de
Bahía Magdalena
Sepúlveda, 1999
Adulto
1.566
0.75 - 1.985
3.37
0.968,9
0.7210
0.99 - 1.4511
2.4 12a
0.48-6.7212b
Método de Beverton-
Holt (1959) 6-7,9-11
Silliman (1943)8
Ricker (1958)10
Ricker (1975)12
Sonora sur6
Sonora norte7,10,11,12a
Mazatlán,
Sinaloa8,9,5
Topolobampo,
Sinaloa12b
Rodríguez de la
Cruz, 19766
García de Quevedo,
19907
Lluch,19748
Saenz y Lluch,
19909
García-Gómez,
197610
Ross-Terrazas,
198811
Sepúlveda, 199912
c) Farfantepenaeus californiensis
Huevo 1,559.513 Modelo gnomónico Bahía Magdalena García-Borbón ,
200913
Nauplio 55313 Modelo gnomónico Bahía Magdalena
Protozoea 106.813
Modelo gnomónico Bahía Magdalena
Misis
Postlarva 47.613 Modelo gnomónico Bahía Magdalena
Juvenil 13.613 Modelo gnomónico Bahía Magdalena
Preadulto 7.8 – 8.512
8.113
Ricker (1975)12
Modelo gnomónico13
Topolobampo,
Sinaloa12
Bahía Magdalena13
Adulto
2.28 - 3.63
2.284
0.24-1.935
1.328
1.08 – 310
2.23 - 2.3611
2.3812a, 3.7212b
Pauly (1980)3,16a
Ricker (1958)5,10
Silliman (1943)8
Ricker (1975)11,12ab
Modelo gnomónico13
Rikhter y Efanov
(1976)15a.16b
Golfo de
Tehuantepec3,4
Mazatlán, Sinaloa5,8
Sonora norte10,11,12a
Topolobampo,
Sinaloa12b
Litoral
Sonorense15abc
Rodríguez de la
Cruz, 197614
López-Martínez,
200015abc
García-Borbón,
200716ab

21
2.713
1.214
2.1115a, 2.7215b,
2.82 15c
1.2216a, 2.1316b
Pauly (1980) 15b
Jensen (1996)15c
Bahía
Magdalena13,16ab
2.1.4. Variabilidad ambiental y reclutamiento
Dentro de las pesquerías y la dinámica poblacional existen procesos que
aumentan la biomasa de la población, como es la entrada de nuevos individuos o
reclutas a la pesquería, y el aumento de su biomasa mediante el crecimiento individual.
Las pesquerías en general se ven afectas por la variabilidad del reclutamiento que se
presenta cada año y obedece en su gran mayoría a causas ambientales. En especies
de vida corta como el camarón, pelágicos menores y el calamar el efecto del clima
sobre la reproducción es notorio y se puede observar en escalas anuales y la biomasa
capturada refleja esta relación reclutamiento-ambiente.
Las especies de ciclo de vida corto son muy sensibles a los cambios
ambientales y reflejan una gran plasticidad adaptativa observada en su alta variabilidad
reproductiva en función del ambiente. Se ha observado acortamiento o amplitud de
sus épocas reproductivas (% hembras maduras) y la temporalidad del desove (Leal,
1999; Leal-Gaxiola et al., 2001; López-Martínez et al., 2005; Romero-Sedano et al.,
2004; Aragón-Noriega y Alcántara-Razo, 2005; Aragón-Noriega, 2007; Aragón-
Noriega et al., 2012), la variación en el tamaño de la hembras sexualmente maduras
(López-Martínez, 2000; López-Martínez et al., 2005) y tamaños y densidades de las
postlarvas (Edwards, 1975; Macías-Regalado y Calderón-Pérez, 1979; Poli y
Calderón-Pérez, 1985), todas en función de las variables ambientales más importantes
que reportan en la literatura: la temperatura superficial del mar (TSM), la precipitación
y la salinidad (Lluch, 1974; Arosemena, 1976; Castro-Aguirre, 1976; Castro-Ortiz y
Sánchez-Rojas, 1976; Sepúlveda, 1976; Lluch, 1977; Sepúlveda, 1991; Días del Léon,
1993; Del Valle y Martín, 1995) en algunos otros mencionan la productividad marina
(Cervantes-Hernández et al. 2008) y los índices climáticos (Álvarez-León et al.,2009;

22
Castro-Ortiz y Lluch-Belda, 2008). La TSM es la variable más significativa porque actúa
directamente en la fecundidad de la población coadyuvando la maduración de los
ovarios, el número de desoves parciales, viabilidad de huevos y del desarrollo
embrionario y la supervivencia de las fases larvarias (duración del periodo larvario)
(Del Valle y Martín, 1995), y además que se correlaciona en cierto grado con el resto
de variables ambientales. En la tabla 11 se hace un breve resumen por especie de la
relación abundancia-reclutamiento con las principales variables ambientales.

23
Tabla 11– Principales variables ambientales y su efecto sobre la abundancia y reclutamiento de los peneidos dentro
del Golfo de California.
Relación variable con abundancia
T°C óptima
(reproducción)
T°C
letal T°C Precipitación Salinidad
Hábitat
adultos
Hábitat
juveniles Referencias
Lv
26°C13;
24-28°C(límites de
tolerancia)13,14
23-30°C (para
crecimiento)16;
<15°C y
>33°C16 (+)13,17 (+)13 (-)1,5,18
Mar
abierto1
Lagunas
costeras1
Lluch, 19771; Salgado,
19762; Arosamena,
19763; Rodríguez de la
Cruz, 19734; Macías-
Regalado y Calderón-
Pérez, 19795; Leal,
19996; López-Martínez,
20007; López-Martínez
et al., 20058; Romero-
Sedano et al., 20049;
Leal-Gaxiola et al.,
200110; Aragón-Noriega
y Alcántara-Razo,
200511; Aragón-Noriega,
200712; Castro-Ortiz y
Lluch-Belda 200813;
Sierra et al., 200014;
Cervantes-Hernández et
al., 200815; Vázquez-
Melchor, 201116;
Aragón-Noriega et al.,
201217; Poli y Calderón-
Pérez, 198518
Ls
24-29°C2; 25-30°C4;
27-29°C12; 26°C13
24-28°C(límites de
tolerancia)13,14
23-30°C (para
crecimiento)16;
>30°C2;
<15°4
<15°C y
>33°C16
(+)12
(+) ≈reclutamiento
marino máximo1;
(-) en lluvias
invernales13
(-)1,5,18 Mar
abierto1
Lagunas
costeras1
Fc
25-30°C;
24.5-25-5°C7;
27-30°C9;
23-27°C11;
<5°C y
>35°C3
<10°C4;
<19°C y
>27°C7
(+)6,8,11,15
hasta su
límite
fisiológico
(+) ≈su máximo
reclutamiento en
sep1; (-)15
(+) en PL5 Mar
abierto1
Lagunas
costeras
hasta
juvenil9 o
puede
prescindir
de ellas10

24
26°C13
24-28°C(límites de
tolerancia)13,14.
*Lv: Litopenaeus vannamei ó camarón blanco; Ls: L. stylirostris o camarón azul; Fc: Farfantepenaues californiensis o
camarón café. (+) correlación positiva; (-) correlación negativa.

25
Sus ciclos reproductivos (desoves de hembras maduras) pueden presentarse
todo el año como se observa en zonas tropicales en el Golfo de Tehuantepec (Aragón-
Noriega y Alcántara-Razo, 2005; Aragón-Noriega, 2007; Cervantes-Hernández et al.,
2008; Aragón-Noriega et al., 2012) o disminuir su amplitud temporal a picos
reproductivos estacionales en épocas de primera-verano como sucede en zonas
subtropicales-templadas como es el Alto Golfo de California (Aragón-Noriega y
Alcántara-Razo, 2005; Aragón-Noriega, 2007). Existen zonas intermedias donde
dependiendo del tipo de ambiente dominante (dominio cálido o frío) puede tener una
reproducción continua o presentar picos estacionales de reproducción como sucede
en Mazatlán, Sinaloa (Aragón-Noriega y Alcántara-Razo, 2005; Aragón-Noriega, 2007;
Aragón-Noriega et al., 2012). El modelo base del ciclo reproductivo para el camarón
en el Golfo de California es un pico reproductivo en primavera-verano y otro secundario
en otoño-invierno (Leal 1999; Castro-Ortiz y Lluch-Belda, 2008). Cada uno es muy
importante para los fines pesqueros porque el de primavera-verano sostiene la
producción de la pesquería de camarón de todo el pacífico mexicano, y los de otoño-
invierno forman los reproductores de primavera- verano que sostienen la pesquería.
Algunos estudios logran asociar la intensidad de la magnitud y la distribución
del reclutamiento reproductivo, empleando variables como densidad de larvas,
porcentaje de hembras maduras y pulsos de reclutas pesqueros con retro-cálculos
para estimar su nacimiento, relacionándolas con índices climáticos de alta frecuencia
como son el fenómeno El Niño y La Niña. En la tabla 12 se indica el efecto de cada
evento sobre la reproducción en camarones peneidos incluyendo las variables que se
emplean para estudiar la relación ambiente y reproducción (Leal, 1999; Leal-Gaxiola
et al., 2001; López-Martínez, 2000; López-Martínez et al., 2003; López-Martínez et al.,
2005; Castro-Ortiz y Lluch-Belda 2008). Donde se observa que en épocas de
calentamiento hay mayor porcentaje de hembras maduras, los periodos de
reproducción aumentan y existe un efecto positivo sobre el reclutamiento de individuos;
y ocurre la disminución de las misma variables cuando los eventos de calentamiento
son muy extremos o en eventos de enfriamiento (evento Niña) cuando se supera el
límite fisiológico de la especie (tabla 11).
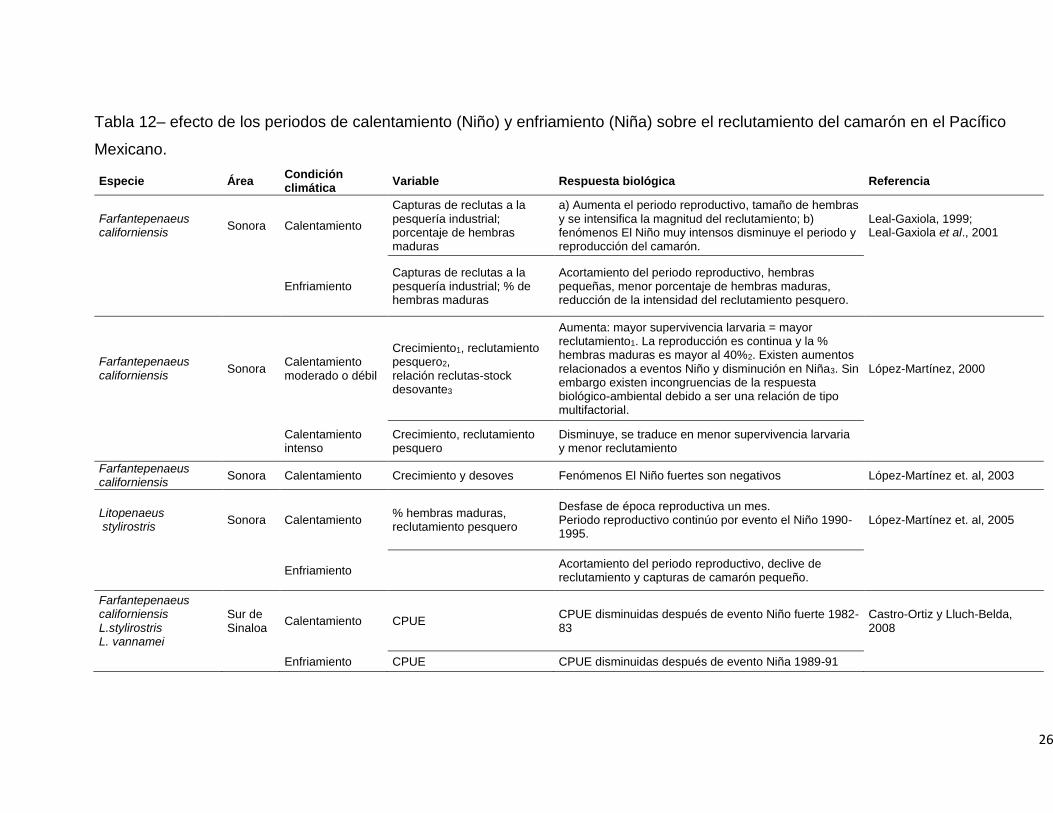
26
Tabla 12– efecto de los periodos de calentamiento (Niño) y enfriamiento (Niña) sobre el reclutamiento del camarón en el Pacífico
Mexicano.
Especie Área Condición climática
Variable Respuesta biológica Referencia
Farfantepenaeus californiensis
Sonora Calentamiento
Capturas de reclutas a la pesquería industrial; porcentaje de hembras maduras
a) Aumenta el periodo reproductivo, tamaño de hembras y se intensifica la magnitud del reclutamiento; b) fenómenos El Niño muy intensos disminuye el periodo y reproducción del camarón.
Leal-Gaxiola, 1999; Leal-Gaxiola et al., 2001
Enfriamiento Capturas de reclutas a la pesquería industrial; % de hembras maduras
Acortamiento del periodo reproductivo, hembras pequeñas, menor porcentaje de hembras maduras, reducción de la intensidad del reclutamiento pesquero.
Farfantepenaeus californiensis
Sonora Calentamiento moderado o débil
Crecimiento1, reclutamiento pesquero2, relación reclutas-stock desovante3
Aumenta: mayor supervivencia larvaria = mayor reclutamiento1. La reproducción es continua y la % hembras maduras es mayor al 40%2. Existen aumentos relacionados a eventos Niño y disminución en Niña3. Sin embargo existen incongruencias de la respuesta biológico-ambiental debido a ser una relación de tipo multifactorial.
López-Martínez, 2000
Calentamiento intenso
Crecimiento, reclutamiento pesquero
Disminuye, se traduce en menor supervivencia larvaria y menor reclutamiento
Farfantepenaeus californiensis
Sonora Calentamiento Crecimiento y desoves Fenómenos El Niño fuertes son negativos López-Martínez et. al, 2003
Litopenaeus stylirostris
Sonora Calentamiento % hembras maduras, reclutamiento pesquero
Desfase de época reproductiva un mes. Periodo reproductivo continúo por evento el Niño 1990-1995.
López-Martínez et. al, 2005
Enfriamiento Acortamiento del periodo reproductivo, declive de reclutamiento y capturas de camarón pequeño.
Farfantepenaeus californiensis L.stylirostris L. vannamei
Sur de Sinaloa
Calentamiento CPUE CPUE disminuidas después de evento Niño fuerte 1982-83
Castro-Ortiz y Lluch-Belda, 2008
Enfriamiento CPUE CPUE disminuidas después de evento Niña 1989-91
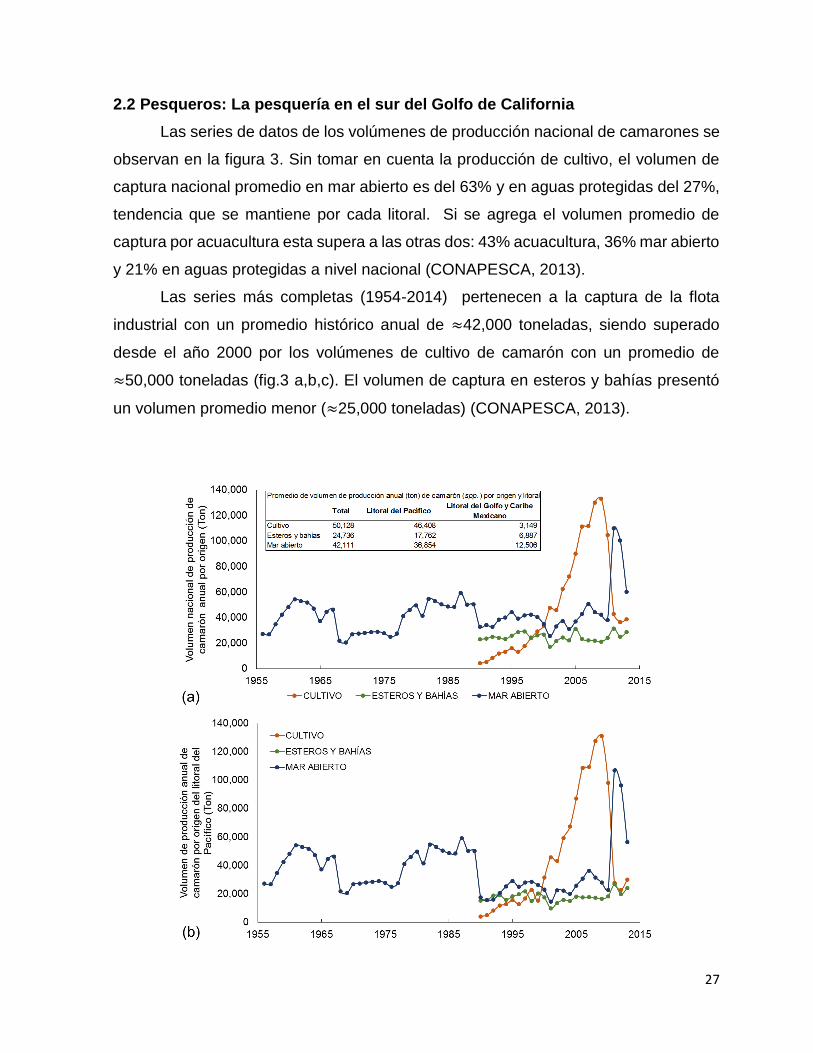
27
2.2 Pesqueros: La pesquería en el sur del Golfo de California
Las series de datos de los volúmenes de producción nacional de camarones se
observan en la figura 3. Sin tomar en cuenta la producción de cultivo, el volumen de
captura nacional promedio en mar abierto es del 63% y en aguas protegidas del 27%,
tendencia que se mantiene por cada litoral. Si se agrega el volumen promedio de
captura por acuacultura esta supera a las otras dos: 43% acuacultura, 36% mar abierto
y 21% en aguas protegidas a nivel nacional (CONAPESCA, 2013).
Las series más completas (1954-2014) pertenecen a la captura de la flota
industrial con un promedio histórico anual de ≈42,000 toneladas, siendo superado
desde el año 2000 por los volúmenes de cultivo de camarón con un promedio de
≈50,000 toneladas (fig.3 a,b,c). El volumen de captura en esteros y bahías presentó
un volumen promedio menor (≈25,000 toneladas) (CONAPESCA, 2013).

28
Figura 3– Volúmenes de producción de camarón por origen: cultivo, esteros y bahías
y altamar a nivel nacional (a), litoral del pacífico (b) y Golfo y Caribe Mexicano (c).
La producción pesquera de camarón en el Pacífico Mexicano (1990-2013) es la
más importante a nivel nacional, produciendo el 86% del volumen total nacional y
dentro de esta, el Golfo de California equivale al 80% del volumen total nacional
(CONAPESCA, 2013). El promedio anual (1990-2013) de captura en el Pacífico
mexicano para la flota de altamar es de 32,200 tons y para esteros de 17,900
tons (tabla 13).
Dentro del volumen de producción estatal promedio para el litoral del Pacífico
(tabla 13) la flota de altamar del estado de Sinaloa presentó la mayor producción
(�� ≈15,000 tons/año) seguido de Sonora (�� ≈11,000 tons/año). En cuanto a la captura
de la flota de pequeña escala nuevamente Sinaloa fue el principal productor (�� ≈7,500
tons/año) seguido de Nayarit (�� ≈3,300 tons/año) y Sonora (�� ≈2,500 tons/año).
Para el 2013 (CONAPESCA, 2013) la producción total de camarón para el Golfo
de California fue de 74,697 toneladas (sin contar la producción acuícola, figura 4)
donde Sinaloa represento el 56% del total de la producción seguido de Sonora con
el 25%.
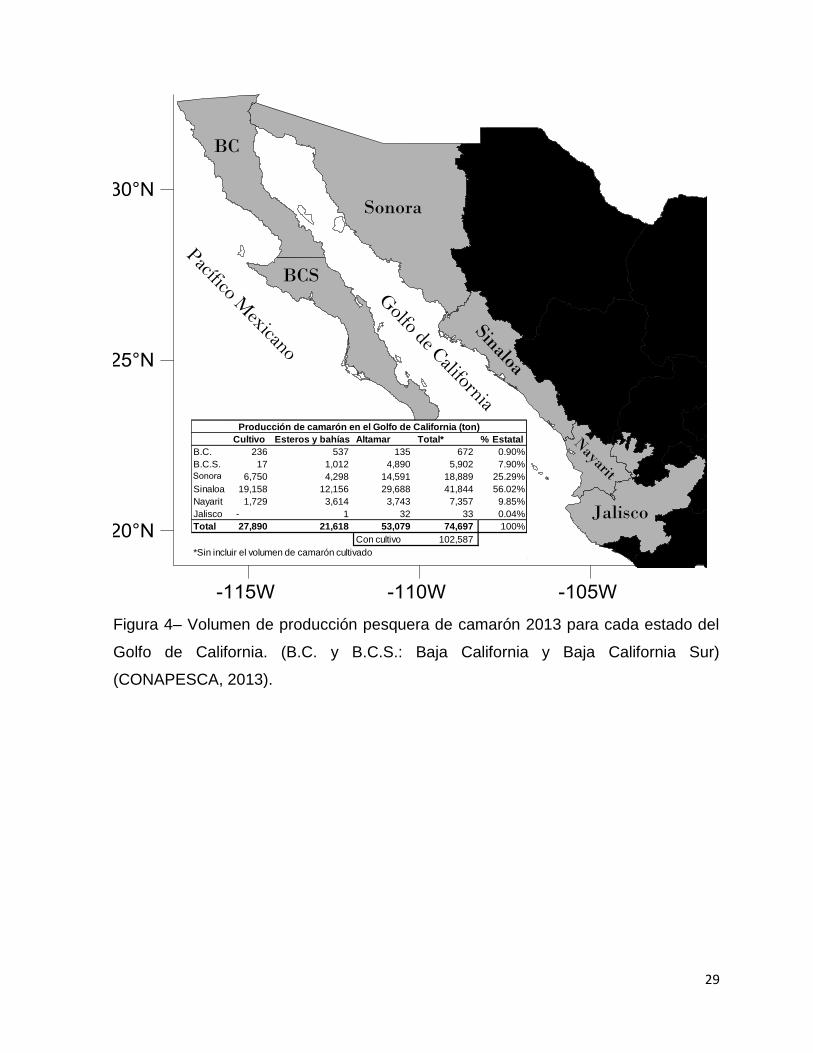
29
Figura 4– Volumen de producción pesquera de camarón 2013 para cada estado del
Golfo de California. (B.C. y B.C.S.: Baja California y Baja California Sur)
(CONAPESCA, 2013).
Cultivo Esteros y bahías Altamar Total* % Estatal
B.C. 236 537 135 672 0.90%
B.C.S. 17 1,012 4,890 5,902 7.90%
Sonona 6,750 4,298 14,591 18,889 25.29%
Sinaloa 19,158 12,156 29,688 41,844 56.02%
Nayarit 1,729 3,614 3,743 7,357 9.85%
Jalisco - 1 32 33 0.04%
Total 27,890 21,618 53,079 74,697 100%
Con cultivo 102,587
*Sin incluir el volumen de camarón cultivado
Producción de camarón en el Golfo de California (ton)

30
Tabla 13 – Volumen de producción peso vivo de camarón en toneladas para el litoral del Pacífico Mexicano de 1990-2013
incluyendo la referencia nacional (CONAPESCA, 2013).
a) Producción en mar abierto
Nacional Litoral del
Pacífico B.C. B.C.S. SON SIN NAY JAL COL MICH GUE OAX CHIS
Ʃ 1,068,657 768,525 11,782 21,636 277,705 362,863 34,430 1,214 9,749 10 1,870 22,351 24,915
�� 44,814 32,224 491 902 11,571 15,119 1,435 110 406 10 78 1,064 1,038
𝜎 3,200 5,231 255 1,794 8,592 11,325 2,284 322 554 - 111 639 578
n 17 11 24 24 24 24 24 11 24 1 24 21 24
% - - 1.53% 2.82% 36.13% 47.22% 4.48% 0.16% 1.27% 0.00% 0.24% 2.91% 3.24%
b) Producción en esteros y bahías
Ʃ 591,565 426,276 3,052 18,747 64,481 179,265 78,799 958 568 1 397 33,882 46,126
�� 25,486 17,916 254 781 2,687 7,469 3,283 60 24 1 23 1,412 1,922
𝜎 682 2,258 260 557 1,386 2,049 1,518 203 34 - 28 927 895
n 17 11 12 24 24 24 24 16 24 1 17 24 24
% - - 0.72% 4.40% 15.13% 42.05% 18.49% 0.22% 0.13% 0.00% 0.09% 7.95% 10.82%

31
La proporción de especies capturadas varía por zona (i.e. altamar/aguas
protegidas) y hábitat en el Pacífico Mexicano. Se relaciona a factores ambientales
como la salinidad y las precipitaciones (INP, 2013; INP, 2014a,b). Las especies
eurihalinas como el camarón blanco toleran bajas salinidades encontradas en las
aguas protegidas y se vuelve la especie dominante en las capturas de aguas
interiores a partir del sur de Sinaloa hasta Tehuantepec (figura 5b). Hacia la parte
norte de Sinaloa, zona del Alto Golfo de California y la zona de Bahía Magdalena,
en la costa occidental de BCS, la salinidad rebaza la media promedio y el dominio
en los esteros y bahías someras es del camarón azul (figura 5b).
La especie dominante en las capturas de altamar (Fig. 5a) es del camarón
café en todo el Golfo de California incluyendo la costa occidental de BCS. Se tienen
registros que hacia el sur de Sinaloa el camarón blanco puede llegar a ser la especie
dominante en las capturas industriales (Núñez et al., 1951) y en Sinaloa centro el
camarón azul puede dominar las capturas industriales (INP, 2014a,b).
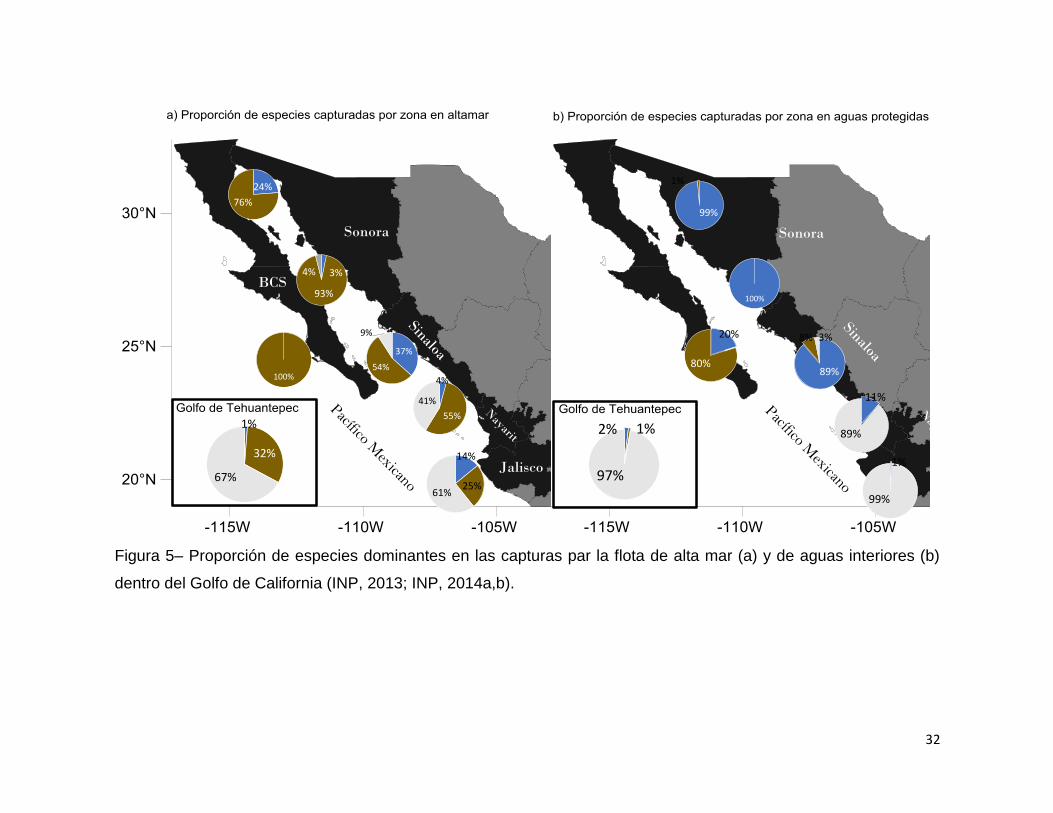
32
Figura 5– Proporción de especies dominantes en las capturas par la flota de alta mar (a) y de aguas interiores (b)
dentro del Golfo de California (INP, 2013; INP, 2014a,b).
24%
76%
3%
93%
4%
37%
54%
9%
4%
55%
41%
100%
14%
25%61%
1%
32%
67%
99%
1%
100%
89%
8% 3%
11%
89%
1%
99%
20%
80%
2% 1%
97%

33
Como se ha observado Sinaloa es el estado más importante a nivel nacional
en cuanto a producción camaronera tanto silvestre como acuícola (Fig. 4). Debido
a que su litoral costero cuenta con grandes extensiones de esteros y sistemas
lagunares costeros en todo su litoral. La CONAPESCA (2007) dividió el estado en
norte, centro y sur, donde la región centro es la que más producción aporta al estado
(fig. 6b).
Figura 6 - Proporción de volúmenes (ton) de captura de camarón silvestre en el
estado de Sinaloa. CONAPESCA (2007).
En un reporte que generó la CONAPESCA (2007) del comportamiento
histórico de la producción total de camarón en los esteros de Sinaloa (2000-2007)
(tabla 14) se reportó que la parte centro históricamente tiene la máxima producción

34
(50%) del total estatal con 26,000 tons y promedio anual de 1,073 tons, siendo el
sistema más productivo Bahía de Santa María-La Reforma; le sigue la parte norte
con 19,000 tons con promedio anuales de 1,180 tons, siendo los sistemas más
productivos las Bahías de Ohuira-El Colorado-Topolobampo-Agiabampo; y al final
la región menos productiva fue la zona sur con 7,137 tons y promedios de 149 tons
anuales siendo el sistema más productivo Huizache-Caimanero.

35
Tabla 14- Producción pesquera del estado de Sinaloa (tons) en bahías, esteros y altamar (2000 – 2007).
Sinaloa Ecosistema Lagunr 2000 2001 2002 2003 2004 2005 2006 2007 Total Media anual
Norte
Bahías de Ohuira-El
Colorado-Topolobampo-
Agiabampo
1,859 1,275 1,772 1,478 1,572 2,036 1,916 1,877
18,903 1,181±611 DE
Bahía de Navachiste-
Macapule-San Ignacio 811 677 637 440 431 507 1,160 456
Centro
Bahía de Santa María- La
Reforma 2,548 2,042 1,496 1,254 1,579 2,250 3,038 1,713
25,748 1,073±881 DE Bahía de Altata-Ensenada
del Pabellón 1,319 1,157 949 1,058 811 1,115 1,767 1,040
Bahía de Ceuta 34 12 35 89 246 84 64 45
Sur
Estero Mazatlán-San
Ignacio 103 44 35 82 45 65 44 38
7,137 149±161 DE
Laguna Huizache 211 143 143 200 177 349 109 106
Laguna Caimanero 884 348 451 334 240 205 548 30
Marismas las Cabras 82 27 47 106 228 268 181 92
Agua Grande-Revolución-
Teacapán 76 9 24 75 74 154 262 3
Marisma de Palmillas 44 39 86 126 63 42 42 57
Subtotal 7,971 5,774 5,674 5,241 5,466 7,076 9,130 5,457 51,788 2,403
Altamar 9,631 7,034 11,029 11,120 6,440 13,516 16,725 23,679 99,174 11,541

36
La producción pesquera reportada por CONAPESCA 2014 y CONAPESCA
2015 para el estado de Sinaloa por origen mensual (esteros lagunas y bahías vs
mar abierto) solo incluye el valor del recurso “camarón” (figura 7 y tabla 15). La
producción de camarón de mar abierto fue casi el doble (56%) que en aguas
interiores (tabla 15). Se observa en la figura 7a un máximo de producción en
septiembre y su caída hasta diciembre, con ligero repunte en enero a marzo. Por
otro lado la producción industrial (Fig. 7b) presenta un máximo de producción en
octubre (25%) seguido de diciembre (20%) y un repunte en febrero y marzo (≈15%).
Se reportó para la temporada 2014-15 una producción total de “camarón” en aguas
interiores de 11,000 tons y en aguas marinas de 20,900 tons (tabla 15).
Figura 7 – Captura de camarón para el estado de Sinaloa reportado por el Anuario Estadístico de Acuacultura y Pesca para los años 2014 y 2015. a) Producción de aguas interiores y b) producción de mar abierto.

37
Tabla 15 - Peso vivo reportado para las capturas silvestres por el Anuario Estadístico
de Acuacultura y Pesca en Sinaloa para los años 2014 y 2015 para el recurso camarón.
(CONAPESCA 2014, 2015).
Año Mes Origen del camarón Peso vivo (tons)
2014 septiembre esteros, lagunas y bahías 3,246
2014 octubre esteros, lagunas y bahías 2,289
2014 noviembre esteros, lagunas y bahías 1,084
2014 diciembre esteros, lagunas y bahías 702
2015 enero esteros, lagunas y bahías 1,491
2015 febrero esteros, lagunas y bahías 1,488
2015 marzo esteros, lagunas y bahías 1,463
Total 11,763
2014 octubre mar abierto 5,282.73
2014 noviembre mar abierto 3,061.77
2014 diciembre mar abierto 4,247.44
2015 enero mar abierto 2,174.61
2015 febrero mar abierto 3,124.60
2015 marzo mar abierto 3,010.32
Total 20,901

38
2.3. Pesqueros: variables pesqueras
2.3.1. Esfuerzo pesquero en Sinaloa sur
Se considera una unidad de pesca como la entidad básica, elemental de captura
y se define esfuerzo pesquero como la cantidad de unidades de pesca que se dedican
a la captura de un recurso determinado en una cierta área durante un cierto tiempo
determinado (Lluch, 1977). Para el caso de altamar las variables barcos registrados,
barcos en operación, días de pesca, días efectivos de pesca, lances, horas de pesca,
y área barrida.
Para el caso de Sinaloa sur la única medida de esfuerzo con la que se cuenta
es el número de barcos reportados para la flota de altamar por estado en los Anuarios
de Pesca de 1980-2013 (Tabla 16). Para el caso de Sinaloa se observa como se ha
ido disminuyendo el esfuerzo en número de barcos presentando un máximo de 785
barcos hasta 469 barcos (2013). Para la flota de aguas interiores no existe información
completa de esfuerzo y solo existen volúmenes mensuales o anuales por especie.

39
Tabla 16- Embarcaciones de altamar registradas por estado para el litoral del Pacífico
mexicano (Anuarios de Pesca).
AÑO SUBTOTAL B.C. B.C.S SON. SIN. NAY. JAL. COL. MICH. GRO. OAX. CHIS.
1980 1,540 51 13 664 509 4 - 74 7 14 166 38
1981 1,692 68 21 743 557 3 - 77 - 11 164 48
1982 1,657 60 34 712 580 5 1 31 6 8 187 33
1983 1,681 64 45 686 588 19 6 37 8 8 186 34
1984 1,557 58 38 593 570 24 13 29 8 8 181 35
1985 1,551 59 36 611 563 24 13 29 8 7 171 30
1986 1,489 53 36 587 532 25 13 29 9 8 170 27
1987 1,510 62 41 595 524 37 13 28 9 9 166 26
1988 1,512 61 38 591 528 44 11 29 9 14 160 27
1989 1,521 66 38 591 529 44 11 29 9 17 160 27
1990 1,552 88 29 602 504 65 13 33 9 25 138 46
1991 1,505 69 21 593 496 54 18 34 10 29 137 44
1992 1,504 69 21 592 496 56 18 34 10 28 136 44
1993 1,509 69 21 596 498 56 18 34 9 28 135 45
1994 1,519 57 26 631 528 34 11 32 10 11 138 41
1995 1,463 47 27 621 549 16 5 30 6 7 129 26
1996 1,475 48 27 616 568 17 5 26 4 7 133 24
1997 1,313 36 16 514 567 11 - 25 - 3 109 32
1998 1,313 36 16 514 567 11 - 25 - 3 109 32
1999 1,313 36 16 514 567 11 - 25 - 3 109 32
2000 1,649 48 25 615 761 18 - 35 - 6 113 28
2001 1,665 41 28 622 775 18 - 35 1 6 104 35
2002 1,674 43 28 625 776 20 - 34 1 7 103 37
2003 1,673 42 28 613 786 21 - 35 1 7 101 39
2004 1,674 43 27 616 785 21 - 35 1 7 101 38
2005 1,594 43 27 568 764 20 - 34 1 7 95 35
2006 1,536 41 27 521 767 20 - 34 1 6 86 33
2007 1,508 41 27 511 752 20 - 34 1 6 83 33
2008 1,508 41 27 511 752 20 - 34 1 6 83 33
2009 1,428 41 27 484 718 18 - 31 1 6 77 25
2010 1,352 38 27 454 682 16 - 31 1 6 72 25
2011 1,326 38 27 441 672 16 - 31 1 6 69 25
2012 1,321 38 27 441 668 16 - 31 1 6 69 24
2013 850 21 16 281 469 5 - 17 - 1 37 3

40
2.3.2. Dinámica del esfuerzo de alta mar
La dinámica de la flota industrial registrada en cada puerto también es
importante porque lo barcos no son exclusivos del área marítima territorial (relativa a
los estados litorales (INP, 2002). La flota más “invasora” es la de Mazatlán (barras
negras, Fig. 8), pero en general todas las flotas registradas en cada puerto migran a
otras zonas en busca del recurso. Esto trae consecuencias en la calidad de los datos
públicos oficiales de captura porque al no ser exclusivos de su área de pesca no se
puede conocer con certeza las captura y la proporción real de las especies por zona.
Figura 8– Porcentaje de lances de cada flota por zonas de pesca asignadas por el INP.
Donde los puertos son: Guaymas (GY); Mazatlán (MZ); Pto Chiapas (PC); Pto Madero
(PM); Pto Peñasco (PP); Salina Cruz (SZ); San Blas (SB); San Carlos (SC); San Felipe
(SF); Topolobampo (TB); y Yavaros (YV). Las zonas son: Las zonas estudiadas por
cada CRIP son: Zona 10 (Ensenada); Zona 50 (La Paz); Zona 20 (Guaymas); Zonas
30 (Sinaloa centro y norte),40 (Sinaloa sur) y 60 (Bahía de Banderas, Nay.); y Zona 90
(Salina Cruz).
2.3.3. Selectividad
La selectividad es la probabilidad diferencial de captura de individuos de cierta
talla en función de al arte de pesca, siendo para el caso de redes la abertura de malla.

41
Siendo las redes un arte de pesca selectivo, esto es permiten el escape de individuos,
y la probabilidad de escape para una red dada disminuye directamente en función a la
talla de individuos (Lluch, 1977).
Para una abertura de red existe una curva sigmoidea que define la probabilidad
de retención a cada talla, representando las probabilidades acumulativas.
𝑆𝐿 =1
(1+𝐸𝑋𝑃(𝑆1−𝑆2∗𝐿)) (1)
𝑆𝐿: Selectividad a la talla L
𝑆1 = 𝑎 y 𝑆2 = 𝑏: constantes
Con las constantes se puede obtener un rango de probabilidades de selección
donde:
𝐿25% = (𝑆1 − ln 3) 𝑆2⁄ (2)
𝐿50% = 𝑆1 𝑆2⁄ (3)
𝐿75% = (𝑆1 + ln 3) 𝑆2⁄ (4)
Ya existen curvas de selectividad por artes de pesca (i.e. red de arrastre con
abertura de malla oficial de 2 ½ pulgadas y atarraya con abertura oficial de 1 ½
pulgadas) para la zona de Mazatlán (Fig. 9; Lluch, 1974; Lluch, 1977).
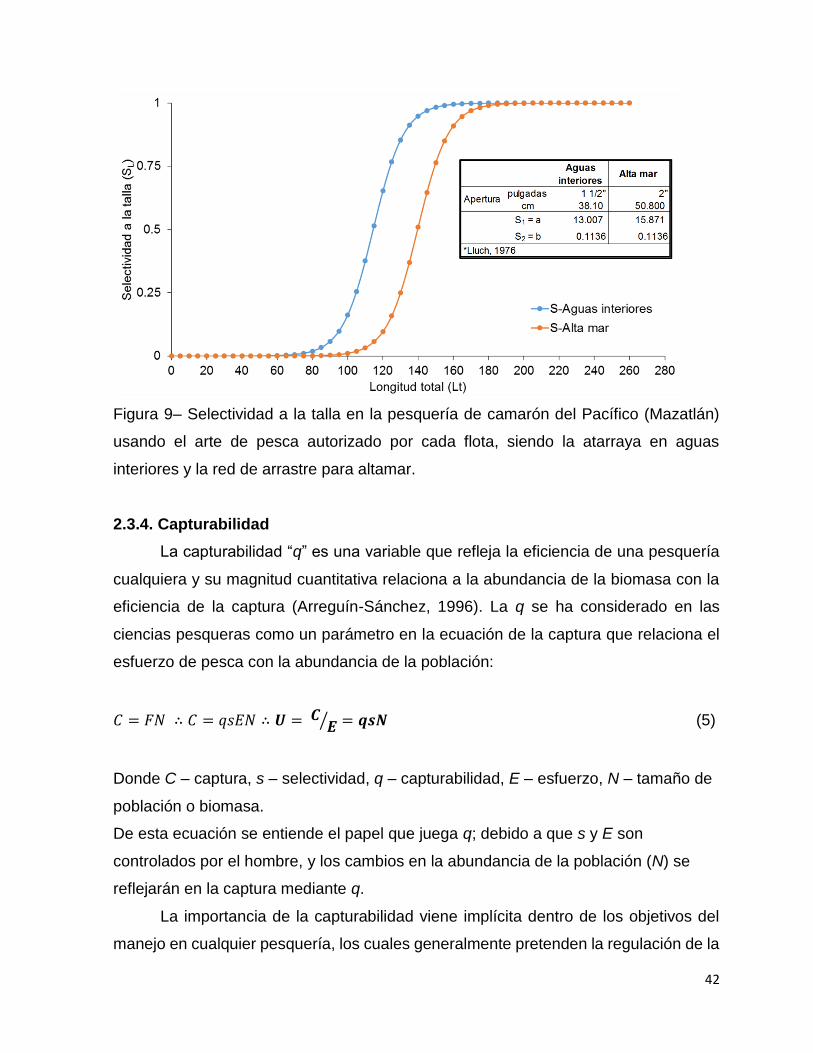
42
Figura 9– Selectividad a la talla en la pesquería de camarón del Pacífico (Mazatlán)
usando el arte de pesca autorizado por cada flota, siendo la atarraya en aguas
interiores y la red de arrastre para altamar.
2.3.4. Capturabilidad
La capturabilidad “q” es una variable que refleja la eficiencia de una pesquería
cualquiera y su magnitud cuantitativa relaciona a la abundancia de la biomasa con la
eficiencia de la captura (Arreguín-Sánchez, 1996). La q se ha considerado en las
ciencias pesqueras como un parámetro en la ecuación de la captura que relaciona el
esfuerzo de pesca con la abundancia de la población:
𝐶 = 𝐹𝑁 ∴ 𝐶 = 𝑞𝑠𝐸𝑁 ∴ 𝑼 = 𝑪 𝑬⁄ = 𝒒𝒔𝑵 (5)
Donde C – captura, s – selectividad, q – capturabilidad, E – esfuerzo, N – tamaño de
población o biomasa.
De esta ecuación se entiende el papel que juega q; debido a que s y E son
controlados por el hombre, y los cambios en la abundancia de la población (N) se
reflejarán en la captura mediante q.
La importancia de la capturabilidad viene implícita dentro de los objetivos del
manejo en cualquier pesquería, los cuales generalmente pretenden la regulación de la

43
mortalidad de pesca (F) ó la captura, comúnmente sobre el componente de adultos o
a veces sobre otras etapas del ciclo de vida de cada recurso, como sucede en las
pesquerías secuenciales (e.g. camarón y mero rojo). Esta regulación muchas veces
se logra implementando vedas (donde no hay relación con la capturabilidad); y/o
regulando el esfuerzo pesquero, donde q es fundamental por su relación entre la
captura y la abundancia de la población (ecuación 5). Así, la abundancia del recurso y
la captura son estimaciones elementales en la evaluación pesquera y la q está
involucrada en ambas estimaciones (Arreguín-Sánchez, 1996; Seijo et al., 1997).
Con lo anterior se puede definir a la capturabilidad como una medida de
interacción entre el esfuerzo pesquero y la abundancia del recurso (Arreguín-Sánchez,
1996). Otros autores la define como la fracción promedio de la población total que es
capturada por unidad de esfuerzo pesquero (Lluch, 1977); también la definen como la
proporción de individuos en el área de pesca “A” removidos por el arte de pesca en un
área “a” con una eficiencia “c”, (Baranov, 1918; Gulland 1964,1983; Caddy, 1979),
siendo:
𝑞 = 𝑐(𝑎 𝐴⁄ ) (6)
Esta relación no toma en cuenta el comportamiento de la población o la estrategia de
pesca, de tal modo que la capturabilidad se definiría mejor como la mortalidad por
pesca por unidad de esfuerzo o unidad de intensidad de pesca:
𝐹 = 𝑞 ∗ 𝐸 ∴ 𝑞 = 𝐹/𝐸 (7)
Donde 𝐹: mortalidad por pesca; 𝑞: capturabilidad, 𝐸: esfuerzo pesquero.
Lo anterior se puede interpretar de dos maneras: a) como el número de peces o
biomasa por unidad de esfuerzo pesquero, que puede reflejar las variaciones en su
abundancia (Beverton y Holt, 1957); ó b) si la abundancia poblacional y el esfuerzo se
encuentran constantes o en equilibrio, las variaciones en q reflejarán la vulnerabilidad
del recurso al equipo de pesca, estrategia de pesca y biología propia del recurso,

44
incluyendo la respuesta de la población a factores ambientales (Arreguín-Sánchez,
1996).
La literatura sobre la dinámica pesquera de los recursos (e.g. pesquería de
camarón en México; tabla 15) no estudian a la q per se, sino su aplicación directa en
los modelos de producción donde normalmente toma un valor constante, lo cual afecta
negativamente la calidad y resolución de los modelos pesqueros. Y además se ha
hecho la observación (Arreguín-Sanchez, 1996) que los coeficientes de capturabilidad
son solo válidos para las condiciones en las cuales fueron estimados.
Así la aplicación de un valor constante de capturabilidad implicaría:
a) las estimaciones aplican solo a individuos con capturabilidad similar;
b) las estimaciones aplican en condiciones bajo las cuales se obtuvieron los datos (i.e.
temperatura, área, etc);
c) el esfuerzo pesquero es de la misma calidad;
d) la población es cerrada.
El problema con el supuesto de constancia es que en realidad la capturabilidad
es muy variable, y este supuesto es violado continuamente por pesquerías con
recursos pelágicos (MacCall, 1976, 1990; Csirke, 1989), crustáceos (Ye y Mohammed,
1999) y moluscos (Chávez, 2000). Las agregaciones reproductivas (Arreguín-
Sánchez, 1996; Arreguín Sánchez y Pitcher 1999), variación espacio-tiempo en la
distribución (Prince, 1992), cambios en la estrategia pesquera (Arreguín-
Sánchez,1996; e.g. cambio en potencia de pesca: Gulland, 1983), factores
ambientales (Castro-Ortiz y Lluch-Belda, 2008), y el comportamiento del propio
pescador (Chávez, 2000) también producen fuentes de variación en la capturabilidad.
Por ejemplo, para el caso de organismos que se agregan para protegerse (i.e.
pelágicos menores), al disminuir su biomasa por factores como la depredación
(incluida la pesca) o ambientales, esto causaría un aumento en q reflejando la
agregación de los sobrevivientes de varios cardúmenes pero conllevaría a una rápida
sobrexplotación y posible extinción local de la subpoblación del recurso por la
disminución en su abundancia, lo cual no se puede observar bajo el supuesto de q
constante (Arreguín-Sánchez, 1996; Pérez y Defeo, 2003).

45
Otra manera de citar las fuentes de variación en la estimación de q es la
propuesta por Arreguín-Sánchez (1996) donde menciona que las fuentes son
asociadas a la selectividad, accesibilidad, disponibilidad y vulnerabilidad del recurso (*
Vulnerabilidad: es la probabilidad de encuentro entre el arte de pesca y el recurso. Si
hay disponibilidad del recurso, la vulnerabilidad dependerá de la eficiencia del arte. i.e.
la selectividad). Otro factor que afecta tanto la captura por unidad de esfuerzo como
medida de abundancia, y la capturabilidad, es el esfuerzo de pesca, sobre todo cuando
el poder de pesca es significativamente diferente entre barcos o flotas, donde es
recomendable la estandarización del esfuerzo (Arreguín-Sánchez, 1996).
La mayoría de las estimaciones del coeficiente de capturabilidad (Leslie y
Davies, 1939; Silliman.1943; DeLury, 1947; Paloheimo, 1958) suponen: una estructura
poblacional constante entre todos los individuos en el tiempo; un esfuerzo pesquero
uniformemente distribuido y constante; y un tamaño poblacional constante (Arreguín-
Sanchéz, 1996). Una diferente interpretación de la capturabilidad surge cuando se
trabaja con modelos por estructura de edades/longitudes (Arreguín-Sanchéz, 1992;
Arreguín-Sanchéz, 1996; Anderson y Seijo, 2010) y algunos empleando los análisis de
población virtual (VPA) (Gulland 1965; Murphy, 1965; Pope,1972; Jones, 1974, 1981),
donde si se asume que la mortalidad por pesca cambia con la edad. Al trabajar con
estos modelos se debe evaluar desde la perspectiva de cada clase de edad o talla
porque cada pez o individuo se comporta diferencialmente de acuerdo con su etapa
del desarrollo, área de distribución y actitud con respecto al arte de pesca; todos los
aspectos anteriores más la variabilidad en el tiempo afectan la estimación de
capturabilidad.
Arreguín-Sánchez (1992, 1996) propuso un modelo de capturabilidad a la edad
(qi) basado en una matriz de transición para estimar la capturabilidad a la edad de
donde se obtiene la relación q = f(L), donde conjuga los elementos de la estructura de
las longitudes/edades de la captura, probabilidades de crecimiento y sobrevivencia,
mortalidad natural y esfuerzo para obtener q(L). Pudiendo solventar los problemas
relacionados a modelos de VPA donde a priori a) se debe asumir un patrón de
capturabilidad en forma de una función exponencial (Pope y Shepherd, 1985); b) el

46
error involucrado en la estimación de q se acumulan durante el retro cálculo de VPA
introduciendo errores en la evaluación de la mortalidad por pesca.
2.3.4.1. Modelos de capturabilidad aplicados a la pesquería de camarón en
México
La gran mayoría de las estimaciones de q en los modelos pesqueros nacionales
fueron estimadas a partir de datos de captura para la flota de altamar (tabla 17). Existe
una gran variabilidad en los valores estimados de q por factores biológicos, espacio-
temporales, diferencias de poder de pesca y modelos de estimación. La gran mayoría
de los modelos aplicados en la pesquería nacional del camarón (Tabla 17) estiman la
q como un índice que mide la relación de la CPUE y la abundancia de la población
(e.g. método de Leslie); o un índice que relaciona a la mortalidad por pesca con el
tamaño poblacional (método de Paloheimo). Otros autores usan el modelo de
producción Schaefer donde la q viene implícita dentro del modelo. Son muy escasos
los estudios que incluyen a la capturabilidad tomando en cuenta a la estructura de
edades de la captura y en el tiempo dentro de la pesquería nacional (INP, 2002;
García-Borbón, 2009).
47
Tabla 17 – Estimaciones independientes de capturabilidad en la literatura haciendo
énfasis en la pesquería de camarón en México.
Modelo de capturabilidad Flota: valores q/spp Lugar Referencia
q=(Ct+1+Et+1)/Nt+1
Leslie y Davies, 1939
Altamar (mes):
Azul: 0.00004-0.003
Blanco: 0.00005-0.00003
Café: 0.00001-0.00005
Mazatlán,
Sinaloa
Jacquemin,
1976
q = pendiente de la relación C/E
y K
DeLury (Ricker, 1958)
Altamar (temporada)
azul: 0.0006-0.0009
café: 0.0002 - 0.0004
Peñasco, Sonora García-Gómez,
1976
q’t=qmin + f(µ)
μ = madurez (talla) NA Mazatlán, Sin Lluch,1977
F=qf ∴ Ct/ft = qN0
Método de Leslie (Ricker, 1975)
Altamar (q temporada):
c. roca: 0.03 - 0.043
q mensual:
0.00559 - 0.00711
Quintana Roo,
Isla Mujeres
Arreguín-
Sánchez, 1981
Método de Leslie (Ricker, 1975)
Altamar (q temporada)
azul: 0.00041 - 0.00008
café: 0.00024 - 0.00007
Guaymas,
Sonora
Ross-Terrazas,
1988
Método de Leslie (Ricker, 1975)
Altamar (q /temporada)
azul (PP): 0.0020 -
0.0036
azul (T):0.00018 -
0.000053
café (PP): 0.0010 -
0.0015
café (T): 0.00010 -
0.00004
Puerto Peñasco
"PP", Sonora
Topolobampo
"T", Sinaloa
Sepúlveda-
Medina, 1991
Zc = Mprom + qprom*fprom c
C = cohort
Paloheimo (1958)
Altamar
(q /temporada):
P. aztecus: 0.0012 -
0.0021
Golfo de México,
Tamaulipas
Castro y
Arreguín-
Sánchez, 1991
Método de Leslie (Ricker, 1975)
Altamar
(q /temporada):
azul: 0.000057 -
0.0000105
Mazatlán,
Sinaloa
Díaz-Corral,
1993
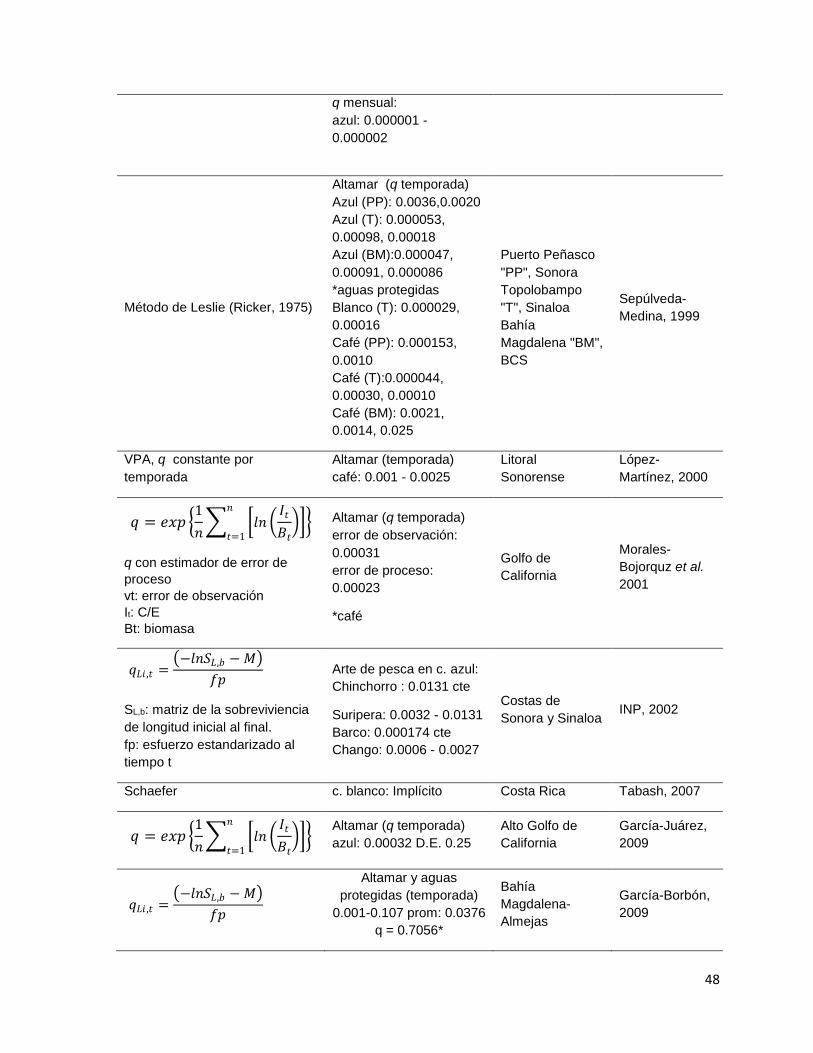
48
q mensual:
azul: 0.000001 -
0.000002
Método de Leslie (Ricker, 1975)
Altamar (q temporada)
Azul (PP): 0.0036,0.0020
Azul (T): 0.000053,
0.00098, 0.00018
Azul (BM):0.000047,
0.00091, 0.000086
*aguas protegidas
Blanco (T): 0.000029,
0.00016
Café (PP): 0.000153,
0.0010
Café (T):0.000044,
0.00030, 0.00010
Café (BM): 0.0021,
0.0014, 0.025
Puerto Peñasco
"PP", Sonora
Topolobampo
"T", Sinaloa
Bahía
Magdalena "BM",
BCS
Sepúlveda-
Medina, 1999
VPA, q constante por
temporada
Altamar (temporada)
café: 0.001 - 0.0025
Litoral
Sonorense
López-
Martínez, 2000
q con estimador de error de
proceso
vt: error de observación
It: C/E
Bt: biomasa
Altamar (q temporada)
error de observación:
0.00031
error de proceso:
0.00023
*café
Golfo de
California
Morales-
Bojorquz et al.
2001
SL,b: matriz de la sobreviviencia
de longitud inicial al final.
fp: esfuerzo estandarizado al
tiempo t
Arte de pesca en c. azul:
Chinchorro : 0.0131 cte
Suripera: 0.0032 - 0.0131
Barco: 0.000174 cte
Chango: 0.0006 - 0.0027
Costas de
Sonora y Sinaloa INP, 2002
Schaefer c. blanco: Implícito Costa Rica Tabash, 2007
Altamar (q temporada)
azul: 0.00032 D.E. 0.25
Alto Golfo de
California
García-Juárez,
2009
Altamar y aguas
protegidas (temporada)
0.001-0.107 prom: 0.0376
q = 0.7056*
Bahía
Magdalena-
Almejas
García-Borbón,
2009
𝑞 = 𝑒𝑥𝑝 1
𝑛 𝑙𝑛
𝐼𝑡
𝐵𝑡
𝑛
𝑡=1
𝑞𝐿𝑖 ,𝑡 =
(−𝑙𝑛𝑆𝐿,𝑏 −𝑀)
𝑓𝑝
𝑞 = 𝑒𝑥𝑝 1
𝑛 𝑙𝑛
𝐼𝑡
𝐵𝑡
𝑛
𝑡=1
𝑞𝐿𝑖 ,𝑡 =
(−𝑙𝑛𝑆𝐿,𝑏 −𝑀)
𝑓𝑝

49
exp (-0.0228 *Lt)
Altamar q(temporada
1999-2005) café:
Schaefer: 0.2357-0.01210
Fox: 0.00769 - 0.00009
Oaxaca
Cervantes-
Hernández et
al., 2010
Z = M + (q*Eprom)
Modelo Paloheimo (1961) c. blanco: Implicito
Sistema lagunar
Huave, Oxaca
Cervantes-
Hernández et
al. 2012
q con estimador de error de
proceso; vt: error de
observación; It: índice
proporcional a la biomasa
Bt: biomasa
Altamar (q/temporada):
café: 0.00034 - 0.00041
café: 0.00021 - 0.00023
Sinaloa y Nayarit Meráz-Sánchez
et al. 2013
Modelo de Schaefer
Flota aguas interiores
qmedio temporada 1987-
2009
L. vannamei: 0.00062
Chiapas Ramos-Cruz,
2013
Las implicaciones de los supuestos de constancia de la capturabilidad son
mencionadas en la tabla 18. Englobando la problemática de considerar una
vulnerabilidad constante en el tiempo, no tomar en cuenta la biología de los recursos
o su comportamiento en función de su edad, y la variabilidad temporal en la abundancia
del recurso por efectos ambientales o biológicos.
𝑞 = 𝑒𝑥𝑝 1
𝑛 𝑙𝑛
𝐼𝑡
𝐵𝑡
𝑛
𝑡=1
𝐶/𝐸 = 𝐼𝑡 = 𝑞𝐵𝑡𝑒𝑣𝑡 ∴
𝑞 = 𝑒𝑥𝑝
1𝑛 𝑙𝑛
𝐼𝑡𝐵𝑡 𝑛
𝑡=1

50
Tabla 18 – Supuesto de capturabilidad (q) más comunes empleados en los modelos
poblacionales y de producción con sus implicaciones (Arreguín-Sánchez, 1996).
Supuesto Implicación
1) q independiente de la abundancia
de la población/biomasa (N)
1) La vulnerabilidad de pesca se vuelve constante ≈ CPUE
independiente de la densidad poblacional. Esto causa una
sobrexplotación rápida del recurso al no tomar en cuenta el
comportamiento intrínseco de cada clase de edad/talla.
2) Se espera que q varíe con N y la vulnerabilidad de manera
similar; pero el comportamiento de algunos stocks es
inverso. Donde en clupeidos la q aumenta cuando disminuye
la abundancia por efecto de la reducción de cardúmenes
formando como estrategia de protección uno solo. Trayendo
como consecuencia principal la sobreexplotación.
2) La abundancia no cambia en el
tiempo, y toda la población se
comporta de manera similar
De manera natural el recurso presenta fluctuaciones en el
tiempo (i.e. interanuales, estacionales), por causas
ambientales, así como cambios en el comportamiento de
acuerdo a su etapa de desarrollo, ambas afectando la
vulnerabilidad del recurso.
2.3.5. Mortalidad por pesca, F
La mortalidad por pesca (F) es el coeficiente de proporcionalidad por muertes
debido a la pesca, representado por un esfuerzo pesquero, con una capturabilidad y
una selectividad en el arte de pesca empleado:
𝐹 = 𝑞𝑠𝐸 (8)
Las estimaciones de la mortalidad por pesca en los modelos pesqueros de
camarón en el Pacífico Mexicano (Tabla 19) se basan en la estimación del parámetro
de capturabilidad constante (generalmente usando método de DeLury o Leslie)
multiplicado por un esfuerzo medido en viajes, barcos o días de pesca. Estas
estimaciones de F normalmente se generan para una temporada de pesca donde F se
mantiene constante; en otros casos se estima una F mensual constante (Sepúlveda-
Medina, 1999); y finalmente existen otras estimaciones de F donde se toma en cuenta
la estructura de la población con F’s diferenciales entre las diferentes

51
edades/longitudes, siendo el caso cuando se emplean los análisis de la población
virtual (López-Martínez, 2000) o cuando se estima una capturabilidad que varíe con la
edad (García-Borbón, 2009). Lo anterior se resume en que dependiendo de la calidad
de la información pesquera disponible es que tan fino es la salida de la mortalidad por
pesca.
De manera global los valores mensuales de F variaron de 1.71 a 0.0002. En
altamar fueron de 1.71 – 0.004 y en aguas interiores 0.89 a 0.0002 para el Pacífico
Mexicano (tabla 19).
Tabla 19 – Valores de mortalidad por pesca (F) para la pesquería de camarón en el
Pacífico Mexicano.
Especie Ecuación Valores (F) mensuales
Flota Sitio Periodo Referencia
L. stylirostris
0.17 Altamar Sonora Sur 1970-1973
Rodríguez de la Cruz, 1976
L.vannamei
0.06 - 0.36 Aguas interiores
Laguna Huizache y Caimanero, Sinaloa
1972 - 1975
Sepúlveda-Medina, 1976
L.vannamei
0.016 - 0.25
Altamar Golfo de California
1970-75 Jacquemin, 1976
L. stylirostris 0.018 - 0.077
F. californiensis 0.020 - .029
L. stylirostris
0.07 - 0.10
Altamar Puerto Peñasco, Sonora
1968-1972
García-Gómez, 1976 F. californiensis 0.04 - 0.06
L.vannamei
0.052 - 0.212 Aguas interiores
Caimanero, Sinaloa 1976-
1977
Sepúlveda-Medina, 1981 0.1 - 0.64
Huizache, Sinaloa
L. stylirostris
0.539 - 0.238
Altamar Norte y Centro del GC
1976-1985
Ross-Terrazas, 1988 F. californiensis 0.541 - 0.065
F. californiensis VPA 0.004 - 0.08 Altamar Sonora 1978-1995
López-Martínez, 2000
F. californiensis
0.2 - 0.31 Altamar
Puerto Peñasco, Sonora
1987-1994 Sepúlveda-
Medina, 1999
L. stylirostris 0.3 - 0.86
F. californiensis 0.3 - 0.53 Altamar Topolobampo, Sinaloa
1987-1994
𝑍 = 𝑀 + 𝐹
𝑍 = 𝐿𝑛
(𝑁𝑡)
𝑁0
𝑍 = 𝑀 + 𝐹
𝑍 = 𝐿𝑛
(𝑁𝑡)
𝑁0
𝑍 = 𝐿𝑛
(𝑁𝑡)
𝑁0
𝑍 = 𝐿𝑛
(𝑁𝑡)
𝑁0
𝑍 = 𝐿𝑜𝑔 𝑒𝑥𝑝 𝑁𝑡
𝑁0

52
L. stylirostris 0.19 - 1.71
L.vannamei 0.081 - 0.28
F. californiensis 0.32 - 1.5 Altamar costa occidental, BCS
L. stylirostris 0.18 - 0.31 Aguas interiores
1987-1994
F. californiensis VPA
0.18-1.61 Altamar Tehuantepec
1989-1998
Ramos-Cruz, et al., 2006 L. vannamei 0.02 - 0.79
F. californiensis
0.06 - 0.15 Altamar costa occidental, BCS
1987-1994
García-Borbón, 2007
F. californiensis
sin vulnerabilidad 0.0745 - 0.002 prom: 0.016 con vulnerabilidad 0.89 - 0.0002 prom: 0.068
Aguas interiores
Bahía Magdalena
1991-2007
García-Borbón, 2009.
𝐹𝑎 ,𝑡 = −𝑙𝑛 𝑁𝑎+1,𝑡+1
𝑁𝑎 ,𝑡 −𝑀
𝐹𝑎 ,𝑡 = 𝑠𝑎𝑞 𝑎 ,𝑡𝐸𝑡

53
2.3. Normatividad
La pesquería de camarón en México está regida por la Ley General de Pesca y
Acuacultura Sustentables (2015) y su correspondiente reglamento que es una ley
reglamentaria del Artículo 27 de la Constitución General de los Estados Unidos
Mexicanos. En ella se establecen las condiciones de acceso al recurso así como los
derechos y obligaciones de los usuarios y/o armadores. Otras leyes concurrentes son:
a) Ley de Metrología y Normalización (2009); b) Ley General de Sociedades
Cooperativas (2009); y c) Ley General del Equilibrio Ecológico y Protección al
Ambiente (LGEEPA, 2014).
Como instrumentos normativos de la pesquería de camarón se encuentran a) la
Norma Oficial Mexicana NOM-002-SAG/PESC-2013 y sus modificaciones, con el
objetivo de ordenar el aprovechamiento de las especies de camarón en aguas de
jurisdicción federal de los Estados Unidos Mexicanos; b) la Carta Nacional Pesquera
(2012) que es un instrumento jurídico normativo donde contiene el estado que guardan
las poblaciones y sus pesquerías, así como los lineamientos y estrategias de manejo
por región de las pesquerías en México; y c) el Plan de Manejo para la Pesquería de
Camarón en el Litoral del Océano Pacífico Mexicano del 2004 con énfasis en la
problemática junto con los objetivos y estrategias de manejo de la pesquería. A manera
de resumen se incluye las principales características de la normatividad en la pesca
de camarón (tabla 20).

54
Tabla 20 - Normatividad en la pesquería de camarón del Pacífico Mexicano en la NOM-
002-SAG/PESC-20131, Carta Nacional Pesquera (2012)2, y Plan de Manejo para la
Pesquería de Camarón en el Litoral del Océano Pacífico Mexicano del 20043.
Unidades de pesca Normatividad
a) Embarcación
mayor
Características: capacidad de bodega >10 ton métricas,
>10.5m de eslora, motor > 80 HP y autonomía hasta por
50 días, equipo de refrigeración, equipo de navegación
y exploración, operado hasta por seis personas1,2,3.
Dónde: Aguas marinas de jurisdicción federal, solo
operan entre los 18 y los 108m≈10 - 60 brazas
(excepción: embarcaciones menores en zona de
amortiguamiento de la Reserva de la Biosfera del Alto
Golfo de California y Delta del Río Colorado con red de
arrastre RS-INP-MEX y excluidor de peces y tortugas).
***Restricción en radio de 9,25 km≈ 5 millas náuticas
alrededor de bocas de bahías, lagunas y esteros del
Pacífico Mexicano1,2,3.
*Obligatorio: Permiso de pesca; Bitácora de pesca;
aviso de Arribo en la Oficina Federal hasta 72 hrs
después de arribar en puerto; sistema de localización y
monitoreo satelital (desde 2008); dispositivo excluidor
de tortugas (DET) y de peces (DEP) para reducir la
fauna de acompañamiento (FAC) 1,2,3.
a.1) Artes de pesca Dos redes de arrastre2: luz de malla de alas cielo,
cuerpo y ante bolso no > 50.8mm ≈2 pulgadas y bolso
de 38.1mm ≈1 ½ pulgadas; relija superior no > 36.6m
≈120 pies. Formas de red de arrastre más usadas en el
Pacífico: balón, semibalón, fantasma, hawaiana y la
voladora3.
b) Embarcación
menor
Solo con embarcaciones menores de <10m de eslora
con motores fuera de ≤ 110 HP1 ó 55-150 HP2,
capacidad de 0.5 – 3.5 tons, de 2 a 3 tripulantes; existen

55
cayucos de madera con remo de 5m con 2 tripulantes
en el sur del Golfo de California incluyendo Sinaloa3.
Dónde: aguas de jurisdicción federal dentro de bahías,
marismas y sistemas lagunarios-estuarinos.
*Obligatorio: Permiso de pesca; Bitácora de pesca;
aviso de Arribo en la Oficina Federal hasta 72 hrs
después de arribar en puerto 1,2,3.
b.1) Artes de pesca a) Atarraya con luz de malla de 37.5mm ≈ 1 ½ pulgadas
en todas las áreas1,3.
b) 1 red “suripera”: luz de malla 31.75mm ≈1¼
pulgadas; para embarcaciones ≤ 7.6m (25 pies); solo en
lagunas, estuarios y bahías del centro y norte de
Sinaloa1,3.
c) 1 red de enmalle o “Chinchorro de línea”: luz de malla
mínimo de 63.5mm ≈ 2 ½ pulgadas y 200m de longitud;
solo aguas marinas costeras y lagunas-estuarios-bahías
de Sonora; en la zona de amortiguamiento de la
Reserva de la Biosfera del Alto Golfo de California y
Delta del Río Colorado; en sistemas lagunarios-
estuarinos y bahías del norte de Sinaloa; Restricción de
uso: distancia menor a los 2 km por ambos lados de las
bocas de lagunas, esteros y ríos y hasta los 9.14m1,3.
d) Tapos3.
e) Red de arrastre tipo chango3.
c) Especies objetivo
y Fauna de
Acompañamiento del
Camarón (FAC)
Se capturan cinco especies objetivo: Litopenaeus
stylirostris (camarón azul), L. vannamei (c. blanco), L.
occidentalis (blanco del sur), Farfantepenaeus
californiensis (c. café) y F. brevirostris (c. cristal). La
composición de la especies varía por zona de captura y
como parte de la fauna de acompañamiento del
camarón se capturan 600 especies: peces, moluscos,
equinodermos y crustáceos, y cuatro se incluyen en la
NOM-059-SEMARNAT-2010 (Protección a especies
nativas de México)2.

56
e) Estatus - Pesquería aprovechada al máximo sustentable, las
capturas varían por zona y especie, y por condiciones
ambientales2.
- Esfuerzo pesquero al límite sustentable2.
j) Extracción para
acuacultura
a) Prohibida la extracción de larvas y postlarvas, y
reproductores (en época de veda) excepto para
investigación con previo Dictamen del INAPESCA1.
b) La vigilancia del cumplimiento de la NOM:
CONAPESCA, SEMARNAT, PROFEPA Y SEMAR1.

57
2.4. Ordenación Pesquera
2.4.1. Ordenación y manejo pesquero a través de modelos pesqueros en
camarón
De acuerdo a la FAO (1999) el proceso de ordenación pesquero es “el proceso
integrado de recogida de información, análisis, planificación, consulta, adopción de
decisiones, asignación de recursos y, formulación y ejecución, así como la imposición
de reglamentos o normas que rijan las actividades pesqueras para asegurar la
productividad pesquera de los recursos (i.e. uso sustentable) y la consecución de otros
objetivos”.
De acuerdo a Seijo et al., (1997) las estrategias de ordenación en las pesquerías
por parte de las autoridades competentes deben estar definidas en objetivos
sustentados en uno o varios criterios de bienestar socioeconómicos dependiendo de
cada caso. Los objetivos más comunes de ordenación en las pesquerías son a) de
conservación, b) económico y c) equidad de recurso (tabla 21).
Tabla 21– Objetivos de ordenación en las pesquerías.
Objetivos Características
Conservación
Objetivo: sustentabilidad del recurso regulando el esfuerzo
pesquero mejorando su abundancia en el tiempo.
Ventaja: conservación del capital natural.
Desventajas: 1) falta de información para las evaluaciones;
2) proyectar cada recurso; implicaciones sociales y
económicas importantes al aplicar medidas de regulación.
Económico
Objetivo: incrementar la renta económica.
Ventaja: tendencia a maximizar la renta económica entre
los participantes.
Desventajas: 1) sobrecapitalización; 2) no toma en cuenta
la biología del recurso; 3) En algunos casos el alto precio
del recurso compensa los altos costos de producción; 3)
No considera factores ambientales y el equilibrio
“determinístico” no existe.

58
Equidad del
recurso
Objetivo: igualdad de oportunidades entre los usuarios de
la pesquería.
Ventajas: bienestar social.
Desventajas: 1) acceso abierto y su efecto sobre los
usuarios dependerá de su capacidad de adaptación; 2)
descontento social; 3) no garantiza una distribución
equitativa de los recursos naturales.
El postulado fundamental de la teoría del bienestar económico aplicado a
pesquerías (Hannesson, 1978) establece que la maximización del bienestar social
(llámese utilidad o renta de la pesquería) es la finalidad de la política pesquera. El
concepto básico de esta teoría es el de Pareto optimalidad, esto significa que una
pesquería está en un estado Pareto óptimo si los cambios en los instrumentos de
manejo mejoran el bienestar de los usuarios sin empeorar el estado de los demás.
Las condiciones de Pareto optimalidad en las pesquerías de acuerdo a Seijo et
al., (1997) son: a) que no aumente la captura y la renta económica de algunas especies
(s) y afecten otras que estén ecológica o tecnológicamente interdependientes; y 2) que
no se redistribuya las capturas entre los consumidores de tal forma que el bienestar
de un consumidor mejore y el de otro disminuya.
En la teoría neoclásica del bienestar aplicada a los recursos naturales
(Bergstrom y Randall, 2016) se plantean diferentes criterios de bienestar cada uno con
su particular manera de distribuir los ingresos/recursos en la sociedad (tabla 22) y esto
depende de lo que la sociedad permite o requiere para alcanzar la eficiencia
económica a veces permitiendo el daño económico, definido como la reducción de la
utilidad individual como resultado directo o indirecto de una decisión de política pública.
Finalmente los criterios de la tabla 22 pueden combinarse para diseñar
estrategias de manejo para lograr los objetivos que cada manejador define en su
pesquería. Estos criterios de manejo empleados por las instituciones responsables de
administrar los recursos marinos deberán hacer explícitos los juicios de valor
distribucionales inherentes en cada uno de ellos.

59
Las herramientas que usa la ordenación pesquera para alcanzar sus objetivos
(tabla 21) basados en algún criterio de bienestar social (tabla 22) se basan en
estrategias de manejo que regulan las variables involucradas en las pesquerías. Estas
se llaman variables control (sobre las que el manejador controla) y las variables estado
(son la variables descriptoras de un sistema dinámico).
2.4.2. Estrategias de manejo pesquero globales
De manera general existen cuatro tácticas no excluyentes de implementar
tácticas de manejo en una pesquería indicadas en la figura 10 (Anderson, 1977;
Pearse, 1980; Seijo, 1986).

60
Tabla 22– Criterios de bienestar aplicados a pesquerías marinas (Seijo et al., 1997; Bergstrom y Randall, 2016).
Criterio de
bienestar
Característica
distribucional Impacto Observaciones
Pareto eficiencia o
eficiencia
económica
Permite daño
económico
Neutral para el
pescador
menos eficiente
Selección de varias soluciones Pareto eficientes con diferentes distribuciones
de los ingresos entre los usuarios; la eficiencia económica (i.e. soluciones
Pareto eficientes) se localiza en la frontera de la gran utilidad (FGU).
Pareto seguridad No permite daño
económico
Neutral
absoluto entre
pescadores
Este criterio solo busca las soluciones que mejoren (i.e. Pareto mejora) ó
aumenten el ingreso o capital natural de al menos una persona sin reducir el
ingreso de nadie.
Son de las políticas óptimas para alcanzar la eficiencia económica.
El Pareto seguridad protege a los individuos de empeorar por políticas
impuestas por el gobierno.
Valor máximo del
producto social
Permite daño
económico si la
sumatoria del
bienestar es máxima
y exceda a los
costos y sea >0
Neutral
Este criterio se calcula sin el peso distribucional entre los usuarios (curvas de
indiferencia con pendiente -1) y la solución resultante será el máximo valor
del bienestar social.
La solución Pareto óptima es la tangente entre la FGU y la curva de
indiferencia más cercana llamado “punto de la dicha o felicidad”.
Este criterio es totalmente neutro en la redistribución del ingreso
Partes
proporcionales
constantes
No permite el daño
absoluto ni relativo
entre pescadores
Proporcional
En este criterio la mejoría es un aumento en el ingreso para alguien mientras
el de todos aumente proporcionalmente a partir de su ingreso inicial.
No hay daño económico porque ninguna parte empeora.
Máximo bienestar
social
Permite daño
económico Neutral
Este criterio es problemático por la dificultad en llegar a un consenso
respecto a lo que es el bienestar social y establecerlo en una función del
beneficio social.
Se puede evaluar este criterio estableciendo lo que la sociedad requiere y
permite (daño económico) para alcanzar la eficiencia económica. La solución
óptima es la tangente entre la curva de indiferencia y la FGU. Se permite el
daño económico mientras exista consenso social que la solución óptima
mejora el bienestar social

61
Figura 10– Tácticas de manejo pesquero empleadas en el manejo de las pesquerías
en el mundo.

62
2.4.3. Estrategias de manejo aplicadas en la pesquería de camarón
A nivel mundial los objetivos generales de la ordenación de la pesquería del
camarón se basan en la conservación del recurso para maximizar la producción y
disminuir los costos globales para mejorar la situación socioeconómica redistribuyendo
las utilidades a la sociedad (García y Le Reste, 1997). El otro objetivo es el económico,
donde se pretende aumentar la renta de los usuarios al disminuir la mortalidad por
pesca de la flota de pequeña escala, además de disminuir el descarte y aprovechar lo
mejor posible las especies secundarias.
En la práctica, las políticas de manejo serán definidas para cada caso particular
en base a la información y los objetivos inmediatos, teniendo en cuenta que los datos,
suficientes y precisos, rara vez están disponibles. De acuerdo a García y Le Reste
(1997) los principios de su ordenación están basados en los puntos de referencia de
máximo rendimiento sostenible (estado capacidad de carga de biomasa virgen “K”/2),
máximo rendimiento económico (punto donde la diferencia entre los ingresos
marginales y costos marginales es máxima) y equilibrio bioeconómico (punto donde
los ingresos marginales son iguales a los costos marginales). Las principales
estrategias de regulación pesquera para alcanzar estos puntos de referencia en
general son dos: control sobre el recurso y control sobre el esfuerzo (figura 11).

63
Figura 11 - Estrategias de manejo en la pesquería de camarón.

64
Gulland y Rothschild (1984) menciona cinco problemas globales inherentes en
el manejo pesquero del camarón que aplican en las pesquerías mexicanas : (1) no se
toma en cuenta el sistema pesquero completo; (2) al ser una pesca multiespecífica no
se da seguimiento a cada especie; (3) existen múltiples artes de pesca involucrados y
no hay una estandarización del esfuerzo; (4) conflictos por operaciones entre las flotas
participantes; (5) se debe tener un monitoreo en tiempo real de la captura y esfuerzo
por localidad, así como del estado de las poblaciones. A lo que agrega que los
principales signos de depleción en la pesquería de camarón son: disminución
constante de las capturas en varios años, la reducción de las tallas de captura, y una
proporción de sexos no balanceada. La ventaja en especies de ciclo de vida corto
anual y muy fecundas, como el camarón, es la recuperación de los stocks en poco
tiempo y alcanzar la estabilidad productiva en pocos años (Lluch, 1977).
2.4.4. Manejo pesquero del camarón en México
La pesquería de camarón en México se basa en su manejo en seis instrumentos
observados en la figura 12. Dentro de estos en el “Plan de manejo para la pesquería
de camarón en el litoral del océano Pacífico mexicano” se plantean seis objetivos (fig.
13) de manejo con sus objetivos particulares (CONAPESCA, 2004). El punto de
referencia objetivo (PRO) de la pesquería mexicana es el máximo rendimiento
sostenible (MRS) usando como variable estado la abundancia relativa y la biomasa de
la población, como variable control el esfuerzo pesquero, y bajo el supuesto de una
tasa de explotación constante al 50% de la biomasa virgen (fig. 14). Las estrategias de
manejo para conseguir el PRO que se plantean son vedas, restricción de áreas de
pesca entre flotas, magnitud del esfuerzo, tasas de explotación óptimas, y usar
modelos que involucren el análisis de vulnerabilidad de tallas como son los modelos
rendimiento por recluta y estructurado por edades. Con el primer modelo las
autoridades pesqueras establecen el punto de referencia límite (PRL) que es la
mortalidad por pesca que genera la máxima producción (FMAX). Existen unas acciones
emergentes del manejo para apoyar la consecución de PRO y PRL que se basan en
la regulación del esfuerzo pesquero basados en evaluaciones periódicas durante la
temporada de pesca (fig. 14).

65
Figura 12– Instrumentos de manejo para la pesquería de camarón (CONAPESCA, 2004).

66
Figura 13- Objetivos de manejo para la pesquería de camarón en el Pacífico mexicano (CONAPESCA, 2004).

67
Figura 14 - Puntos de referencia en manejo de la pesquería de camarón en el Pacífico mexicano.

68
Todos estos objetivos de manejo deben ser analizados por medio de modelos
pesqueros que puedan ofrecer simulaciones en la aplicación de las estrategias de
manejo para obtener aproximaciones del efecto en su implementación sobre la
pesquería y pueda escogerse la estrategia óptima basada en los objetivos de
ordenación y en un criterio de bienestar social (tablas 21 y 22).

69
2.5. Modelos bioeconómicos empleados en las pesquerías de camarón
Las evaluaciones de los recursos pesqueros se basan en la modelación de las
pesquerías en función de la calidad de datos disponibles. Estos modelos fueron
elaborados para especies de ciclos de vida largos como los peces, y no fueron
diseñados para evaluar especies de ciclos cortos como los peneidos (García y Le
Reste, 1997). En función de la calidad de datos existen modelos globales o de
producción, modelos analíticos y métodos aproximativos (García y Le Reste, 1997).
El modelo más sencillo que utiliza la mínima información pesquera es el global
o de producción (Schaefer, 1954 y derivados, Fox, 1970; Pella y Tomlison, 1969).
Debido a esta característica es de los más usados en las pesquerías (Anderson y Seijo,
2010) incluida las pesquerías de los peneidos, aun cuando sus supuestos son
inaplicables en su biología y dinámica poblacional: 1) toda la variación de la biomasa
o población se le adjudica al esfuerzo pesquero, siendo que en los peneidos la
variabilidad ambiental juega un papel muy importante y regulatorio en sus poblaciones
(ver sección 2.1.4.); 2) se asume condiciones de equilibro ambiental las cuales no
existen por la variabilidad tan alta que presentan la biomasa de los peneidos
anualmente; 3) se asume que se explota una sola población sin tomar en cuenta
interacciones con otras especies, cuando la pesquería es multiespecífica y las
especies están estructuradas en subpoblaciones dentro del Golfo de Californian
(Aubert y Lightner, 2000 para L. stylirostris ; Valles-Jiménez et al., 2004 para L.
vannamei); 4) se asume estabilidad poblacional donde la producción y/o captura son
independientes a las edades, o en otras palabras una misma vulnerabilidad para toda
la estructura de edades. Sin embargo la población de peneidos consiste en varias
clases de edad (varias cohortes) que presenta una naturaleza extremadamente
dinámica observada en la gran variabilidad en el tiempo de la biomasa, edad media de
captura, fertilidad, etc. Además la naturaleza secuencial en la pesquería de camarón
ofrece dificultades en el sentido que la sobrexplotación de un componente juvenil o
adulto afectará la producción del otro.
Los modelos de producción son los más comunes en peneidos, aun con la
violación de los supuestos, y siempre buscando alcanzar como punto objetivo el
Máximo Rendimiento Sustentable (MSY), el cual al basarse en los supuestos del

70
modelo tampoco va acorde con la biología de los peneidos (Garcia y LeReste, 1997).
En la tabla 23 se puede observar algunos modelos propuestos en la pesquería de
camarón dentro del Golfo de California donde la mayoría son basados en modelos de
producción, sobre la especie camarón café y con la flota de altamar.
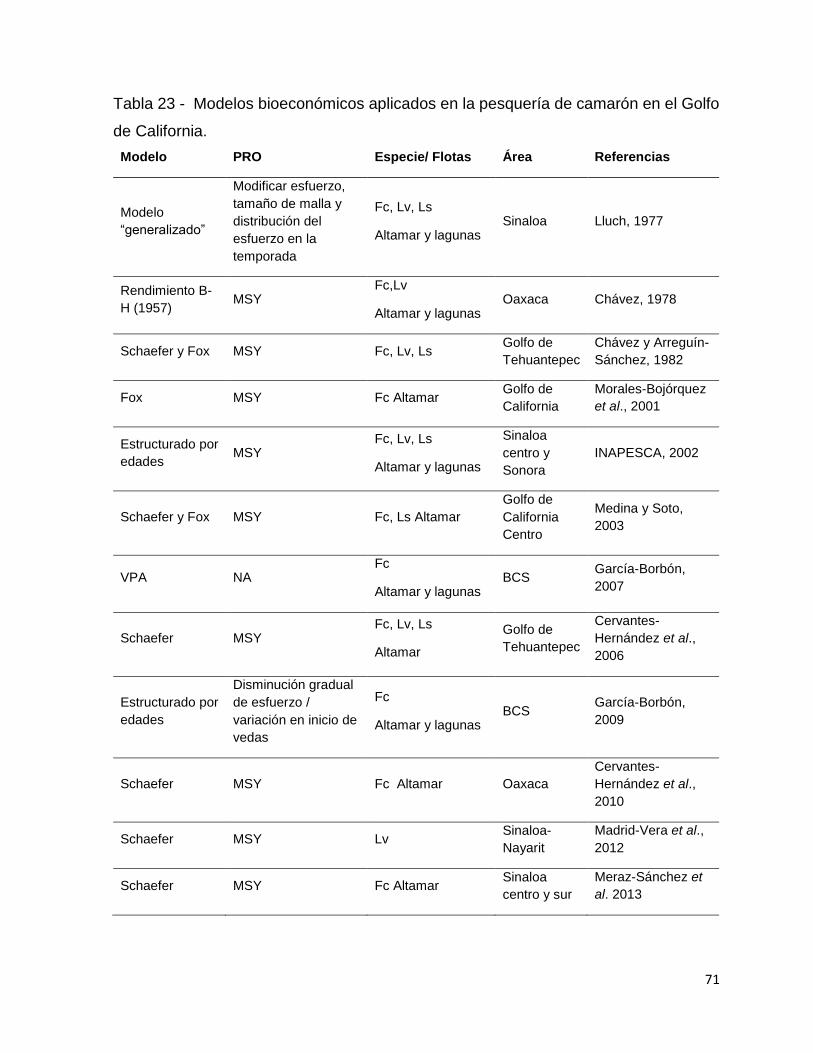
71
Tabla 23 - Modelos bioeconómicos aplicados en la pesquería de camarón en el Golfo
de California.
Modelo PRO Especie/ Flotas Área Referencias
Modelo
“generalizado”
Modificar esfuerzo,
tamaño de malla y
distribución del
esfuerzo en la
temporada
Fc, Lv, Ls
Altamar y lagunas Sinaloa Lluch, 1977
Rendimiento B-
H (1957) MSY
Fc,Lv
Altamar y lagunas Oaxaca Chávez, 1978
Schaefer y Fox MSY Fc, Lv, Ls Golfo de
Tehuantepec
Chávez y Arreguín-
Sánchez, 1982
Fox MSY Fc Altamar Golfo de
California
Morales-Bojórquez
et al., 2001
Estructurado por
edades MSY
Fc, Lv, Ls
Altamar y lagunas
Sinaloa
centro y
Sonora
INAPESCA, 2002
Schaefer y Fox MSY Fc, Ls Altamar
Golfo de
California
Centro
Medina y Soto,
2003
VPA NA Fc
Altamar y lagunas BCS
García-Borbón,
2007
Schaefer MSY Fc, Lv, Ls
Altamar
Golfo de
Tehuantepec
Cervantes-
Hernández et al.,
2006
Estructurado por
edades
Disminución gradual
de esfuerzo /
variación en inicio de
vedas
Fc
Altamar y lagunas BCS
García-Borbón,
2009
Schaefer MSY Fc Altamar Oaxaca
Cervantes-
Hernández et al.,
2010
Schaefer MSY Lv Sinaloa-
Nayarit
Madrid-Vera et al.,
2012
Schaefer MSY Fc Altamar Sinaloa
centro y sur
Meraz-Sánchez et
al. 2013

72
Schaefer MSY Ls Altamar Alto Golfo de
California
García-Juárez et
al., 2009; García-
Juárez et al., 2014
*PRO: Punto de referencia objetivo del manejo propuesto; VPA: análisis de población virtual;
MSY: Rendimiento Máximo Sostenible; Fc: Farfantepenaeus californiensis; Ls: Litopenaeus stylirostris
Lv: Litopenaeus vannamei.
Dentro de los supuestos técnicos pesqueros en los modelos de producción que
tampoco aplican en peneidos (Garcia y Le reste, 1987) se encuentra: 1) la aplicación
de una capturabilidad contante (todas la estructura de tallas en la población tiene la
misma vulnerabilidad); 2) una mortalidad natural constante (todas las edades y fases
del camarón tienen la misma probabilidad de muerte por causas naturales); y 3) la
población reacciona inmediatamente al cambio en el esfuerzo pesquero (en peneidos
al ser de ciclo de vida corto se asume que no hay tiempo de retraso y las poblaciones
reaccionan relativamente rápido). Con todas la evidencias anteriores se afirma que
este modelo no es el óptimo para el manejo de camarones porque no permite integrar
los datos de juveniles con los de adultos y tampoco permite predecir las consecuencias
variando la tasa de mortalidad por pesca en función de la edad, en escenarios
aplicando una veda o en regulaciones de la luz de malla de las redes (García y Le
Reste, 1997).
Dentro de los modelos analíticos se emplean el modelo de rendimiento por
recluta y las técnicas de simulación. El objetivo general del primer modelo es obtener
la mortalidad por pesca (F) que genere la máxima producción de la pesquería llamado
FMAX. Para el primero modelo los más usados son Beverton-Holt (1957), Ricker (1958)
≈ Thompson y Bell (1934) y en Ricker, (1975) donde el ciclo vital se divide en periodos
cortos de tiempo y los parámetros se toman como constantes. Los supuestos son
mortalidad natural constante, un reclutamiento constante y vulnerabilidad constante
sobre la estructura de edades y existen condiciones de equilibrio. García y Le Reste
(1997) menciona que si se cuenta con la información pesquera suficiente el modelo
puede predecir consecuencias del uso de distintas regulaciones pesqueras. Sin
embargo de acuerdo a el mismo autor es errático cuando existen variaciones
estacionales (crecimiento, reclutamiento y capturabilidad) y para prever consecuencias

73
precisas de la distribución estacional esfuerzo como son la aplicación de vedas sobre
el vector de F promedio en función de la edad.
Dentro de las técnicas de simulación se encuentra el modelo de estructura por
edades. Este tiene una gran aplicabilidad de análisis pesquero y su único limitante es
la calidad y disponibilidad de los datos. Los modelos de estructura por edades
consideran factores como el crecimiento, reclutamiento, mortalidad natural y
capturabilidad que afectan la biomasa de la población ó cohorte, los cuales son
tomados como implícitos dentro de los modelos globales. Estos modelos pueden
representar la dinámica poblacional de los peneidos con excelentes resultados
(Anderson y Seijo, 2010).
Los primeros pioneros en el modelo estructurado por edades en su forma
estática fueron Beverton y Holt (1957) definiendo a la captura como el producto de F
y la biomasa de una población, esta es producto de un peso individual por el número
de individuos presentes en el tiempo. El modelo dinámico de estructura de edades ya
toma en cuenta los influjos y eflujos para cada cohorte de la estructura de la población
aplicando los principales conceptos de tablas de vida i.e. procesos de crecimiento,
reclutamiento y mortalidad (Begon et al., 1990). La variable estado del modelo es un
vector del número de individuos en cada clase de edad y la captura se compone de un
número de individuos de diferentes clases de edad tomadas en cualquier año (Anderso
y Seijo, 2010).
Los supuestos que manejan son (Seijo et al., 1997): 1) el reclutamiento y la
selección del arte de pesca presentan una forma de filo de navaja; 2) toda la cohorte
se captura al mismo tiempo; 3) distribución homogénea y misma probabilidad de
captura para todos los individuos; 4) la tasa de explotación es constante durante la
vida de los organismos, donde el rendimiento será estable en el tiempo si el patrón de
captura se mantiene y hay reclutamiento constante; 5) mortalidad natural constante e
independiente de la edad; 6) tradicionalmente el reclutamiento era considerado
independiente al tamaño del stock parental y no se afecta por la variación de la
intensidad de pesca, este supuesto es el más irreal de todos (Seijo et al., 1997), pero
después se incorporaron los modelos de reclutamiento denso independientes y

74
dependientes de Beverton y Holt (1957) y Ricker (1975), siendo el primero de los más
usados en peneidos (Ramírez y Arreguín, 2003; Chen et al., 2007).
De acuerdo a Seijo et al., (1997) y Anderson y Seijo (2010) los modelos de
estructura por edades permiten observar consecuencias en la aplicación de
regulaciones como el establecimiento de una edad de primera captura y/o escenarios
de variabilidad en las relaciones reclutamiento y stock parental variando el esfuerzo
pesquero. Se puede estimar el tamaño del stock y los niveles de capturas usando
diferentes niveles de pesca, llamado proyección del stock. Si se aplicara al revés se
puede calcular las tasas de F para llegar a un nivel de captura deseado, observando
como el nivel de captura afecta el tamaño del stock y el reclutamiento. Así mismo el
modelo se puede emplear para a calcular las tasas de F para alcanzar un objetivo de
recuperación de stocks, p.e. “alcanzar un +25% en el stock desovante en un periodo
de 10 años”.
En México existen pocos ejemplos que apliquen el modelo estructurado por
edades para el manejo de la pesquería de camarón (ver tabla 23, INAPESCA, 2003;
García-Borbón, 2007,2009).

75
3. Justificación
Como se observó en la sección anterior se necesita generar un modelo
adecuado a la biología en peneidos y además que use otro punto de referencia objetivo
que no se base en los principios de los modelos de producción. Más aún que tome en
cuenta a todas las especies comerciales y que incluya la fase lagunar y marina para
tener así todo el sistema de la pesquería de camarón. Todo lo anterior depende
únicamente en la calidad de la información que existe para generar un modelo con
esta complejidad.
Para generar un modelo estructurado por edades multiespecies, multiflota con
parámetros dinámicos en el tiempo y espacio y además con reclutamiento estacional
ocupamos analizar toda la información biológica-pesquera y obtener los parámetros
relevantes para la zona de estudio (para este caso: Sinaloa sur). Por lo que la primer
parte del trabajo es analizar los parámetros de crecimiento, obtener la clave edad-
longitud, la mortalidad natural por etapa de vida y el reclutamiento estacional por cada
especie, obteniendo la dinámica poblacional por especie. Aquí se debe recalcar que
debido a que la pesquería es de tipo secuencial, distintas flotas capturan distintos
componentes del ciclo de vida en peneidos, esto implica mortalidades naturales
diferenciales entre sus distintos estadios de vida, hábitat, ecología, etc, y esto no se
aplica en la gran mayoría de los trabajos de manejo pesquero en México al usar
mortalidades naturales constantes y para adultos. Otro punto importante es el
reclutamiento, la innovación es que se ajustará una función al reclutamiento para
simular la estacionalidad por especie lo cual no existe en otro modelo pesquero en
México. Además, basados en efectos ambientales conocidos sobre el reclutamiento
por especie, se puede explorar escenarios ambientales con distintos reclutamientos,
lo cual tampoco ha sido explorado bajo la perspectiva bioeconómica.
La parte pesquera y económica es la más empobrecida en calidad de
información y este trabajo pretende por primera vez obtener los datos necesarios para
ser empleados en modelos pesqueros complejos (no de producción excedente). Se
buscará obtener los esfuerzos pesqueros más realistas tanto de la flota industrial y
sobre todo generar un dato de esfuerzo ribereño confiable (para la zona) que nunca
se maneja en la literatura porque no existe. Además se debe obtener la captura

76
estructurada por tallas/edades por flota por especie en al menos una temporada. Con
la combinación de esfuerzo y captura por edad se puede tener un análisis más fino de
la capturabilidad a cada talla/edad y romper con la tendencia del uso de
capturabilidades constantes las cuales no aplican en peneidos y se podrá observar
tallas vulnerables durante la temporada de pesca.
Los modelos pesqueros que analizan la pesquería de peneidos con el modelo
estructurado por edades son tres no publicados (reporte del INAPESCA, tesis García
Borbón 2007, 2009) pero el primero sigue usando capturabilidad constante en la parte
marina bajo el PRO de MSY, y los segundos usan solo una especie y usan como
manejo escenarios de % de esfuerzo e inicio de veda. El modelo propone el manejo
de Máximo Rendimiento Económico, MEY, el cual es el recomendado por las máximas
autoridades en pesquerías de camarón (García y LeReste, 1997) y además usan
parámetros pesqueros dinámicos que superan los modelos actuales en México.
Por último en lo relacionado al manejo pesquero tomando en cuenta la
variabilidad climática y las variaciones del reclutamiento no existe mucha información
en México del tema y esta es nula en camarones. Es muy importante estudiar la
manera como la pesquería puede adaptarse a los cambios ambientales que afectan el
reclutamiento-producción y poder abordar que estrategia de manejo podrían aplicarse
para contrarrestar estos efectos para mantener el bienestar socio-económico.

77
4. Objetivos
4.1. Objetivo general
Desarrollar un modelo bioeconómico estructurado por edades, multi-especies,
multi-flotas, con mortalidad natural y capturabilidad dinámicos a la edad y tiempo, y
con una función de reclutamiento estacional por cada especie para la pesquería del
camarón en la temporada 2014-15 en el sur del Golfo de California.
4.2. Objetivo particular
a) Estimar los vectores de mortalidad natural para cada intervalo de edad en las
especies comerciales L. vannamei, L, stylirostris y Farfantepenaeus californiensis
del sur de Sinaloa, México.
b) Elaborar la clave edad-longitud para las especies Litopenaeus stylirostris (azul) y L.
vannamei (blanco) para Sinaloa sur.
c) Determinar las variables de esfuerzo por cada flota así como la estructura de edades
de su captura correspondiente.
d) Determinar los vectores de capturabilidad a la edad (𝑞𝑖) para las flotas que explotan
el camarón azul (L. stylirostris), blanco (L. vannamei) y café (F. californiensis) en la
zona de Sinaloa sur para la temporada de pesca 2014-2015.
e) Crear un modelo bioeconómico con parámetros dinámicos de coeficiente de
capturabilidad y mortalidad natural por edad y por especie que describa la
temporada 2014-15 para la pesquería del camarón en Sinaloa sur.
f) Establecer un modelo de reclutamiento estacional mensual por especie que permita
manipular el reclutamiento en función de su repuesta ante el ambiente y que varíe
con el esfuerzo pesquero.
g) Establecer el manejo óptimo de la temporada 2014-15 usando el punto de referencia
objetivo de máximo rendimiento económico (MEY) sobre la flota de altamar.
h) Generar escenarios de variabilidad interanual del reclutamiento en función de las
condiciones ambientales para establecer el manejo óptimo a la pesquería de
camarón usando el punto de referencia objetivo de máximo rendimiento económico
(MEY) sobre la flota de altamar como medida de resiliencia pesquera ante la
variabilidad climática.

78
5. Materiales y métodos
La estrategia de investigación será abordada primero resolviendo las cuestiones
de la dinámica poblacional, después resolviendo los parámetros pesqueros y terminar
con la construcción del modelo bioeconómico estructurado por edades.
5.1. Parámetros de la dinámica poblacional
5.1.1. Crecimiento para las especies de camarón Litopenaeus stylirostris, L.
vannamei y Farfantepenaeus californiensis.
Se construyó una base de datos a partir de la literatura para encontrar las
estimaciones de las ecuaciones de crecimiento de von Bertalanffy (1934) por cada
especie (tabla 8). Después se evaluaron las relaciones peso-longitud de cada modelos
y se validaron con valores biométricos observados para la zona de Sinaloa sur en las
especies L. vannamei y L. stylirostris, no hubo muestra biológica para F. californiensis.
Para lo anterior se realizó un solo muestreo biométrico (octubre – noviembre,
2015) tomando los datos de longitud y peso total en las lagunas de Huizache (n=269),
Chametla (n=588), Teacapán (n=590), Palmilla (n=417) y en la zona de costera de
Teacapán (n=195) (fig 15). Y se completaron con otras medidas biométricas
independientes de la literatura en la zona (Hernández-Covarrubias et al., 2003 para
camarón blanco y Galicia, 1976 para camarón azul).

79
Figura 15– Sitios de muestreo único en Sinaloa sur. 1: Huizache, 2: Chametla, 3:
Teacapán, 4: Palmilla y 5: zona de costera de Teacapán.
5.1.2. Clave edad-longitud para las especies de camarón Litopenaeus stylirostris,
L. vannamei
1) Para la obtención de la clave edad-longitud se realizó la digitalización de varias
bases de datos de reportes de frecuencias de tallas en la captura tanto de aguas
protegidas como de altamar que estuvieran cercanas a la zona de Sinaloa sur (Tabla
24).
2) Se sistematizaron las frecuencias de tallas de cada estudio en una hoja Excel en
frecuencias de longitudes totales con rangos de 5mm.
3) Para cada distribución de frecuencia de tallas se identificaron los componentes
normales (modas) que se diferenciaban claramente, mediante el método de
Bhattacharya (1967) dentro del software de análisis pesqueros FISAT II - FAO-
ICLARM Stock Assessment (v.1.2.2).

80
4) La edad relativa correspondiente a la longitud total media de cada grupo o moda se
estimó usando las ecuaciones de crecimiento previamente seleccionadas en la
sección (a) de von Bertalanffy por cada especie.
5) Se construyó una función cuadrática representando la relación entre la longitud total
media (x) con su desviación estándar (y) y se le ajustó una función para todos los
grupos modales de Lt.
6) Se construyó en un archivo Excel la clave edad-longitud representando las
distribuciones de cada grupo de edad en términos de la normal estándar para
asignar probabilidades relativas que un individuo de longitud L pertenezca a una o
varias edades.
7) La estructura de edades de la captura se obtiene como 𝐿𝑇𝑃 = 𝐸, donde 𝐿𝑇 es el
vector transpuesto de la frecuencia de las longitudes, P es la matriz de la clave edad
– longitud, y E es el vector de la estructura de edades.

81
Tabla 24– Estudios empleados en la elaboración de la clave edad por especie.
Especie de
camarón Área estudiada Referencia
Litopenaeus
vannamei
Altamar - Mazatlán, Sin. Chávez ,1973
Laguna – Huizache-Caimanero, Sin. Soto-López,1973
Laguna - Laguna Cabras, Sin. Gutiérrez,1980
Laguna – Laguna Oriental y
Occidental, Oax. Barrera,1976
Laguna - Huizache-Caimanero, Sin. Sepúlveda-Medina,1976
Laguna - Huizache-Caimanero, Sin. Menz y Bowers,1980
Altamar – Mazatlán, Sin. Sepúlveda-Medina,1991
Altamar - Topolobampo, Sin. Sepúlveda-Medina,1999
Laguna – Mar Muerto Oax-Chi. Ramos-Cruz, 2000
Laguna – Carretas-Pereyra, Chi. Rivera-Velázquez, 2010
Laguna - Huizache-Caimanero, Sin. Muñoz-Rubí et al. ,2012
Litopenaeus
stylirostris
Laguna - Bahía Guaymas y Lobos Márquez,1976
Lagunas Bahía de Santa María,
Altata-Ensenada del Pabellón, Bahía
Ceuta
Castro-Ortiz y Sánchez-
Rojas ,1976
Altamar - Mazatlán Sáenz y Lluch ,1990
Altamar – Mazatlán. INP,1974 a-d; 1975a-g
5.1.3. Uso de intervalos gnomónicos para estimar la mortalidad natural a la edad
(Mi)
Para calcular los valores de la mortalidad natural (M) por cada estadio de vida
para las tres especies de camarón se empleó el modelo de intervalos gnomónicos con
el programa “Gnomonic Intervals (GIM)” (Caddy, 1996; Morales-Bojórquez et al. 2003;
Martínez-Aguilar et al. 2005). La estimación de M para cada intervalo de tiempo
gnomónico requiere de la información sobre el número de etapas de desarrollo en el
ciclo de vida, el tiempo transcurrido durante el primer estadio que corresponde al
primer intervalo de tiempo gnomónico, y la fecundidad promedio de vida de la especie
(tabla 5 y 25).

82
El modelo presenta los supuestos de (Caddy, 1991;1996): 1) condiciones
estables en la población y ambiente; 2) existe en la población una proporción de sexos
1:1 con las mismas probabilidades de M; 3) existe “reemplazo poblacional estable”
donde al comenzar con una hembra con fecundidad media, un vector realista de M
resultaría en la sobrevivencia de al menos una hembra y un macho; 4) su ciclo de vida
se puede dividir en intervalos de tiempo donde la probabilidad de muerte por causas
naturales se puede asumir constante, y incrementando proporcionalmente la duración
de cada intervalo conforme aumenta la edad, llamados intervalos gnomónicos; 5) El
patrón de variación de M con la edad sigue el patrón generalizado donde su magnitud
decae agudamente en la primer parte del ciclo de vida, y se nivela ó alcanza una
asíntota al alcanzar la madurez sexual.
Los intervalos comienzan en t = 0 lo que sería el tiempo que tardaría la eclosión
del huevo por especie (tabla 6), donde el primer intervalo de tiempo ∆1 acaba en 𝑡1. La
duración del segundo intervalo de tiempo se estimó como ∆2= 𝛼 ∗ 𝑡2−1, donde 𝛼 es
una constante de proporcionalidad y los intervalos gnomónicos sucesivos son
calculados como ∆𝑖= (𝛼 ∗ 𝑡𝑖−1) + 𝑡𝑖−1, donde 𝑖 ≥ 3 hasta el n-ésimo intervalo
gnomónico. Las especies de camarón tienen ciclos de vida anuales, por lo que 𝑡𝑛 =
∆𝑖𝑛𝑖=1 = 365 𝑑í𝑎𝑠. La duración de ∆1 para cada especie de camarón así como el resto
de los intervalos gnomónicos de su ciclo de vida (∆𝑖; 𝑖 = 2,3,4…… , 𝑛) se basaron en
hechos biológicos reportados en la literatura (tabla 25), donde se ajustó a un ciclo de
vida con siete estadios de vida donde se fusionaron el estadio protozoea y mysis, que
a pesar de tener morfotipos diferentes, ambas son de hábitat planctónico, su duración
es similar y su agrupación permite darle continuidad a la condición establecida en el
concepto de intervalo gnomónico.
Caddy (1991) menciona que cada intervalo gnomónico debe tener una
proporción constante de muertes para el tiempo de vida total. Por lo que el producto
de 𝑀𝑖 y ∆𝑖 es constante para todos los intervalos, 𝐺 = (𝑀𝑖 ∗ ∆𝑖), donde la 𝐺 es la
proporción constante de muertes para cada intervalo y la tasa de mortalidad natural se
estima como 𝑀 = 𝐺/𝜃𝑖 − 𝜃𝑖−1, donde 𝜃 = ∆𝑖/𝑡𝑛, y representa la duración del intervalo
gnomónico como una proporción del año para una especie anual.

83
El número de individuos (𝑁𝑖) al inicio de cada intervalo gnomónico es el número
de sobrevivientes del intervalo previo (𝑁𝑖+1 = 𝑁𝑖 ∗ 𝑒(−𝑀𝑖∗𝜃𝑖); 𝑑𝑜𝑛𝑑𝑒 𝑖 ≥ 2), excepto el
primer intervalo donde se asume que los números eclosionados se asemejan a la
fecundidad media del ciclo de vida (𝑁1 = 𝐹𝑀 ∗ 𝑒(−𝑀𝑖∗𝜃𝑖)).
Los parámetros 𝛼 y 𝐺 son calculados numéricamente por el algoritmo de Newton
(Neter et al. 1996). Donde los valores de 𝛼 son elegidos cuando la suma de la duración
del ciclo de vida es igual a (𝑡𝑛 ∗ 𝐺), de modo que 𝑁𝑖 en el último intervalo sea igual a
dos, aplicando el supuesto de “reemplazo poblacional estable”.
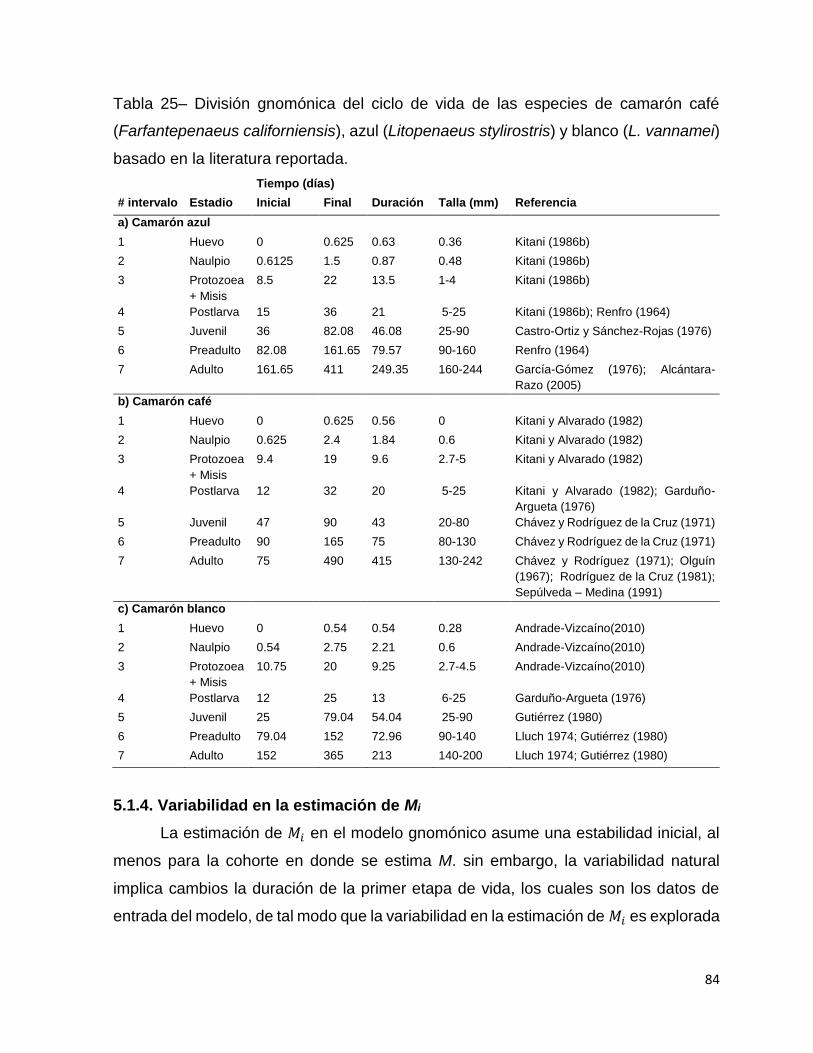
84
Tabla 25– División gnomónica del ciclo de vida de las especies de camarón café
(Farfantepenaeus californiensis), azul (Litopenaeus stylirostris) y blanco (L. vannamei)
basado en la literatura reportada.
Tiempo (días)
# intervalo Estadio Inicial Final Duración Talla (mm) Referencia
a) Camarón azul
1 Huevo 0 0.625 0.63 0.36 Kitani (1986b)
2 Naulpio 0.6125 1.5 0.87 0.48 Kitani (1986b)
3 Protozoea
+ Misis
8.5 22 13.5 1-4 Kitani (1986b)
4 Postlarva 15 36 21 5-25 Kitani (1986b); Renfro (1964)
5 Juvenil 36 82.08 46.08 25-90 Castro-Ortiz y Sánchez-Rojas (1976)
6 Preadulto 82.08 161.65 79.57 90-160 Renfro (1964)
7 Adulto 161.65 411 249.35 160-244 García-Gómez (1976); Alcántara-
Razo (2005)
b) Camarón café
1 Huevo 0 0.625 0.56 0 Kitani y Alvarado (1982)
2 Naulpio 0.625 2.4 1.84 0.6 Kitani y Alvarado (1982)
3 Protozoea
+ Misis
9.4 19 9.6 2.7-5 Kitani y Alvarado (1982)
4 Postlarva 12 32 20 5-25 Kitani y Alvarado (1982); Garduño-
Argueta (1976)
5 Juvenil 47 90 43 20-80 Chávez y Rodríguez de la Cruz (1971)
6 Preadulto 90 165 75 80-130 Chávez y Rodríguez de la Cruz (1971)
7 Adulto 75 490 415 130-242 Chávez y Rodríguez (1971); Olguín
(1967); Rodríguez de la Cruz (1981);
Sepúlveda – Medina (1991)
c) Camarón blanco
1 Huevo 0 0.54 0.54 0.28 Andrade-Vizcaíno(2010)
2 Naulpio 0.54 2.75 2.21 0.6 Andrade-Vizcaíno(2010)
3 Protozoea
+ Misis
10.75 20 9.25 2.7-4.5 Andrade-Vizcaíno(2010)
4 Postlarva 12 25 13 6-25 Garduño-Argueta (1976)
5 Juvenil 25 79.04 54.04 25-90 Gutiérrez (1980)
6 Preadulto 79.04 152 72.96 90-140 Lluch 1974; Gutiérrez (1980)
7 Adulto 152 365 213 140-200 Lluch 1974; Gutiérrez (1980)
5.1.4. Variabilidad en la estimación de Mi
La estimación de 𝑀𝑖 en el modelo gnomónico asume una estabilidad inicial, al
menos para la cohorte en donde se estima M. sin embargo, la variabilidad natural
implica cambios la duración de la primer etapa de vida, los cuales son los datos de
entrada del modelo, de tal modo que la variabilidad en la estimación de 𝑀𝑖 es explorada

85
usando cambios la duración de la primer etapa de vida. Los rangos de duración de las
etapas del ciclo de vida se muestran en las tabla 25.
5.2. Parámetros del modelo pesquero
5.2.1. Estimación del esfuerzo pesquero por tipo flota
5.2.1.1. Esfuerzo pesquero de la flota industrial camaronera
El esfuerzo pesquero observado en el área de Sinaloa sur para la temporada
2014-15 fue obtenido de la base de datos satelital otorgada por la SAGARPA-
CONAPESCA, oficinas de Mazatlán. Este se determinó de manera mensual, tomando
en cuenta solo los barcos bajo velocidad de arrastre (4-6 nudos ≈12km/hr) en el área
de Sinaloa sur.
El esfuerzo en días efectivos de la temporada 2014-15 se obtuvo de otra base
de capturas de la flota industrial para el puerto de Mazatlán de SAGARPA-
CONAPESCA, oficinas de Mazatlán. Se buscaron los nombres de los barcos que se
reportaron en la base satelital y se promediaron sus viajes en días efectivos
mensualmente.
5.2.1.2. Esfuerzo pesquero de la flota camaronera de pequeña escala
El valor estimado del esfuerzo ribereño fue obtenido a partir de entrevistas en
campo de las principales cooperativas pesqueras de camarón en la zona Mazatlán –
Palmilla (frontera con Nayarit) y de fuentes oficiales en las oficinas de pesca de Sinaloa
sur (Mazatlán, Rosario y Escuinapa). Hay que hacer notar que algunas oficinas no
contaban con el padrón actualizado o desconocían el número de socios bajo permisos
de pesca de camarón. Por lo que las entrevistas de campo fueron muy útiles para
generar un valor de esfuerzo pesquero ribereño congruente. De igual modo a través
de las entrevistas se obtuvo la dinámica del esfuerzo pesquero en el tiempo. Los
primeros dos meses de la temporada el esfuerzo pesquero aplicado es máximo,
después para el tercer mes con el descenso de la abundancia del recurso el esfuerzo
disminuye ≈ 50%, y para diciembre quedan pocos pescadores atarrayeros y la pesca
es más enfocada al autoconsumo.

86
El número de lances diarios por pescador fue estimado a partir de los muestreos
mensuales del trabajo CRIP-Mazatlán (Muñoz-Rubi et al., 2011) convirtiendo los
kilogramos de la captura y el kg/lance mensual a lances por día.
Los días de pesca por mes se obtuvieron a partir de la fecha de inicio de la
temporada de pesca de camarón para las aguas interiores y se asumió que el esfuerzo
de pesque fue diario y constante por mes y decreciente hasta el último mes que tiene
registrado captura.
El valor del esfuerzo total mensual por lances se obtuvo a partir del trabajo
CRIP-Mazatlán (Muñoz-Rubi et al., 2011) en el sistema lagunar Huizache-Caimanero,
Sinaloa y la captura total reportada en la base de datos de SAGARPA-CONAPESCA-
Mazatlán usando la fórmula:
𝑐𝑚
𝐸𝑚=
𝐶𝑇
𝐸𝑇∴ 𝐸𝑇 =
𝐸𝑚×𝐶𝑇
𝑐𝑚 (9)
Donde cm: captura del muestreo; Em esfuerzo en lances del muestreo; CT captura total
de la base de SAGARPA-CONAPESCA-Mazatlán; ET: esfuerzo total en lances
5.2.2. Bases de datos de captura por flotas
5.2.2.1. Base de la flota industrial
Con los nombres de los barcos de la base satelital se buscó su captura en peso
vivo por mes y por especie en la base de captura de la flota de altamar 2014-15 del
puerto de Mazatlán proporcionada por las oficinas de SAGARPA-CONAPESCA-
Mazatlán para la zona de Sinaloa Sur. La captura está estructurada mensualmente (de
octubre a marzo) por barco, desagregada por especie en peso vivo e incluye valores
de los días efectivos de pesca correspondientes a cada barco.
Como muchos barcos no estaban registrados para el puerto de Mazatlán se
estimó la captura de la muestra satelital con la siguiente fórmula:
𝑐𝑚
𝐸𝑚=
𝐶𝑇
𝐸𝑇∴ 𝑐𝑚 =
𝐶𝑇
𝐸𝑇× 𝐸𝑚 (10)

87
Donde cm: captura de la muestra satelital; Em esfuerzo de la muestra satelital; CT
captura total de la base de SAGARPA-CONAPESCA-Mazatlán; ET: esfuerzo total de
la base de Mazatlán.
Esto implicó valores más bajos de captura que si se tomara toda la captura
reportada en la mencionada base de datos, esto disminuye la sobrestimación de la
mortalidad por pesca (F). Además por mes se obtuvo la proporción de especies
capturadas la cual fue multiplicada a la captura estimada (cm) para no perder la
información cualitativa por especie de la captura.
5.2.2.1. Base de datos de flota camaronera de pequeña escala
La base de captura de la flota de pequeña escala 2014-15 fue proporcionada
por las oficinas de SAGARPA-CONAPESCA-Mazatlán. La captura es mensual
(septiembre a febrero), sin datos de esfuerzo, solo esta reportada el camarón blanco
en peso vivo.
5.2.3. Obtención de la estructura de edades de la captura
5.2.3.1. Flota industrial de arrastre
La estructura de longitudes fue proporcionada por la maquila Mexican Shrimp
Paradise, siendo la principal maquiladora de barcos camaroneros del puerto de
Mazatlán (trabaja para 200 barcos industriales de 380 barcos totales que operaron en
el puerto de Mazatlán 2014-15). La información de maquila es mensual (octubre a
marzo), por especie, peso de colas en libras y solo maquilan camarones U/8 a 41/50.
La base se transformó a kilogramos y se obtuvo las frecuencias relativas
mensuales de la captura maquilada por especie. Cada clasificación de maquila fue
transformada a peso total (Wt) y longitud total (Lt) por especie de acuerdo a sus
parámetros biológicos (tabla 26). Esta se graficó y se transformó a una nueva base
con intervalos de clase - longitud de 5mm estimando cada valor con los valores
obtenidos a través del área bajo la curva. Una vez estimada la base con los valores
del área bajo la curva se volvió a obtener la frecuencia relativa de los valores para
tener la correspondiente proporción de longitudes de la captura por mes y por especie

88
para la flota industrial. Cada frecuencia por talla por especie fue multiplicada por la
captura total de la muestra satelital para ese mes.
Finalmente se transformó de kilogramos de captura a individuos capturados por
clase de longitud. Se apoyó en el software Microsoft Excel y Surfer 12.
Tabla 26– Parámetros de crecimiento y peso empleadas para las especies de
camarones café (Farfantepenaeus californiensis), azul (Litopenaeus stylirostris) y
blanco (L. vannamei).
Especie 𝑾∞ 𝑳∞ 𝑲𝒎𝒆𝒔 𝒕𝟎 a1 b1 Referencia
(gr) (mm)
L. vannamei 107 230 0.2304 0.413 2.47E-07 3.65 Este estudio
L. stylirostris 135 245 0.179 0.477 6.99E-07 3.46 Rodríguez de la Cruz, 1972
F. californiensis 119 238 0.1358 0.759 3.27E-06 3.18 Galicia, 1976
1 parámetros a y b de la ecuación 𝑊𝑡 = 𝑎𝐿𝑏
5.2.3.2. Base de flota de pequeña escala
La estructura de longitudes de la captura se obtuvo mediante el informe técnico
de muestreos generado por el CRIP-Mazatlán (Muñoz-Rubi et al., 2011) en el sistema
lagunar Huizache-Caimanero, Sinaloa, para los meses de septiembre a diciembre. Se
asume que la estructura de la captura no cambio significativamente de la temporada
2011-12 al 2014-15 y es congruente para el análisis de la temporada 2014-15.
De los muestreos mensuales se obtuvo la frecuencia relativa por clase de
longitud y esta se multiplicó por la captura total reportada en el mes por SAGARPA-
CONAPESCA-Mazatlán. Para finalmente hacer la conversión de kilogramos de
captura a individuos captura por clase de longitud.
5.2.4. Modelo de capturabilidad a la edad (qi)
Para estimar la capturabilidad se empleó el modelo del programa
CATCHABILITY (Martínez-Aguilar et al., 1999). El método y la teoría fue desarrollado
por Arreguín-Sánches (1996) y se ha aplicado exitosamente en diversos recursos

89
como el camarón café del Pacífico Mexicano (García-Borbón, 2009), sardina
(Martínez-Aguilar et al., 2009), el mero rojo (Arreguín-Sanchez y Pitcher, 1999).
El método del modelo se basó de Arreguín-Sanchez y Pitcher (1999) en donde
el coeficiente de capturabilidad se estima para cada clase de longitud en un tiempo
dado 𝑞(𝑙,𝑡). La forma para representar la transformación de una distribución de
frecuencias de longitudes (representando la estructura de longitudes de la captura)
hacia otra en el tiempo es por medio de la matriz de transición (Shepherd, 1987;
Caswell, 1989), de tal manera:
𝑁(𝑙,𝑡+1) = 𝐴(𝑙,𝑘)𝑁(𝑙,𝑡) (11)
Donde 𝑘 y 𝑙 = son los intervalos de longitud sucesivos;
𝑁(𝑙,𝑡) = es el número de individuos de la captura al tiempo t; y
𝐴 = es la matriz de transición que depende del crecimiento y la
mortalidad;
La matriz de transición se expresa como:
𝐴(𝑙,𝑘) = 𝐺(𝑙,𝑘)𝑆𝑘 (12)
Donde 𝐺 = es la matriz de crecimiento en ausencia de la mortalidad; y
𝑆 = la matriz de supervivencia expresado en términos de mortalidad
natural y la selectividad del arte de pesca.
La matriz 𝐺(𝑙,𝑘) se puede estimar con el criterio definido por Shepherd (1987) donde
asigna las probabilidades de crecimiento a cada intervalo de clase de longitud. El
elemento de 𝑆𝑘 puede ser definido en términos de la mortalidad como:
𝑆(𝑘) = 𝑒−𝑍(𝑘)𝑡 = 𝑒−[𝑀+𝑞(𝑘,𝑡)𝑠(𝑘)𝐸(𝑡)] (13)

90
Donde 𝑆(𝑘)representa los valores de los elementos en la diagonal principal de la
matriz de sobrevivencia (y los demás elementos se vuelven cero; ver Caswell, 1988);
𝑍(𝑘) es la tasa instantánea de mortalidad total para el 𝑘 − é𝑠𝑖𝑚𝑜 grupo de longitud al
tiempo t; 𝑀 es la tasa instantánea de mortalidad natural ponderada (se explica más
adelante); 𝑠(𝑘) es la probabilidad de la selección del equipo de pesca para la longitud
𝑘; y 𝐸(𝑡) es el esfuerzo de pesca al tiempo t ( el cual se asume que no está dirigido a
tamaños específicos dentro de un rango de longitudes capturadas por el arte de
pesca).
Por tanto la ecuación (11) puede ser representada como:
𝑁(𝑙,𝑡+1) = 𝐺(𝑙,𝑘)𝑒−[𝑀+𝑞(𝑘,𝑡)𝑠(𝑘)𝐸(𝑡)]𝑁(𝑘,𝑡)𝑘 (14)
Ahora si 𝑁(𝑙,𝑡+1), 𝑁(𝑘,𝑡) y 𝐺(𝑙,𝑘) son conocidas, así como 𝑀, 𝑠(𝑘) y 𝐸(𝑡), entonces
𝑞(𝑘,𝑡) puede ser estimada. La ecuación (11) puede ser resuelta mediante iteraciones
para 𝑞(𝑘,𝑡) usando un algoritmo que minimice las diferencias entre los valores
observados y calculados de 𝑁(𝑙,𝑡+1).
Una vez estimados los valores de 𝑞(𝑘,𝑡), el patrón de la capturabilidad a la
longitud puede ser observado y su tendencia puede ser cuantificada. Arreguín-
Sánchez y Pitcher (1999) mencionan que aun cuando la capturabilidad a la longitud
puede determinarse empíricamente, la interpretación debe hacerse en base a la
biología del recurso.
Una vez explicado el modelo, el programa CATCHABILITY ocupa los valores
de entrada para la matriz 𝐺(𝑙,𝑘), estos son los valores máximos y mínimos de las
longitudes observadas de la estructura de longitudes de la captura por cada especie
(camarón azul, café y blanco), de todos los meses para la flota industrial y para la flota
de pequeña escala. Así mismo pide los valores de los modelos de crecimiento de von
Bertalanffy por especie indicados en la tabla 8.
Para la matriz de sobrevivencia o 𝑠(𝑘) pide como valores de entrada la
mortalidad natural constante (𝑀), el esfuerzo 𝐸(𝑡) y la estructura de longitudes de los
organismos de la captura por unidad de esfuerzo de dos meses consecutivos

91
(𝑈𝑡, 𝑈𝑡+1); como se conocen ya los valores de la mortalidad natural a la longitud (𝑀𝑖)
(Aranceta et al., 2016) se estimó una mortalidad natural ponderada 𝑀𝑝 por cada
especie de camarón. Para lo anterior:
1) Se estimó el promedio de la captura por unidad de esfuerzo en individuos por
clase de longitud (𝑈𝑖) para todos los meses (octubre – marzo para la flota de
altamar y septiembre – diciembre para la flota de pequeña escala) y se realizó su
sumatoria total ( ��𝑖).
2) Después se multiplicó cada 𝑈𝑖 por su correspondiente 𝑀𝑖 𝑎𝑛𝑢𝑎𝑙 y se hizo la
correspondiente sumatoria ��𝑖 ∗ 𝑀𝑖;
3) Finalmente se obtiene el valor de 𝑀𝑝 = ��𝑖 ∗ 𝑀𝑖
��𝑖 anual, y se convierte a valores
mensuales para ingresarlos al modelo.
El valor de esfuerzo se obtuvo 𝐸(𝑡) multiplicando el número de barcos reportados
por mes por los días efectivo de pesca (siendo días efectivos de pesca / barcos en el
área por cada mes).
Para la estimación de la 𝑞(𝑘,𝑡) del último mes (marzo para la flota industrial y
diciembre para la flota de pequeña escala) se estimó empíricamente a partir de los
patrones anteriores de la 𝑞𝑖 y de la temporalidad, biología y comportamiento de las
poblaciones por especie de camarón.
Una vez obtenidas las salidas mensuales de la capturabilidad a la longitud por
cada flota para cada especie se le ajustó una función logística para obtener una
ecuación general por especie-flota de 𝑞(𝐿,𝑡):
𝑞(𝐿,𝑡) = 𝑎
1+𝑒𝑥𝑝 −(𝑐+𝑏𝐿𝑖)
(15)
Donde 𝑎, 𝑏, 𝑐: son parámetros estimados del ajuste; 𝐿𝑖 es el intervalo de longitud.
Cabe de mencionar que para mejorar el ajuste y obtener los patrones o la
tendencia de 𝑞(𝐿,𝑡) se eliminaron datos extremos o muy cercanos a 0.
Se estimó una función general logística para todos los datos de 𝑞(𝐿,𝑡) por flota y
para cada especie. Debido a que la dispersión era muy alta se transformó los datos de
𝑞(𝐿,𝑡) a logaritmos base 10. Una vez estimada la función “general” de todos los 𝑞(𝐿,𝑡) se
le aplicó el antilog.

92
El ajuste al modelo de capturabilidad (ecuación 15) se realizó con apoyo del
programa STATISTICA 10.
5.3. Modelo bioeconómico para la pesquería multi-específica y multi-flota de
camarón
Se desarrolló un modelo dinámico de largo plazo de la pesquería y fue
desarrollado considerando la estacionalidad del reclutamiento y del esfuerzo e
integrando la dinámica de la estructura poblacional de las especies de crustáceos
capturadas por flotas heterogéneas.
5.3.1. Sub-modelo biológico
Para estimar la dinámica de las poblaciones de peneidos presentes en la
pesquería del sur del Golfo de California (Sur de Sinaloa) es fundamental poder seguir,
a través del tiempo, los cambios en el número de individuos de cada cohorte de la
población. Los individuos sobrevivientes de las diferentes especies a través de las
temporadas de pesca se calculan mediante la siguiente ecuación:
𝑑𝑁𝑖,𝑗
𝑑𝑡= −( 𝐹𝑖,𝑗,𝑚,𝑡 +𝑀𝑖,𝑗,𝑡
𝑘
)𝑁𝑖,𝑗,𝑡 (16)
donde 𝑁𝑖,𝑗,𝑡 es el número de individuos de la especie i de camarón de edad j en el
tiempo t. 𝐹𝑖,𝑗,𝑚,𝑡 es la mortalidad por pesca de individuos de la especie i, edad j causada
por la flota m en tiempo t. 𝑀𝑖,𝑗,𝑡 es la mortalidad natural de individuos de la especie i,
edad j en tiempo t. La estimación del tamaño de los stocks es necesario como dato de
entrada en el tiempo t = 0, a fin de inicializar el modelo.
Resolviendo la ecuación (16) para los individuos de la especie i, de la cohorte j
sobreviviendo en el tiempo t tenemos que:
𝑁𝑖,𝑗+1,𝑡+1 = 𝑁𝑖,𝑗,𝑡 ∙ 𝑒 −(𝑀𝑖,𝑗,𝑡+ 𝐹𝑖,𝑗,𝑚,𝑡𝑚 ) (17)
La biomasa por especie y edad en el tiempo se determina a través de:
𝐵𝑖,𝑗,𝑡 = 𝑁𝑖,𝑗,𝑡 ∙ 𝑎𝑖 ∙ 𝐿𝑖,𝑗𝑏𝑖 (18)

93
Donde, 𝐿𝑖,𝑗 es la longitud del camarón de la especie i a la edad j y 𝑎𝑖 y 𝑏𝑖 son constantes
de la relación de peso-longitud de las especies i. La longitud a diferentes edades para
las diversas especies se calcula utilizando la ecuación de crecimiento de von
Bertalanffy:
𝐿𝑖,𝑗 = 𝐿𝑖,∞ ∙ (1 − 𝑒−𝑘𝑖(𝑗−𝑗0)) (19)
La biomasa total de cada especie de camarón al final de cada mes se determina a
través de:
𝐵𝑇𝑠,𝑡 = 𝐵𝑖,𝑡
𝑖=15
𝑖=2
(20)
La estacionalidad en el reclutamiento fue modelada utilizando el modelo de
retraso distribuido (Manetsch, 1976; Seijo et al., 1997). El modelo se describe a
continuación:
)( ,1,1 ttiIDEL
gd (21)
)( ,2,2 tti
DEL
gd (22)
. . . .
. . . .
. . . .
)( ,, tgtigg
DEL
gd (23)
donde:
It = postlarvas de camarón en el área de reclutamiento
g,t= reclutas de camarón de edad 1
1,t, 2,t, ..., g,t son las tasa intermedias de retraso utilizadas en el proceso para
representar la distribución de la estacionalidad del reclutamiento.
DEL = tiempo promedio de maduración
g = orden del retraso distribuido

94
Estas ecuaciones diferenciales fueron resueltas utilizando integración numérica
de Euler para ser introducidas en el modelo desarrollado en ambiente Excel.
Integrando en el tiempo se tiene que:
(24)
(25)
(26)
Integrando numéricamente con el método de Euler se tiene que:
(27)
(28)
(29)
Las integrales numéricamente resueltas son incorporadas a la hoja de cálculo de
Excel para representar la estacionalidad en el reclutamiento de individuos a las
poblaciones de camarón.
Para estimar el número de individuos reclutas mensuales por especie se ajustó
en base a la similitud de la captura observada y estimada mensual. Esta estimación
además se basó en la distribución estacional de la información biológica de la
frecuencia relativa de hembras maduras mensuales promedio de 1981 a 2013
obteniendo además una proporción base estacional por especie. La cual es multiplica
por el valor de reclutamiento semilla por especie que genera una ecuación Beverton-
Holt que alimenta al modelo de retraso distribuido en función del aumento o
dtt
t
tttdtt dtIDEL
g))(( ,1,1,1
dtt
t
tttdtt dtDEL
g))(( ,2,1,2,2
dtt
t
tgtgtgdttg dtDEL
g))(( ,,1,,
))(( ,1,1,1 tttdtt IDEL
gDT
))(( ,2,1,2,2 tttdttDEL
gDT
))(( ,,1,, tgtgtgdttgDEL
gDT

95
disminución del stock desovante y su consecuente reclutamiento por consecuencia de
la variación en el esfuerzo pesquero. La ecuación Beverton-Holt:
𝑅 = 𝛼𝑏ℎ𝑆𝑆𝐵
[𝑋𝑠𝑝 + 𝛽𝑏ℎ] (30)
Los recluta por unidad de tiempo estarán en función de las constantes 𝛼𝑏ℎ, el
máximo número posible de reclutas, y 𝛽𝑏ℎ, la biomasa del stock desovante (𝑆𝑆𝐵)
necesario para producir en promedio 𝛼𝑏ℎ 2⁄ reclutas. Las cuales son estimadas con la
herramienta de optimización de Excel SOLVER.
5.3.2. Sub-modelo Tecnológico
Para iniciar este sub-modelo, es necesario el esfuerzo de la temporada 2014-15
(número de barcos o atarrayas y días totales de pesca) para cada flota. Se requiere
también la longitud de la temporada de veda. El primer paso es calcular la mortalidad
por pesca estacional por tipo de flota. La mortalidad por pesca se calcula por edades
de acuerdo a la siguiente ecuación
𝐹𝑖,𝑗,𝑚,𝑡 = 𝑓𝑡,𝑚 ∙ 𝑆𝑒𝑙𝑖,𝑗,𝑚 ∙ 𝑞𝑖,𝑗,𝑚,𝑡 (31)
Donde 𝑓𝑡,𝑚 representa los días de pesca por magnitud mensual de tipo de flota m (e.i.
pequeña escala o industrial); 𝑆𝑒𝑙𝑖,𝑗,𝑚 representa el patrón de selectividad por especie
i, por edad j y por tipo de flota m. 𝑞𝑖,𝑗,𝑚,𝑡 representa el coeficiente de capturabilidad por
especie i, por edad j, por tipo de flota m en el tiempo t (meses de la temporada. El
número actual de días de pesca se requiere para iniciar el modelo.
Para estimar la captura en el tiempo (meses) por especie, edad y tipo de
embarcación, se utiliza la siguiente ecuación de captura
𝐶𝑖,𝑗,𝑚,𝑡 = 𝐹𝑖,𝑗,𝑚,𝑡
𝐹𝑖,𝑗,𝑚,𝑡+𝑀𝑖,𝑗,𝑡 [1 − 𝑒−(𝐹𝑡,𝑖,𝑗,𝑚+𝑀𝑖,𝑗,𝑡)] ∙ 𝑁𝑖,𝑗,𝑡 (32)
La captura durante el año se estima de la siguiente manera

96
𝐶𝑚,𝑡 = 𝐶𝑖,𝑗,𝑚,𝑡
𝑗=12
𝑗=1
(33)
5.3.3 Sub-modelo económico
Todos los costos e ingresos se reportan en dólares (US$) con una tasa de cambio
de 14.33 pesos / dólar (promedio de la temporada 2014-15). El ingreso por flota se
calcula utilizando la ecuación:
𝑇𝑅𝑚,𝑡 = 𝐶𝑖,𝑗,𝑚,𝑡 ∙ 𝑝𝑖,𝑗,𝑚 + 𝐼𝑡𝑖,𝑗
(34)
En donde 𝑝𝑖,𝑗,𝑚 es un vector de los precios de playa por especie i, talla j y flota m. este
vector multiplica a la captura dependiente de la especie i, edad j, flota m y tiempo t y
se le suma 𝐼𝑡, el ingreso por captura incidental por flota.
Las ganancias generadas para cada flota por temporada de pesca se calculan
de la siguiente manera:
∏ = 𝑇𝑅𝑚,𝑡 − 𝑇𝐶𝑚,𝑡𝑚,𝑡
(35)
Donde 𝑇𝐶𝑚,𝑡 son los costos totales del tipo de flota m en la temporada t. Las utilidades
(o pérdidas) anuales se determinan agregando las ganancias o pérdidas mensuales
durante el año.
Los costos totales por embarcación por día de pesca se estimaron para la flota
industrial mediante la información aportada por el armador Jesús Ortiz de la flota de
Pesca Mar de Manzanillo, S.A. de C.V. con 13 embarcaciones (tabla 27). Para el caso
de la flota de pequeña escala se realizaron encuestas a distintos presidentes de
cooperativas (tabla 28). También se incluye el costo por tipo de embarcación / flota
(tabla 29) y el costo de oportunidad de cada flota (tabla 30).
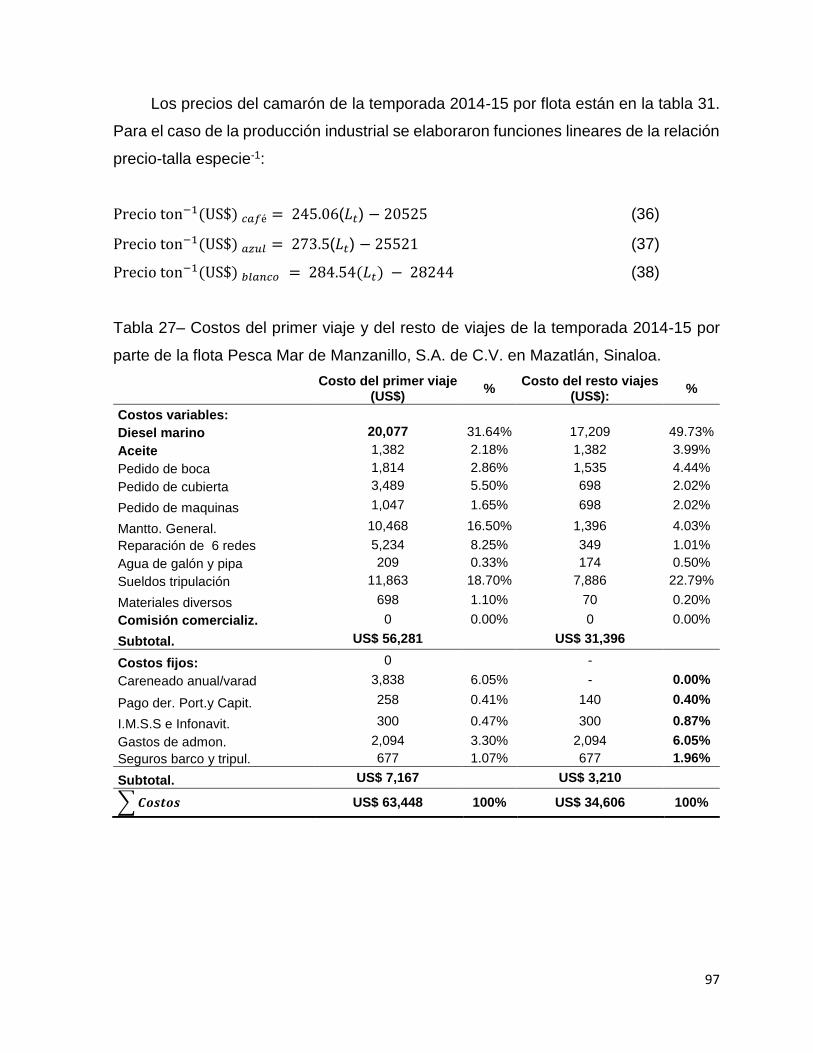
97
Los precios del camarón de la temporada 2014-15 por flota están en la tabla 31.
Para el caso de la producción industrial se elaboraron funciones lineares de la relación
precio-talla especie-1:
Precio ton−1(US$) 𝑐𝑎𝑓é = 245.06(𝐿𝑡) − 20525 (36)
Precio ton−1(US$) 𝑎𝑧𝑢𝑙 = 273.5(𝐿𝑡) − 25521 (37)
Precio ton−1(US$) 𝑏𝑙𝑎𝑛𝑐𝑜 = 284.54(𝐿𝑡) − 28244 (38)
Tabla 27– Costos del primer viaje y del resto de viajes de la temporada 2014-15 por
parte de la flota Pesca Mar de Manzanillo, S.A. de C.V. en Mazatlán, Sinaloa.
Costo del primer viaje
(US$) %
Costo del resto viajes (US$):
%
Costos variables:
Diesel marino 20,077 31.64% 17,209 49.73%
Aceite 1,382 2.18% 1,382 3.99%
Pedido de boca 1,814 2.86% 1,535 4.44%
Pedido de cubierta 3,489 5.50% 698 2.02%
Pedido de maquinas 1,047 1.65% 698 2.02%
Mantto. General. 10,468 16.50% 1,396 4.03%
Reparación de 6 redes 5,234 8.25% 349 1.01%
Agua de galón y pipa 209 0.33% 174 0.50%
Sueldos tripulación 11,863 18.70% 7,886 22.79%
Materiales diversos 698 1.10% 70 0.20%
Comisión comercializ. 0 0.00% 0 0.00%
Subtotal. US$ 56,281 US$ 31,396
Costos fijos: 0 -
Careneado anual/varad 3,838 6.05% - 0.00%
Pago der. Port.y Capit. 258 0.41% 140 0.40%
I.M.S.S e Infonavit. 300 0.47% 300 0.87%
Gastos de admon. 2,094 3.30% 2,094 6.05%
Seguros barco y tripul. 677 1.07% 677 1.96%
Subtotal. US$ 7,167 US$ 3,210
𝑪𝒐𝒔𝒕𝒐𝒔 US$ 63,448 100% US$ 34,606 100%

98
Tabla 28– Costos variables de la flota de pequeña escala para la temporada 2014-15
en Sinaloa sur.
Costos variables Costo temporada-1 (US$)
a) Materiales de pesca nocturna:
Candil de diésel 4.88
Tanque de gas 23.26
Gas L.P. (recarga) 203.21
Capuchones 1.95
Bombillas (2/temporada) 6.98
Garzas 34.89
Subtotal 275.18
b) Alimento 746.69
Subtotal 746.69
c) Reparación de equipo 279.13
Subtotal 279.13
Total 1,301
*temporada de cuatro meses.
Tabla 29- Costo de pesca industrial y de pequeña escala por día (US$).
a) Días efectivos pesca SAGARPA flota de alta mar
Sep Oct Nov Dic Ene Feb Mar
Camarón azul - 17.69 21.02 27.25 22.61 31.35 21.28
Camarón blanco - 17.60 22.19 27.81 23.72 30.84 24.00
Camarón café - 17.71 19.75 27.51 22.89 31.58 20.95
Promedio - 17.67 20.98 27.52 23.07 31.26 22.08
Costo industrial de pesca/día (US$) 3,589 1,648 1,257 1,499 1,107 1,567
b) Días efectivos pesca SAGARPA flota de alta mar
Sep Oct Nov Dic Ene Feb Mar
Camarón blanco 23 25 25 25 - - -
Costo de pequeña escala de pesca/día (US$) 17.18 15.80 15.80 15.80
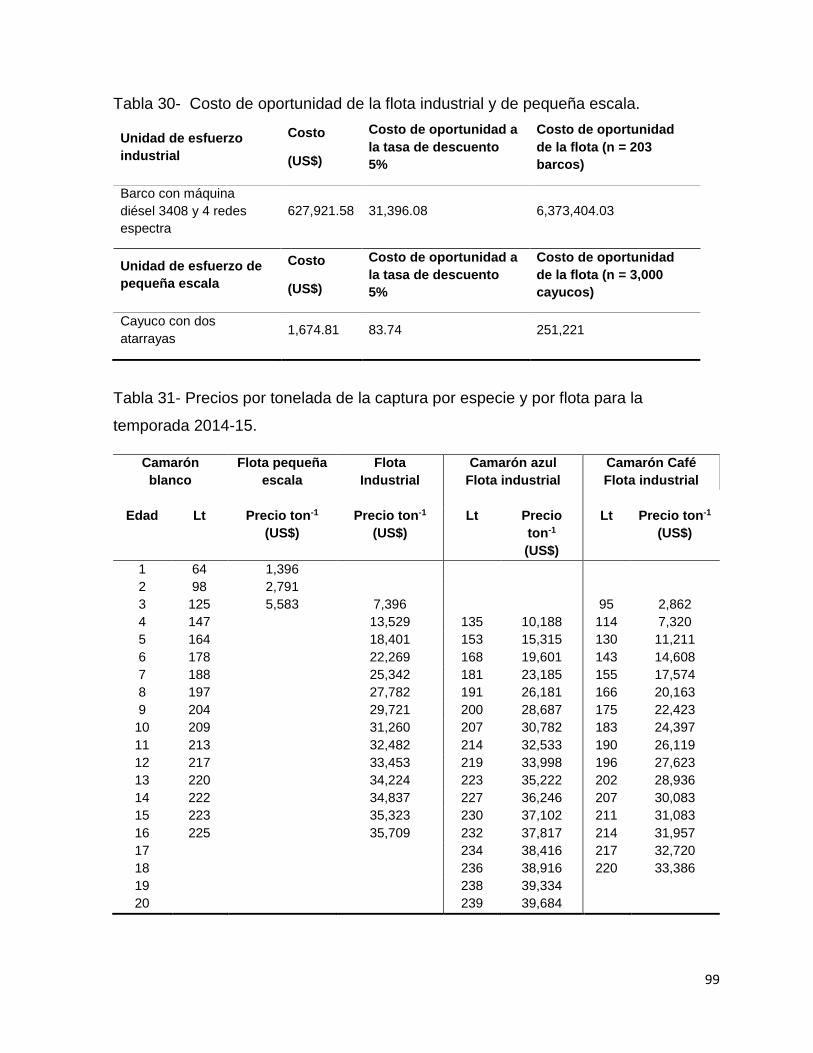
99
Tabla 30- Costo de oportunidad de la flota industrial y de pequeña escala.
Unidad de esfuerzo
industrial
Costo
(US$)
Costo de oportunidad a
la tasa de descuento
5%
Costo de oportunidad
de la flota (n = 203
barcos)
Barco con máquina
diésel 3408 y 4 redes
espectra
627,921.58 31,396.08 6,373,404.03
Unidad de esfuerzo de
pequeña escala
Costo
(US$)
Costo de oportunidad a
la tasa de descuento
5%
Costo de oportunidad
de la flota (n = 3,000
cayucos)
Cayuco con dos
atarrayas 1,674.81 83.74 251,221
Tabla 31- Precios por tonelada de la captura por especie y por flota para la
temporada 2014-15.
Camarón
blanco
Flota pequeña
escala
Flota
Industrial
Camarón azul
Flota industrial
Camarón Café
Flota industrial
Edad Lt Precio ton-1
(US$)
Precio ton-1
(US$)
Lt Precio
ton-1
(US$)
Lt Precio ton-1
(US$)
1 64 1,396
2 98 2,791
3 125 5,583 7,396 95 2,862
4 147 13,529 135 10,188 114 7,320
5 164 18,401 153 15,315 130 11,211
6 178 22,269 168 19,601 143 14,608
7 188 25,342 181 23,185 155 17,574
8 197 27,782 191 26,181 166 20,163
9 204 29,721 200 28,687 175 22,423
10 209 31,260 207 30,782 183 24,397
11 213 32,482 214 32,533 190 26,119
12 217 33,453 219 33,998 196 27,623
13 220 34,224 223 35,222 202 28,936
14 222 34,837 227 36,246 207 30,083
15 223 35,323 230 37,102 211 31,083
16 225 35,709 232 37,817 214 31,957
17 234 38,416 217 32,720
18 236 38,916 220 33,386
19 238 39,334
20 239 39,684

100
5.3.3.1 Valor presente neto para la pesquería
El valor presente neto (VPN) fue calculado a 4 años mediante la siguiente ecuación:
Tt
tt
tmmNPV
0 1 (39)
donde es la tasa de descuento.
5.3.3.2. Ingresos por pesca incidental
Mediante los avisos de arribo de 13 barcos (flota de Pesca Mar de Manzanillo,
S.A. de C.V.) se estimó un aproximado de la pesca incidental para la pesca industrial
del sur de Sinaloa (tabla 32). Este ingreso su adicionado a la renta generada por la
captura comercial de las especies de camarón objetivo.
Con toda la información anterior se realizó la tabla de parámetros
bioeconómicos que se emplearán en el modelo bioeconómico para la pesquería de
camarón en Sinaloa sur (tabla 33).

101
Tabla 32 – Captura incidental por barco y flota, especie y mes en toneladas al día y en ingresos (US$).
Especie precio/ton
(US$) Oct Nov Dic Ene Feb Mar
Total (ton día-1)
Camarón botalón cola 5,583 - 0.0004 - - - -
C. botalón entero 5,583 0.00028 - - - - -
C. cristal cola 5,583 - 0.0040 - 0.0285 0.0579 -
C. cristal entero 5,583 - - - - - -
Huachinango fresco 4,187 0.0105 0.0101 0.0020 0.0316 0.0044 0.0013
Berrugata fresco 2,791 0.0500 0.0753 0.0213 0.0154 0.0346 0.0142
Calamar chico fresco 1,396 0.0002 0.0029 - 0.0011 0.0020 -
Jurel entero fresco 2,791 - 0.0032 - - - -
Burro fresco 2,791 - 0.0022 0.0015 0.0066 - -
Pescado revuelto 2,094 0.0106 - - - - -
Lenguado 2,791 - - - - - 0.0033
Ton/día/barco 0.0717 0.0981 0.0248 0.0832 0.0989 0.0188 0.3955
Ton/día/flota 13.55 9.32 3.48 13.73 20.07 2.54 62.68
Ingresos:
US$/día/barco
208 296 72 354 441 54 1,426
US$/día/flota 39,285 28,136 10,097 58,489 89,486 7,347 232,840
Precio ponderado por
flota (US$/Ton) 2,900 3,019 2,905 4,260 4,458 2,890 20,433
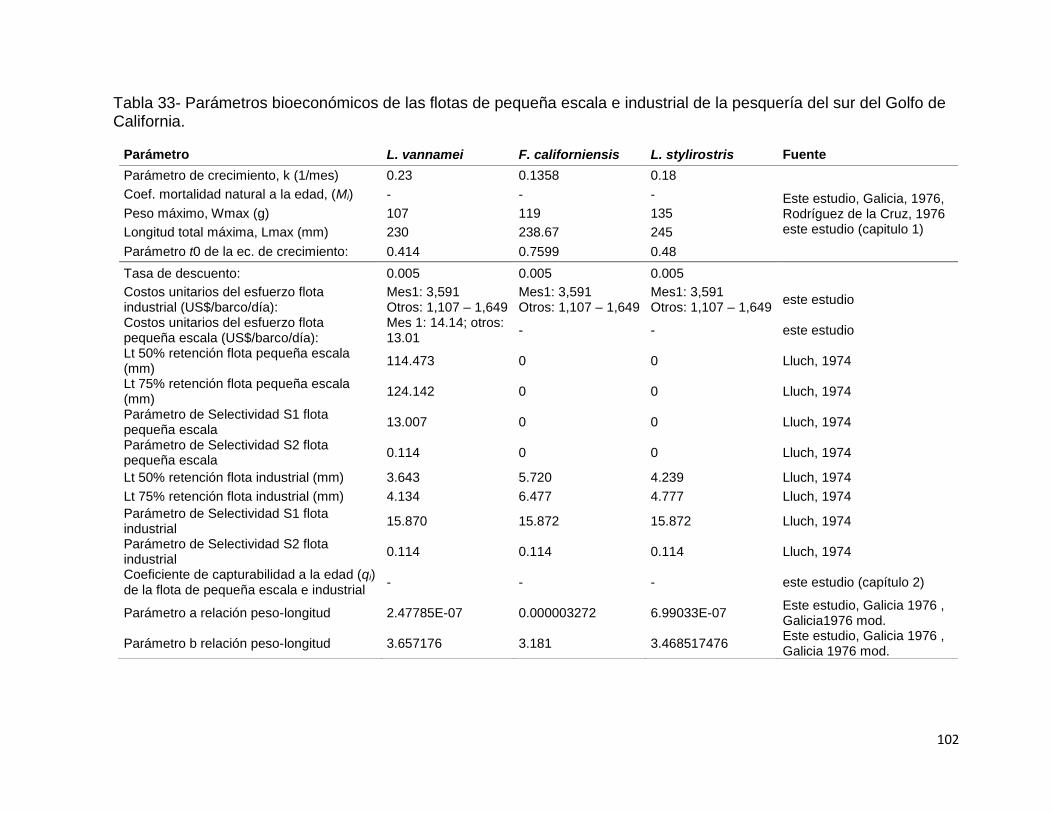
102
Tabla 33- Parámetros bioeconómicos de las flotas de pequeña escala e industrial de la pesquería del sur del Golfo de California.
Parámetro L. vannamei F. californiensis L. stylirostris Fuente
Parámetro de crecimiento, k (1/mes) 0.23 0.1358 0.18
Este estudio, Galicia, 1976, Rodríguez de la Cruz, 1976 este estudio (capitulo 1)
Coef. mortalidad natural a la edad, (Mi) - - -
Peso máximo, Wmax (g) 107 119 135
Longitud total máxima, Lmax (mm) 230 238.67 245
Parámetro t0 de la ec. de crecimiento: 0.414 0.7599 0.48
Tasa de descuento: 0.005 0.005 0.005
Costos unitarios del esfuerzo flota industrial (US$/barco/día):
Mes1: 3,591 Otros: 1,107 – 1,649
Mes1: 3,591 Otros: 1,107 – 1,649
Mes1: 3,591 Otros: 1,107 – 1,649
este estudio
Costos unitarios del esfuerzo flota pequeña escala (US$/barco/día):
Mes 1: 14.14; otros: 13.01
- - este estudio
Lt 50% retención flota pequeña escala (mm)
114.473 0 0 Lluch, 1974
Lt 75% retención flota pequeña escala (mm)
124.142 0 0 Lluch, 1974
Parámetro de Selectividad S1 flota pequeña escala
13.007 0 0 Lluch, 1974
Parámetro de Selectividad S2 flota pequeña escala
0.114 0 0 Lluch, 1974
Lt 50% retención flota industrial (mm) 3.643 5.720 4.239 Lluch, 1974
Lt 75% retención flota industrial (mm) 4.134 6.477 4.777 Lluch, 1974
Parámetro de Selectividad S1 flota industrial
15.870 15.872 15.872 Lluch, 1974
Parámetro de Selectividad S2 flota industrial
0.114 0.114 0.114 Lluch, 1974
Coeficiente de capturabilidad a la edad (qi) de la flota de pequeña escala e industrial
- - - este estudio (capítulo 2)
Parámetro a relación peso-longitud 2.47785E-07 0.000003272 6.99033E-07 Este estudio, Galicia 1976 , Galicia1976 mod.
Parámetro b relación peso-longitud 3.657176 3.181 3.468517476 Este estudio, Galicia 1976 , Galicia 1976 mod.

103
5.3.4. Sub-modelo ambiental - Escenarios de reclutamiento en función de su
variabilidad interanual
Para representar el reclutamiento a las lagunas (individuos postlarvas) nos
basamos en la información disponible y más actual que generó el Instituto Nacional de
Pesca (Fig 16). El único índice reproductivo en el área de estudio que presenta datos
de variabilidad interanual mensual y en el tiempo es el porcentaje de hembras maduras
o en fase III de su ciclo gonádico. Para este modelo se asume que el porcentaje de
hembras maduras son proporcionales al reclutamiento que se observan en la
temporada de captura simulada en el modelo, este supuesto es apoyado por estudios
que relacionan la abundancia del stock reproductor con l magnitud reclutamiento en
pesquerías de camarón (Ye, 2000).
Para lo anterior se basó en la información contenida en el informe técnico de la
temporada 2013 para el “Dictamen de inicio de veda 2013” (INP, 2013). El instituto
manejó la información como diagramas de cajas (en inglés Box plots) por especie y
por mes con promedios históricos donde incluye los percentiles 75-25 y valores
máximos y mínimos (fig.16) del porcentaje de hembras maduras.

104
Figura 16 – Porcentaje de hembras maduras en fase III para el camarón blanco (a),
azul (b) y café (c).

105
5.3.5 Escenarios de manejo pesquero
La pesquería secuencial de camarón en el Pacifico mexicano involucra el
manejo de dos flotas. La flota de pequeña escala o artesanal es prácticamente
inmanejable por la cantidad tan grande de usuarios que dependen de ella. Por ese
preciso detalle es la que presenta más conflictos de ordenamiento (se desconoce el
número de usuarios), de vigilancia (pesca ilegal y captura en época de vedas, com.
personal) y de falta de imposición de la normatividad (no hay control del esfuerzo; la
flota no declara el 100% de su captura; uso de artes de pesca prohibidos: uso de redes
de arrastre tipo changos por pescadores ilegales, y uso de luz de malla chica; com.
personal). Además de los potenciales conflictos sociales causales de imponer la
correcta regulación para las miles de personas que están involucrada en ella.
Por otro lado la flota industrial si está en términos generales regulada: hay un
control del esfuerzo en barcos, existen programas de retiro de embarcaciones, existe
la localización satelital para observar las operaciones, hay revisión del uso de redes y
luz de malla legales así como excluidores de tortugas y peces, y declaran las capturas
con avisos de arribo en los puertos.
Con todo lo anterior para establecer el escenario más cercano a la realidad se
hará un manejo sobre la flota industrial y se dejara a la flota de pequeña escala en su
condición actual o de status quo (por las dificultades antes mencionadas). El manejo
propuesto tiene como objetivos la conservación del recurso y maximizar las utilidades
económicas (tabla 21) en base al criterio de bienestar de Pareto seguridad (tabla 22),
donde se busca mejorar la utilidad de la pesquería regulando el esfuerzo pesquero
industrial llevándola a una solución Pareto o a la frontera de la gran utilidad (punto de
máxima eficiencia económica) dejando la flota artesanal en su status quo pero se
espera reciban una externalidad positiva por el aumento del reclutamiento de las
especies. Dentro de las ventajas al usar este criterio antes mencionadas es que no
permite un daño económico a ningún usuario y de manera global todos igualan o
mejoran su situación de bienestar (i.e. su utilidad en la pesquería).
Para llegar a la frontera de la gran utilidad se maximizará el valor presente neto
(VPN) de toda la pesquería controlando el esfuerzo industrial, en días o barcos, bajo
el punto de referencia de manejo llamado Máximo Rendimiento Económico (siglas en

106
inglés MEY). Esta estrategia de manejo propuesto es viable de aplicar porque ya
existen programas encaminados al retiro de embarcaciones industriales. En teoría la
estrategia de manejo pretende, al maximizar la renta económica con las variables
control, disminuir el esfuerzo pesquero, trayendo consigo un aumento en la producción
derivado de una disminución de barcos y una menor explotación del recurso, esto trae
como efecto un aumento en el stock desovante y por consiguiente mayor número de
reclutas. Este último se trabajó con la ecuación B-H donde una vez establecido el
escenario status quo para cada especie con los parámetros preestablecido de la
ecuación B-H (ec. 30), este se modificaba automáticamente al cambiar el esfuerzo
pesquero, mostrando aumento o disminución del reclutamiento en función del tamaño
del stock desovante sobreviviente. La estrategia global del manejo propuesta se
encuentra en la tabla 34.

107
Tabla 34 – Manejo pesquero propuesto para la pesquería de camarón del sur de Sinaloa en la temporada 2014-15.
Flota Criterio Punto de referencia
objetivo del manejo
Variable estado Variable control Objetivos empatados con el plan
de manejo del camarón (5/6)
Industrial Pareto seguridad Máximo rendimiento
económico
Valor Presente Neto
(VPN) a cuatro años
Utilidades por flota
Reclutamiento
Stock desovante
Esfuerzo:
a) Barcos
b) Días de pesca
1) Conducir pesquería a niveles
sustentables;
2) conservar rendimiento y beneficio
económico;
3) reducir interacciones entre
sectores y
4) ambientales;
5) promover beneficios económicos
sociales.
Artesanal Pareto seguridad Status quo - -
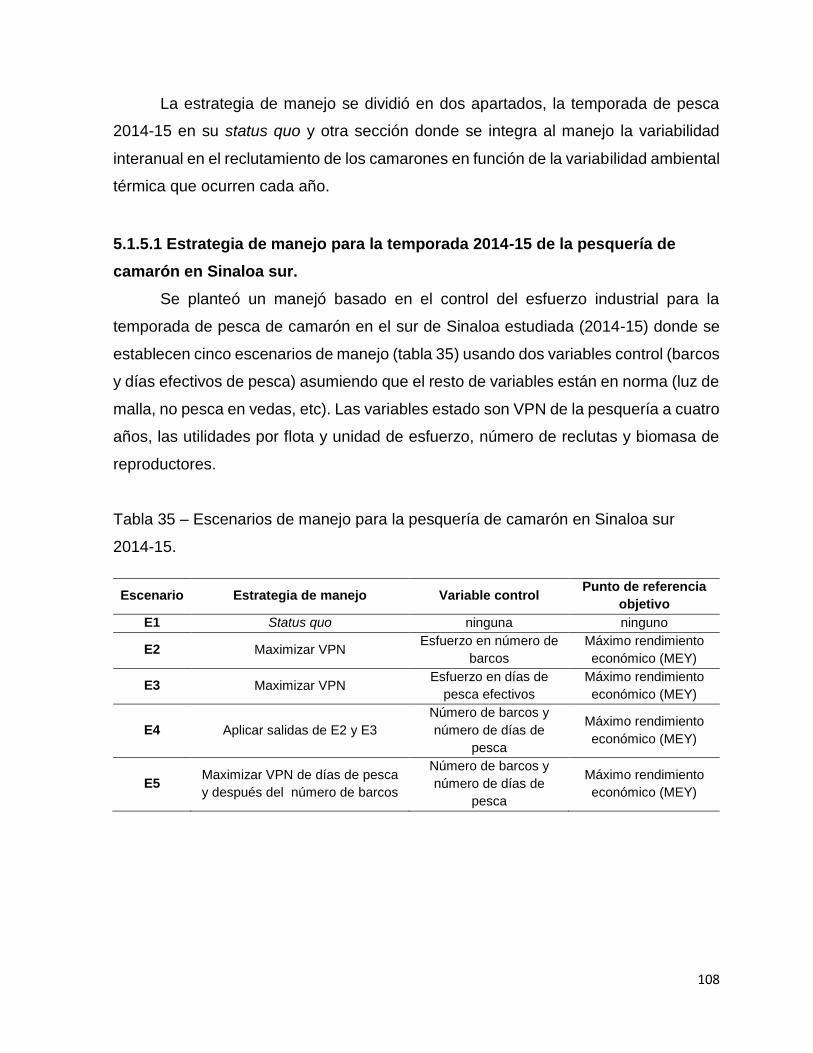
108
La estrategia de manejo se dividió en dos apartados, la temporada de pesca
2014-15 en su status quo y otra sección donde se integra al manejo la variabilidad
interanual en el reclutamiento de los camarones en función de la variabilidad ambiental
térmica que ocurren cada año.
5.1.5.1 Estrategia de manejo para la temporada 2014-15 de la pesquería de
camarón en Sinaloa sur.
Se planteó un manejó basado en el control del esfuerzo industrial para la
temporada de pesca de camarón en el sur de Sinaloa estudiada (2014-15) donde se
establecen cinco escenarios de manejo (tabla 35) usando dos variables control (barcos
y días efectivos de pesca) asumiendo que el resto de variables están en norma (luz de
malla, no pesca en vedas, etc). Las variables estado son VPN de la pesquería a cuatro
años, las utilidades por flota y unidad de esfuerzo, número de reclutas y biomasa de
reproductores.
Tabla 35 – Escenarios de manejo para la pesquería de camarón en Sinaloa sur
2014-15.
Escenario Estrategia de manejo Variable control Punto de referencia
objetivo
E1 Status quo ninguna ninguno
E2 Maximizar VPN Esfuerzo en número de
barcos
Máximo rendimiento
económico (MEY)
E3 Maximizar VPN Esfuerzo en días de
pesca efectivos
Máximo rendimiento
económico (MEY)
E4 Aplicar salidas de E2 y E3
Número de barcos y
número de días de
pesca
Máximo rendimiento
económico (MEY)
E5 Maximizar VPN de días de pesca
y después del número de barcos
Número de barcos y
número de días de
pesca
Máximo rendimiento
económico (MEY)

109
5.1.5.2 Estrategia de manejo con escenarios de variabilidad interanual del
reclutamiento para la temporada 2014-15 de la pesquería de camarón en Sinaloa
sur
Para el manejo de la pesquería bajo escenarios de variabilidad de reclutamiento
en función de las condiciones climáticas se plantearon cuatro escenarios que
relacionan la variabilidad climática con el reclutamiento (tabla 36).
La manera de integrar la variabilidad del reclutamiento fue obteniendo la
diferencia entre la frecuencia relativa de hembras maduras promedio con los máximos
reportados (simulando condiciones cálidas moderadas donde se reporta el máximo de
hembras maduras); un punto intermedio (percentil 75) simulando condiciones cálidas
menos intensas o débiles. y por debajo del promedio en el percentil 25 se simulan
condiciones de enfriamiento débiles donde el porcentajes de hembras disminuye del
promedio. Esta diferencia se le suma o resta en porcentaje a la frecuencia relativa del
escenario base (Status quo) que multiplica al valor semilla de reclutamiento B-H
estimado (ec. 30) por cada escenario e integrando así la variabilidad interanual al
modelo.
Tabla 36- Escenarios de reclutamiento basados en el efecto de las variables
ambientales sobre la reproducción de las especies de camarón del Pacífico.
Escenario Tipo de variabilidad ambiental
1) Status quo Ambiente promedio (sin calentamiento-enfriamiento)
2) Percentil 75 Calentamiento débil ≈ Niño débil: aumento en la
anomalía de 0.5 a 1.0°C en la TSM.
3) Percentil 25 Enfriamiento débil ≈ Niña débil: disminución en la
anomalía de 0.5 a 1.0°C en la TSM.
4) Máximo valor Calentamiento moderado ≈ Niño moderado: aumento
en la anomalía de 1.0 a 1.5°C en la TSM.
*TSM= temperatura superficial del mar.

110
6. Resultados
6.1. Parámetros de la dinámica poblacional
6.1.1. Selección de ecuaciones de crecimiento para las especies de especies
de camarón Litopenaeus stylirostris, L. vannamei y Farfantepenaeus
californiensis.
a) Camarón blanco
Las ecuaciones propuestas por otros autores no daban el mejor ajuste a los
datos observados de los camarones en Sinaloa sur. Para el caso de Chávez (1973)
hubo una subestimación del peso total infinito (𝑊∞) y de la longitud infinita (𝐿∞),
donde el autor reporta 𝑊∞= 26grs y 𝐿∞=200mm contra las mediciones observadas
de 𝑊∞=100grs y el valor teórico de 𝐿∞=230mm de FAO, (2006) y Romero-Moreno,
(2009). El ajuste de los nuevos valores propuestos en este estudio se encuentra en
la tabla 37.
Tabla 37– Parámetros de la ecuación de crecimiento estimados en este estudio para
el camarón blanco de Sinaloa sur.
Parámetro Valor estimado
𝑊∞ 107.481
𝐿∞ 230
𝐾𝑎ñ𝑜 2.7649
𝑡0 -0.4136
λ𝑎ñ𝑜 1.5 años
* Parámetros a y b de relación longitud peso: a= 2.47E-07, b= 3.65
Se comparó con el modelo de crecimiento propuesto por Romero-Moreno
(2009) para Nayarit y Sinaloa (fig. 17; ver tabla 8). Los cuales no muestran mayores
diferencias, excepto que el tiempo 0 del modelo propuesto para camarón blanco
equivale a individuos de 20mm ó a la fase postlarva en este estudio.

111
Figura 17– Ecuaciones de crecimiento de von Bertalanffy aplicadas al camarón
blanco en el Pacífico Mexicano.
b) Camarón azul
Para el caso del camarón azul las estimaciones de las ecuaciones de
crecimiento no muestran mucha variación entre los distintos autores, y los
parámetros que mejor se ajustaron con los datos observados fueron los de
Rodríguez de la Cruz (1976) [ver figura 18]. Además estos parámetros son
empleados en otros estudios pesqueros (Saenz y Lluch, 1990). Los parámetros
peso – longitud se estimaron con los datos de Galicia (1976) junto con las
mediciones in situ para Sinaloa sur obteniendo a= 6.9903E-07 y b= 3.46.
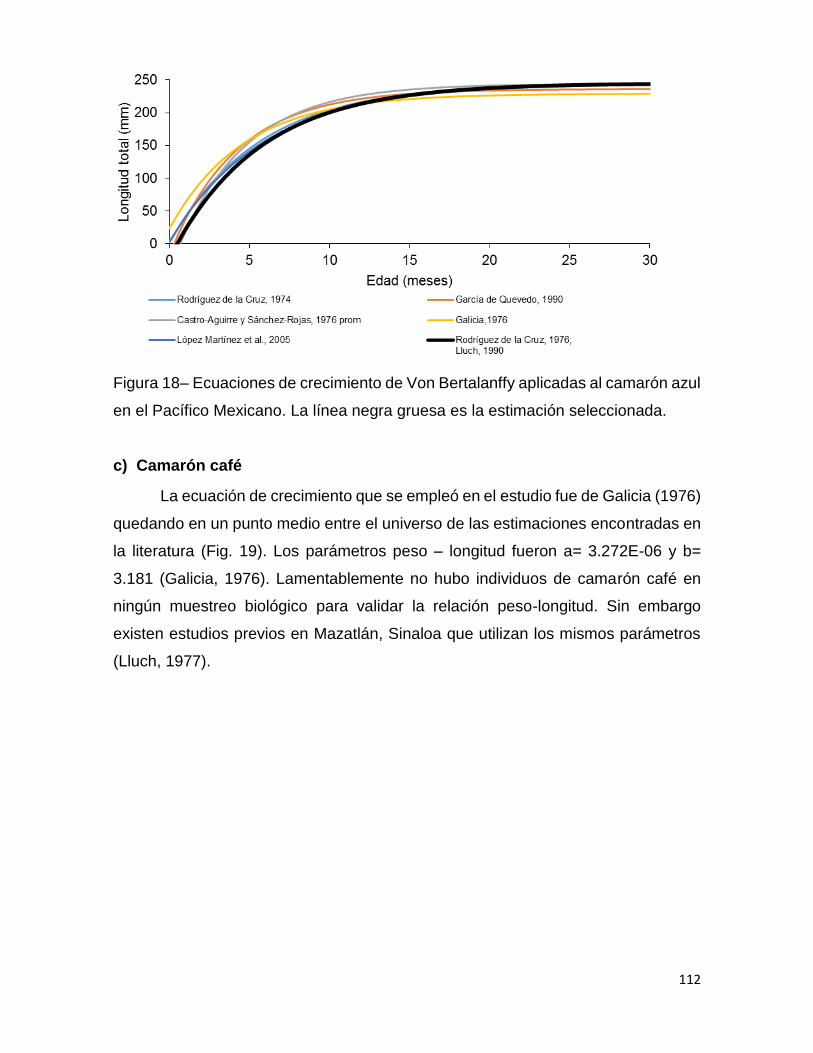
112
Figura 18– Ecuaciones de crecimiento de Von Bertalanffy aplicadas al camarón azul
en el Pacífico Mexicano. La línea negra gruesa es la estimación seleccionada.
c) Camarón café
La ecuación de crecimiento que se empleó en el estudio fue de Galicia (1976)
quedando en un punto medio entre el universo de las estimaciones encontradas en
la literatura (Fig. 19). Los parámetros peso – longitud fueron a= 3.272E-06 y b=
3.181 (Galicia, 1976). Lamentablemente no hubo individuos de camarón café en
ningún muestreo biológico para validar la relación peso-longitud. Sin embargo
existen estudios previos en Mazatlán, Sinaloa que utilizan los mismos parámetros
(Lluch, 1977).

113
Figura 19– Ecuaciones de crecimiento de von Bertalanffy aplicadas al camarón café
en el Pacífico Mexicano.
Finalmente el modelo de crecimiento para las tres especies se puede
observar en la figura 20. Se puede observar que la especie de hábitos marinos
dominantes crece más rápido que sus contra partes de hábitos estuarino-marinos.
Así mismo el modelo muestra que la especie más grande fue el camarón azul.

114
Figura 20 – Comparación entre las ecuaciones de crecimiento de von Bertalanffy
seleccionadas para el presente trabajo en Sinaloa sur.
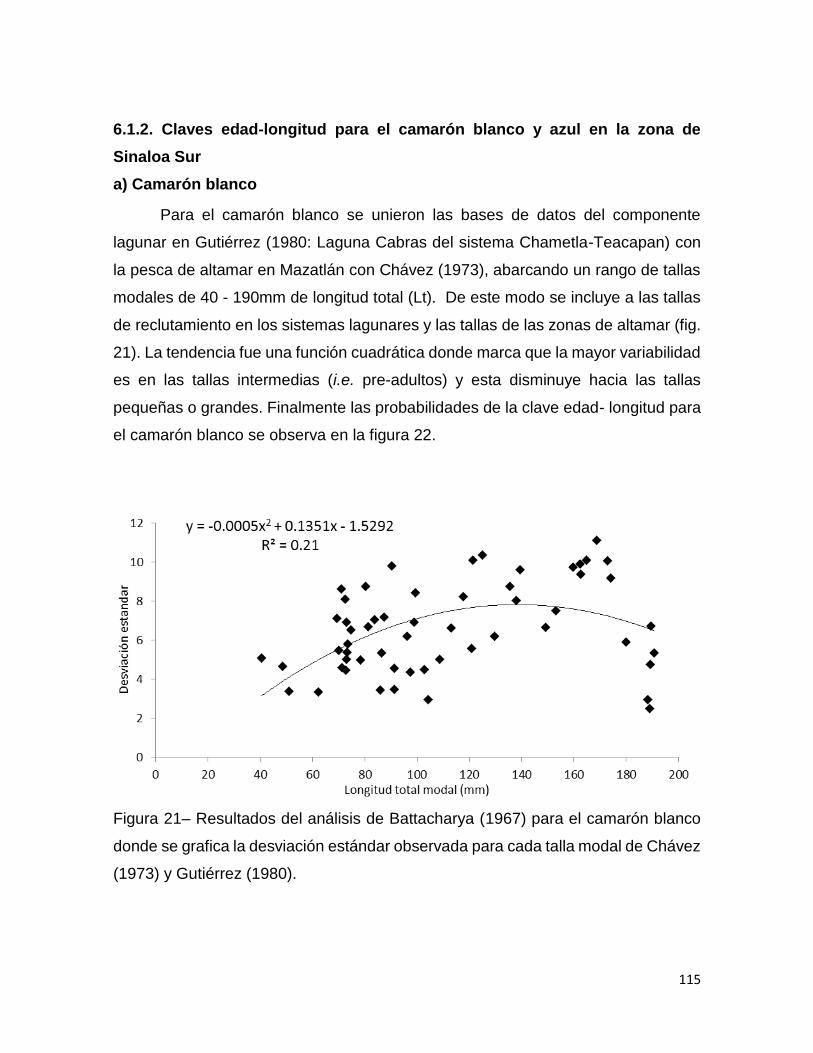
115
6.1.2. Claves edad-longitud para el camarón blanco y azul en la zona de
Sinaloa Sur
a) Camarón blanco
Para el camarón blanco se unieron las bases de datos del componente
lagunar en Gutiérrez (1980: Laguna Cabras del sistema Chametla-Teacapan) con
la pesca de altamar en Mazatlán con Chávez (1973), abarcando un rango de tallas
modales de 40 - 190mm de longitud total (Lt). De este modo se incluye a las tallas
de reclutamiento en los sistemas lagunares y las tallas de las zonas de altamar (fig.
21). La tendencia fue una función cuadrática donde marca que la mayor variabilidad
es en las tallas intermedias (i.e. pre-adultos) y esta disminuye hacia las tallas
pequeñas o grandes. Finalmente las probabilidades de la clave edad- longitud para
el camarón blanco se observa en la figura 22.
Figura 21– Resultados del análisis de Battacharya (1967) para el camarón blanco
donde se grafica la desviación estándar observada para cada talla modal de Chávez
(1973) y Gutiérrez (1980).

116
Figura 22- Clave edad-longitud en términos de probabilidad para el camarón blanco
Litopenaeus vannamei.
b) Camarón azul
Para el camarón azul se unieron las bases de datos del componente lagunar
del estudio en la Bahía Santa María, en el centro de Sinaloa de Castro-Ortiz y
Sánchez-Rojas (1976) e informes técnicos de la pesquería de altamar (INP, 1974a-
d; 1975a-g) para el puerto de Mazatlán; incluyendo todas las tallas modales
posibles del ciclo de vida del camarón azul (i.e.14 mm hasta 224 mm Lt; fig.23). De
igual manera se ajustó una ecuación cuadrática al conjunto de datos generado la
clave edad-longitud de la figura 23 y finalmente la clave edad-longitud en la figura
24.
A) CAMARÓN BLANCO (Litopenaeus vannamei)
EDAD 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 sumas
LONGITUD
30 1.000 1.000
35 1.000 1.000
40 1.000 1.000
45 1.000 1.000
50 1.000 0.000 1.000
55 1.000 0.000 1.000
60 1.000 0.000 1.000
65 1.000 0.000 1.000
70 0.999 0.001 1.000
75 0.969 0.031 1.000
80 0.253 0.747 1.000
85 0.002 0.998 1.000
90 0.000 1.000 0.000 1.000
95 0.000 1.000 0.000 1.000
100 0.000 0.996 0.004 1.000
105 0.956 0.044 1.000
110 0.654 0.346 1.000
115 0.131 0.868 0.000 1.000
120 0.011 0.986 0.003 1.000
125 0.001 0.982 0.017 1.000
130 0.000 0.901 0.098 0.000 1.000
135 0.000 0.594 0.406 0.001 1.000
140 0.000 0.189 0.805 0.006 1.000
145 0.000 0.035 0.930 0.035 1.000
150 0.005 0.838 0.156 0.000 1.000
155 0.001 0.529 0.466 0.005 1.000
160 0.000 0.193 0.771 0.036 0.000 1.000
165 0.046 0.793 0.161 0.001 1.000
170 0.007 0.537 0.442 0.013 0.000 1.000
175 0.001 0.220 0.690 0.088 0.001 1.000
180 0.000 0.056 0.626 0.305 0.012 0.000 1.000
185 0.009 0.332 0.567 0.090 0.002 1.000
190 0.001 0.102 0.557 0.312 0.028 0.001 1.000
195 0.000 0.018 0.290 0.517 0.163 0.013 0.000 1.000
200 0.002 0.079 0.405 0.397 0.106 0.010 0.001 0.000 1.000
205 0.000 0.011 0.148 0.404 0.320 0.099 0.015 0.002 0.000 0.000 1.000
210 0.001 0.025 0.168 0.347 0.290 0.125 0.035 0.007 0.001 0.000 1.000
215 0.000 0.002 0.028 0.132 0.261 0.269 0.175 0.085 0.035 0.014 1.000
220 0.000 0.002 0.019 0.079 0.169 0.226 0.217 0.170 0.118 1.000
225 0.000 0.002 0.013 0.052 0.126 0.213 0.281 0.313 1.000
230 0.000 0.002 0.014 0.055 0.147 0.297 0.485 1.000
235 0.000 0.005 0.026 0.097 0.273 0.599 1.000
240 0.073 0.252 0.676 1.000
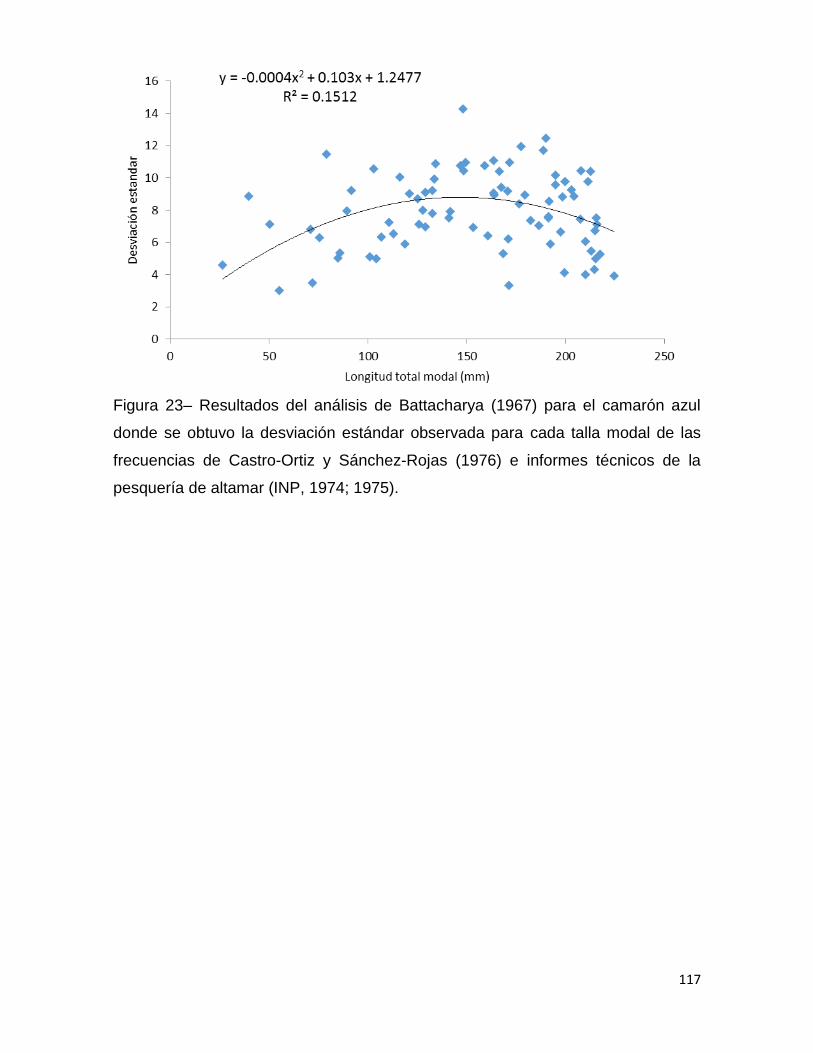
117
Figura 23– Resultados del análisis de Battacharya (1967) para el camarón azul
donde se obtuvo la desviación estándar observada para cada talla modal de las
frecuencias de Castro-Ortiz y Sánchez-Rojas (1976) e informes técnicos de la
pesquería de altamar (INP, 1974; 1975).

118
Figura 24 – Clave edad-longitud en términos de probabilidad para el camarón azul
Litopenaeus stylirostris.
6.1.3. Vectores de mortalidad natural en camarón del Pacífico mexicano
generados por modelo gnomónico (Caddy, 1996)
El ajuste de los intervalos gnomónicos a los datos observados resultó en siete
intervalos para las tres especies de camarón: café (Farfantepenaeus californiensis),
azul (Litopenaeus stylirostris) y blanco (L. vannamei) (tabla 38). Cada uno de los
intervalos presentó características intrínsecas a su etapa de desarrollo y hábitat.
Los primeros tres intervalos son planctónicos, pero son diferenciables entre sí por
las características propias de sus larvas. El resto de los cuatro intervalos son
bentónicos. De estos los primeros dos intervalos, postlarva y juvenil, se diferencian
por sus morfología y comportamiento; ambas presentes en las aguas estuarinas.
Los siguientes dos intervalos, el pre-adulto y adulto, habitan la plataforma
continental y se diferencian por su comportamiento y distribución a partir de la costa.
B) CAMARÓN AZUL (Litopenaeus stylirostris )
EDAD 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 sumas
LONGITUD
30 1.000 1.000
35 1.000 1.000
40 1.000 1.000
45 1.000 1.000
50 1.000 1.000
55 1.000 0.000 1.000
60 0.999 0.001 1.000
65 0.984 0.016 1.000
70 0.676 0.324 1.000
75 0.054 0.946 1.000
80 0.001 0.999 0.000 1.000
85 0.000 0.998 0.001 1.000
90 0.988 0.012 1.000
95 0.911 0.089 1.000
100 0.535 0.465 0.000 1.000
105 0.108 0.890 0.001 1.000
110 0.012 0.980 0.008 1.000
115 0.001 0.958 0.041 1.000
120 0.000 0.810 0.190 0.000 1.000
125 0.437 0.561 0.002 1.000
130 0.122 0.865 0.013 1.000
135 0.023 0.914 0.063 0.000 1.000
140 0.003 0.765 0.231 0.001 1.000
145 0.000 0.432 0.560 0.008 1.000
150 0.000 0.148 0.807 0.045 0.000 1.000
155 0.035 0.788 0.176 0.001 1.000
160 0.006 0.532 0.452 0.010 1.000
165 0.001 0.228 0.708 0.063 0.000 1.000
170 0.000 0.064 0.700 0.232 0.004 1.000
175 0.012 0.442 0.513 0.033 0.000 1.000
180 0.001 0.174 0.665 0.157 0.003 1.000
185 0.000 0.043 0.512 0.414 0.031 0.000 1.000
190 0.007 0.233 0.595 0.160 0.005 0.000 1.000
195 0.001 0.062 0.467 0.415 0.053 0.001 1.000
200 0.000 0.010 0.200 0.533 0.237 0.020 0.000 1.000
205 0.001 0.046 0.339 0.463 0.140 0.010 0.000 1.000
210 0.000 0.006 0.106 0.401 0.377 0.101 0.009 0.000 1.000
215 0.000 0.016 0.153 0.394 0.326 0.098 0.012 0.001 1.000
220 0.000 0.001 0.025 0.160 0.353 0.311 0.125 0.026 1.000
225 0.000 0.002 0.027 0.138 0.307 0.331 0.196 1.000
230 0.000 0.003 0.028 0.137 0.343 0.489 1.000
235 0.000 0.005 0.047 0.239 0.709 1.000
240 0.001 0.017 0.154 0.828 1.000

119
Las diferencias en los M-vectores se pueden explicar por diferencias en etapa de
desarrollo, como el comportamiento, uso de hábitat y vulnerabilidad a los
depredadores, específicos de cada etapa.
La especie más longeva fue el camarón café (565 días) así como las fecunda
(Fecundidad media “FM” = 550,000) en comparación con el camarón blanco (365
días / FM = 265,000), quedando el camarón azul como intermedio (411/días / FM =
350,000; Tabla 5).
La comparación de los M-vectores por cada especie con los valores de M
independientes de la literatura (fig. 25) mostró que los M-vectores obtenidos ajustan
a los rangos reportados, donde se observa también que ciertos valores extremos
pudieran representar un sesgo en su estimación (i.e. valor de M = 8.9 en camarón
café para adultos esta sobrestimado).
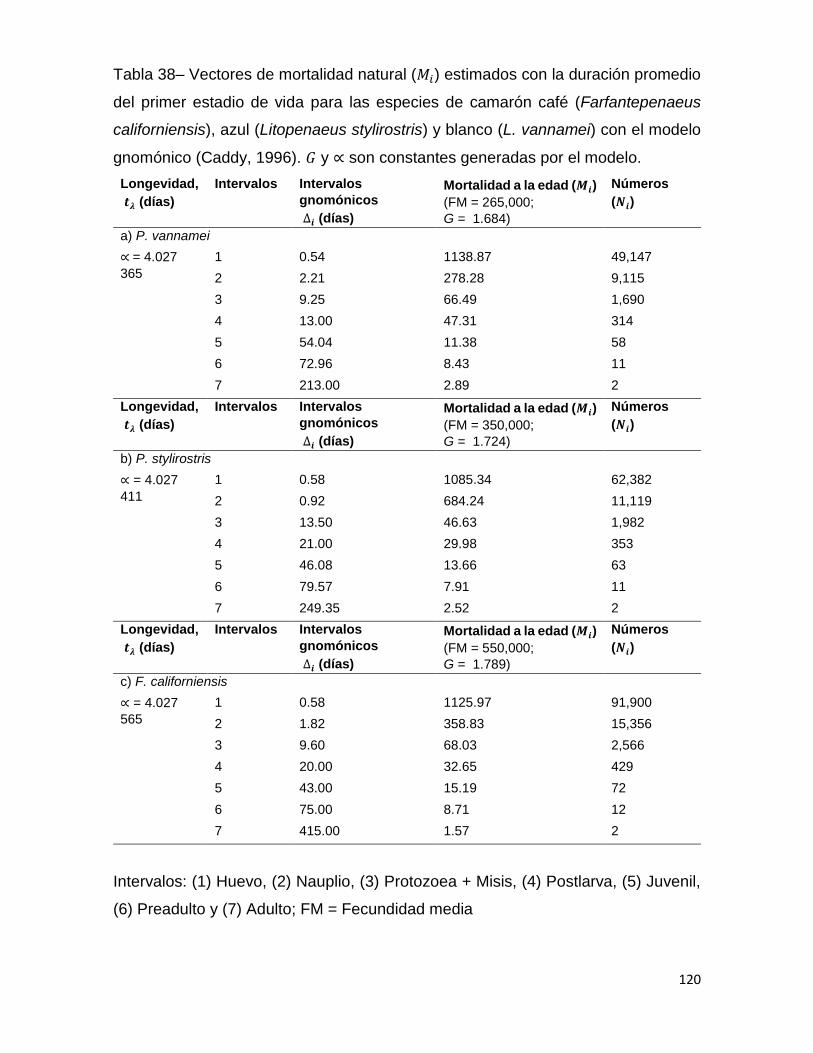
120
Tabla 38– Vectores de mortalidad natural (𝑀𝑖) estimados con la duración promedio
del primer estadio de vida para las especies de camarón café (Farfantepenaeus
californiensis), azul (Litopenaeus stylirostris) y blanco (L. vannamei) con el modelo
gnomónico (Caddy, 1996). 𝐺 y ∝ son constantes generadas por el modelo.
Longevidad,
𝒕𝝀 (días)
Intervalos Intervalos
gnomónicos
∆𝒊 (días)
Mortalidad a la edad (𝑴𝒊)
(FM = 265,000;
G = 1.684)
Números
(𝑵𝒊)
a) P. vannamei
∝ = 4.027
365
1 0.54 1138.87 49,147
2 2.21 278.28 9,115
3 9.25 66.49 1,690
4 13.00 47.31 314
5 54.04 11.38 58
6 72.96 8.43 11
7 213.00 2.89 2
Longevidad,
𝒕𝝀 (días)
Intervalos Intervalos
gnomónicos
∆𝒊 (días)
Mortalidad a la edad (𝑴𝒊)
(FM = 350,000;
G = 1.724)
Números
(𝑵𝒊)
b) P. stylirostris
∝ = 4.027
411
1 0.58 1085.34 62,382
2 0.92 684.24 11,119
3 13.50 46.63 1,982
4 21.00 29.98 353
5 46.08 13.66 63
6 79.57 7.91 11
7 249.35 2.52 2
Longevidad,
𝒕𝝀 (días)
Intervalos Intervalos
gnomónicos
∆𝒊 (días)
Mortalidad a la edad (𝑴𝒊)
(FM = 550,000;
G = 1.789)
Números
(𝑵𝒊)
c) F. californiensis
∝ = 4.027
565
1 0.58 1125.97 91,900
2 1.82 358.83 15,356
3 9.60 68.03 2,566
4 20.00 32.65 429
5 43.00 15.19 72
6 75.00 8.71 12
7 415.00 1.57 2
Intervalos: (1) Huevo, (2) Nauplio, (3) Protozoea + Misis, (4) Postlarva, (5) Juvenil,
(6) Preadulto y (7) Adulto; FM = Fecundidad media

121
Figura 25– Vectores de mortalidad natural estimados para el camarón café (a), azul
(b) y blanco (c) en círculos. La estimaciones independientes de la literatura están
con taches.

122
6.1.3. Variabilidad en M
Basados en los datos reportados en la literatura, la variabilidad de 𝑀𝑖
asociada con la variación en la duración de la etapa huevo fue analizada (tabla 6).
Debido a que la duración del primer intervalo gnomónico o etapa huevo es medido
en horas, se consideraron intervalos de 0.5 horas dentro del rango observado de la
literatura y se adecuaron las duraciones de las demás etapa de vida, resultando en
las estimaciones del valor medio, desviación estándar y coeficiente de
determinación para 𝑀𝑖 (tabla 39).
Tabla 39– Variaciones debido a diferentes duraciones en la etapa huevo para el
camarón blanco (L. vannamei), azul (L. stylirostris) y café (F. californiensis) usando
el modelo gnomónico; n, número de diferentes duraciones de la etapa huevo usadas
en la estimación; DE, desviación estándar; CV, coeficiente de la variación.
Intervalos
gnomónicos
F. californiensis (n=5) L. stylirostris (n=5) L. vannamei (n=13)
Media DE CV Media DE CV Media DE CV
1 Huevo 1724.35 103.65 0.06 1212.71 62.71 0.051 884.25 520.64 0.589
2 Nauplio 550.59 10.29 0.018 761.37 25.29 0.033 240.46 95.85 0.399
3 Protozoea + Mysis 104.24 0.1 0.001 51.89 0.062 0.001 57.35 22.77 0.397
4 Post-larva 50.03 0.05 0.001 33.36 0.04 0.001 40.81 16.2 0.397
5 Juvenil 23.27 0.02 0.001 15.2 0.018 0.001 9.82 3.89 0.396
6 Pre-adulto 13.34 0.01 0.001 8.8 0.011 0.001 7.27 2.88 0.396
7 Adulto 2.41 0 0.001 2.81 0.003 0.001 2.49 0.98 0.394

123
6.2. Parámetros del modelo pesquero
6.2.1. Estimación del esfuerzo pesquero por tipo flota
Los resultados del esfuerzo de altamar observado para el área de Sinaloa
sur vía satélite se observan en la tabla 40 y los estimados para la flota de pequeña
escala se observan en la tabla 41. Para la flota de pequeña escala se usó el informe
técnico del muestro de Muñoz et al., 2011 para determinar los lances diarios. El
problema fue al estimar el número de pescadores por mes en la temporada, el cual
fue muy por debajo de los números reportados en campo (“reales” tabla 41 y 42), lo
que hace pensar que la captura total reportada esta subestimada y esto va acorde
con la visita a los centros regionales pesqueros donde informaron que los
pescadores no reporta su captura. Por lo que se decidió trabajar con el número real
de pescadores para el modelo pesquero.
Tabla 40– Estimación del esfuerzo pesquero para la flota industrial para la
temporada 2014-15 en Sinaloa sur.
Esfuerzo pesquero industrial observado vía satélite en Sinaloa Sur
Sep Oct Nov Dic Ene Feb Mar
Número de barcos -- 189 95 140 165 203 135
Días efectivos pesca por barco/especie
Camarón azul -- 17.69 21.02 27.25 22.61 31.35 21.28
Camarón blanco -- 17.60 22.19 27.81 23.72 30.84 24.00
Camarón café -- 17.71 19.75 27.51 44.10 31.58 20.95
Promedio -- 18 21 28 23 31 22
Días efectivos pesca por flota/especie
Camarón azul -- 3,343 1,996 3,815 3,731 6,364 2,873
Camarón blanco -- 3,326 2,108 3,894 3,914 6,261 3,240
Camarón café -- 3,348 1,876 3,851 3,777 6,410 2,828
Días de pesca máximos -- 3,348 2,108 3,894 3,914 6,410 3,240

124
Tabla 41– Estimación del esfuerzo pesquero para la flota de pequeña escala para
la temporada 2014-15 en Sinaloa sur.
Sep Oct Nov Dic
Captura total (ton)a 424.53 183.48 40.99 30.29
Captura por muestreo (ton dia-1)b 0.0073 0.0140 0.0038 0.0033
Ton/lance por muestreob 0.000148 0.000093 0.000056 0.000044
Lances por muestreo (lance dia-1)b 49.41 150.96 67.14 73.56
Días efectivos de pesca 23 25 25 25
Lances totales al mes estimadosb 2,873,546 1,978,337 733,818 681,259
Pescadores/día estimadosb 2,528 524 437 370
Pescadores/día reales 6000 6000 4000 1000
Días efectivos de pesca mes-1 estimados 58,155 13,105 10,930 9,261
Días efectivos de pesca mes-1 reales 138,000 150,000 100,000 25,000 aBase de datos SAGARPA; b informe técnico Muñoz et al., 2011; reales: obtenidos de oficinas de
pesca locales y entrevistas a las cooperativas de la zona de Sinaloa sur.
Los valores de número de pescadores en la flota de pequeña escala se
observan en la tabla 42. Donde existen 51 cooperativas pesqueras con permiso
para explotar el camarón en las aguas de Sinaloa sur. Esto se refleja en un número
registrado de ≈4,000 pescadores y 2,000 pangas en total. Además, todas las
cooperativas (comunic. personal) reportan que contratan pescadores “libres” o sin
permiso (generalmente familiares y amigos cercanos) para trabajarle a las
cooperativas la extracción del recurso. Se estimó que alcanzan el 50% del número
total de socios, estimando un valor de 6,000 pescadores para el área.

125
Tabla 42– Número de pescadores para la temporada 2014-15 reportados en Sinaloa Sur.
Sitio # COOPERATIVA SOCIOS Pangas/cayucos
Mazatlán 1 S.C.P. AGROPECUARIA PESQUERA Y ACUICOLA PESCADORES DE MEDINA S.C.L
69 35
2 S.C.P.P. BARRAS DE PIAXTLA, S.C. DE R. L 15 8 3 S.C.P.P. CAMARONEROS DEL POZOLE S.C. DE R.L 52 26 4 S.C.P.P. EL PATOLE, S.C. DE R.L 15 8 5 S.C.P.P. LABRADAS DE BARRAS DE PIAXTLA, S.C. DE R. L 25 13 6 S.C.P.P. VETERANOS DE LA REVOLUCION MEXICANA, S.C.L 24 12
Huizache 1 S.C.P.P. EJIDO VILLA UNION S. DE R.L. DE C.V 134 67 2 S.C.P.P. FRENTE PESQUERO S. DE R.L. DE C.V 186 93 3 S.C.P.P. JOSE MARIA CANIZALES S.C. DE R.L. DE C.V 25 13 4 S.C.P.P. LA NUEVA SINALOENSE , S.C. DE R.L. DE C.V 114 57 5 S.C.P.P. LA SINALOENSE, S. DE R.L 123 62 6 S.C.P.P. PUNTA TIBURON S. DE R.L. DE C.V 16 8 7 S.C.P.P.R EJERCITO DEL TRABAJO S. DE R.L. DE C.V 98 49
Caimanero 1 S.C. OSTIONEROS DE CHAMETLA S.C. DE R.L. DE C.V 14 7 2 S.C.P.P. EJIDO CAJON OJO DE AGUA S. DE R.L. DE C.V 90 45 3 S.C.P.P. FRANCISCO I MADERO S.C. DE R.L 122 61 4 S.C.P.P. FRATERNIDAD PESQUERA, S. DE R.L.DE C.V 61 31 5 S.C.P.P. GENERAL ALVARO OBREGÓN S.C.L 230 115 6 S.C.P.P. INDEPENDENCIA DE AGUA VERDE S. DE R.L. DE C.V 62 31 7 S.C.P.P. INDIGENAS MARISMAS DE LAS CABRAS S.C. DE R.L 45 23 8 S.C.P.P. PESCADORES DE LA GUASIMA S.C. DE R.L. DE C.V 79 40 9 S.C.P.P. PESCADORES DE LA LAGUNA DEL CAIMANERO S.C DE R.L. DE C.V 73 37 10 S.C.P.P. PESCADORES DE LA PEDREGOZA S. DE R.L. DE C.V 83 42 11 S.C.P.P. PESCADORES DE LOS POZOS, S. DE R.L. DE C.V 76 38 12 S.C.P.P. PESCADORES DE POTRERILLOS S. DE R.L. DE C.V 102 51 13 S.C.P.P. PESCADORES DEL EJIDO, S. DE R.L. DE C.V 96 48 14 S.C.P.P. PESCADORES DEL IGUANITO S. DE R.L. DE C.V 77 39 15 S.C.P.P. PESCADORES RIBEREÑOS DE MATADERO S. DE R.L. DE C.V 210 105

126
16 S.C.P.P. UNIDOS DEL BALOARTE S.C. DE R.L. DE C.V 62 31 17 S.C.P.P. VENA EL QUELELE S.C. DE R.L 42 21 18 S.C.P.P. VICENTE LOMBARDO TOLETANO S. DE R.L. DE C.V 76 38 19 S.C.P.P.A. MARISMAS DE LAS CABRAS DE CHAMETLA SC DE R.L 24 12 20 S.C.P.P.R EJIDO TEODORO BELTRAN S. DE R.L. DE C.V 97 49 21 S.C.P.P.R. FRANCISCO JAVIER S.C. DE R.L 48 24 22 S.C.P.P.R. LA NUEVA FRANCISCO I. MADERO S.C DE R.L. DE C.V 29 15 23 S.C.P.P.R. PESCADORES DE VAZQUEZ MORENO S.C.L 62 31 24 S.C.P.P.R. PUNTA PODEROSA S. DE R.L. DE C.V 91 46
Teacapan-Chametla 1 S.C. DE ALTAMAR DE DIVERSAS ACTIVIDADES Y TURISMO LA BOCA DE TEACAPAN S.C. DE R.L DE C.V
40 20
2 S.C.P. Y C. DE PRODUCTOS DEL MAR DE TEACAPAN 165 83
3 S.C.P.P JOSE OSUNA CRESPO S. DE R.L. DE C.V 44 22 4 S.C.P.P. ATARRALLEROS CECEISTAS S.C. DE R.L 91 46 5 S.C.P.P. PALMITO DEL VERDE S RL DE CV 68 34 6 S.C.P.P. PESCADORES DE LA BRECHA S.C.L 53 27 7 S.C.P.P. PESCADORES DEL CAMICHIN S.C. DE R.L. DE C.V 46 23 8 S.C.P.P. PESCADORES DEL NANCHITO S.C.L. DE C.V 128 64
9 S.C.P.P. PESCADORES UNIDOS DE ESCUINAPA GENERAL LAZARO CARDENAS S. DE R.L. DE C.V
223 112
10 S.C.P.P. PESCADORES Y OSTIONEROS DE TEACAPAN S DE R.L D C.V 48 24 11 S.C.P.P. SUR DE SINALOA, S.DE R.L. DE C.V 87 44 12 S.C.P.P. TATA LAZARO CARDENAS S DE RL DE CV 87 44 13 S.C.P.P. TRIUNFO PESQUERO DE PALMILLAS S.C.L. DE C.V 105 53 14 S.C.P.P.R GERMINAL ARAMBURO CRISTERNA S.C.L. DE C.V 66 33
Total oficial 51 4,098 2,049
Total oficial + 50% de pescadores libres reportados 6000 ≈3,000

127
6.2.2. Bases de datos de captura por flotas
Los valores de la captura industrial provenientes de la base de capturas de
SAGARPA del puerto de Mazatlán (Esfuerzo industrial total) y de la base satelital
(Esfuerzo industrial depurado a arrastres en Sinaloa sur) para la temporada 2014-
15 se observan en la tabla 43. Cabe de mencionar que solo se usarán los valores
de la base satelital para los análisis.
Se reporta para la temporada 2013-14 una captura total para Sinaloa sur de
≈5,500 tons para la pesquería de camarón, con ≈4,900 tons para la flota industrial
y ≈680 tons para la flota de pequeña escala (tabla 43). La base industrial de todo el
puerto de Mazatlán (figura 26a) muestra la tendencia promedio por temporada de
pesca de camarón: la captura más alta en octubre (30%) con otro pico menor en
diciembre (24%) y con los valores mínimos en los últimos meses. La base industrial
estimada a partir del satélite para Sinaloa sur (figura 26a) muestra una tendencia
similar pero presenta tres picos de captura en octubre (22%), diciembre (24%) y
febrero (24%), disminuyendo al final de la temporada.
Finalmente la captura en aguas interiores fue un solo mes con altas capturas
(septiembre) y la disminución progresiva para los meses sucesivos (figura 26b). Los
valores de la captura de la flota de pequeña escala se localizan en la tabla 43. La
flota captura las especies de azul y blanco, y raramente el café. La especie que
domina las capturas (más de un 90%) es el camarón blanco y las demás especies
tienen una participación mínima y no se tomaron en cuenta para los análisis
sucesivos.

128
Tabla 43 - Capturas reportadas por base de datos generada en toneladas de peso vivo: a) Puerto de Mazatlán 2014-15; b)
Captura estimada vía satelital 2014-15; y c) Captura de la flota de pequeña escala 2014-15.
a) Captura industrial total reportada para el puerto de Mazatlán temporada 2014-15
Sep Oct Nov Dic Ene Feb Mar Total
Camarón Azul -- 1,679 319 497 89 224 215 3,023
Camarón Blanco -- 200 39 56 18 87 33 433
Camarón Café -- 1,446 1,561 2,194 468 1,252 1,118 8,039
Total -- 3,325 1,918 2,747 575 1,563 1,366 11,494
b) Captura industrial estimada vía satélite para el área de Sinaloa sur temporada 2014-15
Sep Oct Nov Dic Ene Feb Mar
Camarón Azul -- 582 195 274 92 178 104 1,425
Camarón Blanco -- 69 24 25 35 63 12 228
Camarón Café -- 437 340 890 348 909 299 3,223
Total -- 1,088 559 1,189 475 1,150 414 4,875
c) Captura de pequeña escala estimada para el área de Sinaloa sur temporada 2014-15
Sep Oct Nov Dic Ene Feb Mar
Camarón Azul
Camarón Blanco 425 183 41 30 679
Camarón Café
Total 425 183 41 30 679

129
Figura 26– Frecuencia relativa de las capturas por base de datos [a: base industrial
para el puerto de Mazatlán, Sinaloa y para Sinaloa sur (satélite); b: flota de pequeña
escala).
El comportamiento dinámico de la captura por especie para la base puerto de
Mazatlán se observa en la figura 27. Donde al inicio de la temporada se captura
camarón azul y su abundancia cae abruptamente en los meses sucesivos, siendo
sustituido por el camarón café. Finalmente el camarón con menos participación
proporcional en la captura industrial es el camarón blanco.
a)
b)

130
En la figura 28 se observa como está repartida la captura de aguas interiores
por municipio, donde el municipio de El Rosario (donde están las lagunas Huizache y
Caimanero) es donde se captura la mayor parte de los camarones al inicio de la
temporada. En los meses siguientes comienza a tener mayor participación el municipio
de Escuinapa (Chametla, Teacapán, Palmilla) y al final de los meses Mazatlán se
observa que participa con la mayor captura relativa (diciembre).
Figura 27- Captura industrial por especie para el puerto de Mazatlán (SAGARPA
2014-15).
Figura 28 – Captura de la flota de pequeña escala para la temporada 2014-15 en
Sinaloa sur (SAGARPA 2014-15).

131
6.3. Obtención de la estructura de longitudes de la captura
6.3.1. Estructura de longitudes de la captura para la flota de pequeña escala
La estructura de tallas para la flota de pequeña escala se observa en las figuras
29-31 donde solo se consideró a la especie dominante, el camarón blanco, para el
análisis (≈90% de las capturas).
Las frecuencias relativas de los muestreos de Huizache-Caimanero mostraron (fig.
29) en septiembre un rango de tallas de los 60 a los 180mm, con una moda a los
130mm y con las máximas frecuencias de tallas grandes 160-180mm. Este rango se
redujo en los meses de octubre (con modas entre los 120-130 y 150mm) y noviembre
(moda 130mm) a 80-160mm, y para diciembre la moda disminuyó a 110mm con un
aumento de frecuencias hacia las tallas pequeñas (hasta los 40mm) donde la
frecuencia de individuos de tallas grandes 135-170mm fue mínima.
La captura en número de individuos (fig. 30) fue máxima para el primer mes (hasta
14 millones de individuos) en septiembre con una moda principal entre los 65-80mm
y otra menor a los 130mm. En los siguientes meses se observa como la captura en
individuos cae drásticamente en todos los meses (<2 millones de individuos).
La captura por unidad de esfuerzo (CPUE: lance/día) en individuos (fig. 31) mostró
que el primer mes se registró la mayor CPUE la cual fue cayendo hasta diciembre.
Finalmente la CPUE estimada para el pescador y la flota en Sinaloa sur para la
temporada 2014-15 se observa en la tabla 44. Donde se ajustó a la captura observada
total reportada por SAGARPA al número de pescadores reales reportada en la tabla
41 y 42.
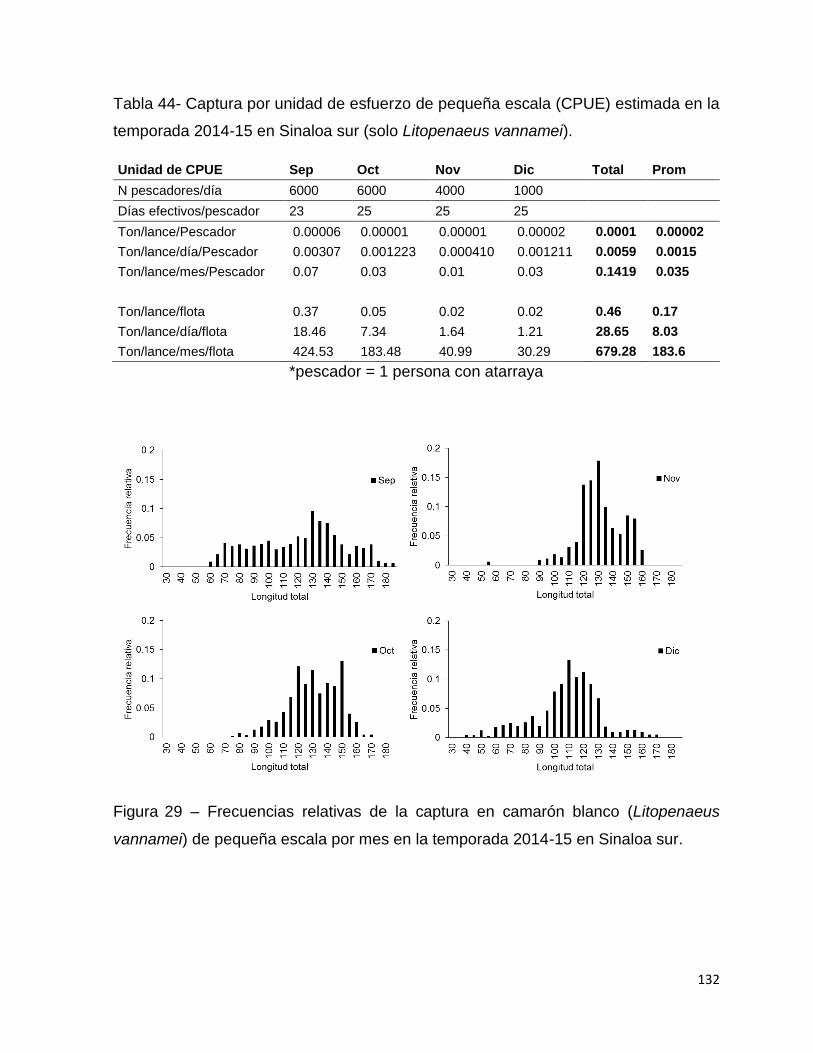
132
Tabla 44- Captura por unidad de esfuerzo de pequeña escala (CPUE) estimada en la
temporada 2014-15 en Sinaloa sur (solo Litopenaeus vannamei).
Unidad de CPUE Sep Oct Nov Dic Total Prom
N pescadores/día 6000 6000 4000 1000
Días efectivos/pescador 23 25 25 25
Ton/lance/Pescador 0.00006 0.00001 0.00001 0.00002 0.0001 0.00002
Ton/lance/día/Pescador 0.00307 0.001223 0.000410 0.001211 0.0059 0.0015
Ton/lance/mes/Pescador 0.07 0.03 0.01 0.03 0.1419 0.035
Ton/lance/flota 0.37 0.05 0.02 0.02 0.46 0.17
Ton/lance/día/flota 18.46 7.34 1.64 1.21 28.65 8.03
Ton/lance/mes/flota 424.53 183.48 40.99 30.29 679.28 183.6
*pescador = 1 persona con atarraya
Figura 29 – Frecuencias relativas de la captura en camarón blanco (Litopenaeus
vannamei) de pequeña escala por mes en la temporada 2014-15 en Sinaloa sur.

133
Figura 30– Individuos totales en la captura de pequeña escala en camarón blanco
(Litopenaeus vannamei) por mes en la temporada 2014-15 en Sinaloa sur.
Figura 31 – Captura por unidad de esfuerzo (lance/día) en individuos para la flota de
pequeña escala en camarón blanco (Litopenaeus vannamei) por mes en la
temporada 2014-15 en Sinaloa sur.

134
6.3.2. Estructura de longitudes de la captura para la flota industrial
Las frecuencias relativas de la estructura de la captura industrial obtenidas de
la maquiladora Mexican Shrimp Paradise se observan en las figuras 32-34. Se observa
que no están presentes las tallas de ≈90-130 Lt por el proceso de descarte de tallas
chicas en las embarcaciones para todas las especies.
Para el camarón blanco (fig.32) las frecuencias de tallas de la temporada
muestran modas alrededor de los 175mm en todos los meses y conforme avanza la
temporada a partir de diciembre se capturan las tallas de 200-220mm.
Para el camarón azul (fig.33) las frecuencias de tallas de la temporada de
octubre a enero la moda esta sobre los 180-185mm. En los meses de febrero y marzo
la moda incrementa a 185-190mm con mayor presencia de longitudes de hasta
210mm.
Para el camarón café (fig. 34) las frecuencias de tallas de la temporada
muestran modas con picos más obtusos que las demás especies (mayor presencia de
individuos por clase de longitud). Para noviembre y diciembre se muestran dos modas
una alrededor de los 150mm y otra a los 170mm. Para enero esta alrededor de los
150mm y en febrero y marzo cambia a los 170mm. Solo hasta marzo se observan
longitudes hasta los 205mm.

135
Figura 32– Frecuencias relativas por longitud total de maquila obtenidas de la
maquiladora Mexican Shrimp Paradise para camarón blanco (Litopenaeus vannamei)
para la temporada 2014-15.

136
Figura 33– Frecuencias relativas por longitud total de maquila obtenidas de la
maquiladora Mexican Shrimp Paradise para camarón azul (Litopenaeus stylirostris)
para la temporada 2014-15.

137
Figura 34– Frecuencias relativas por longitud total de maquila obtenidas de la
maquiladora Mexican Shrimp Paradise para camarón café (Farfantepenaeus
californiensis) para la temporada 2014-15.

138
Las capturas mensuales por individuos de la flota por especie para Sinaloa sur
se observan en la figuras 35-37. La especie con menos individuos representantes por
clase de longitud en la captura fue el camarón blanco con hasta 400,000 individuos
(figura 35). Sus meses más importantes fueron octubre y febrero.
La especie que inicio la temporada con más individuos capturados por clase de
longitud fue el azul alcanzando hasta 3 millones de individuos, después para octubre
y noviembre mantuvo el millón de individuos y para el resto de la temporada estuvo
por debajo de los 500 mil individuos, con un ligero aumento en febrero (figura 36).
Para el camarón café las capturas de sus individuos siempre se mantuvieron
por encima del millón y no fue hasta diciembre y en febrero donde sus capturas fueron
superiores a los 3 millones de individuos, y marzo presentó el menor número de
capturas (<1 millon por clase de longitud) (figura 37).
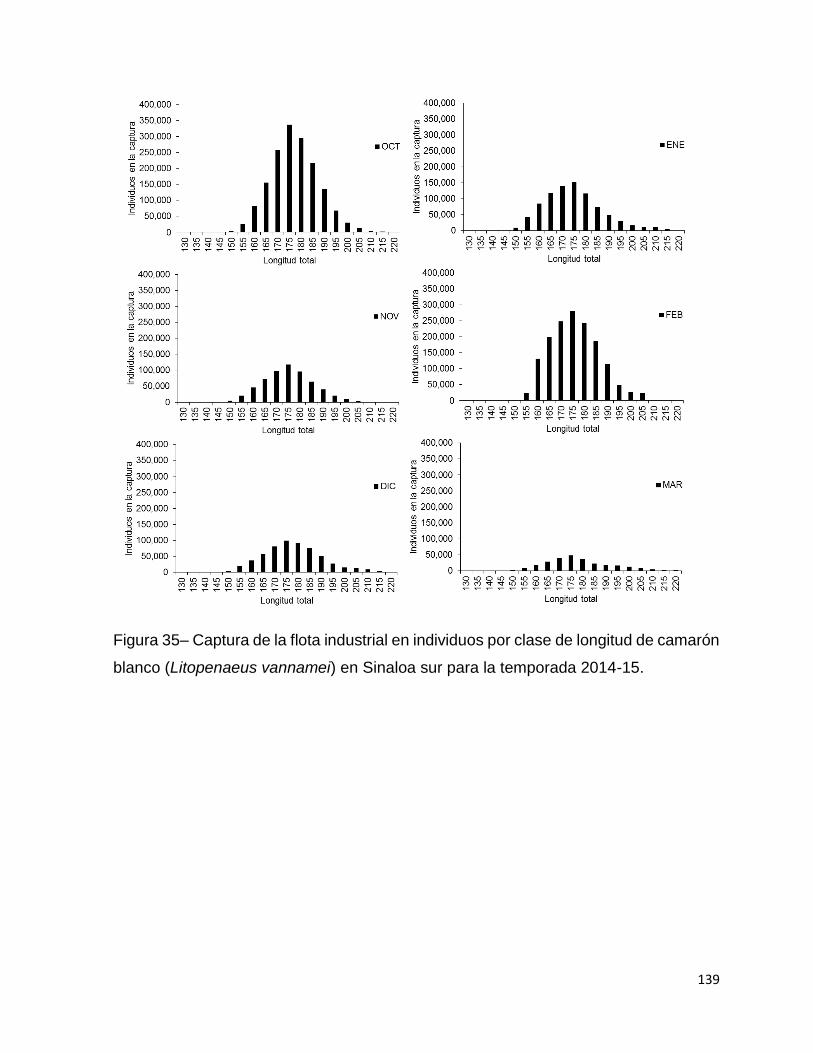
139
Figura 35– Captura de la flota industrial en individuos por clase de longitud de camarón
blanco (Litopenaeus vannamei) en Sinaloa sur para la temporada 2014-15.

140
Figura 36 - Captura de la flota industrial en individuos por clase de longitud de camarón
azul (Litopenaeus stylirostrisi) en Sinaloa sur para la temporada 2014-15.

141
Figura 37- Captura de la flota industrial en individuos por clase de longitud de camarón
café (Farfantepenaeus californiensis) para la flota que pesco en Sinaloa sur la
temporada 2014-15.

142
La captura por unidad de esfuerzo (CPUE) industrial estructurada por longitudes
de individuos en días de pesca por barco se muestra en las figuras 38-40. La tendencia
general por cada especie es la disminución de individuos en la captura conforme
avanza la temporada. Particularmente en el camarón café se presentaron incrementos
en la captura de individuos en los meses de diciembre y febrero.
La CPUE por barco y por flota en toneladas se observa en la tabla 45. La
especie con los menores rendimientos de CPUE fue el blanco y que dio los máximos
fue el café. En promedio para todas las especies para la temporada 2014-15 por
barco/día se captura 0.23 (DE 0.09) toneladas/día y 5.46 toneladas (DE 2.05)
barco/mes. Para la flota en promedio por día se captura 40 toneladas (DE 24.12)
flota/día y 831 (DE 341.56) toneladas flota/mes.
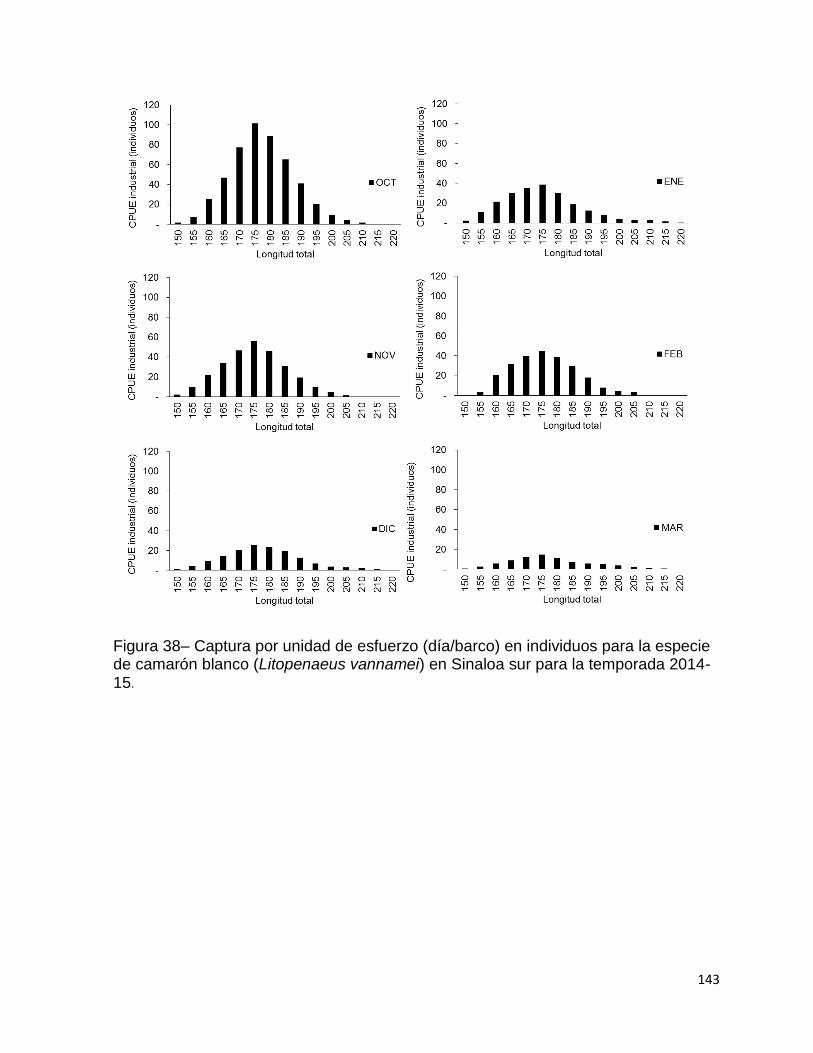
143
Figura 38– Captura por unidad de esfuerzo (día/barco) en individuos para la especie de camarón blanco (Litopenaeus vannamei) en Sinaloa sur para la temporada 2014-15.

144
Figura 39– Captura por unidad de esfuerzo (día/barco) en individuos para la especie
de camarón azul (Litopenaeus stylirostris) en Sinaloa sur para la temporada 2014-15.
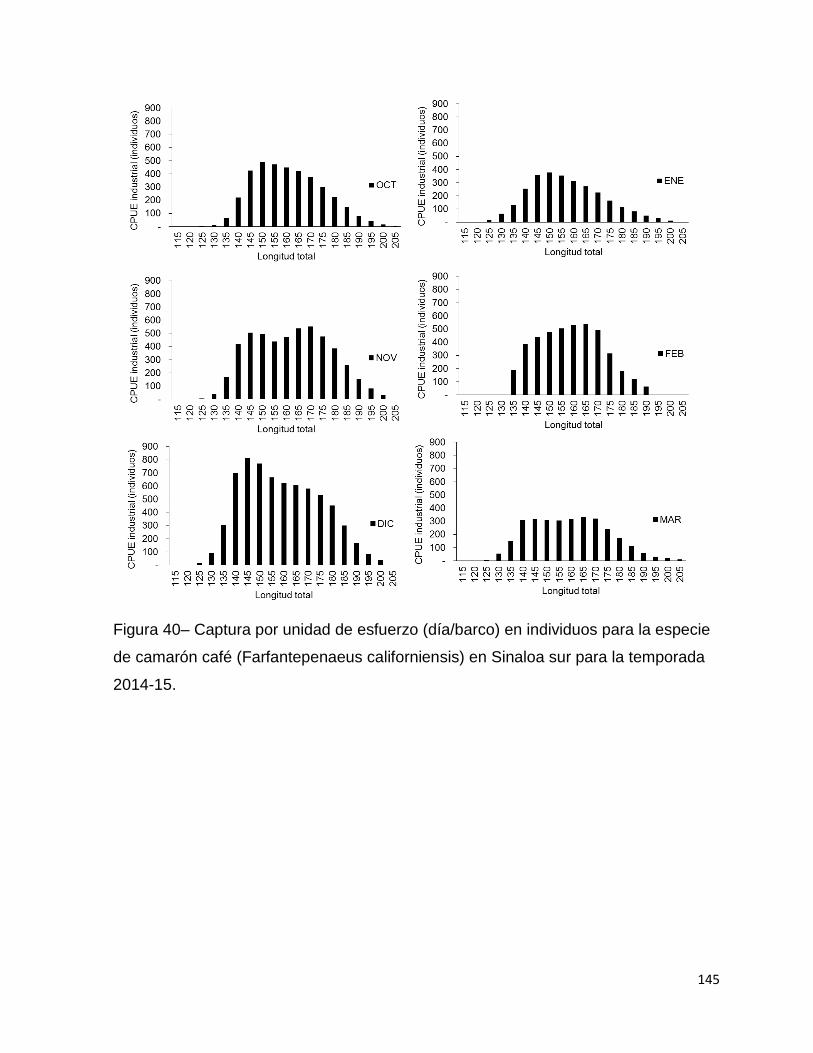
145
Figura 40– Captura por unidad de esfuerzo (día/barco) en individuos para la especie
de camarón café (Farfantepenaeus californiensis) en Sinaloa sur para la temporada
2014-15.

146
Tabla 45– Captura por unidad de esfuerzo industrial (CPUE) por día/barco/especie en
la temporada 2014-15 en Sinaloa sur.
a) Captura por unidad de esfuerzo (barco/día/tonelada)
Oct Nov Dic Ene Feb Mar Total Promedio
Blanco 0.02 0.01 0.01 0.01 0.01 0.00 0.06 0.0
Azul 0.17 0.10 0.07 0.02 0.03 0.04 0.43 0.1
Café 0.13 0.18 0.23 0.09 0.14 0.11 0.88 0.1
Total 0.33 0.29 0.31 0.13 0.18 0.15 1.38 0.2
b) Captura por unidad de esfuerzo (barco/mes/tonelada)
Blanco 0.36 0.25 0.18 0.21 0.31 0.09 1.40 0.2
Azul 3.07 2.17 2.00 0.58 0.86 0.87 9.54 1.6
Café 2.30 4.02 6.43 2.18 4.38 2.53 21.84 3.6
Total 5.73 6.44 8.61 2.98 5.55 3.49 32.79 5.5
c) Captura por unidad de esfuerzo (flota/día/tonelada)
Blanco 4 1 1 1 2 0 10 1.5
Azul 0 0 0 0 0 0 1 -
Café 69 24 25 35 63 12 227 38.0
Total 73 25 26 37 65 12 239 39.5
d) Captura por unidad de esfuerzo (flota/mes/tonelada)
Blanco 69 24 25 35 63 12 227 38.0
Azul 580 206 279 96 175 117 1,453 242.2
Café 434 382 900 360 888 342 3,307 551.0
Total 1,083 612 1,205 492 1,126 471 4,988 831.2
6.4. Modelo de capturabilidad a la edad (𝒒𝑳,𝒕) por especie
6.4.1. Capturabilidad a la longitud de la flota de pequeña escala
La capturabilidad estimada a la longitud 𝑞(𝐿,𝑡) para la flota de pequeña escala se
aplicó únicamente a la especie del camarón blanco (fig. 41). La capturabilidad a la
longitud promedio (��𝐿,𝑡) para la temporada 2014-15 en Sinaloa sur por lance de
atarraya fue de 5.37 E-07 (tabla 46). La 𝑞𝐿,𝑡 mensual por lance promedio se mantuvo
similar en los meses de septiembre (𝑞𝐿,𝑡 =5.85E-07) y octubre (𝑞𝐿,𝑡 =5.01 E-07), donde
para septiembre y octubre las tallas más vulnerables fueron de 150-175mm ó edades
4-5 meses; para noviembre se presentó un aumento (𝑞𝐿,𝑡 =1.30E-06) pero la
vulnerabilidad se redujo a tallas de 105-140mm ó edades 2-3 meses; los valores de
diciembre se estimaron por fuera del modelo de CATCHABILITY por la ausencia de
estructura de tallas para enero. Se basó en el CPUE del mismo mes y la dinámica
pesquera de la temporada. Así se estimó que en diciembre la abundancia poblacional

147
es la mínima (𝑞𝐿,𝑡 =6.10E-07) capturando tallas pequeñas de 50-130mm ó edades 1-
3. Finalmente de estos valores se estimaron las funciones de q mensuales (tabla 47)
y sus parámetros de ajuste se observan en la tabla 48. Del mismo modo se estimó una
función logística universal para la temporada (fig.41) cuyos parámetros se observan
en la tabla 48. Se observa que la vulnerabilidad del recurso aumenta
exponencialmente al alcanzar las tallas de la migración (90-140mm o edades de la 2
meses en adelante).
Figura 41– Función logística general de capturabilidad a la edad 𝑞(𝐿,𝑡) en camarón
blanco para la flota de pequeña escala en Sinaloa sur temporada 2014-15.

148
Tabla 46– Valores mensuales de la capturabilidad a la longitud del modelo de
CATCHABILITY en camarón blanco estimados para la flota de pequeña escala por
lance en Sinaloa sur para la temporada 2014-15.
Sep Oct Nov Dic
60 2.63E-07 1.06E-06 *
65 1.71E-07 8.58E-07 *
70 8.43E-08 7.47E-07 *
75 6.37E-09 3.63E-07 6.07E-07 *
80 1.10E-07 9.71E-08 6.87E-07 *
85 1.74E-07 3.63E-07 9.08E-07 *
90 3.20E-07 2.40E-08 5.19E-07 2.77E-07 *
95 3.05E-07 1.82E-07 5.69E-07 8.98E-07 *
100 1.84E-07 2.79E-07 4.79E-08 1.07E-06 *
105 7.27E-08 2.23E-07 1.11E-06 8.88E-07 *
110 2.35E-08 2.25E-07 1.78E-06 1.12E-06 *
115 6.97E-08 9.43E-08 2.11E-06 1.09E-06 *
120 1.25E-07 3.02E-07 2.14E-06 8.38E-07 *
125 2.19E-07 2.14E-07 2.01E-06 4.67E-07 *
130 4.85E-07 3.60E-07 2.06E-06 5.67E-07 *
135 2.94E-07 4.05E-07 3.04E-06 2.77E-07 *
140 4.96E-08 2.12E-07 1.02E-06 1.37E-07 *
145 3.35E-07 3.27E-07 7.71E-07 3.37E-07 *
150 1.05E-06 7.73E-07 1.20E-06 1.97E-07 *
155 6.60E-07 2.08E-06 6.24E-08 2.17E-07 *
160 2.42E-06 6.66E-07 1.09E-06 1.17E-07 *
165 2.17E-06 9.84E-07 1.06E-06 *
170 2.21E-06 1.85E-06 8.58E-07 *
175 2.56E-06 7.47E-07 *
180 4.35E-07 6.07E-07 *
185 2.73E-07
�� 5.80E-07 5.01E-07 1.30E-06 6.10E-07
max 2.56E-06 2.08E-06 3.04E-06 1.12E-06
min 6.37E-09 2.40E-08 4.79E-08 6.65E-08
Global 6.73E-07
* Valores estimados fuera del modelo

149
Tabla 47– Valores mensuales estimados de la capturabilidad a la longitud/edad de
camarón blanco para la flota de pequeña escala por lance en Sinaloa sur para la
temporada 2014-15.
Edad
meses L(t) media
Sep Oct Nov Dec
1 64 5.00E-08 8.43E-08
2 98 1.80E-07 1.87E-07 4.31E-07 8.43E-08
3 125 4.85E-07 3.85E-07 1.56E-06 8.28E-08
4 147 1.02E-06 6.57E-07 1.56E-06 5.44E-08
5 164 1.73E-06 9.69E-07 1.56E-06 9.31E-09
6 178 2.51E-06 1.28E-06 1.23E-09
7 188 3.22E-06
Tabla 48– Parámetros de la función logística de la capturabilidad a la longitud de
camarón blanco para la flota de pequeña escala en Sinaloa sur para la temporada
2014-15.
Parámetros Sep Oct Nov Dic Fx
general (Log)
a 0.000007 0.000004 1.56E-06 8.43E-08 -7.29231
b 0.038060 0.028649 0.2848 23.59 -0.01508
c -7.4204 -5.78973 -28.9126 -0.16 3.84848
Varianza
explicada 0.810 0.66 0.053 0.50 0.3139
*Intervalo de confianza al 95% (𝛼 = 0.050).
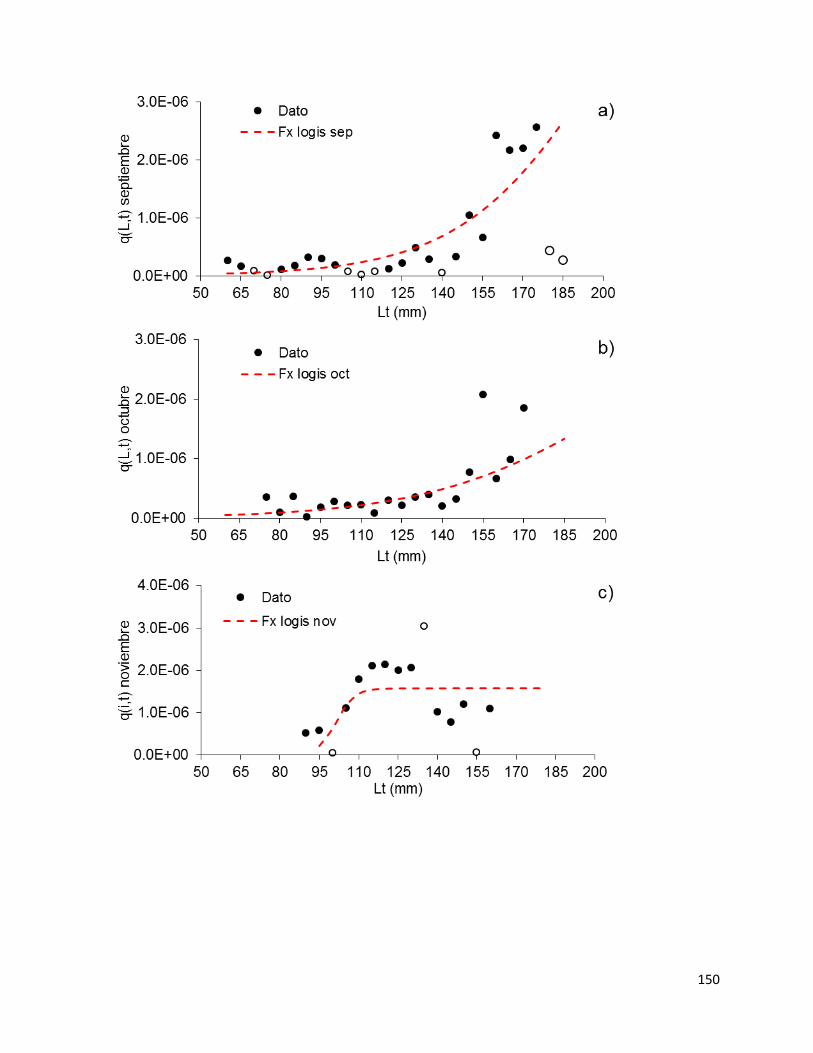
150

151
Figura 42 - Patrón mensual de capturabilidad por longitud de camarón blanco para la
flota de pequeña escala en Sinaloa sur temporada 2014-15 (a-d). El resumen de los
patrones de 𝑞(𝐿,𝑡) mensual se observa en (e). Los círculos vacíos representan valores
extremos eliminados para fines del ajuste.

152
6.4.2. Capturabilidad a la longitud de la flota industrial
a) Capturabilidad a la longitud del camarón blanco
La capturabilidad a la longitud promedio (��𝐿,𝑡) del camarón blanco para la flota
industrial para la temporada 2014-15 en Sinaloa sur por día efectivo/barco fue de 𝑞𝐿,𝑡 =
0.0002 (tabla 49).
La función logística general estimada para la capturabilidad a la longitud del
camarón blanco para la flota industrial en la temporada 2014-15 se observa en la figura
43 y sus parámetros en la tabla 51. Se puede notar que la vulnerabilidad a la pesca
aumenta hacia el componente reproductor.
La 𝑞𝐿,𝑡 mensual (fig. 44 Y tabla 50).presentó sus valores más altos en los meses
de octubre (𝑞𝐿,𝑡 =0.00036) y noviembre (𝑞𝐿,𝑡 =0.00032); pero la vulnerabilidad por talla
presentó diferencias donde para octubre fueron los reproductores de tallas 190-
220mm≈edades 7 en adelante; y en noviembre fueron tallas de reclutas a la pesquería
(150 – 160mm≈edades 4 a 5). La 𝑞𝐿,𝑡 cayó en diciembre ((𝑞𝐿,𝑡 =0.00011) y se
presentan altas vulnerabilidades en tanto tallas de reclutas como de reproductores,
pero si se observa la figura 29c la tendencia es hacia el reclutamiento a la pesquería.
Para los siguientes meses (ene-mar) se estabiliza la 𝑞𝐿,𝑡 en valores de 𝑞𝐿,𝑡 =0.00012-
0.00013. En enero la mayor vulnerabilidad fue para las tallas más grandes y en febrero-
marzo se invirtió hacia tallas menores
Los valores estimados de 𝑞(𝐿,𝑡), para ambas flotas que explotan el camarón
blanco (fig.45), lo que representaría la pesquería de camarón blanco en Sinaloa sur
(figura 30), se observa que la vulnerabilidad del camarón blanco a la pesca aumenta
desde que alcanza sus tallas de migración, siendo explotadas las tallas 150-170mm
por ambas flotas y la vulnerabilidad continua aumentando hacia tallas de organismos
reproductores (>150mm).

153
Figura 43– Función logística general de capturabilidad a la edad 𝑞(𝐿,𝑡) en camarón
blanco para la flota industrial en Sinaloa sur temporada 2014-15.

154
Tabla 49 - Valores mensuales de la capturabilidad a la longitud del modelo de
CATCHABILITY en camarón blanco estimados para la flota industrial por día efectivo
de pesca/barco en Sinaloa sur para la temporada 2014-15.
Oct Nov Dic Ene Feb Mar
150 0.00010 0.00040 0.00018 0.00001 0.00026 *
155 0.00015 0.00038 0.00014 0.00003 0.00019 0.00021 *
160 0.00017 0.00038 0.00012 0.00003 0.00017 0.00018 *
165 0.00018 0.00035 0.00009 0.00005 0.00018 0.00015 *
170 0.00020 0.00028 0.00004 0.00009 0.00020 0.00013 *
175 0.00023 0.00019 0.00002 0.00013 0.00024 0.00011 *
180 0.00022 0.00019 0.00001 0.00011 0.00020 0.00010 *
185 0.00021 0.00015 0.00002 0.00003 0.00011 0.00009 *
190 0.00020 0.00023 0.00004 0.00002 0.00003 0.00008 *
195 0.00023 0.00006 0.00002 0.00002 0.00002 0.00007 *
200 0.00034 0.00045 0.00007 0.00011 0.00009 0.00006 *
205 0.00107 0.00084 0.00012 0.00055 0.00005 0.00006 *
210 0.00078 0.00022 0.00033
215 0.00063 0.00027 0.00021
220 0.00074 0.00030 0.00030
�� 0.00036 0.00032 0.00011 0.00013 0.00013 0.00012
max 0.00107 0.00084 0.00030 0.00055 0.00024 0.00026
min 0.00010 0.00006 0.00001 0.00001 0.00002 0.00006
Global 0.00020
* Valores estimados fuera del modelo.

155
Tabla 50- Valores mensuales estimados de la capturabilidad a la longitud/edad de
camarón blanco para la flota industrial por día efectivo/barco en Sinaloa sur para la
temporada 2014-15.
Edad Lt media Oct Nov Dic Ene Feb Mar**
1 64 -- -- -- -- -- --
2 98 -- -- -- -- -- --
3 125 -- -- -- -- -- --
4 147 1.47E-04 1.61E-04 4.22E-04 2.63E-05 4.37E-04
5 164 1.54E-04 1.62E-04 7.07E-05 3.41E-05 2.40E-04 8.42E-05
6 178 1.67E-04 1.65E-04 6.15E-05 4.70E-05 1.52E-04 5.55E-05
7 188 1.87E-04 1.73E-04 6.06E-05 6.68E-05 1.04E-04 4.94E-05
8 197 2.17E-04 1.92E-04 6.05E-05 9.51E-05 7.65E-05 4.74E-05
9 204 2.58E-04 2.30E-04 6.04E-05 1.33E-04 5.93E-05 4.67E-05
10 209 3.09E-04 3.00E-04 6.04E-05 1.80E-04 4.82E-05 4.63E-05
11 213 3.70E-04 6.04E-05 2.33E-04 4.61E-05
12 217 4.37E-04 6.04E-05 2.91E-04 4.60E-05
13 220 5.07E-04 6.04E-05 3.51E-04 4.60E-05
*Intervalo de confianza al 95% (𝛼 = 0.050).** valores estimados fuera del modelo
Tabla 51 - Parámetros de la función logística de la capturabilidad a la longitud de
camarón blanco para la flota industrial en Sinaloa sur para la temporada 2014-15.
Parámetros Oct Nov Dic Ene Feb
a -3.84458 -3.79266 -4.218 -4.74386 -143.943
c 13.56235 25.23977 --21.967 8.07681 -4.313
b -0.05362 -0.10841 0.1591 -0.03230 0.004
Varianza
explicada 0.857 0.329 0.346 0.473 0.503
Mar Fx general
(Log)
a 0.00010 -3.97650
c 18.70586 28.88170
b -0.09272 -0.12430
Varianza
explicada 0.48 0.14

156
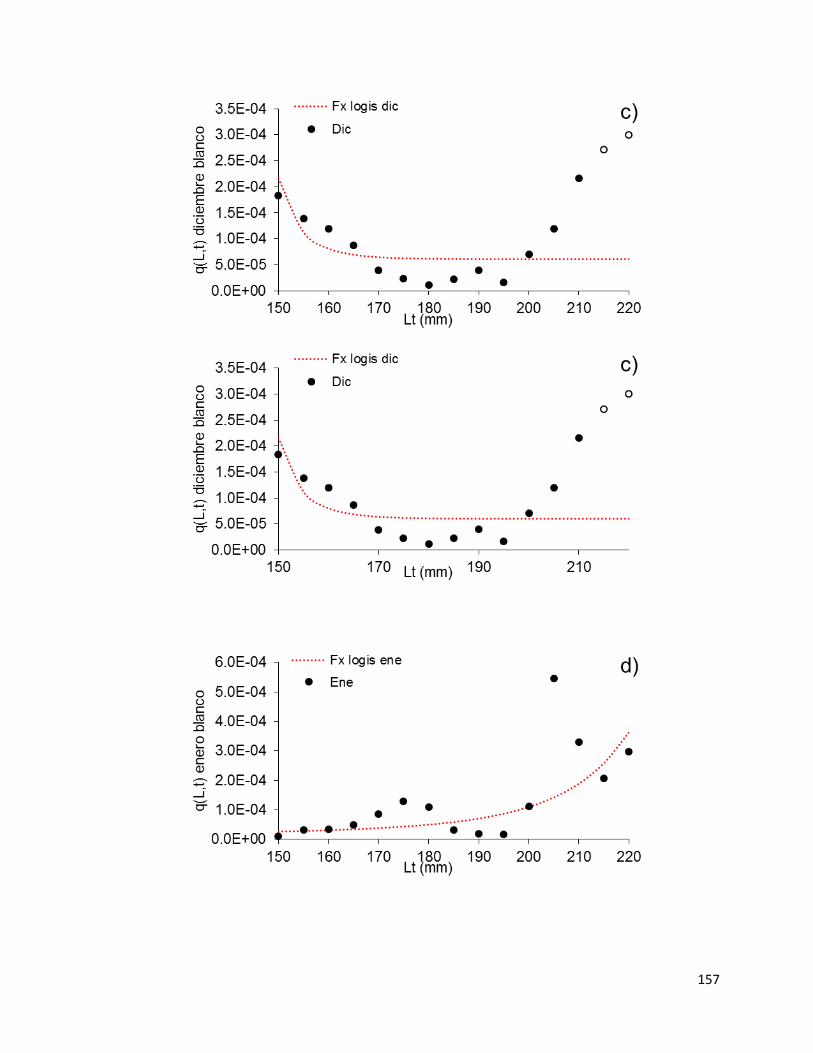
157
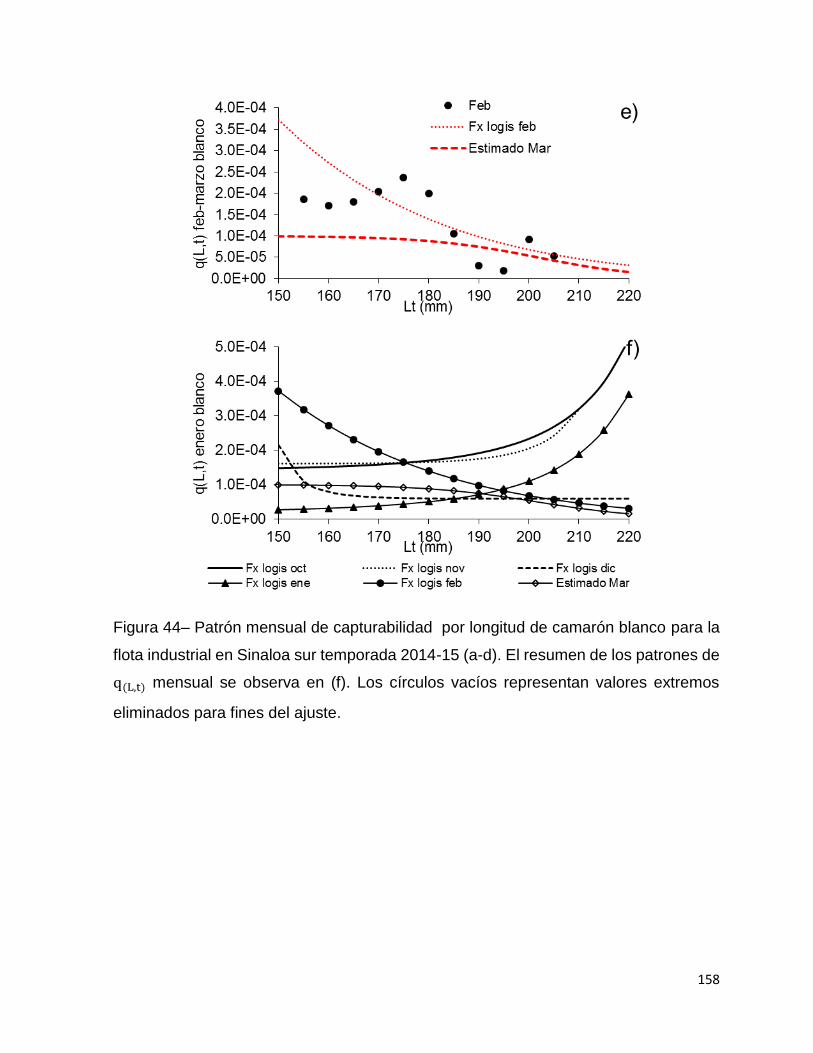
158
Figura 44– Patrón mensual de capturabilidad por longitud de camarón blanco para la
flota industrial en Sinaloa sur temporada 2014-15 (a-d). El resumen de los patrones de
q(L,t) mensual se observa en (f). Los círculos vacíos representan valores extremos
eliminados para fines del ajuste.

159
Figura 45– Vulnerabilidad del recurso en función de la capturabilidad por cada longitud
para la pesquería de camarón blanco en Sinaloa sur temporada 2014-15.
b2) Capturabilidad a la longitud del camarón azul
La capturabilidad a la longitud promedio (��𝐿,𝑡) mensual del camarón azul para la
flota industrial para la temporada 2014-15 en Sinaloa sur por día efectivo/barco fue de
𝑞𝐿,𝑡 = 0.00016 (tabla 52).
La función logística general estimada para la capturabilidad a la longitud del
camarón azul para la flota industrial en la temporada 2014-15 se observa en la figura
46 y sus parámetros en la tabla 54. Se puede notar que la vulnerabilidad a la pesca
aumenta hacia el componente reproductor.
La 𝑞𝐿,𝑡 mensual (fig. 47) presentó sus valores más altos en los meses de octubre
(𝑞𝐿,𝑡 =0.00020), noviembre (𝑞𝐿,𝑡 =0.00018) y el pico más alto de diciembre
(𝑞𝐿,𝑡 =0.00023); además la vulnerabilidad por talla presentó diferencias mensuales
donde en octubre fueron los reproductores de tallas 170-215mm≈edades 6 en
adelante; y en noviembre fueron tanto tallas de reclutas a la pesquería (155 – 165mm ≈
edades 5 a 6 meses) y otro aumento a tallas de reproductores (195-200mm ≈ edades
8-9 meses), pero la tendencia fue hacia tallas de reclutas (fig.47b). Para diciembre de
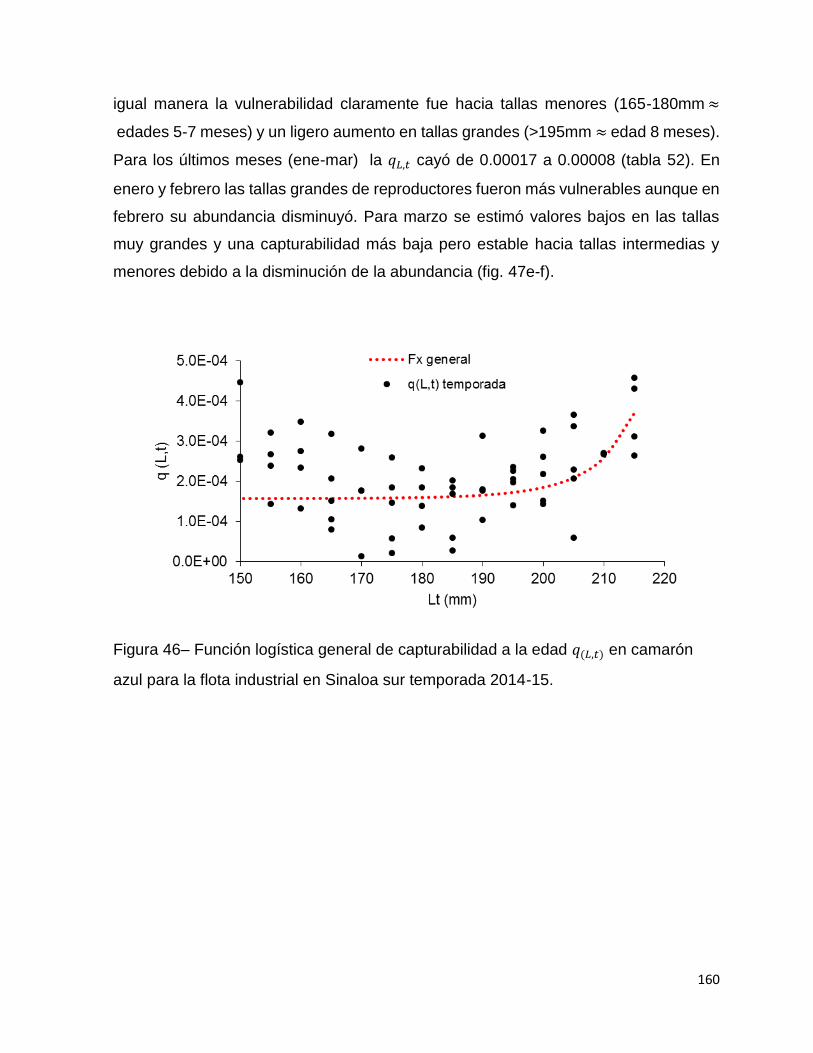
160
igual manera la vulnerabilidad claramente fue hacia tallas menores (165-180mm ≈
edades 5-7 meses) y un ligero aumento en tallas grandes (>195mm ≈ edad 8 meses).
Para los últimos meses (ene-mar) la 𝑞𝐿,𝑡 cayó de 0.00017 a 0.00008 (tabla 52). En
enero y febrero las tallas grandes de reproductores fueron más vulnerables aunque en
febrero su abundancia disminuyó. Para marzo se estimó valores bajos en las tallas
muy grandes y una capturabilidad más baja pero estable hacia tallas intermedias y
menores debido a la disminución de la abundancia (fig. 47e-f).
Figura 46– Función logística general de capturabilidad a la edad 𝑞(𝐿,𝑡) en camarón
azul para la flota industrial en Sinaloa sur temporada 2014-15.

161
Tabla 52- Valores mensuales de la capturabilidad a la longitud del modelo de
CATCHABILITY en camarón azul estimados para la flota industrial por día efectivo de
pesca/barco en Sinaloa sur para la temporada 2014-15.
Oct Nov Dic Ene Feb
150
155 0.00014 0.00024
160 0.00013 0.00023
165 0.00015 0.00021 0.00032 0.00010 0.00008
170 0.00018 0.00018 0.00028 0.00001 0.00001
175 0.00019 0.00015 0.00026 0.00006 0.00002
180 0.00018 0.00008 0.00023 0.00014 0.00000
185 0.00017 0.00006 0.00019 0.00020 0.00003
190 0.00018 0.00010 0.00018 0.00031 0.00039
195 0.00023 0.00024 0.00020 0.00021 0.00014
200 0.00026 0.00033 0.00022 0.00015 0.00014
205 0.00021
0.00023 0.00037 0.00006
210
215 0.00043
�� 0.00020 0.00018 0.00023 0.00017 0.00010
Max 0.00043 0.00033 0.00032 0.00037 0.00039
Min 0.00013 0.00006 0.00018 0.00001 0.00000
Global 0.00016
* Valor de marzo fue estimado sin el programa CATCHABILITY.

162
Tabla 53- Valores mensuales estimados de la capturabilidad a la longitud/edad de
camarón azul para la flota industrial por día efectivo/barco en Sinaloa sur para la
temporada 2014-15.
Edad Lt media Oct Nov Dic Ene Feb Mar*
1 57 -- -- -- -- -- --
2 88 -- -- -- -- -- --
3 114 -- -- -- -- -- --
4 135 7.71E-05 2.80E-04 3.09E-04 4.30E-05 1.03E-04 7.34E-05
5 153 1.09E-04 2.56E-04 2.90E-04 7.73E-05 1.03E-04 7.34E-05
6 168 1.44E-04 2.22E-04 2.73E-04 1.21E-04 1.03E-04 7.34E-05
7 181 1.83E-04 1.82E-04 2.57E-04 1.68E-04 1.03E-04 7.34E-05
8 191 2.24E-04 1.44E-04 2.44E-04 2.14E-04 1.03E-04 7.33E-05
9 200 2.65E-04 1.13E-04 2.32E-04 2.54E-04 1.03E-04 7.15E-05
10 207 3.04E-04 8.95E-05 2.22E-04 2.88E-04 3.84E-07 5.72E-05
11 214 3.42E-04 7.20E-05 2.13E-04 3.15E-04 1.26E-13 2.38E-05
12 219 3.77E-04 5.93E-05 2.06E-04 3.37E-04 4.77E-19 6.10E-06
13 223 4.09E-04 5.01E-05 2.00E-04 3.54E-04 1.40E-23 1.62E-06
14 227 4.38E-04
1.95E-04
* Valor de marzo fue estimado sin el programa CATCHABILITY.
Tabla 54 - Parámetros de la función logística de la capturabilidad a la longitud de
camarón azul para la flota industrial en Sinaloa sur para la temporada 2014-15.
Parámetros Oct Nov Dic Ene Feb
a 0.236061 0.000299 0.000401 0.000515 0.0001
c -10.5881 9.154840 3.104506 -7.32747 497.675
b 0.0190 -0.051182 -0.013911 0.036948 -2.426
Varianza
explicada 0.739 0.343 0.275 0.116 0.09
Mar Fx general
(Log)
a 0.000083 -3.80536
c 66.21178 -0.11601
b -0.31516 27.16709
Varianza
explicada 0.145 0.06
*Intervalo de confianza al 95% (𝛼 = 0.050).

163

164
Figura 47- Patrón mensual de capturabilidad por longitud de camarón azul para la flota
industrial en Sinaloa sur temporada 2014-15 (a-d). El resumen de los patrones de 𝑞(𝐿,𝑡)
mensual se observa en (f). Los círculos vacíos representan valores extremos
eliminados para fines del ajuste.

165
b3) Camarón café
La capturabilidad a la longitud promedio (��𝐿,𝑡) del camarón café para la flota
industrial para la temporada 2014-15 en Sinaloa sur por día efectivo/barco fue de 𝑞𝐿,𝑡 =
0.0002 (tabla 55-56) siendo la más alta de las tres especies.
La función logística general estimada para la capturabilidad a la longitud del
camarón azul para la flota industrial en la temporada 2014-15 se observa en la figura
48 y sus parámetros en la tabla 57. Se puede notar que la vulnerabilidad a la pesca
aumenta hacia el componente reproductor.
El caso del camarón café fue presento diferencias respecto a las otras dos
especies, debido a que siempre hubo una señal de reclutamiento hacia aguas marinas,
a continuación se describe su tendencia mensual. La 𝑞𝐿,𝑡 mensual (fig. 49) presentó
sus valores más altos en los meses de diciembre (𝑞𝐿,𝑡 =0.00028), noviembre
(𝑞𝐿,𝑡 =0.00021), febrero (𝑞𝐿,𝑡 =0.00021) y octubre (𝑞𝐿,𝑡 =0.00020); pero la
vulnerabilidad por talla presentó diferencias donde para octubre, noviembre y enero
(𝑞𝐿,𝑡 =0.000121) fueron la talles grandes o reproductores (tallas >150 ≈ edades 6
meses en adelante); para noviembre, febrero y marzo (𝑞𝐿,𝑡 =0.0001) la vulnerabilidad
de las tallas fue hacia los reclutas a la pesquería (130 – 150mm ≈ edades 5 y 7
meses).

166
Figura 48– Función logística general de capturabilidad a la edad 𝑞(𝐿,𝑡) en camarón
café para la flota industrial en Sinaloa sur temporada 2014-15.
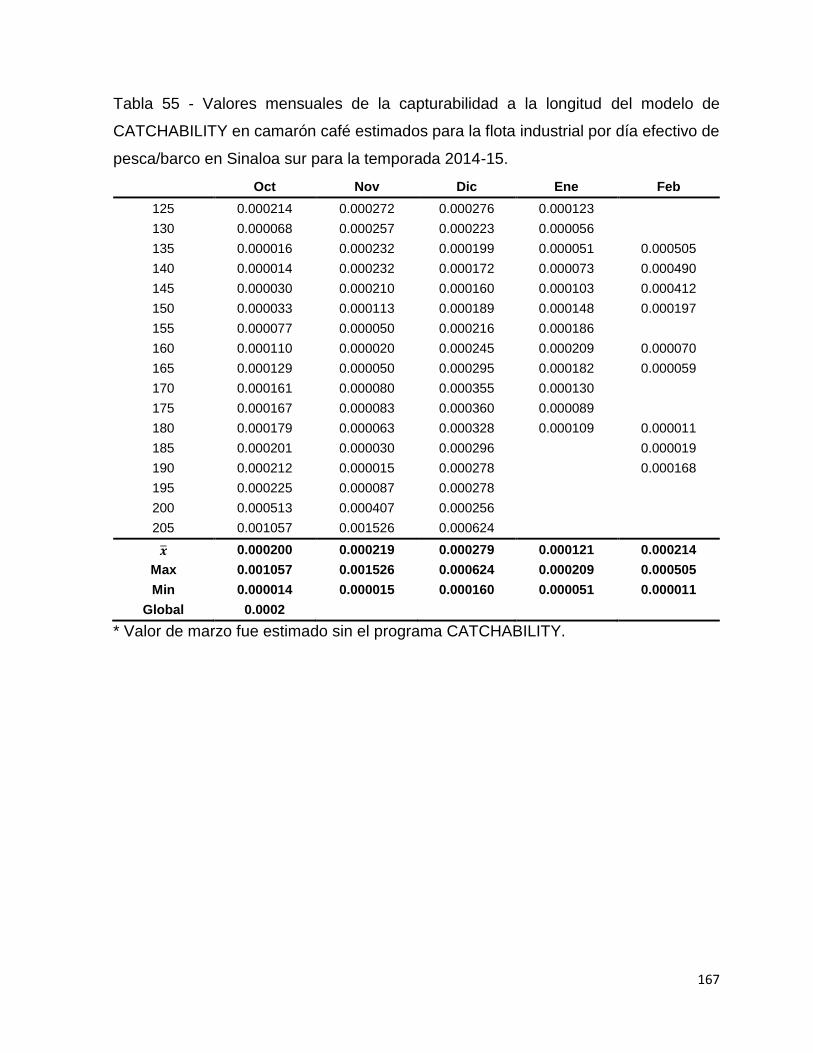
167
Tabla 55 - Valores mensuales de la capturabilidad a la longitud del modelo de
CATCHABILITY en camarón café estimados para la flota industrial por día efectivo de
pesca/barco en Sinaloa sur para la temporada 2014-15.
Oct Nov Dic Ene Feb
125 0.000214 0.000272 0.000276 0.000123
130 0.000068 0.000257 0.000223 0.000056
135 0.000016 0.000232 0.000199 0.000051 0.000505
140 0.000014 0.000232 0.000172 0.000073 0.000490
145 0.000030 0.000210 0.000160 0.000103 0.000412
150 0.000033 0.000113 0.000189 0.000148 0.000197
155 0.000077 0.000050 0.000216 0.000186
160 0.000110 0.000020 0.000245 0.000209 0.000070
165 0.000129 0.000050 0.000295 0.000182 0.000059
170 0.000161 0.000080 0.000355 0.000130
175 0.000167 0.000083 0.000360 0.000089
180 0.000179 0.000063 0.000328 0.000109 0.000011
185 0.000201 0.000030 0.000296 0.000019
190 0.000212 0.000015 0.000278 0.000168
195 0.000225 0.000087 0.000278
200 0.000513 0.000407 0.000256
205 0.001057 0.001526 0.000624
�� 0.000200 0.000219 0.000279 0.000121 0.000214
Max 0.001057 0.001526 0.000624 0.000209 0.000505
Min 0.000014 0.000015 0.000160 0.000051 0.000011
Global 0.0002
* Valor de marzo fue estimado sin el programa CATCHABILITY.

168
Tabla 56 - Valores de la capturabilidad a la longitud de camarón café obtenidos de
individuo/días/flota para la flota industrial en Sinaloa sur para la temporada 2014-15.
Edad (meses) Lt media Oct Nov Dic Ene Feb Mar*
1 51 -- -- -- -- -- --
2 75 -- -- -- -- -- --
3 95 -- -- -- -- -- --
4 114 3.14E-06 3.12E-04 1.04E-04 6.57E-05 1.15E-04
5 130 1.19E-05 2.56E-04 1.62E-04 8.57E-05 5.48E-04 1.02E-04
6 143 3.51E-05 1.78E-04 2.12E-04 1.05E-04 4.15E-04 7.65E-05
7 155 7.81E-05 1.08E-04 2.48E-04 1.23E-04 1.16E-04 4.65E-05
8 166 1.30E-04 6.17E-05 2.72E-04 1.39E-04 1.77E-05 2.47E-05
9 175 1.72E-04 3.55E-05 2.87E-04 1.52E-04 2.95E-06 1.28E-05
10 183 1.99E-04 2.12E-05 2.96E-04 1.64E-04 6.07E-07 6.83E-06
11 190 2.13E-04 1.33E-05 3.02E-04 1.73E-04 1.52E-07 3.87E-06
12 196 2.22E-04 8.81E-06 3.06E-04 1.80E-04 4.54E-08 2.33E-06
13 202 2.26E-04 6.12E-06 3.08E-04 1.87E-04 1.50E-06
14 207 2.29E-04 4.44E-06 3.10E-04 1.92E-04 1.01E-06
15 211 7.18E-07
16 214 5.32E-07
17 217 4.10E-07
18 220 3.26E-07
* Valor de marzo fue estimado sin el programa CATCHABILITY.

169
Tabla 57 - Parámetros de la función logística de la capturabilidad a la longitud de
camarón café para la flota industrial en Sinaloa sur para la temporada 2014-15.
Parámetros Oct Nov Dic Ene Feb
a 0.000235 0.000354 0.000284 0.000248 0.00056
c -14.0899 9.997721 -3.40800 -3.75042 29.28
b 0.0862 -0.069496 0.03069 0.02393 -0.1970
Varianza
explicada 0.942 0.815 0.230 0.199 0.895
Mar Fx general
(Log)
a 0.00012 -3.95796
c 12.6725 12.10307
b -0.08445 -0.04907
Varianza
explicada 0.98 0.087
*Intervalo de confianza al 95% (𝛼 = 0.050).
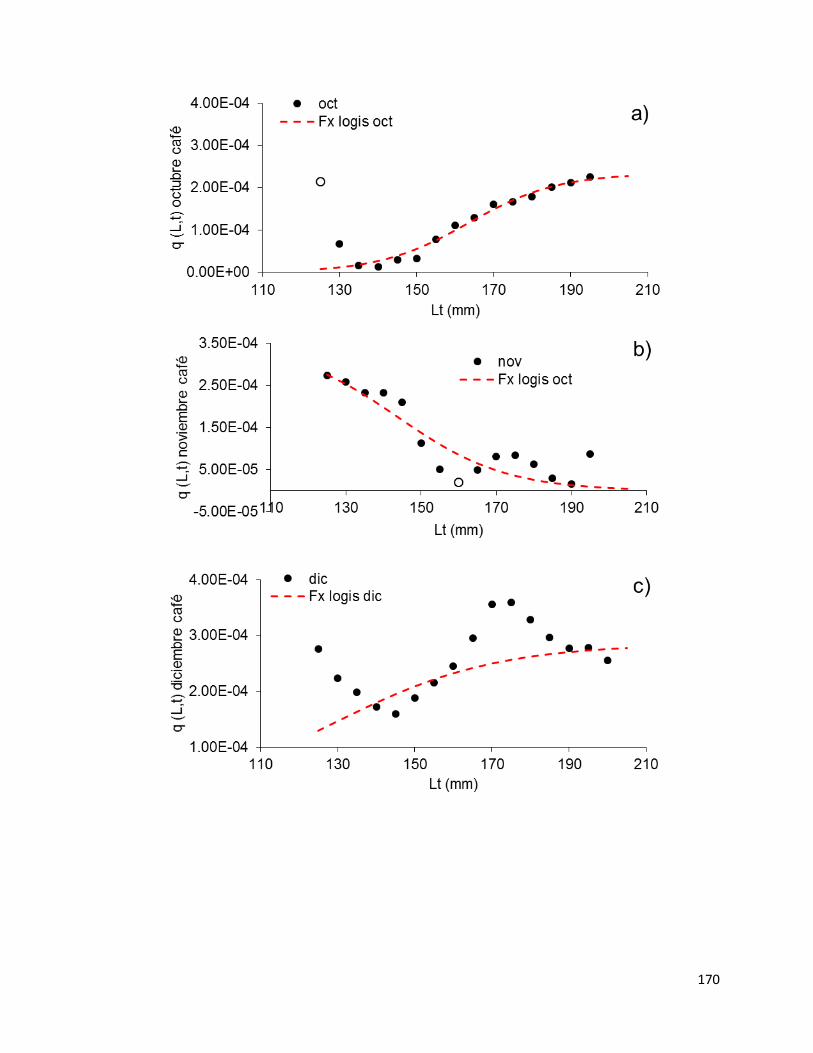
170

171
Figura 49- Patrón mensual de capturabilidad por longitud de camarón café para la flota
industrial en Sinaloa sur temporada 2014-15 (a-d). El resumen de los patrones de 𝑞(𝐿,𝑡)
mensual se observa en (f). Los círculos vacíos representan valores extremos
eliminados para fines del ajuste.

172
6.5 Relación Captura por unidad de esfuerzo y capturabilidad a la longitud por
flota.
La relación entre la capturabilidad a la longitud (𝑞𝐿,𝑡) y la captura por unidad de
esfuerzo (CPUE) se observan en la figura 50a-d. Se puede observar de manera
general una tendencia similar para la flota industrial (fig. 50 b-d), reflejando la
abundancia del recurso y su captura excepto en el mes de febrero para el camarón
café y en la flota de pequeña escala para el mes de noviembre (fig. 50a).

173
Figura 50 – Relación entre la capturabilidad a la talla (q(L,t) mensual promedio con la
captura por unidad de esfuerzo (CPUE) mensual para la flota de pequeña escala para
camarón blanco (a), y en la flota industrial para el camarón blanco (b), azul (c) y café
(d).

174
6.5. Modelo bioeconómico para la pesquería multi-específica y multi-flota de
camarón
6.5.1. Salidas del modelo bioeconómico de estructura por edades para la
pesquería del camarón en Sinaloa sur temporada 2014-15
El modelo bioeconómico descrito incorpora datos de las dos flotas que inciden
en diferentes estructuras de la población de las tres principales especies de camarón
capturados en el Pacífico Mexicano. De las cuales solo el camarón blanco estuvo
presente en las capturas de la flota de pequeña escala y las tres especies estuvieron
presentes como preadultos y adultos en las capturas de la flota industrial para la
temporada 2014-15 en Sinaloa sur. Se comparan los datos de observados de la
captura mensual con los estimados por el modelo por cada flota y especie en las
figuras 51-53.
El reclutamiento estacional se aproximó a los índices de maduración de
hembras estacionales reportados en la figura 2 donde los picos máximos coinciden
con los datos observados por el Instituto Nacional de Pesca. Se puede observar que
las especies costeras (Fig. 54 a-b) presentaron sus picos de desoves en primavera y
en verano. Para el caso del camarón café los mayores reclutamientos fueron
observados hacia verano otoño y presentó la mayor magnitud de individuos reclutados
(Fig. 54 c).
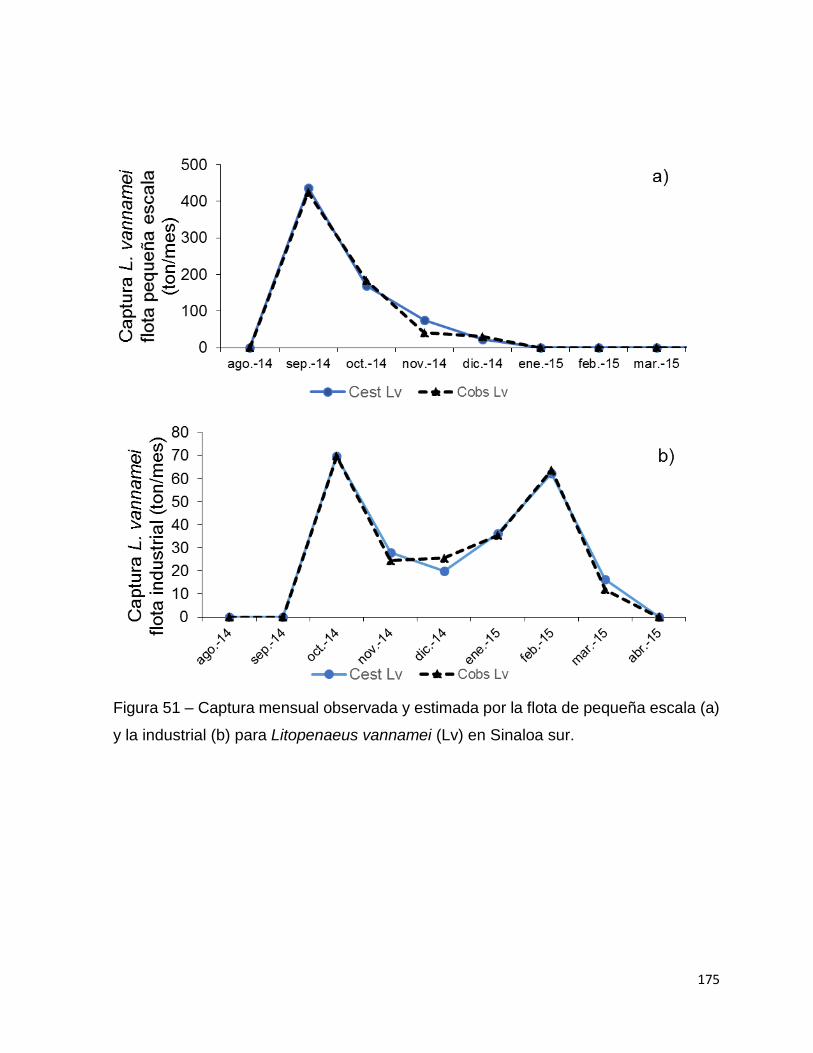
175
Figura 51 – Captura mensual observada y estimada por la flota de pequeña escala (a)
y la industrial (b) para Litopenaeus vannamei (Lv) en Sinaloa sur.
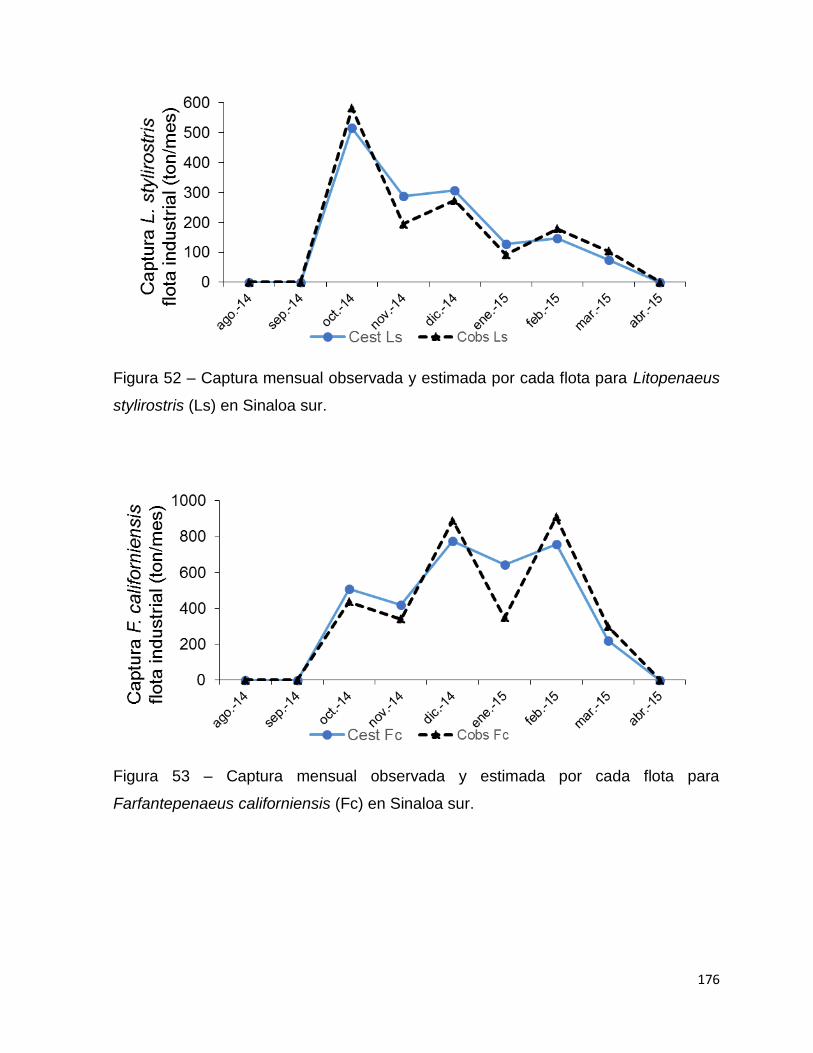
176
Figura 52 – Captura mensual observada y estimada por cada flota para Litopenaeus
stylirostris (Ls) en Sinaloa sur.
Figura 53 – Captura mensual observada y estimada por cada flota para
Farfantepenaeus californiensis (Fc) en Sinaloa sur.

177
Figura 54 – Reclutamiento estacional mensual estimado para las especies de
Litopenaeus vannamei, L. stylirostris y Farfantepenaeus californiensis (Fc) en Sinaloa
sur.
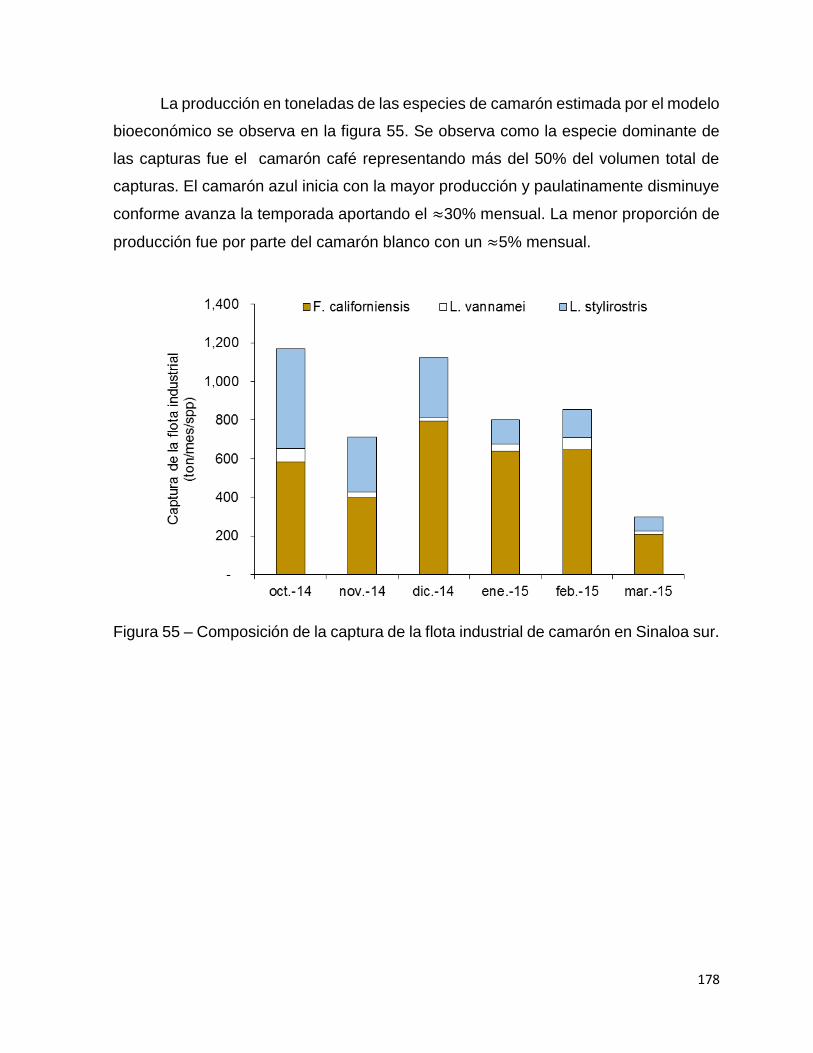
178
La producción en toneladas de las especies de camarón estimada por el modelo
bioeconómico se observa en la figura 55. Se observa como la especie dominante de
las capturas fue el camarón café representando más del 50% del volumen total de
capturas. El camarón azul inicia con la mayor producción y paulatinamente disminuye
conforme avanza la temporada aportando el ≈30% mensual. La menor proporción de
producción fue por parte del camarón blanco con un ≈5% mensual.
Figura 55 – Composición de la captura de la flota industrial de camarón en Sinaloa sur.

179
6.5.2. Escenarios de manejo para temporada 2014-2015 de la pesquería de
camarón en Sinaloa sur
La tabla 58 muestra todos los resultados del análisis de la temporada de pesca
por el modelo bioeconómico estructurado por edades. Cabe de mencionar que la
unidad año se refiere en el caso de utilidades y esfuerzo a la temporada de pesca
(septiembre-marzo) y en el caso del stock reproductor y reclutas al año de doce meses.
Cabe de mencionar que cada vez que se maximizaba el VPN usando el optimizador
SOLVER sobre el esfuerzo mensual de barcos, el mes de febrero siempre terminaba
con 1 barco, después la sumatoria final del total de barcos se repartía
proporcionalmente de acuerdo al esfuerzo mensual observado en status quo. Pero
esta transformación causaba una disminución en el VPN.
El valor presente neto (VPN) de la pesquería (fig.56) fue superior al escenario
status quo en todos los escenarios de manejo demostrando que el esfuerzo pesquero
se excede ≈ 60% tanto para los barcos como los días efectivos de pesca (tabla 58).
En cuanto a los escenarios de manejo el controlar el esfuerzo tanto en barcos como
en días de pesca generó resultados similares en todas las variables estado (fig. 56).
Hubo un aumento considerable de casi tres veces el valor base al juntar las salidas del
esfuerzo en días y barcos (en MEY) de los escenarios E2 y E3. Finalmente el escenario
con el mayor VPN fue el E4, donde se maximiza el VPN para días de pesca y luego se
maximiza para el número de barcos (fig. 56).
En las utilidades por barco (fig. 57) se observa que si reduces el esfuerzo en
barcos el escenario económicamente más óptimo es el ≈ E3 (US$600,000/barco año-
1); y en el escenarios E4 (MEY días seguido MEY barcos) se puede alcanzar un
escenario con un esfuerzo en barcos similar al actual (927 barcos año-1 vs 1,125
barcos año-1) con la gran diferencia que las utilidades por barco son tres veces
mayores que el escenario base.
Para la flota de pequeña escala (fig. 58) en todos los escenarios mejoraron sus
utilidades de un 5-7%, pero manteniendo el mismo esfuerza de pesca de 6,000
atarrayas en 3,000 embarcaciones únicamente para el camarón blanco.
Para las variables de reclutamiento y biomasa de reproductores (figs.59-60) el
escenario más conservador fue el E3 donde ambas variables son máximas, y para los
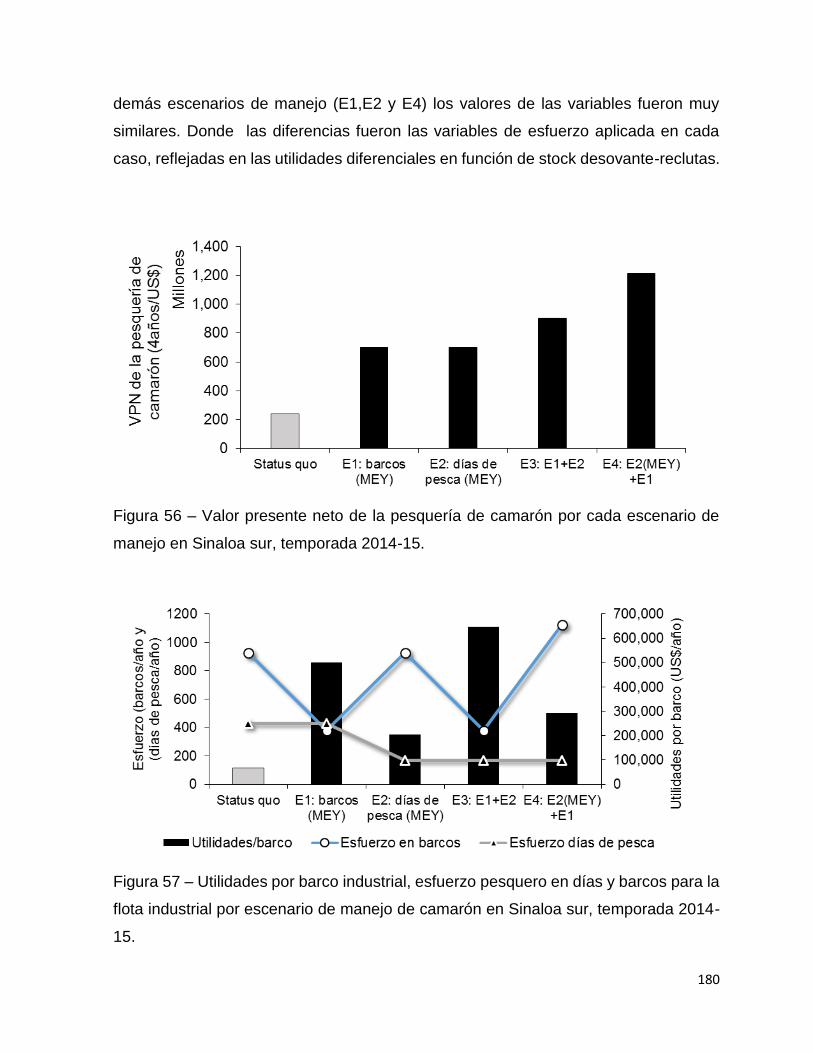
180
demás escenarios de manejo (E1,E2 y E4) los valores de las variables fueron muy
similares. Donde las diferencias fueron las variables de esfuerzo aplicada en cada
caso, reflejadas en las utilidades diferenciales en función de stock desovante-reclutas.
Figura 56 – Valor presente neto de la pesquería de camarón por cada escenario de
manejo en Sinaloa sur, temporada 2014-15.
Figura 57 – Utilidades por barco industrial, esfuerzo pesquero en días y barcos para la
flota industrial por escenario de manejo de camarón en Sinaloa sur, temporada 2014-
15.

181
Figura 58 – Utilidades por cayuco y esfuerzo total para la flota de pequeña escala por
escenario de manejo de camarón en Sinaloa sur, temporada 2014-15.
Figura 59 – Reclutas anuales por escenario de manejo junto a las distintas
combinaciones de esfuerzo estimado.

182
Figura 60 – Biomasa de reproductores anuales por escenario de manejo junto a las
distintas combinaciones de esfuerzo estimado.

183
Tabla 58 – Escenarios de manejo pesquero para el camarón en Sinaloa sur para la temporada de pesca 2014-15.
Status quo E1: barcos E2: días de pesca E3: E1+E2 E4: E2 + E1
MEY (barcos) MEY (días de
pesca)
(salidas E1 y
E2)
(E2 + MEY con
barcos)
Variables control
Esfuerzo días de pesca (año) 428 428 170 170 170
Esfuerzo número de barcos industriales (año)
927 378 927 378 1125
Esfuerzo de pequeña escala (atarrayas/año)
3,000 3,000 3,000 3,000 3,000
Variables estado
Valor Presente Neto (4años/US$) 238,197,567 701,615,555 699,438,657 904,105,276 1,214,238,853
Valor Presente Neto Fi (4años/US$) 225,486,522 688,269,730 686,040,319 889,865,360 1,200,609,540
Valor Presente Neto Fpe(4años/US$) 12,711,045 13,345,825 13,398,338 14,239,915 13,629,314
Utilidades Fi (US$/año) 62,022,507 189,453,972 188,835,619 244,939,440 330,386,830
Utilidades barco (US$/año) 66,907 500,965.42 203,706.17 647,683 293,527.06
Utilidades Fpe (US$/año) 3,462,015 3,634,953 3,649,258 3,878,510 3,712,169
Utilidades cayuco (US$/año) 1,154 1,212 1,216 1,293 1,237.39
Reclutas/año blanco (individuos/año) 2,736,000,000 2,895,803,601 2,909,008,833 3,082,039,944 2,890,959,227
Reclutas/año café (individuos/año) 16,900,000,00
0
28,506,440,58
5 28,947,826,259 33,376,768,445 29,038,890,925
Reclutas/año azul (individuos/año) 1,910,000,000 6,813,769,896 6,547,981,692 14,000,617,310 10,661,017,786
Biomasa reproductora blanco (ton/año) 10,989 13,314 13,537 17,046 13,234
Biomasa reproductora café (ton/año) 67,662 229,380 242,234 466,060 245,015
Biomasa reproductora azul (ton/año) 10,189 47,574 44,990 175,077 97,753
*Punto de referencia pesquero MEY: siglas en inglés para Máximo Rendimiento Económico; Fpe.: Flota de pequeña escala, Fi: flota industrial.

184
6.5.3. Escenarios de variabilidad interanual del reclutamiento para la temporada
2014-2015 de la pesquería de camarón en Sinaloa sur.
Una vez establecido el escenario base o status quo se procedió a incluir
únicamente la variabilidad en el reclutamiento estacional que se muestra en la tabla
59 sin modificar el esfuerzo simulando las condiciones status quo. Se observa bajo
escenarios base cuando ocurren los aumentos en los porcentajes de hembras
maduras observados en el aumento de los reclutamientos el VPN de la pesquería y la
biomasa del recurso puede llegar a aumentar de manera considerable (fig. 61-63). En
este punto hay utilidades superiores al escenario status quo para ambas flotas. Los
barcos industriales presentan aumentos de utilidades de US$66,000 año-1 a casi
US$450,000 año-1 por barco en el escenario de calentamiento moderado “CM” (tabla
60, fig. 62). La flota de pequeña escala por cayuco presentó un aumento de US$1,150
año-1 a US$4,370 año-1 bajo escenario de calentamiento moderado.
En cuanto a los reclutamientos y biomasa de stock desovantes (fig. 64-65) el
calentamiento favoreció su aumento en donde en el escenario de calentamiento
moderado todos los camarón aumentaron la biomasa del stock desovante 3-5 veces,
siendo el más favorecido el camarón blanco. Para los reclutamientos todos
aumentaron en diferentes proporciones siendo el más dominante el café duplicando
su reclutamiento.
Para el escenario de enfriamiento el resultado global fueron pérdidas a partir del
escenarios status quo. Por barco año-1 se pierde el 52% de la utilidad y para la flota de
pequeña escala se pierde solo el 30% (fig. 62 y 63). El stock desovante tiene pérdidas
del 35 al 45% en su biomasa y el reclutamiento del 20 al 44%, siendo el camarón café
y el blanco los más afectados.
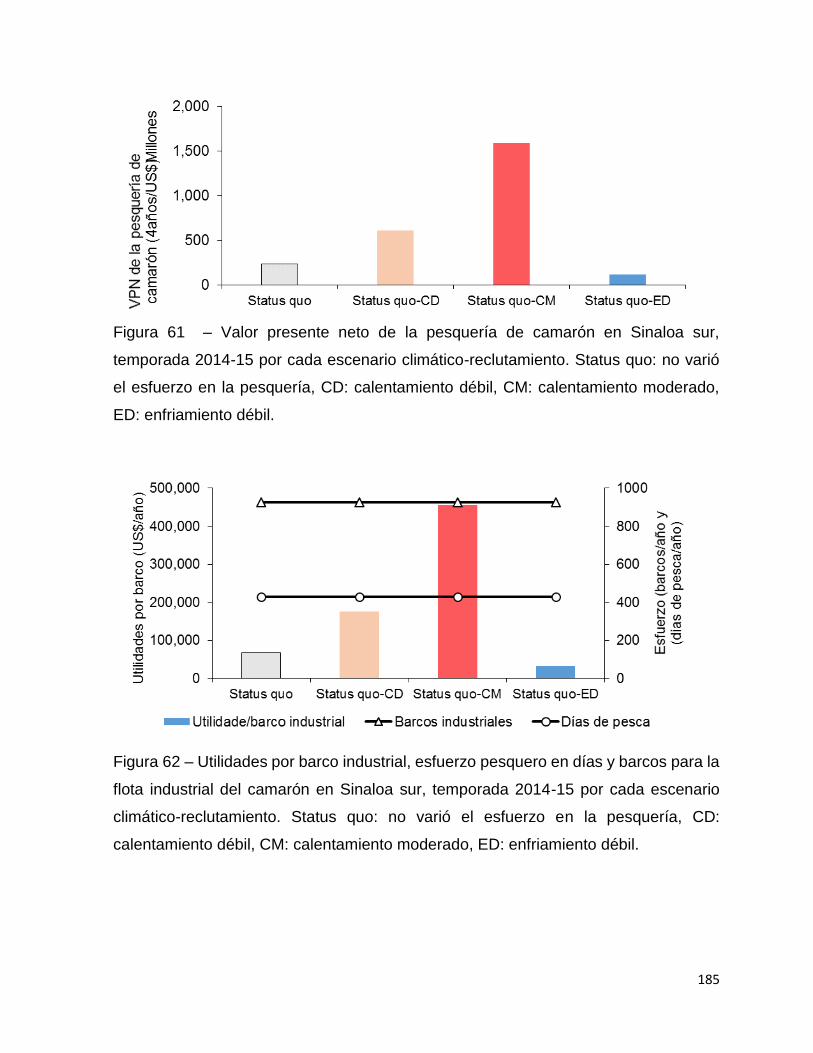
185
Figura 61 – Valor presente neto de la pesquería de camarón en Sinaloa sur,
temporada 2014-15 por cada escenario climático-reclutamiento. Status quo: no varió
el esfuerzo en la pesquería, CD: calentamiento débil, CM: calentamiento moderado,
ED: enfriamiento débil.
Figura 62 – Utilidades por barco industrial, esfuerzo pesquero en días y barcos para la
flota industrial del camarón en Sinaloa sur, temporada 2014-15 por cada escenario
climático-reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD:
calentamiento débil, CM: calentamiento moderado, ED: enfriamiento débil.
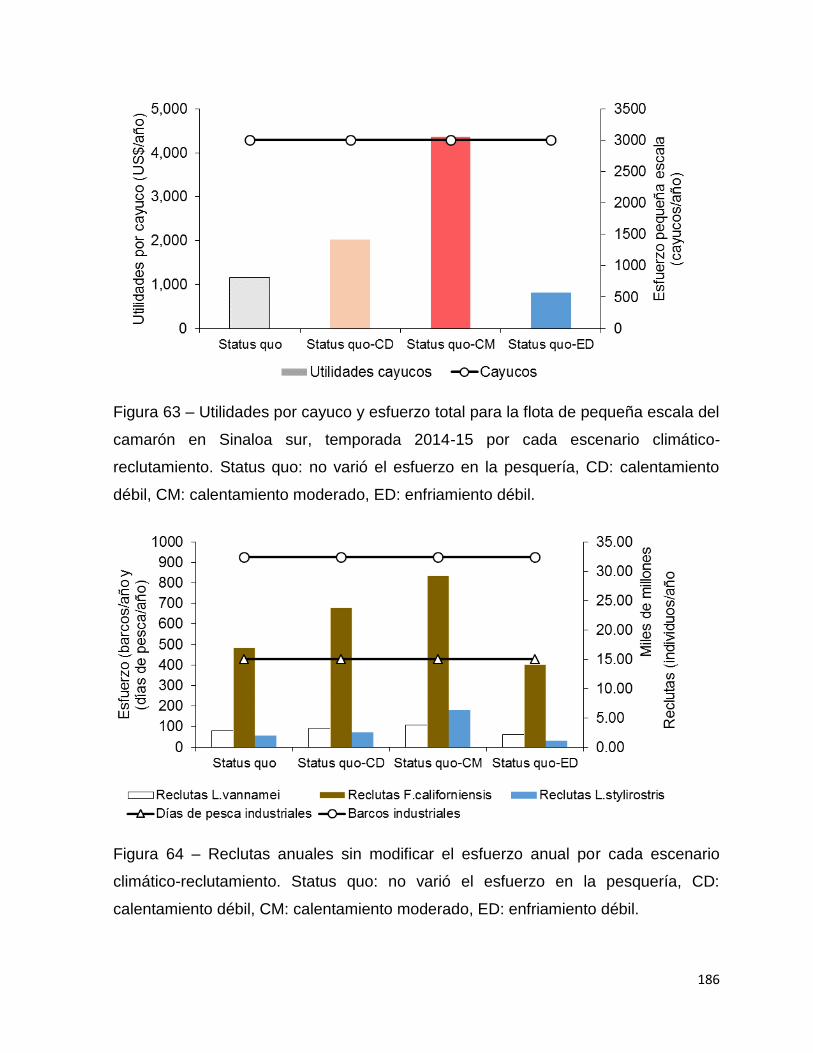
186
Figura 63 – Utilidades por cayuco y esfuerzo total para la flota de pequeña escala del
camarón en Sinaloa sur, temporada 2014-15 por cada escenario climático-
reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD: calentamiento
débil, CM: calentamiento moderado, ED: enfriamiento débil.
Figura 64 – Reclutas anuales sin modificar el esfuerzo anual por cada escenario
climático-reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD:
calentamiento débil, CM: calentamiento moderado, ED: enfriamiento débil.
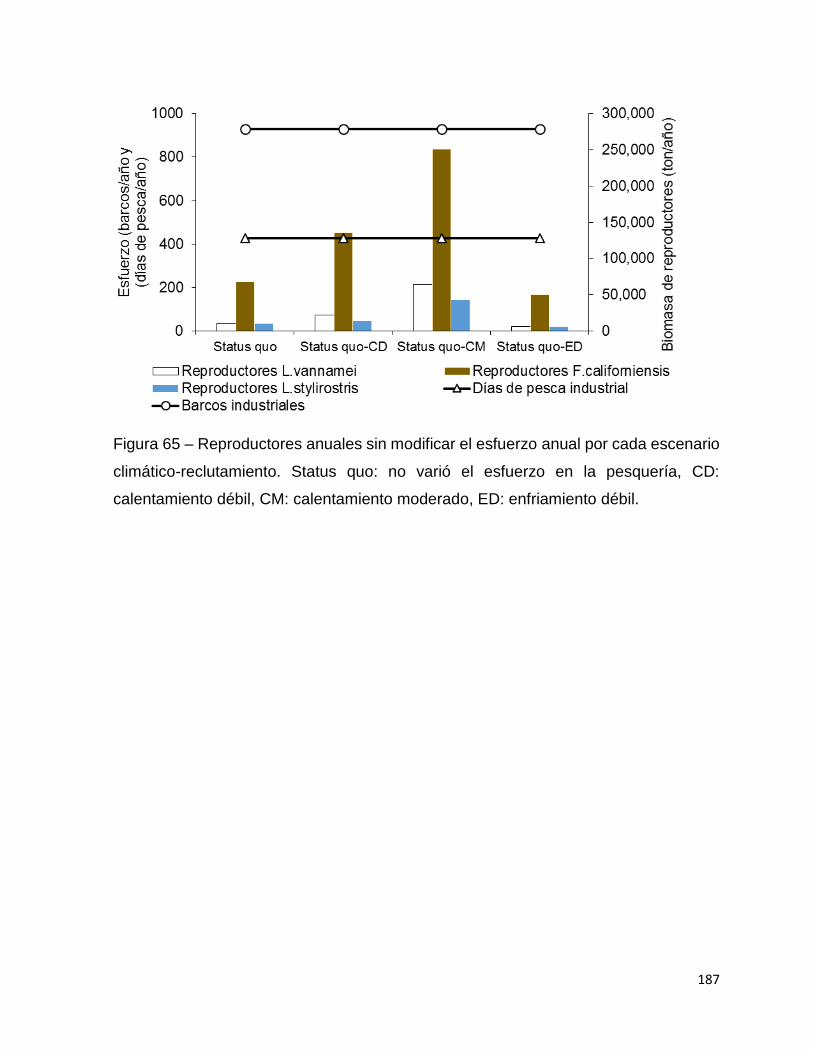
187
Figura 65 – Reproductores anuales sin modificar el esfuerzo anual por cada escenario
climático-reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD:
calentamiento débil, CM: calentamiento moderado, ED: enfriamiento débil.

188
Tabla 59– Frecuencias relativas de hembras maduras (promedio 1980-81 a 2012-13) y porcentaje de aumento (+) o
disminución (-) de hembras maduras de acuerdo a cada escenario climático.
Hembras maduras de camarón blanco Hembras maduras de camarón azul Hembras maduras de camarón café
Status quo Cal. débil
Cal. moderado
Enf. débil
Status quo Cal. débil
Cal. moderado
Enf. débil
Status quo Cal. débil
Cal. moderado
Enf. débil
Ene 0.039 1% 10% -1% 0.040 1% 7% -2% 0.136 6% 25% -5%
Feb. 0.067 3% 20% -3% 0.054 1% 15% -3% 0.118 4% 23% -5%
Mar. 0.088 2% 13% -4% 0.078 2% 9% -3% 0.134 5% 23% -5%
Abr. 0.133 8% 35% -9% 0.147 6% 13% -6% 0.040 2% 5% -1%
May. 0.081 6% 25% -6% 0.093 7% 23% -7% 0.082 4% 15% -4%
Jun. 0.055 5% 25% -4% 0.123 5% 19% -6% 0.172 8% 21% -7%
Jul. 0.085 5% 28% -3% 0.155 5% 24% -4% 0.172 5% 19% -5%
Ago. 0.122 11% 55% -6% 0.122 3% 11% -3% 0.202 5% 19% -4%
Sep. 0.020 15% 28% -2% 0.020 2% 4% -1% 0.219 7% 14% -6%
Oct. 0.010 8% 19% -1% 0.010 0% 2% -1% 0.155 6% 27% -6%
Nov. 0.013 6% 27% -1% 0.013 1% 4% -1% 0.193 8% 42% -8%
Dic. 0.035 5% 26% -3% 0.035 3% 32% -2% 0.148 7% 31% -6%
*Cal: Calentamiento, Enf: Enfriamiento

189
Tabla 60 - Escenarios de manejo pesquero para el camarón en Sinaloa sur con escenarios climáticos para la temporada
de pesca 2014-15.
Calentamiento débil Calentamiento moderado Enfriamiento débil Status quo Status quo % Status quo % Status quo %
Variables control
Esfuerzo días de pesca (año) 428 428 0% 428 0% 428 0%
Esfuerzo número de barcos industriales (año) 927 927 0% 927 0% 927 0%
Esfuerzo de pequeña escala (atarrayas/año) 3,000 3,000 0% 3,000 0% 3,000 0%
Variables estado
Valor Presente Neto (4años/US$) 238,197,567 614,802,293 158% 1,588,432,623 567% 117,984,021 -50%
Valor Presente Neto Fi (4años/US$) 225,486,522 592,476,740 163% 1,540,361,584 583% 109,042,952 -52%
Valor Presente Neto Fpe(4años/US$) 12,711,045 22,325,553 76% 48,071,039 278% 8,941,069 -30%
Utilidades Fi (US$/año) 62,022,507 162,908,437 163% 423,477,719 583% 30,001,499 -52%
Utilidades barco (US$/año) 66,907 175,737 163% 456,826.02 583% 32,364.08 -52%
Utilidades Fpe (US$/año) 3,462,015 6,089,596 76% 13,111,389 279% 2,434,107 -30%
Utilidades cayuco (US$/año) 1,154 2,030 76% 4,370.46 279% 811.37 -30%
Reclutas/año blanco (individuos/año) 2.74.E+09 3.25E+09 19% 3.71E+09 35% 2.16E+09 -21%
Reclutas/año café (individuos/año) 1.69.E+10 2.38E+10 41% 2.92E+10 73% 1.40E+10 -17%
Reclutas/año azul (individuos/año) 1.91.E+09 2.49E+09 30% 6.36E+09 233% 1.07E+09 -44%
Biomasa reproductora blanco (ton/año) 10,989 22,114 101% 64,009 482% 5,974 -46%
Biomasa reproductora café (ton/año) 67,662 135,638 100% 250,478 270% 50,038 -26%
Biomasa reproductora azul (ton/año) 10,189 13,728 35% 42,550 318% 5,575 -45%
*E: esfuerzo; VPN: Valor presente neto; U: Utilidades; F.i. : Flota industrial; F.p.e: Flota de pequeña escala; ind: individuos.

190
6.5.4. Escenarios de manejo para temporada 2014-2015 de la pesquería de
camarón en Sinaloa sur incluyendo la variabilidad interanual del reclutamiento.
Se compararon las salidas de los escenarios entre el escenario estatus quo
optimizado a MEY con los escenarios climáticos optimizados a MEY y se incluye de
referencia el estado sin optimizar de la pesquería (Status quo) (tabla 18).
De manera global con el calentamiento se pudo aumentar la flota industrial de
1 a 17% (378 barcos año-1 a 442 barcos año-1 ) obteniendo ganancias de hasta 9 veces
sobre el escenario status quo optimizado (MEY) en escenarios de calentamiento
moderado (figs.22-23). La flota de pequeña escala (fig. 24) también presenta un
aumento cuatro veces mayor (tabla 18) derivado de la disminución del esfuerzo
industrial y por el consiguiente aumento del reclutamiento en el camarón blanco. Bajo
escenarios de calentamiento moderado y con el ajuste de la flota los stocks desovante
aumentan en todas las especies y el más beneficiado es el camarón café y se refleja
la misma tendencia en el reclutamiento (tabla 18; fig. 25-26).
Para el caso del escenario de enfriamiento se tiene que disminuir el esfuerzo en
un 10% a 339 barcos año-1 y todas las variables estado presentan pérdidas sobre el
escenario status quo – MEY barcos de entre 30-45%, sin embargo sobre el escenario
status quo sin optimizar únicamente la flota industrial presenta ganancias tres veces
mayores (utilidades US$66,907 barco año-1 a US$310,573 barco año-1) al reducir su
esfuerzo de barcos en un 63%.
En cuanto al stock desovante y el reclutamiento el camarón blanco fue muy
sensible al enfriamiento con pérdidas del 34% en el stock desovante, por su parte el
camarón café y azul presentan ganancias del 160 y 111% (recordando que es bajo
escenarios de optimización de la flota en función del VPN). Pero todas las especies
presentan perdidas en los individuos reclutados que van del 8% café al 46% azul.

191
Figura 66 – Valor presente neto bajo condiciones de máximo rendimiento económico (MEY) de la pesquería de camarón en Sinaloa sur, temporada 2014-15 por cada escenario climático-reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD: calentamiento débil, CM: calentamiento moderado, ED: enfriamiento débil.
Figura 67 – Utilidades por barco industrial, esfuerzo pesquero en días y barcos para
la flota industrial del camarón bajo condiciones de máximo rendimiento económico
(MEY) en Sinaloa sur, temporada 2014-15 por cada escenario climático-

192
reclutamiento. Status quo: no varió el esfuerzo en la pesquería, CD: calentamiento
débil, CM: calentamiento moderado, ED: enfriamiento débil.
Figura 68 – Utilidades por cayuco y esfuerzo total para la flota de pequeña escala del
camarón bajo condiciones de máximo rendimiento económico (MEY) en Sinaloa sur,
temporada 2014-15 por cada escenario climático-reclutamiento. Status quo: no varió
el esfuerzo en la pesquería, CD: calentamiento débil, CM: calentamiento moderado,
ED: enfriamiento débil.
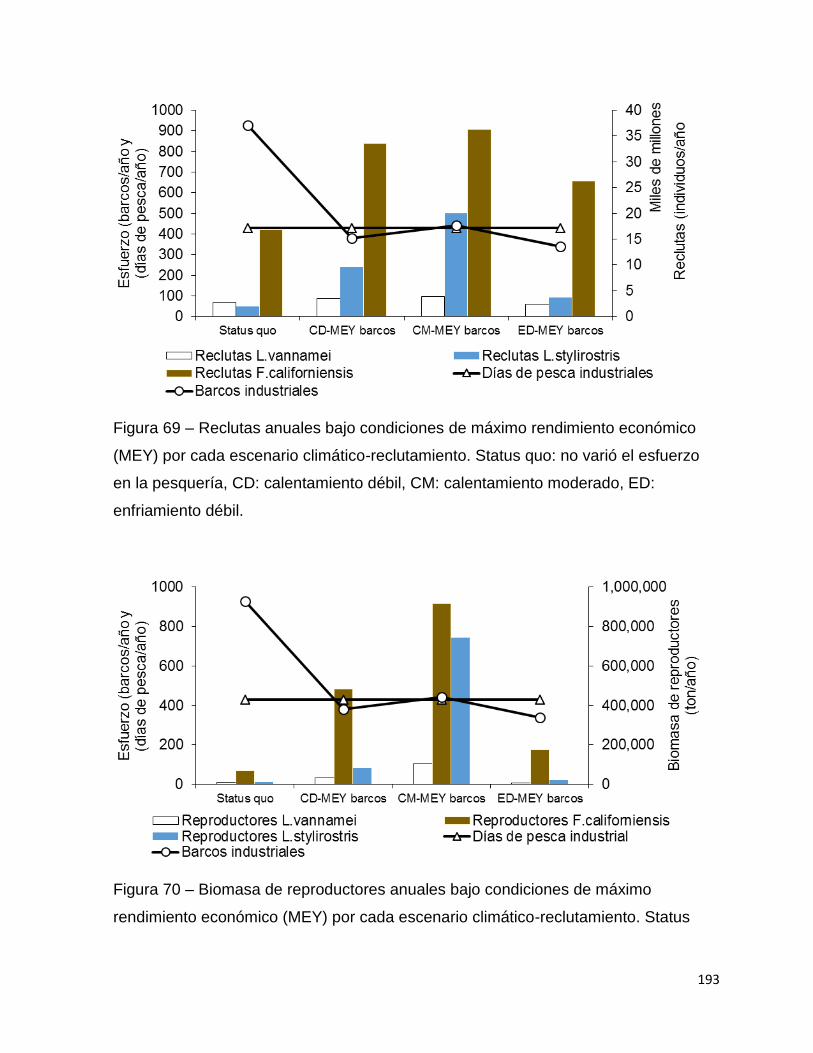
193
Figura 69 – Reclutas anuales bajo condiciones de máximo rendimiento económico
(MEY) por cada escenario climático-reclutamiento. Status quo: no varió el esfuerzo
en la pesquería, CD: calentamiento débil, CM: calentamiento moderado, ED:
enfriamiento débil.
Figura 70 – Biomasa de reproductores anuales bajo condiciones de máximo
rendimiento económico (MEY) por cada escenario climático-reclutamiento. Status

194
quo: no varió el esfuerzo en la pesquería, CD: calentamiento débil, CM:
calentamiento moderado, ED: enfriamiento débil.

195
Tabla 61 - Escenarios de manejo pesquero para el camarón en Sinaloa sur con escenarios climáticos con manejo de
esfuerzo a MEY para la temporada de pesca 2014-15.
Status quo Status quo Calentamiento débil Calentamiento moderado Enfriamiento débil MEY barcos MEY barcos % MEY barcos % MEY barcos %
Variables control
Esfuerzo días de pesca año-1 428 428 428 0% 428 0% 0%
Esfuerzo número de barcos año-1 927 378 381 -59% 442 -52% 339 -63%
Esfuerzo cayucos año-1 3000 3000 3000 0% 3000 0% 3000 0%
Variables estado
Valor Presente Neto (4 años/US$)
238,197,567 701,615,555 1,647,811,347 592%
8,047,685,596 3279% 391,127,640 64%
Valor Presente Neto Fi (4años/US$)
225,486,522 688,269,730 1,620,106,333 618%
7,980,337,033 3439% 382,160,837 69%
Valor Presente Neto Fpe (4años/US$)
12,711,045 13,345,825 27,705,014 118%
67,348,563 430% 8,966,803 -29%
Utilidades año-1 Fi (US$) 62,022,507 189,453,972 445,852,651 619% 2,194,772,680 3439% 105,205,485 70%
Utilidades año-1 barco (US$) 66,907 500,965 1,170,314 1649% 4,963,063 7318% 310,573 364%
Utilidades año-1 Fpe. (US$) 3,462,015 3,634,953 7,557,057 118% 18,369,589 431% 2,441,134 -29%
Utilidades año-1 cayuco (US$) 1,154 1,212 2,519 118% 6,123 431% 814 -29%
Reclutas blanco (individuos/año) 2.74.E+09 2.90.E+09 3.47E+09 27% 3.81E+09 39% 2.35E+09 -14%
Reclutas café (individuos/año) 1.69.E+10 2.85.E+10 3.36E+10 99% 3.63E+10 115% 2.62E+10 55%
Reclutas azul (individuos/año) 1.91.E+09 6.81.E+09 9.67E+09 406% 2.00E+10 947% 3.67E+09 92%
Biomasa reproductora blanco (ton/año)
10,989 13,314 33,436 204%
104,525 851% 7,225 -34%
Biomasa reproductora café (ton/año)
67,662 229,380 482,978 614%
916,836 1255% 176,462 161%
Biomasa reproductora azul (ton/año)
10,189 47,574 82,129 706%
742,327 7186% 21,525 111%
*MEY – Máximo rendimiento económico.

196
7. Discusión
La discusión se estructura por cada componente del modelo y al final una
integración global.
7.1. Parámetros de la dinámica poblacional
El parámetro esencial dentro de la dinámica de poblaciones son los modelos de
crecimiento edad porque de ellos depende el correcto análisis posterior en los modelos
estructurados por edades. Se escogieron los modelos basados en el crecimiento de
los camarones de acuerdo a von Bertalanffy sobre otros modelos porque estos se
basan en la propiedad fisiológica del crecimiento orgánico y describen correctamente
el crecimiento en peneidos (Tian et al. 1993). Donde se asume que el crecimiento
alcanza un máximo teórico de longitud o peso, y la tasa de crecimiento se reduce al
mínimo o ≈0 conforme se aproxime al valor máximo (von Bertalanffy, 1951; Pauly,
1981).
De manera general las tres especies de peneidos presentaron variabilidad en
las estimaciones de los parámetros para la ecuación de von Bertalanffy (tabla 8). Esto
era de esperarse debido a que el crecimiento en peneidos depende de factores
intrínsecos como el sexo (las hembras alcanzan mayores tamaños después de la
madurez e.g. Chávez, 1973), la etapa de vida (en la fase de aguas interiores el
crecimiento es más rápido que en aguas marinas e.g. Lluch, 1977) y por factores
externos como el ambiente (e.g. precipitación: Gutiérrez, 1980; fenómeno del Niño,
índice PDO: Castro-Ortiz y Lluch-Belda, 2008), calidad y cantidad de alimento,
densidad del stock (McGurk, 1986), temperatura del agua (e.g. Golfo de California:
Aragón-Noriega y Alcántara-Razo, 2005; Aragón-Noriega 2007; y la salinidad (Dall et
al., 1990).
Las ecuaciones de crecimiento seleccionadas (camarón azul: Rodríguez de la
Cruz, 1972 y café: Galicia, 1976) han sido empleadas y validadas en otros estudios
sobre la dinámica pesquera (e.g. Lluch, 1977; Saenz y Lluch, 1990) y a falta de un
muestreo sistemático para determinar los parámetros, se hace la suposición que la
tasa de crecimiento en cada especie de camarón no ha variado considerablemente y
es aplicable en la actualidad. Además que se validaron las salidas de los modelos de

197
crecimiento seleccionados con observaciones de muestras biológicas in situ en el átea
de estudio.
Con los modelos de crecimiento por especie se construyeron las clave edad-
longitud para azul y blanco porque solo se encontraron claves para el camarón café
(López-Martínez 2000; García-Borbón, 2009). Para las tres especies la función general
fue cuadrática, lo que implica que los intervalos de longitud media intermedios son los
de mayor variabilidad (i.e. desviación estándar). Esto concuerda con las etapas de la
edad de migración (i.e. edades 3 - 4 meses) para el caso de los camarones blanco y
azul, donde coexisten las cohortes de camarones pre-adultos en las zonas costeras,
y por otro lado la variación disminuye hacia organismos muy pequeños o muy grandes.
La clave edad del camarón café fue obtenida directamente de López-Martínez (2000)
y reportan la mayor desviación estándar en las edades 5 – 8 meses. Sainsbury (1980)
explica que cuando hay más de una cohorte en la población o una gran variabilidad
individual en los parámetros de crecimiento surge esta relación en forma de domo (i.e.
función cuadrática) entre las tasas de crecimiento y la longitud inicial. Esto reflejaría
que en la construcción de la clave edad-longitud se usaron varios estudios para
conformar una base de datos completa que incluyera a casi todo el ciclo de vida,
usando varias cohortes y observando esta forma domo (Sainsbury, 1980). Sin
embargo para el camarón café (López – Martínez, 2000) se usaron muestreos
biológicos in situ y de igual manera se obtuvo la forma “domo”, lo que indica dos cosas,
que la captura proviene de varias locaciones en el Golfo de California o que de manera
natural las poblaciones de camarones están conformadas por varías cohortes producto
de una reproducción anual continua.
La estimación de la mortalidad natural a la edad (Mi) representa un gran reto
para todos los recursos, incluidos los peneidos, porque en general se desconoce toda
o parte de la biología en su ciclo de vida. Además que los modelos pesqueros usados
en los peneidos comerciales del Pacífico mexicano siempre usan una M constante
para las fases de adultos (tabla 10) aun cuando se manejen etapas más tempranas.
Que como se observó son claramente una subestimación del posible valor real del
correspondiente intervalo (figura 25). Además, debido a que la literatura utiliza distintos

198
estimadores de M basados en diferentes variables correlacionadas a M (e.g.
temperatura, longitud máxima, edad de madurez), estos estimados son dependientes
del lugar donde se obtuvieron las mediciones, por lo que no se recomiendan en
poblaciones de camarones con distintos ambientes.
Los valores de Mi obtenidos difieren entre las especies y podrían ser causa de
la historia de vida de cada especie. Para el intervalo huevo no existen estudios
directamente en crustáceos, pero en peces reportan mortalidades tan altas como M =
1328 anual, indicando valores promedio menores (Bunn et al., 2000), aproximándose
al rango estimado por nuestro estudio (M1 ó huevo = 1085-1139) para camarones.
Para el camarón blanco presentó una menor M promedio en sus estadios
larvarios (��1-4 = 382.73 año-1), haciéndolo menos vulnerable a la depredación del
plancton por un rápido desarrollo larvario dentro de las aguas protegidas; por otra parte
el camarón café cuya vida quasi-marina tuvo una mayor M promedio para la fase
larvaria (��1-4 = 396.37 año-1) acorde a su historia de vida, donde todas sus
metamorfosis ocurren en la columna de agua siendo más vulnerable a depredadores.
El camarón azul presentó la mortalidad promedio para la fase lavaría más alta de los
tres (��1-4 = 461.54 año-1), específicamente se debe a la fase nauplio con un M-vector
de 684 anual, casi duplicando el M-valor de los otros dos (M4 =278 blanco y 358 café).
Esta especie es la dominante en las captura de la ribera o franja costera (INP, 2013),
y en las desembocaduras de bahías o esteros (Rodríguez dela Cruz, 1976),
propiamente la zona de interfase entre aguas protegidas y oceánicas; estas zonas
están más expuesta a los factores ambientales que contribuyen con una mayor
mortalidad en la fase larvaria como son ser zonas de paso de depredadores, altas
densidades de con-específicos donde pueda ocurrir canibalismo, el oleaje y corrientes
costeras que alejen las larvas de zonas favorables (Thorson, 1966; Dahlberg, 1979;
Rumrill, 1990).
Para la fase de aguas interiores en el camarón blanco, el único valor
comparativo encontrado en la literatura fue el de Edwards (1977) en las lagunas de
Huizache y Caimanero, Sinaloa, con una M de 46.8 año-1, siendo similar al valor
encontrado de 46.43 año-1. Las postlarvas además de ser muy vulnerables a los
depredadores, los son a factores abióticos como las altas mortandades observadas

199
debido a las alzas en la temperatura (>35°C) y cambios en las salinidades de las
lagunas por desecación (Sepúlveda-Medina, 1981).
En la búsqueda de valores M de juveniles solo el camarón blanco presentó
estimaciones independientes, esto refleja su importancia pesquera en las zonas
sureñas del pacífico mexicano que abarca casi el total de las capturas (Chapa y Soto,
1969). Estudios locales reportan especies depredadoras de juveniles dentro de las
lagunas de la familia Portunidae (género Callinectes), y varios peces como robalos
(Centropomus robalito y C. nigrescens), bagres (Ariopsis seemani), lenguados
(Cyclopsetta panamensis), pargos (Lutjanus argentiventris y L. novemfasciatus) y
machetes (Chanos chanos) (Sepúlveda-Medina, 1981; Amezcua y Portillo, 2010).
Los estudios de evaluación de stocks se han concentrado en los intervalos
vulnerables a la pesca industrial o de altamar debido a que la información si está
disponible. Estos estudios abarcan organismos pre-adultos y adultos, razón por la que
existen estimaciones de M en estos intervalos de vida para las tres especies (tabla 10
y figura 25). El vector de M6 o pre-adultos estimado no tuvo gran variación en las tres
especies (M6 = 7.91 – 8.71 año-1), posiblemente se deba a que las especies con fase
estuarina (e.g. camaró blanco y azul) al alcanzar cierta edad comienzan la migración
hacia aguas abiertas para la reproducción y es el punto donde las tres convergen en
hábitat marino (García y Le Reste, 1987). Comparando con las estimaciones de
fuentes independientes (tabla 10), los rangos de M para pre-adultos en el camarón
blanco fueron 6.6 – 8.43 año-1, para azul de 0.96 – 10.08 año-1 y para el café de 7.8
– 8.52 año-1, donde claramente se observa una subestimación en los valores para el
camarón azul.
Finalmente el componente de adultos en las tres especies tuvo ligeras
diferencias, donde el rango fue de M7 = 1.57 - 2.89 año-1, siendo la menor mortalidad
natural en el camarón café. Esto se podría explicar porque habita zonas profundas dea
50 hasta 180m (Rodríguez de la Cruz, 1976) donde el ambiente es más estable y
posiblemente exista una reducción en la depredación derivado de: una red trófica
menos compleja, una reducción del espectro de luz y zonas de sedimento sin mucha
heterogeneidad de hábitats. Lo cual cambia en los hábitats más heterogéneos de sus

200
especies hermanas (azul y blanco) habitando a 20m de profundidad (Sepúlveda,
1999).
A nivel mundial se reportan M en stocks de adultos de 1.2 a 5.4 año-1 (García y
Le Reste, 1986) dentro de los cuales se encuentran nuestros valores de referencia.
Los valores de M reportados para camarones adultos del Pacifico mexicano en la
literatura fueron de 0.09 a 8.92 año-1. Caddy (1996) observó que valores en peneidos
adultos tan bajos como 0.2-0.5 de M son posibles pero implican una mortandad
extrema en huevos y larvas. Sin embargo valores mayores de 5 son incompatibles
para especies anuales (Caddy, 1996; Gracia 1997). Por lo tanto las magnitudes de
nuestros valores estimados de M-a la edad parecen ser razonables y compatibles con
las historias de vida de las tres especies.
7.2. Parámetros pesqueros
7.2.1. Sección esfuerzo y captura de la pesquería de camarón en Sinaloa sur.
En este trabajo se reportan por primera vez valores del esfuerzo observado de
las flotas que capturan camarón en el área de Sinaloa sur para la temporada 2014-15
(tabla 40-41), siendo estimados en el tiempo mediante datos satelitales para el caso
de la flota industrial y por medio de varias fuentes de información para el caso de la
flota de pequeña escala. Lo que ayuda a disminuir el error a no sobrestimar la
mortalidad por pesca por cada flota.
En la literatura no se menciona casi ningún dato de esfuerzo puntual ribereño
para Sinaloa sur debido a la dificultad en su obtención. Existen pocos reportes como
informes técnicos del CRIP-Mazatlán (CONAPESCA, 2007) donde mencionan valores
de 13,000 pescadores ribereños para el estado de Sinaloa lo que representa el 30%
en este estudio; otro estudio por Castro-Ortiz y Sánchez-Rojas (1976) mencionan para
el área de Sinaloa centro o el sistema lagunar de Santa María de la Reforma, Altata y
Ceuta un esfuerzo de 2,400 pescadores ribereños e indican que el esfuerzo aumentará
a una tasa del 10% anual, lo cual no es informativo para el presente estudio.
La proporción de especies capturadas para la flota industrial que arrastró en
Sinaloa sur representa otro problema. Para el estudio y los modelos pesqueros en
general se asume que los barcos industriales capturan en la misma área del puerto de

201
atraco. La realidad es que los barcos toman el sitio de Sinaloa sur más como de tránsito
(pero si los usan para pescar) hacia otras zonas donde el recurso se agrupa en
mayores densidades como sucede en Sinaloa centro, (al principio de la temporada), el
Golfo de Ulloa o hacia el Golfo de Tehuantepec (hacia el final de la temporada) (INP,
2002; ver fig.8). Por lo que se puede mencionar con seguridad que las capturas de
camarón reportadas no son exclusivas de Sinaloa sur. En la actualidad, únicamente
mediante muestreos de cruceros dirigidos por zona (Sinaloa sur es la zona 40) por
parte del INAPESCA y los reportados en los años 50 se puede conocer la proporción
real de las especies capturadas por zonas. De este modo se reporta que la proporción
de las especies en la captura en Sinaloa sur es de un 65% para el camarón blanco,
31% azul y 6% de café (INP, 2014a); otro crucero de muestreo (de septiembre hasta
diciembre) por Nuñez et al., (1951) en la zona de Teacapán y Ticama, Sinaloa sur
acentuó más la diferencia hacia un dominio del 92% para el camarón blanco, 7% de
azul y 1% de café. Y esto si contrasta con la proporción observada de las capturas en
la temporada 2014-15 donde el camarón blanco representa el 4%, el azul el 26% y el
café el 70% de la captura total. Este problema sigue presente en los estimados de la
estructura de tallas obtenidas a partir de la maquiladora Shrimp Paradise en Mazatlán
porque no cuentan con información sobre el origen de los arrastres, con su volumen y
la proporción de especies, además que también puede haber sesgo por confundir el
camarón blanco reproductor con azul en borda durante el proceso de seleccionando y
descabezando. Sin embargo si el origen es dentro del Golfo de California podría existir
alguna señal ó tendencia de la dinámica de la población por especie; como se observa
en la figura 9, la flota de altamar de Mazatlán captura a lo largo de la temporada un
50% en Sinaloa centro y norte, un 20% en Sinaloa sur y el resto repartido en Sonora,
Golfo de Ulloa y el Golfo de Tehuantepec; este 70% podría generar tal tendencia y los
resultados ser acordes a la zona.
La dinámica de la flota en el tiempo se ajusta a la comunicación personal que
se tuvo con armadores en Mazatlán donde manifestaron que al principio de la
temporada toda la flota de Sinaloa tiene el objetivo de capturar camarón azul (el sitio
preferido es Sinaloa centro). Esto porque es la especie que alcanza mayores tallas y
mayor precio de exportación (Figura 27) y al compartir el hábitat con el camarón blanco

202
este podría actuar más como una especie no objetivo, explicando sus volúmenes tan
bajos. Después se observa en la misma figura 27 que cuando disminuyen las
densidades del camarón azul en aguas costeras-someras, las embarcaciones cambian
la estrategia hacia aguas más retiradas y profundas donde se encuentra el camarón
café en mayores densidades explicando su aumento de abundancia en sus capturas
en los meses de diciembre y febrero. Se observa de igual forma en la figura 26a una
caída de producción general del camarón café en enero debida a que los barcos
(comunicación personal con armador de Mazatlán Jesús Ortíz) se esperan hasta
febrero para declarar su captura a puerto (Avisos de arribo) y maquilar el camarón.
En lo que se refiere propiamente a la estructura de tallas de la captura reportada
por cada flota se pueden observar algunas tendencias importantes por cada especie.
Para la flota de pequeña escala está el efecto de la presencia de los tapos, estas
barreras son muy eficientes para detener la migración del camarón blanco hacia aguas
abiertas y el efecto inmediato es la depleción del recurso durante la temporada. Esto
se observa en la cohorte principal capturada en septiembre que explica el gran
volumen de captura en toneladas (fig. 30) y las tallas por debajo de la talla óptima de
captura (sobreexplotación de crecimiento; fig. 29); y como en los meses siguientes las
tallas y la abundancia de la población disminuyen drásticamente (tabla 43 y fig.29 y
30). Otro factor causante de las bajas capturas subsiguientes son las fugas de
camarones dentro de las lagunas cuando hay grandes precipitaciones que inundan las
lagunas sobrepasando las barreras tipo tapo o inclusive rompiéndolas, ó tener tapos
dañados (comunicación personal con presidentes cooperativistas de Sinaloa sur).
Para la flota industrial existe un sesgo hacia tallas de reclutamiento hacia aguas
marinas en las capturas el cual ha sido reportado en varios estudios previos (Leal,
1999; López-Martínez, 2000), oscureciendo las señales de los picos de reclutamiento
a la pesquería. Se alcanza a observar los agrupamientos reproductivos que generan
capturas de tallas mayores en diciembre a marzo, y se acentúa en las figuras 32-34
de CPUE; siendo el periodo reproductivo secundario en temporada fría que han
manifestado otros estudios (Lluch, 1977; Leal, 1999; López, 2000; Castro-Aguirre y
Lluch-Belda, 2008), y cuya función es contribuir al mantenimiento de la biomasa de los
reproductores de primavera-verano. La única especie que no se comportó de manera

203
similar en la tendencia de la estructuras de tallas capturada fue el camarón café, donde
se observan picos bimodales (figs. 34) que representan una señal que tanto los
reclutas a la pesquería como de reproductores. Lo que concuerda con la biología
misma del camarón café, que al ser de hábitos profundos se ha indicado que la
estabilidad ambiental aumenta sus periodos de reproducción en contraste con sus
especies hermanas más costero-someras donde los picos son en primavera y verano,
y que esto se relaciona a la presencia anual de hembras maduras en el camarón café
reflejando la entrada continua de reclutas en la estructura de tallas de capturas
reportadas en este estudio (Leal et al., 2001; López-Martínez et al., 2003; Aragón-
Noriega y Alcantara-Razo, 2005; Cervantes-Hernández et al., 2008).
7.2.2. La capturabilidad a la longitud (𝒒(𝑳,𝒕)) en la pesquería de camarón 2014-15
en Sinaloa sur
El problema de aplicar una capturabilidad (q) constante en los modelos
pesqueros generalmente es una respuesta a la falta de información pesquera y
biológica disminuyendo la resolución del modelo; sin embargo este supuesto podría
aplicar en pesquerías donde se extrae el recurso con tallas muy homogéneas (muy
selectiva o con capturabilidad similar), con flotas con el mismo poder de pesca, en un
mismo espacio y tiempo, y con condiciones ambientales constantes (Arreguín-
Sánchez, 1996).
Para el caso de la pesquería de camarón el problema de usar una q constante
es que existe mucha variabilidad asociada a su estimación. Dentro de los factores
detectados en el presente estudio que generan variabilidad a q: 1) la naturaleza de la
pesquería: esta es una pesquería secuencial donde se explota distintos componentes
del ciclo de vida, con diferentes hábitats y distintos comportamientos (en los esteros el
recurso se encuentra disperso con gradientes de edades/tallas (Gutiérrez, 1980; Lluch,
1982) hasta que inicia la migración hacia mar abierto; en aguas marinas forman
agregaciones reproductivas y después se dispersan [Rodríguez de la Cruz, 1976]),
también cada etapa puede reaccionar diferencialmente ante el arte de pesca; 2) el
poder de pesca de cada flota es distinto (red atarraya-tapo vs un barco de arrastre con
dos redes); 3) el recurso se comporta de manera diferencial en el tiempo, pudiendo

204
identificarse en función de la vulnerabilidad diferencial temporal en las tallas de captura
(figs. 41, 43, 47 y 49); 4) el ambiente ocasiona cambios en la abundancia de los
camarones, afectando su comportamiento (dispersión, agregación) y puede hacer que
las poblaciones reproductivas aumenten o disminuyan su periodo reproductivo (Leal-
Gaxiola et al., 2001; López-Martínez et al., 2005; Aragón-Noriega y Alcántara Razo,
2005; Aragón-Noriega 2007; Castro-Aguirre y Lluch-Belda, 2008, Aragón Noriega et
al., 2012); y 5) el mismo comportamiento del pescador o la estrategia de pesca: en
aguas interiores al bloquear con tapos las salidas la mar abierto se está agregando el
recurso y aumenta la q; en aguas costeras los mismos barcos arrastrando en los límites
legales de las bocas de las lagunas donde se agregan los migrantes o dirigiéndose a
capturar el recurso más rentable (camarón azul) ó a mayores profundidades (camarón
café). Arreguín-Sánchez (1996) define estas mismas fuentes de variabilidad y las
asocia con la selectividad (i.e. artes de pesca empleados por flota: poder de pesca); la
accesibilidad (i.e. no hay permiso de pesca para la flota de pequeña escala en mar
abierto y viceversa); la disponibilidad (i.e. patrón de abundancia en el espacio y tiempo
de la población de camarón: hábitats por etapa de ciclo de vida) y la vulnerabilidad del
recurso (aumenta la probabilidad de captura hacia algunas edades o tallas por
características del arte de pesca o de la biología del recurso: agregaciones
reproductivas de adultos; agrupaciones de juveniles y preadultos en los tapos en aguas
protegidas).
Las estimaciones independientes de q con valores constantes empleadas en
modelos pesqueros nacionales de camarón (tabla 17) no toman en cuenta las fuentes
de variación antes mencionadas, siendo que la variabilidad de la q a la longitud o edad
del recurso es de las más importantes en pesquería secuenciales. Además que estos
modelos están bajo el supuesto de una mortalidad por pesca (F) constante para todas
las tallas en el tiempo-espacio, aplicada en los modelos de producción en la literatura
(Leslie y Davies, 1939; De Lury, 1947; Silliman y Paloheimo 1958, entre otros), donde
además manifiestan la dificultad en su estimación por falta de datos (Castro-Aguirre y
Sánchez-Rojas, 1976).
En este punto es claro que por la naturaleza secuencial, multiespecie, multiflota
y multiespacio de la pesquería nacional del camarón estos modelos con capturabilidad

205
constante no son óptimos para abordar el manejo pesquero secuencial en estas
pesquerías. Otras pesquerías que presentan secuencialidad es el mero rojo en
Yucatán (Arreguín-Sánchez y Pitcher, 1999) donde se observaron al igual que en el
presente estudio una variabilidad en q dependiente de la longitud o edad, del tiempo,
por denso dependencia y por la distribución de la flota. Por otro lado el camarón tiende
a formar agregaciones de migración a causa de los tapos y en la plataforma continental
por causas reproductivas que aumentan su vulnerabilidad ante las flotas pesqueras,
aumentado su capturabilidad por factores de denso dependencia naturales o
artificiales. En otras especies con comportamiento agregativo, como la sardina, se ha
observado que cuando su abundancia disminuye su comportamiento es agregarse
para protegerse haciendo que el recurso sea muy propenso a sobreexplotarse
(Martínez-Aguilar et al., 1997; Martínez-Aguilar et al., 2009).
Dentro de los modelos que aplican una capturabilidad a la longitud o edad en la
literatura gris está el empleado para el camarón café en Bahía Magdalena, B.C.S.
(García-Borbón, 2009) y el modelo usado por el Centro Regional de Investigaciones
Pesqueras en Guaymas (INP, 2002) para los camarones blanco, azul y café en aguas
protegidas y frente costero de Sinaloa (norte) y Sonora sur. García-Borbón (2009)
estima una qi a la edad semana-1 para la flota de pequeña escala en camarón café
donde existe una variedad más grande de artes de pesca. Este autor menciona una
relación proporcional entre la captura por unidad de esfuerzo y la q reflejo de la
abundancia del recurso. Esta relación se cumple en el presente estudio para todas las
especies exceptuando el mes de noviembre en camarón blanco para la flota de
pequeña escala y en café en febrero (fig. 50). Esto significa que en estos meses la
relación capturabilidad y biomasa no se cumple y existen otros factores asociados
como son el comportamiento o ambientales que no permitieron la relación en esos
meses.
Para el caso del modelo del Instituto de Pesca (INP, 2002) calculan la
capturabilidad mensual para las artes de red de arrastre tipo “chango” y redes
suriperas con qi variables. Sin embargo indican q constantes para las redes de arrastre
de los barcos y las redes de enmalle. Esto genera un modelo sesgado en su
componente marino al suponer una misma capturabilidad en todas las tallas marinas.

206
Además que es contradictorio con la biología del camarón adulto donde ya se
mencionó que existen eventos agregativos.
Como se observó en la q a la edad en la pesquería de camarón blanco (figura 45)
su vulnerabilidad comienza a aumentar desde que se agregan en los tapos para
realizar su migración hacia aguas abiertas (≈90mm) aumentando la capturabilidad
hacia organismos reproductores. El camarón azul y café no representaron capturas
significativas en la flota de pequeña escala, fungiendo más como especies de bycatch
en esta zona. Esto sugiere una hipótesis donde el origen del camarón azul es en la
zona costera o migran de otros sitios donde son las especies dominantes en las
lagunas como sucede en el centro de Sinaloa y hacia el norte del Golfo de California
(Rodríguez de la Cruz, 1976; Castro-Aguirre y Sánchez-Rojas, 1976; Vázquez-Melchor
et al., 2011). Para el origen del camarón café se ha hecho hincapié que por las
condiciones oceanográficas de las lagunas y la alta variación en la salinidad de Sinaloa
sur llevan acabo su ciclo de vida en aguas marinas (Rodríguez de la Cruz y Rosales,
1973).
En cuanto a la vulnerabilidad general, se observó para las tres especies un
aumento a partir de la edad de primera madurez (fig. 46 en azul y fig. 48 en café). Este
cambio también se había observado antes en otras especies como la sardina del
pacífico (Sardinops caeruleus) en el Golfo de California (Martínez-Aguilar et al., 1997;
Martínez-Aguilar et al., 2009). La vulnerabilidad específica por especie de las capturas
industriales (figs.27 blanco, fig.29 azul y fig.32 café) obedece a la época de
reclutamiento pesquero en octubre y a la época de reproducción invernal secundaria
(Barreiro-Güemes, 1986; Leal, 1999; Cervantes-Hernández et al., 2008; Castro-
Aguirre y Lluch-Belda, 2008; Vázquez-Melchor, 2011). También se observa que el
camarón es un recurso muy dependiente de las condiciones ambientales que desfasan
o alteran la magnitud de su época reproductiva (López-Martínez et al., 2010), pudiendo
traer consecuencias económicas a cada flota.
El aumento en la vulnerabilidad de reproductores del camarón blanco se
observó de octubre a enero (fig.27). Esto corresponde con la época reproductiva
observaba por Vázquez-Melchor (2011) en Mazatlán y Aragón-Noriega (2012) en
Tehuantepec y Mazatlán con sus respectivas variaciones en la época e intensidad

207
reproductivas dependientes del tiempo y espacio. La vulnerabilidad por talla de
camarón café mostró señales de reclutamiento a la pesquería en todos los meses
siendo más claros en noviembre y febrero con edades de 5 a 7 meses (fig. 32). Esto
corresponde con el reclutamiento continuo reportado previamente por Leal (1999) y
López (2000) en Sonora. Además Leal (1999) indica un tipo de reclutamiento
reproductivo con una mayor presencia de reclutas en los meses de noviembre, marzo
y mayo con edades de 4-6 meses lo que concuerda con lo observado en este estudio
y confirmado de igual forma por Cervantes-Hernández et al., (2008) en Tehuantepec
reportando un pico menor de reclutamiento en enero y febrero. Leal (1999) explica que
la temporalidad en los reproductores se debe a una estrategia evolutiva donde la
reproducción ocurre en conjunto con los picos de productividad primaria como
estrategia alimentaria para sus larvas.
La vulnerabilidad de los reclutas a la pesquería en la especie de azul fueron en
noviembre, diciembre y marzo y la de los reproductores fue en los meses de octubre,
enero y febrero (figura 47) esto corresponde con el pico reproductivo de octubre a
marzo reportado para la especie para la costa sur de Sinaloa (Garduño-Argueta y
Calderón Pérez, 1994); y coincide al final del pico reproductivo principal observado por
López-Martínez et al., (2005) en Sonora estableciendo un rango principal de marzo a
septiembre. Por otro lado las alta capturabilidad de adultos en los meses de enero y
febrero corresponden con el pico secundario de reproducción donde Castro-Aguirre y
Lluch-Belda (2008) explican que la influencia climática en este periodo frío (enero –
junio) sobre los organismos reproductores afectará la producción pesquera del a
siguiente temporada de pesca.
7.3. Modelo bioeconómico para la pesquería multi-específica y multi-flota de
camarón
7.3.1. Manejo actual de la pesquería de camarón por parte de las autoridades
pesqueras
De acuerdo con el plan de manejo del camarón para el litoral del Pacífico
Mexicano se establece como objetivo de manejo el conducir la pesquería de camarón
a niveles sustentables usando como herramienta de manejo el punto de referencia

208
objetivo del máximo rendimiento sostenible (siglas en inglés MSY). Este concepto esta
basado de modelos de producción excedentes donde suponen la existencia de
condiciones de equilibrio (poblacional y ambiental), en donde se busca el esfuerzo que
explote la población en su punto que representa la mitad de la capacidad de carga de
la población (K/2). Además este concepto fue originado para especies ícticas
tropicales-templadas longevas, suponiendo una sola población (uni-específico), donde
el componente adulto es explotado y tiene una distribución homogénea (Gulland y
Borema, 1973, García y Le Reste, 1987). Todo lo anterior es incompatible con la
biología, dinámica poblacional y pesquera de los peneidos, los cuales rompen con casi
todos los supuestos por presentar migraciones y agrupaciones estacionales, es una
pesquería multiespecífica y secuencial, y se desarrolla en ambientes es muy variable,
además que estos organismos particularmente son muy sensibles a las fluctuaciones
ambientales observado en la variación de su ciclo reproductivo/reclutamiento en época
amplitud y magnitud (Leal, 1999; López-Martínez, 2000; López-Martínez et al., 2005;
Aragón-Noriega et al., 2012). Por lo que los expertos a nivel mundial recomiendan no
usar el MSY como punto de referencia objetivo en la pesquería de peneidos (Garcia y
Le Reste, 1987).
Aun con toda la violación de supuestos muchos modelos pesqueros de camarón
publicados en el área de estudio (Golfo de California, Mazatlán, Sinaloa, Pacífico
Mexicano) siguen usando modelos de producción excedentes para obtener el MSY
como punto de referencia óptimo del estado de la pesquería, muchas veces
“obligados” por la falta de información disponible para el recurso (Morales-Bojórquez
et al., 2001; Medina y Soto, 2003 en camarón café; García-Juárez et al., 2009 en
camarón azul; Rivera et al., 2009 en camarón blanco; Madrid-Vera et al., 2012 en
camarón blanco), pero esto hace sospechar de su aplicabilidad que podrían resultar
riesgoso para la pesquería.
Para el caso del modelo empleado por el INP (INP, 2003) el cual es la
herramienta del manejo en México para esta pesquería, se emplea un modelo
estructurado por edades pero a diferencia del propuesto en el presente trabajo, este
continúa empleando una mortalidad natural (M) y una capturabilidad (q) en la flota de
arrastre constantes. Estos valores constantes no son acordes con la dinámica

209
poblacional el recurso (juveniles y adultos en diferentes hábitats) donde ya se ha
observado que presentan M a la edad diferenciales (Aranceta-Garza et al., 2016) y
que pueden causar sesgos importantes en estimaciones poblacionales.
7.3.2. Manejo de la pesquería de camarón bajo el punto de referencia de máximo
rendimiento económico (siglas en inglés MEY)
En el presente estudio se propone el uso del MEY bajo el supuesto económico
que la pesquería que maximice su potencial económico y esto usualmente satisface
los objetivos de conservación del recurso (Hannesson, 2007) y se puede observar en
los todas las salidas de manejo propuestas que al optimizar el valor presente neto se
reduce el esfuerzo, aumenta el ingreso y aumenta la biomasa del recurso (figura 56).
Este objetivo además este incluido dentro de los planes de manejo en la
pesquería de camarones de otros países como Australia, los cuales son considerados
a nivel mundial vanguardistas en el manejo pesquero (Dichmont et al., 2010; Kompas
et al., 2010), y además se propone en varios estudios bioeconómicos pesqueros de
camarón el uso del criterio MEY sobre el MSY (Önal et al.,1991; Christensen y
Vestergaard, 1993; Nance et al ., 2006), existiendo ventajas, observadas en este
estudio, como el “efecto stock” (Hannesson, 2007) donde el tamaño del stock será
mayor bajo MEY que MSY produciendo mayores utilidades (i.e. reduciendo costos o
aumentado la producción); conservando el ecosistema (menos impacto de arrastre de
redes) y el recurso (stock más grandes) (Kompas et al., 2010); y da la ventaja de
generar mayor resiliencia pesquera a los fenómenos climáticos.
El modelo bioeconómico bajo su escenario status quo (tabla 58, figs. 56 - 60)
mostró que cuando se optimiza la renta en función del esfuerzo (barcos o días) se
observa que existe un sobrante de esfuerzo del ≈60%, razón por la que cuando se
ajusta a niveles de utilidades máximas por barco industrial esta aumenta de
US$66,000 con un escenario (927 barcos año-1) a US$200,000 - US$600,000 barco
año-1 dependiendo de la variable control modifiques: días de pesca (428 a 170 días
año-1) o barcos (927 a 378 barcos año-1) , aumentando de dos a seis veces la utilidad.
Cuando se utiliza el resultado combinado de fdías de pesca MEY y fbarcos MEY aumenta la
utilidad hasta ocho veces (US$647,000 barco año-1). Existen estudios donde

210
manifiestan que no hay relación entre el stock reproductor y los reclutas en camarones
(García y Le Reste, 1987; Gulland y Borema, 1973) y otros donde confirman que el
reclutamiento en camarones peneidos no es azaroso y demuestran que si existe la
relación significativa con la abundancia del stock desovante (Ye, 2000), tal como se
demuestra en este estudio al liberar la presión pesquera sobre los reproductores y
aumentando los reclutas a la pesquería.
Uno de los principales problemas en la aplicación de punto de referencia MEY
es la poca aceptación social en su implementación. Porque involucra la reducción del
esfuerzo pesquero en pesquerías sobrecapitalizadas, como es el caso de México, y
no es socialmente aceptable, por lo que se dice que no es un objetivo socialmente
óptimo (Nance et al., 2006). En el modelo propuesto únicamente modificamos el
esfuerzo industrial y se dejó en status quo el esfuerzo ribereño, amortiguando un poco
este descontento social. Lo importante es que se trabajó bajo el criterio de bienestar
Pareto seguridad demostrando un aumento en las ganancias para todos los usuarios
de la pesquería. Además este nuevo análisis no está muy alejado de la realidad
pesquera mexicana porque existen actualmente programas que están intentando
reducir el esfuerzo pesquero de los barcos industriales.
Se planteó un cuarto escenario de manejo donde se maximiza la renta en días
(-60% del total de 428 días) y luego se vuelve a maximizar el VPN del esfuerzo
pesquero en barco, ajustándose casi al número de barcos anuales observados
(927status quo v.s. 1125MEY). Este escenario es el óptimo desde un punto de vista social
porque no se reduce el esfuerzo pesquero en barcos si no solo los días de pesca. Este
ajuste causaría que la utilidad por barco aumentara tres veces (≈US$295,000 barco
año-1) sobre su escenario base siendo un buen incentivo para proponerlo como medida
regulatoria.
Por otra parte las utilidades ribereñas no variaron mucho bajo estos escenarios
(de $US1,154 a US$1,300). La razón es que de acuerdo a los gráficos de reclutamiento
y stock desovante (figuras 59 y 60) el stock de camarón blanco no presentó aumentos
tan significativos como el caso del camarón azul y café a los cuales no tiene acceso
por la dominancia ecológica del camarón blanco en los sistemas lagunares del sur de
Sinaloa. Existen varias razones que podrían explicar porque no aumentaron las

211
utilidades en las unidades de esfuerzo de pequeña escala como en las industriales. 1)
Esto efecto podría ser a causa que solo el stock de camarón blanco está siendo
explotado por las dos flotas en Sinaloa sur, donde hay una gran captura de juveniles
(edades tempranas) que no ocurre para el caso del camarón café y azul; 2) porque no
se puede modificar el esfuerzo pesquero de la pequeña escala por tener miles de
usuarios participantes entre personas legales e ilegales, por representar una actividad
socioeconómica muy importante en la zona y rebasa la capacidad de las autoridades
para ejercer una regulación eficiente; y 3) de acuerdo a la carta nacional pesquera las
poblaciones de camarón blanco en Sinaloa están sobreexplotadas y se necesita
aplicar medidas regulatorias hacia la flota de pequeña escala.
7.3.3. Manejo de la pesquería de camarón bajo el punto de referencia de máximo
rendimiento económico (siglas en inglés MEY) con variabilidad en el
reclutamiento pesquero.
En el Pacífico Mexicano se ha demostrado que los camarones presentan
diferentes épocas e intensidades en su reproducción. Se ha demostrado que existen
gradientes latitudinales donde hay presencia de hembras maduras todo el año en
zonas tropicales (+T°C) y reproducción se restringe únicamente a los meses cálidos
de verano (-T°C) en zonas templadas para la misma especie (Aragón-Noriega y
Alcántara-Razo, 2005; Aragón-Noriega, 2007; Aragón-Noriega et al., 2012). Por otro
lado se ha demostrado que en periodos de enfriamiento la reproducción, las tallas de
los camarones y el reclutamiento pesquero disminuyen (López-Martínez, 2000; López-
Martínez et al., 2003; López-Martínez et al., 2004; Castro-Ortiz y Lluch-Belda, 2008)
reflejando una disminución global de la utilidades (Fig. 61-63).
Este efecto se pudo integrar al modelo bioeconómico en el submodelo de
reclutamiento estacional donde se pudo simular el aumentó en el reclutamiento
producto del aumento en la frecuencia de hembras maduras observado en el aumento
de todas las variables estado bajo escenarios de calentamiento (Fig.61-65). Sin
embargo con el escenario de enfriamiento el VPN de toda la pesquería fue menor al
valor status quo bajo un esquema de esfuerzo sin manejo (figura 61-63). Donde se

212
puede observar en las figuras 64-65 como las tres especies disminuyen
considerablemente su biomasa reproductora y su reclutamiento.
Bajo estos escenarios status quo de calentamiento siempre hubo una mayor
ganancia para la flota de aguas interiores en la producción del camarón blanco
(US$1,154status quo a US$4,370CM cayuco año-1), así como el café y azul para el caso
industrial (US$66,907 status quo a US$456,826Cal.Moderado barco año-1).
Caso contrario en los periodos de enfriamiento sin manejo (escenario base)
donde la flota industrial disminuye sus utilidades anuales en un -50% (US$66,907 a
US$32,364 barco año-1) y la flota de pequeña escala presentó perdidas de utilidades
por un -30% (US$1,154status quo a US$811 cayucos año-1). Pero cuando se ajusta el
esfuerzo industrial de barcos al objetivo a MEY (tabla 60) bajo escenarios de
enfriamiento las utilidades industriales cambiaron de US$32,364Status quo- enfriamiento a
US$310,573MEY-enfriamiento barco-1 año-1 pudiendo contrarrestar los efectos negativos del
enfriamiento sobre la reproducción y pudiendo aumentar en teoría hasta 3.5 veces la
utilidad sobre el escenario base.
Este resultado no fue homogéneo entre las flotas y las utilidades de pequeña
escala en condiciones de enfriamiento sin y con manejo bajo MEY presentaron
pérdidas hasta del 30%. Esto se debió de acuerdo a las variables estado a que la
biomasa reproductora más afectada fue la del camarón blanco (tabla 60). La cual
siguió afectada aun después del escenario de enfriamiento de manejo bajo MEY (tabla
61). No se encontraron estudios que aborden directamente el efecto del ambiente
sobre los reproductores de camarón blanco, pero Aragón-Noriega (2012) demuestra
que la reproducción (≈reclutamiento) es continua en zonas tropicales y se reduce
hacía los meses de verano en zonas de transición o sub tropicales. Esto refleja varias
situaciones: 1) la flota de pequeña escala camaronera sin ningún manejo (por su
presunta falta de regulación pesquera que vive hoy en día) resultó ser más vulnerable
ante el ambiente extremos por depender de una especie ambientalmente sensible,
situación que no ocurre en la flota industrial porque su pesca es multiespecífica; 2) La
biomasa del camarón blanco posiblemente se encuentre sobrexplotada y necesite más
de 4 años para poder incrementar su biomasa, esto concuerda con el plan de manejo
(2004) que califica al camarón blanco en estado de deterior y sobrexplotado para la

213
zona de Mazatlán; 3) el camarón blanco al ser explotado por las dos flotas en la zona
esto pudiera sugerir un esfuerzo mayor de manejo sobre las dos flotas obligando a
gestionar una mayor regulación del componente de aguas interiores debido a
sobrepesca de crecimiento (observable en este estudio) y a los reportes de pesca
ilegal.
7.4. Discusión global
Los modelos bioeconómicos son una herramienta básica en la evaluación de
cualquier recurso pesquero. Pero la calidad y resolución de cada modelo dependerá
directamente en la calidad de información disponible del recurso a manejar (García y
Le Reste, 1987; Seijo et al., 1997). La gran mayoría de los modelos bioeconómicos en
camarón continúan empleando modelos de producción excedente bajo objetivos de
máximo rendimiento sustentable (García y Le Reste, 1987; Morales-Bojórquez et al.,
2001; Medina y Soto, 2003 en camarón café; García-Juárez et al., 2009 en camarón
azul; Rivera et al., 2009 en camarón blanco; Madrid-Vera et al., 2012 en camarón
blanco). Los cuales no se deben aplicar en las pesquerías secuenciales, de ciclos de
vida corto y además sensibles al ambiente (López-Martínez, 2000; López-Martínez et
al., 2003; López-Martínez et al., 2004; Castro-Ortiz y Lluch-Belda, 2008). Lo que obliga
buscar otro tipo de modelos como los de estructurados por edades (García y Le Reste,
1987; Seijo et al., 1997; Anderson y Seijo, 2010).
Particularmente para las pesquerías de camarones peneidos se debe de contar
con un submodelo biológico que englobe toda la dinámica propia de estas especies.
Muchos de los modelos pesqueros anteriores en camarón para el Golfo de California
fallaron en obtener parámetros dinámicos tan básicos como es la mortalidad natural
variable a la edad (Lluch,1974; García-Gómez, 1976; Rodríguez de la Cruz, 1976;
Ross-Terrazas, 1988; García de Quevedo, 1990; Saenz y Lluch, 1990; Sepúlveda,
1999) que como se demostró puede crear sesgos muy importantes en pesquerías
secuenciales si se aplican valores constantes de adultos. Otra parte importante a
considerar es el área de estudio, porque debido a la gran variabilidad que presentan
tanto en crecimiento (Lluch et al., 1972; Edwards, 1974; Galicia, 1976; Márquez, 1976;
Gutiérrez, 1980; Menz y Bowers, 1980) como en sus épocas reproductivas (López-

214
Martínez, 2000; López-Martínez et al., 2003; López-Martínez et al., 2004), y no puedes
usar parámetros de crecimiento de áreas tropicales (Tehuantepec) en áreas templadas
(Alto Golfo de California o Bahía Magdalena) (Aragón-Noriega y Alcántara-Razo, 2005;
Aragón-Noriega, 2007; Aragón-Noriega et al., 2012). Esto también generaría una sobre
o subestimación de los periodos reproductivos o del reclutamiento, semilla elemental
que alimenta al modelo bioeconómico estructurado por edades.
Respecto al submodelo tecnológico ya se discutió ampliamente la problemática
que se genera al usar coeficientes de capturabilidad constantes en estos recursos tan
dinámicos. Porque se pierde información muy importante respecto a la vulnerabilidad
del recurso en ciertas épocas del año donde ciertas edades son más capturadas que
otras por comportamiento biológico o pesquero. Desafortunadamente la gran mayoría
de los modelos pesqueros en el Golfo de California optan por seguir usando estos
valores constantes por dificultades que surge en sus estimaciones (Morales-Bojórquez
et al., 2001; Medina y Soto, 2003 en camarón café; García-Juárez et al., 2009 en
camarón azul; Rivera et al., 2009 en camarón blanco; Madrid-Vera et al., 2012 en
camarón blanco). Por lo que este trabajo representa un gran avance en ese sentido y
es aplicable para otras zonas y especies.
Otra parte elemental que aporta este trabajo es el manejo de toda la pesquería
de camarón. Ningún trabajo en México da un manejo pesquero al camarón que incluya
todo el sistema de pesca: aguas interiores (Ramos et al., 2000), o pesquería de altamar
(Saenz y Lluch, 1990; Morales-Bojórquez et al., 2001; Medina y Soto, 2003 en camarón
café; García-Juárez et al., 2009 en camarón azul). Esta última es la más estudiada de
todas por el problema de falta la información de esfuerzo ribereño. Por otro lado los
datos de captura que incluyeran la estructura de la tallas de la captura en el tiempo y
por flota son inexistentes, la información es muy antigua ó no existe ó está fuera del
acceso público.
Otra observación importante en la parte pesquera es que para esfuerzo
industrial se tuvo acceso a los datos satelitales de la flota que arrastró durante la
temporada de pesca 2014-15 y reduciendo la sobrestimación del esfuerzo en barcos
o días. Un problema sin solución fue el supuesto que todos los barcos analizados
arrastraron exclusivamente en el área de Sinaloa sur. Este supuesto claramente es

215
falso porque la flota camaronera de altamar es altamente dinámica en el Pacífico
Mexicano y la flota de Mazatlán es la más invasora (INP, 2002), por lo que se espera
que las tendencias en la captura global reflejen alguna tendencia en la plataforma
continental de Sinaloa sur.
Con todo el trabajo anterior se integró el modelo bioeconómico estructurado por
edades y cabe de mencionar que no solo se usan los precios de la captura por especie
de camarón, sino que se incluye las ganancias reportadas por los barcos de la pesca
incidental o bycatch comercial. El manejo que se le aplicó en el presente estudio fue
pensada en los objetivos de conservación y máximas ganancias económicas (objetivos
globales del manejo de camarón en México), aplicando el criterio de bienestar Pareto
seguridad (Seijo et al., 1997; Bergstrom y Randall, 2016). Y para lograrlo se tomó como
punto de referencia objetivo el Máximo Rendimiento Económico (MEY), donde ya se
han discutido sus ventajas sobre el objetivo actual de manejo mexicano, el Máximo
Rendimiento Sustentable (MSY). Lo importante de este modelo además de ser muy
adaptable hacia condiciones o ambientes distintos en camarones o cualquier especie
con pesquería secuencial es que pudo demostrar que todos los escenarios bajo el
esquema de MEY superaron el escenario base en MSY (Figura 56), lo cual en teoría
se debe cumplir si disminuyen el esfuerzo de pesca. Pero la parte innovadora además
de todo el modelo estructurado por edades, es la introducción del reclutamiento
estacional. Este permitió observar como el escenario base bajo condiciones de
enfriamiento la producción de camarón disminuye hasta un 50% y como si se aplica
un manejo bajo condiciones de MEY este resultado se revierte incluyendo utilidades
en la flota industrial. Por lo que se podría proponer su uso junto a modelos climáticos
predictivos para generar sistemas de alerta para asesorar a los armadores con
respecto a la temporada que podrían esperar.

216
8. Conclusiones
1) Se construyó un modelo bioeconómico estructurado por edades, multiespecies,
multiflotas, con mortalidad natural a la edad, coeficientes de capturabilidad a la edad y
con una función de reclutamiento estacional por especie que tuvo un ajuste muy bueno
con los datos de captura observados en la temporada 2014-15 en Sinaloa sur y podría
ayudar al manejo de la pesquería de camarón en el país.
2) Se lograron estimar los vectores de mortalidad natural a la edad para las tres
especies comerciales más importantes de peneidos en el Pacífico Mexicano listos para
usarse en la pesquería secuencial del camarón.
3) Se logró construir la función cuadrática de la clave edad para el camarón azul y
blanco a partir de las ecuaciones de crecimiento seleccionadas y modificadas (p.e.
camarón blanco) datos de muestreos en la zona y la cual fue aplicada para la convertir
la captura en edades para el modelo bioeconómico.
4) Se pudo determinar un valor de esfuerzo observado en la temporada 2014-15 para
la flota industrial que arrastro en Sinaloa sur así como un valor de esfuerzo ribereño
observado a partir de varias fuentes de información. Del mismo modo mediante los
muestreos a las plantas de maquilas y reportes obtenidos directamente de las oficinas
del INAPESCA en Mazatlán se obtuvo la estructura de tallas de la captura para cada
flota.
5) Se obtuvo una capturabilidad a la edad para las tres especies de camarón comercial
en Sinaloa sur que presento variabilidad en el tiempo, talla/edad, hábitat, flota, arte de
pesca y especie, y el cual fue integrado al modelo estructurado por edades.
6) Se logró obtener ajustar una función gamma al reclutamiento estacional por cada
especie de camarón comercial en Sinaloa sur para la temporada 2014-15. Además se
pudo introducir la variabilidad interanual al modelo con observaciones técnicas del
INAPESCA para cada especie y poder simular condiciones de calentamiento -
enfriamiento y observar cómo afecta la pesquería.

217
7) Bajo el escenario base o status quo (en máximo rendimiento sustentable, MSY) el
modelo estructurado por edades mostró que las flotas industrial y de pequeña escala
no presentaron pérdidas para la temporada 2014-15. Pero al usar el manejo pesquero
bajo el criterio de optimización económica (MEY) todos los escenarios bajo distintas
combinaciones de las variables control superaron el criterio actual de manejo en el
camarón mexicano (MSY), aumentando la utilidad y conservando el recurso.
8) Bajo los escenarios climáticos con el escenario base (MSY) resultó que con
calentamientos moderados a débiles la producción pesquera aumenta para ambas
flotas, pero bajo condiciones de enfriamiento resultó en una disminución de hasta el
50% en el valor presente neto (VPN) de la pesquería. Al introducir el criterio MEY en
la flota industrial resultó que se pudo superar el enfriamiento y generar utilidades 3
veces mayores que el escenario base sin enfriamiento. Este no fue el caso para la flota
de pequeña escala debido a que el camarón blanco resultó ser sensible a las bajas
temperaturas (pérdidas del 30% de utilidades con y sin manejo), con el componente
de biomasa reproductora siendo el más afectado y esto dejaría vulnerable a la flota
ante eventos ambientales extremos por lo que habría que tomar medidas regulatorias
de manejo que incluyeran esta flota.

218
9. Recomendaciones 1) Se debe mejorar el acceso a la información pesquera del país por parte de las
agencias responsables del gobierno. Esto facilitaría el trabajo tanto para
investigadores del INAPESCA y académicos para la generación de nuevas estrategias
de manejo ayudado de modelos pesqueros de alta resolución. Esto mejoraría
significativamente el manejo de los recursos pesqueros nacionales.
2) Una parte importante en el éxito del manejo de las pesquerías del país es poder
ayudar a disminuir la pesca ilegal y concientizar a las partes involucradas en la
pesquería sobre las ventajas de aplicar la normatividad establecida en la pesquería
del camarón en México, el uso de los modelos bioeconómicos con datos actualizados
servirían como línea base para demostrar la magnitud de la pesca ilegal.
3) Realizar estudios sobre el origen de las especies del camarón azul y café, así como
conocer los volúmenes capturados de tallas de reclutas marinos que no se reportan
en la flota industrial que se capturan en Sinaloa sur para incluir la capturabilidad de
estas longitudes y cerrar el sistema pesquero para estas especies.
4) La detección de las especies capturadas por zona y su esfuerzo aplicado mejoraría
la resolución de las capturas reales por cada zona pesquera, sobre todo en la flota
industrial, lo que ayudaría a dar un mejor manejo por cada zona.
5) Se recomienda pedir al INP las proporciones de los índices de gonadales por año
para poder crear una función que relacione la temperatura y % hembras maduras por
cada especie.
6) Se recomienda tener información biológica de la zona de estudio porque la
información a partir de los avisos de arribo no respeta zonas de pesca y se inflan los
valores de captura de las especies, por ejemplo del camarón café.

219
7) Se recomienda explorara medidas de manejo en donde se separen las especies
costeras (azul, blanco) de la marina (café), y que se diseñe una veda específica para
cada hábitat, esta separación espacial natural podría aumentar la producción natural
del recurso camarón café.
6) Se recomienda el manejo del número de barcos sobre el número de días de pesca
por la facilidad de controlar desde el puerto la salida de barcos no autorizados y evitar
gastos de vigilancia a toda la flota del Golfo de California para que cumplan los días
autorizados de pesca (que además que no existe tal infraestructura en vigilancia).
Todos el modelo esta con la suposición que toda la pesca fue desarrollada en el área
de estudio, lo cual es incorrecto pero es la información disponible. Sería de interés a
las autoridades pesqueras el ejercicio del modelo bioeconómico pero con datos
pesqueros exclusivos de cada zona de pesca, explorando alternativas de manejo
pesquero a las aguas estatales y evitando que “flotas invasoras”, las cuales llegan a
superar en tecnología a flotas locales (e.g. Mazatlán v.s. BCS) y ayudar a dar un mejor
ordenamiento de la pesquería de camarón en el Pacífico.
7) Por último se recomienda combinar el modelo bioeconómico con un modelo
ambiental que pueda predecir las condiciones ambientales en escalas mensuales
donde se pudiera tomar decisiones a futuro (p.e. corto-mediano plazo) sobre el manejo
de la pesquería basados en variables ambientales y biológicas. Y Pudiendo dar una
alerta o aviso a los armadores de las llamadas temporadas malas.

220
10. Bibliografía
Alcántara-Razo, E. 2005. Índice de producción de huevos, reclutamiento reproductor
y distribución de la biomasa de camarón azul Litopenaeus stylirostris en el
frente costero de Agiabampo, Sonora-Sinaloa, México. M.Sc. thesis,
Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas,
La Paz, B.C.S., 75 pp.
Álvarez-León, R., Díaz-Ochoa, J. A., y Núñez-Martínez, E. 2009. Ecosistemas de
manglar, variabilidad climática y producción de camarones en el área
costera del Pacífico colombiano. Cienc. Mar. (37): 3-20.
Amezcua, F, Portillo A. 2010. Hábitos alimenticios del lenguado panámico Cyclopsetta
panamensis (Paralichthyidae) en el Sureste del Golfo de California. Rev.
Biol. Mar. Oceanogr. 45: 335-340.
Anderson, L.G. 1977. The economics of fisheries management. The John Hopkins
University Press, New York.
Anderson, L.G. Seijo J.C. 2010. Bioeconomics of fisheries management. Wiley-
Blackwell. Iowa. 305 pp.
Andrade-Vizcaíno, K. 2010. Descripción del desarrollo larval del camarón blanco
Litopenaeus stylirostris (Boone, 1931), y evaluación del índice de desarrollo
en función del régimen de alimentación. B.S., Universidad Autónoma de
Baja California Sur, Marine Biology Department, La Paz, B.C.S., 58 pp.
Aragón-Noriega, E. A. 2007. Coupling the reproductive period of blue shrimp
Litopenaeus stylirostris Stimpson, 1874 (Decapoda: Penaeidae) and sea
surface temperature in the Gulf of California. Rev. Biol. Mar. Ocean. 42(2),
167-175.
Aragón-Noriega, E. A., Alcántara-Razo, E. 2005. Influence of sea surface temperature
on reproductive period and size at maturity of brown shrimp
(Farfantepenaeus californiensis) in the Gulf of California. Mar. Biol. 146(2),
373-379.

221
Aragón-Noriega, E. A., Pérez-Arvizu, E. M., Valenzuela-Quiñonez, W. 2012. Latitudinal
variation in reproduction timing of whiteleg shrimp Litopenaeus vannamei
(Decapoda, Penaeidae) of the Mexican Pacific coast. Crustaceana. 85(3),
287.
Aranceta-Garza, F., Arreguín-Sánchez, F., Ponce-Díaz, G., & Seijo, J. C. 2016. Natural
mortality of three commercial penaeid shrimps (Litopenaeus vannamei, L.
stylirostris and Farfantepenaeus californiensis) of the Gulf of California using
gnomonic time divisions. Scien. Mar. (80): 199-206.
Arosemena, M. 1976. Influencia de la salinidad y temperatura en el comportamiento
de camarones juveniles. In: Castro-Aguirre J.L. (ed), Memorias del Simposio
sobre Biología y Dinámica Poblacional de Camarones. Instituto Nacional
de Pesca, pp. (1): 375-378.
Arreguín-Sánchez, F. 1981. Diagnosis de la pesquería de camarón de roca (Sicyonia
brevirostris Stimpson, 1871) de Contoy, Q. Roo, México. Cienc. Pesq. 1(2),
1-85.
Arreguin-Sanchez, F. 1992. An approach to the study of the catchability coefficient with
application to the red grouper (Epinephelus morio) fishery from the
continental shelf of Yucatan, Mexico. PhD dissertation, CINVESTAV
Mexico. 222 pp.
Arreguín-Sánchez, F. 1996. Catchability: a key parameter for fish stock assessment.
Rev. Fish Biol. Fisher. 6(2), 221-242.
Arreguin-Sanchez, F., Pitcher, T. J. 1999. Catchability estimates and their application
to the red grouper (Epinephelus morio) fishery of the Campeche Bank,
Mexico. Fish. Bull. 97(4), 746-757.
Aubert, H., & Lightner, D. V. 2000. Identification of genetic populations of the Pacific
blue shrimp Penaeus stylirostris of the Gulf of California, Mexico. Mar. Biol.
137(5-6), 875-885.

222
Baranov, T. I. 1918. On the question of the biological basis of fisheries. Nauchnyi
Issledovatelskii Ikhtiologicheskii Institut Isvestia. 1 (1): 81–128.
Barreiro-Güemes, M.T. 1986. Estudio sobre la madurez y desove de Penaeus
californiensis y Penaeus vannamei en la costa sur de Sinaloa. Mem.
Intercambio académico sobre investigaciones del mar de Cortés.
Hermosillo, Son. México.
Barrera-Huerta, R. R. 1976. Algunos aspectos a considerar para la determinación de
vedas, aperturas y cierres de tapos y reglamentación de la luz de malla de
las atarrayas de pesca de camarón en las marismas y lagunas del estado
de Oaxaca. Mem. Simp. Biología y Dinámica Poblacional de
Camarones.124-130.
Begon, M., Harper. J.L., Townsend, C.R. 1990. Ecology. Individulas, Populations and
Communities. Blackwell Scientific Publications, Boston.
Bergstrom, J.C. y Randall, A. 2016. Resource Economics: An Economic Approach to
Natural Resource and Environmental Policy, Fourth Edition. Edward Elgar
Publishing. UK.
Beverton R.J.H., Holt S. J. 1959. A Review of the Lifespans and Mortality Rates of Fish
in Nature, and Their Relation to Growth and Other Physiological
Characteristics. In: Wolstenholme G.E.W. and O'Conner M. (eds), The
Lifespan of Animals (Colloquia on Ageing). John Wiley & Sons, Ltd, pp 142-
180.
Bertalanffy, L. 1934. Untersuchungen über die Gesetzlichkeit des Wachstums. Dev.
Gen. Evo. 131(4), 613-652.
Bertalanffy, L. 1951. Theoretische Biologie, Zweiter Band: Sto¡wechsel, Wachstum. A
Francke AG Verlag, Bern, Switzerland, 418pp.
Beverton, R.J.H., 1957 On the dynamics of exploited fish populations. Fish. Invest.
Minist. Agric. Fish. Food G.B.(2 Sea Fish.). Series II. Vol.19:533 p.

223
Beverton, R.J.H. and Holt, S.J. 1957. On the dynamics of exploited fish populations.
Fish. Invest. Ser. II. Vol. 19. 533 pp.
Brusca, R.C., M.E. Hendrickx. 2008. The Gulf of California Invertebrate Database:
The Invertebrate Portion of the Macrofauna Golfo Database.
http://wwwdesertmuseumorg/center/seaofcortez/databasephp.
Bunn, N. A., Fox, C. J. and Webb, T. 2000. A literature review of studies on fish egg
mortality: implications for the estimation of spawning stock biomass by the
annual egg production method. The Centre for Environment, Fisheries and
Aquaculture Science, Science Series Technical Report, 111, 37 pp.
Caddy J.F. 1991. Death rates and time intervals: is there an alternative to the constant
natural mortality axiom? Rev. Fish Biol. Fisheries. 1: 109-138.
Caddy J.F. 1996. Modelling natural mortality with age in short-lived invertebrate
populations: definition of a strategy of gnomonic time division. Aquat. Living
Resour. 9: 197–207.
Cadima, E. L. 2003. Fish stock assesment manual. FAO Fisheries Technical Paper
393. 66 pp.
Cárdenas, F.M. 1951. Ciclo evolutivo de tres peneidos del Noroeste de México. Rev.
Soc. Mex. Hist. Nat. 12(1-4), 229-258.
Castro, R. G., y Arreguín-Sánchez, F. 1991. Evaluación de la pesquería de camarón
café Penaeus aztecus del litoral mexicano del noroeste del Golfo de México.
Cienc. Mar. 17(4), 147-159.
Castro-Aguirre, J. L. 1976. Efecto de la temperatura y precipitación pluvial sobre la
producción camaronera. In: Castro-Aguirre J.L. (ed), Memorias del
Simposio sobre Biología y Dinámica Poblacional de Camarones. Instituto
Nacional de Pesca. (1) pp. 8-13.
Castro-Ortiz J., Sánchez-Rojas M. 1976. Notas Preliminares del comportamiento y
dinámica poblacional de Litopenaeus stylirostris (Stimpson 1871), en los
sistemas lagunarios del centro de Sinaloa. In: Castro-Aguirre J.L. (ed),

224
Memorias del Simposio sobre Biología y Dinámica Poblacional de
Camarones. Instituto Nacional de Pesca, pp. (2): 213-253.
Castro-Ortiz, J. L., y Lluch-Belda, D. 2008. Impacts of interanual environmental
variation on the shrimp fishery off the Gulf of California. CalCOFI Rep.49,
183-190.
Caswell, H. 1989. Analysis of life table response experiments I. Decomposition of
effects on population growth rate. Ecol. Mod. 46(3-4), 221-237.
Cervantes-Hernández, P., Flores-Gómez, A., Serrano-Guzmán, S. J., Ramos-Cruz, S.,
Gallardo-Berumen, M. I. 2010. Historical exploitation and evaluation of
brown shrimp fishery Farfantepeneaeus californiensis (Decapoda,
Dendrobranchiata) in the Gulf of Tehuantepec, Oaxaca, Mexico. Mexico.
Panamjas. 5(4), 486-494.
Cervantes-Hernández, P., Gallardo-Berumen, M. I., Serrano-Guzmán, S. J. 2012.
Explotación y distribución de la captura artesanal de camarón en el sistema
lagunar Huave, Oaxaca, México. Rev. Cienc. Mar. Cost. 4, 67-81.
Cervantes-Hernández, P., Sánchez-Meraz, B., Serrano-Guzmán, S. J., Frías-Velasco,
A., Ramos-Cruz, S., Gracia, A. 2008. Variación interanual de la abundancia
de Farfantepenaeus californiensis (Holmes 1900) en el Golfo de
Tehuantepec. Hidrobiológica. 18(3), 215-226.
Chapa, S.H. y Soto, R.L. 1969. Relación de algunos factores ecológicos con la
producción camaronera de las lagunas litorales del sur de Sinaloa, México.
En: Coastal Lagoons. A Symposium. (Ayala-Castañares, A. y Phelger, F.B.,
eds) México D.F. 28-30 Noviembre 1967. 653-662.
Chávez E.A. 1973. Estudio sobre la tasa de crecimiento del camarón blanco
(Litopenaeus stylirostris Boone) de la región Sur del Golfo de California.
Cienc. Mex. XXVIII (2): 79-85.

225
Chávez E.A., Rodríguez de la Cruz M.C. 1971. Estudios sobre el crecimiento del
camarón café (Litopenaeus californiensis Holmes) del Golfo de California.
Rev. Soc. Mex. Hist. Nat. 32: 111-127.
Chávez, E.A., Arreguín-Sánchez, F.1982. Evaluación y diagnostico de la pesquería de
camarón del Golfo de Tehuantepec. Informe Definitivo. Centro de
investigación y Estudios Avanzados INP, Unidad Mérida. 60 p.
Chávez, J. 2000. Análisis dinámico del coeficiente de capturabilidad y sus implicancias
en la modelación de pesquerías: Mesodesma donacium en el banco de
bahía Coquimbo, un estudio de caso (Doctoral dissertation, Tesis.
Universidad Católica del Norte. Chile).
Chen, W., Al-Husaini, M., & Al-Foudari, H. M. 2007. Using age-structured models to
develop a stock recovery strategy for Kuwait's shrimp fishery. Fish. Res.
83(2), 276-284.
Christensen, S., & Vestergaard, N. 1993. A bioeconomic analysis of the Greenland
shrimp fishery in the Davis Strait. Mar. Res. Econ. 345-365.
CNP. 2012. Carta Nacional Pesquera. SAGARPA. Diario Oficial de la Federación 24
de agosto de 2012. SAGARPA, Segunda sección. Acuerdo por el que se da
a conocer la Actualización de la Carta Nacional Pesquera.
CONAPESCA. 2004. Plan de Manejo para la Pesquería de Camarón en el Litoral del
Océano Pacífico Mexicano. SAGARPA-CONAPESCA-INP. De:
http://www.inapesca.gob.mx/portal/
CONAPESCA. 2007. Comportamiento histórico de la producción de camarón en
esteros y bahías por Sociedades Cooperativas de Producción Pesqueras
Ribereñas por Ecosistemas Lagunares Costeros. 2000-2007.SAGARPA.
Dirección General de Planeación, Programación y Evaluación.
CONAPESCA, A. E. D. A. 2013. Pesca 2013. Comisión Nacional de Acuacultura y
Pesca. Mazatlán, Sinaloa, México, 2008b. De:

226
https://www.gob.mx/conapesca/documentos/anuario-estadistico-de-
acuacultura-y-pesca
CONAPESCA. 2014. Base de datos de producción anuario. De:
https://www.gob.mx/conapesca/documentos/anuario-estadistico-de-
acuacultura-y-pesca
CONAPESCA. 2015. Base de datos de producción anuario. De:
https://www.gob.mx/conapesca/documentos/anuario-estadistico-de-
acuacultura-y-pesca
Csirke, J. O. R. G. E. 1989. Changes in the catchability coefficient in the Peruvian
anchoveta (Engraulis ringens) fishery. The Peruvian upwelling ecosystem:
dynamics and interactions. 18: 207-219.
Dahlberg M.D. 1979. A review of survival rates of fish eggs and larvae in relation to
impact assessments. MFR. 41: 1-12.
Dall, W., Hill, B.J., Rothlisberg, P.C., Staples, D.J., 1990. Biology of the Penaeidae. In:
Blaxter, J.H.S., Southward, A.J. (Eds.), Advances in Marine Biology, vol. 27.
Academic Press, London, UK. 489 pp.
Del Valle, I., Martin, P. 1995. Interannual variation in the catch and mean length of
penaeid shrimp in the lagoons and coastal waters of Sinaloa, NW Mexico,
and their possible link with environmental factors. In: ICES Mar. Sci. Symp.
Vol (199): 370-378pp.
DeLury, D. B. 1947. On the estimation of biological populations. Biometrics. 3(4), 145-
167.
Días de Leon, A.J.C. 1993. Exploitation and management of the Sinaloa shrimp fishery,
Mexico. PhD dissertation, University of London.
Dichmont, C. M., Pascoe, S., Kompas, T., Punt, A. E., & Deng, R. 2010. On
implementing maximum economic yield in commercial fisheries. PNAS.
107(1), 16-21.

227
Edwards, R.C. 1977. Field experiments on growth and mortality of Litopenaeus
stylirostris in a Mexican coastal lagoon complex. Estuar. Coast. Mar. Sci. 5:
107-121.
Edwards, R.R.C., Menz, A., Nuñez, A. y Silva, J. 1975. The ecology and population
dynamics of penaeid shrimp in a Mexican coastal lagoon. Proceedings of
the International Symposium in Fisheries Science. University of Baja
California, Mexico.
FAO. 1999. La ordenación pesquera. Orientaciones Técnicas para la Pesca
Responsable N.° 4. Roma. FAO. 81 p.
FAO. 2006. Programa de información de especies acuáticas. Penaeus vannamei.
Programa de información de especies acuáticas. En: Departamento de
Pesca y Acuicultura de la FAO [en línea]. Roma.
Fox, W.W. Jr., 1970 An exponential surplus yield model for optimizing exploited fish
populations. Trans.Am.Fish.Soc., 99(1):80–8.
Galicia, R. 1976. Crecimiento del camarón azul (Penaeus stylirostris) y camarón café
(Penaeus californiensis) en la zona de Puerto Peñasco, Son. En: Memorias
del Simposio sobre Biología y Dinámica de Poblaciones de Camarones. (2):
189.
García de Quevedo, R. 1990. Determinación de algunos parámetros poblacionales y
producción máxima sostenible del camarón azul (Litopenaeus stylirostris
Stimpson, 1881) en el Alto Golfo de California. B.S. thesis, Universidad
Autónoma de Baja California. Ensenada, B.C., 59 pp.
García S, Le Reste L. 1987. Ciclos vitales, dinámica, explotación y ordenación de las
poblaciones de camarones peneidos costeros. FAO Doc. Téc. Pesca.
(203):180 pp.
García-Borbón, J.A. 2007. Análisis de población virtual en la pesquería de camarón
café (Farfantepenaeus californiesis, Holmes) en el complejo lagunar Bahía

228
Magdalena-Almejas, Baja California Sur, México. B.S. thesis, Universidad
Autónoma de México, D.F., México, 78 pp.
García-Borbón, J.A. 2009. Construcción de un modelo estructurado por edades para
la determinación del inicio de temporada de captura de camarón café
(Farfantepenaeus californiesis, Holmes) en Bahía Magdalena-Almejas, Baja
California Sur, México. M.Sc. thesis, Instituto Politécnico Nacional, Centro
Interdisciplinario de Ciencias Marinas, La Paz, B.C.S., 194 pp.
García-Gómez, M. 1976. Breve análisis de cuatro temporadas de pesca camaronera
en Puerto Peñasco. In: Castro-Aguirre J.L. (ed), Memorias del Simposio
sobre Biología y Dinámica Poblacional de Camarones. Instituto Nacional
de Pesca, pp. (2): 255-277.
García-Juárez, A. R., Rodríguez-Domínguez, G., y Lluch-Cota, D. B. 2009. Blue shrimp
(Litopenaeus stylirostris ) catch quotas as a management tool in the Upper
Gulf of California La cuota de captura de camarón azul (Litopenaeus
stylirostris ) como instrumento de gestión en el Alto Golfo de California.
Cienc. Mar. 35(3), 297-306.
García-Juárez, A.R. 2009. Bases para el manejo y el ordenamiento del recurso
camarón en el alto golfo de California. Tesis Doctorado, CIBNOR. 306pp.
Garduño-Argueta, H. 1976. Primeras repoblaciones de camarón en aguas protegidas
del litoral del pacífico mexicano. Resultados preliminares. In: Castro-Aguirre
J.L. (ed), Memorias del Simposio sobre Biología y Dinámica Poblacional de
Camarones. Instituto Nacional de Pesca, pp. (2): 140-152.
Garduño-Argueta H, Calderón-Pérez JA 1994. Abundancia y maduración sexual de
hembras de camarón (Penaeus spp.) en la costa sur de Sinaloa, México.
Rev Inv Cient, Universidad. Autónoma de Baja California Sur. Ser. Cienc.
Mar. 1:27–34
Giménez-Hurtado, E., Arreguín-Sánchez, F., & Lluch-Cota, S. E. 2009. Natural
mortality rates during life history stages of the red grouper on Campeche
Bank, Mexico. N.A. J. Fish. Manag. 29(1), 216-222.

229
Gracia, A. 1997. Simulated and actual effects of the brown shrimp, Penaeus aztecus,
closure in Mexico. MFR. 59: 18-24.
Gulland J.A. 1965. Survival of the youngest stages of fish and its relation to year-class
strength. Int. Comm. NW Atlant. Fish. 6: 363-371.
Gulland, J.A. 1964. Manual of methods for fish population analysis. FAO. Fish. tech.
Pap. No. 40-60pp.
Gulland, J.A. 1965. Estimation of mortality rates. Annex to rep. Arctic Fish. Working
Group. ICES,CM. . (3) 9 pp.
Gulland, J.A. 1983. Fish Stock Assessment. A Manual for Basic Methods. New York:
John Wiley and Sons. 223 pp.
Gulland, J.A. y L.K. Boerema. 1973. Scientific advice on catch levels.
Fish.Bull.NOAA/NMFS. 71(2): 325–35
Gulland, J. A., & Rothschild, B. J. 1984. Penaeid shrimps: their biology and
management. Fishing News Books Ltd.
Gutiérrez J.L. 1980. Ecología y crecimiento de Litopenaeus stylirostris (Boone) en
lagunas “Las Cabras” del sistema Chametla Teacapan, Sinaloa. B.S. thesis,
Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas,
La Paz, B.C.S., 95 pp.
Gutiérrez J.L.V. 1980. Ecología y crecimiento de Penaeus vannamei (Bonne) en
lagunas “Las Cabras” del sistema Chametla Teacapan, Sinaloa. Tesis
Licenciatura. CICIMAR-IPN.
Hannesson, R. 1978. A note on the welfare-economic consequences of extended
fishing limits. J. Environ. Econ. Manag. 5(2), 187-197.
Hannesson, R. 2007. A note on the ‘‘stock effect’’. Mar. Res. Econ. 22, 68–74.
Hernández-Covarrubias V., Estrada-Navarrete F.D., Madrid-Ver J., Abad-Rosales
S.M., Medina-Guerrero, R.M. 2003. Fecundidad del camarón azul
(Litopenaeus stylirostris ) de la ribera adyacente a la boca de Baradito de la

230
Bahía Santa María La Reforma, Sinaloa México. In: Instituto Nacional de
Pesca (ed), 2° Foro de Investigación de Camarón del Pacífico: Evaluación
y manejo, Huatulco, Oax. 12-13 Jun 2003. Instituto Nacional de la Pesca,
pp. 25.
Hernández-Covarrubias V., Muñoz-Rubí H.A., Madrid-Vera J. Chávez-Herrera D.
2012. Fecundidad del camarón blanco Litopenaeus stylirostris de la
plataforma continental de Sinaloa, México. Cienc. Pesq. 20: 17-21.
Hjort J. 1926. Fluctuations in the year classes of important food fishes. J. Cons. int.
Explor. Mer. 1: 5-38.
Holthuis, L.B. 1980. FAO species catalogue. Volume 1-Shrimps and prawns of the
world. An annotated catalogue of species of interest to fisheries. (No 125).
INP. 1974a. Resultados del muestreo de altamar junio. INP
INP. 1974b. Resultados del muestreo de altamar julio. INP
INP. 1974c. Resultados del muestreo de altamar agosto. INP
INP. 1974d. Resultados del muestreo de altamar septiembre. INP
INP. 1975a. Resultados del muestreo de altamar mayo. INP
INP. 1975b. Resultados del muestreo de altamar octubre y noviembre 1974 y enero
1975. INP
INP. 1975c. Resultados del muestreo de altamar mayo. INP
INP. 1975d. Resultados del muestreo de altamar junio. INP/SI: i42.
INP. 1975e. Resultados del muestreo de altamar julio. INP/SI: i78.
INP. 1975f. Resultados del muestreo de altamar junio-julio. INP/SI: i43.
INP. 1975g. Resultados del muestreo de altamar agosto. INP

231
INP. 2002. Evaluación de la migración y reclutamiento de las poblaciones de camarón
en aguas protegidas y en el frente costero de Sinaloa y Sonora. SAGARPA.
CRIP GUAYMAS. 179 pp.
INP. 2003. Muestreos de camarón en el Pacifico durante la veda de 2003 y
recomendación de apertura de temporada 2003-2004. SAGARPA.
INP. 2004. Inicio de la temporada de veda 2004 para la pesquería de camarón del
océano Pacifico mexicano. SAGARPA.
INP. 2013. Análisis de las capturas de camarón en la temporada 2012-2013 del litoral
Pacífico. SAGARPA. CRIP Mazatlán. México, 93 pp.
INP. 2014a. Dictamen de fin de veda: evaluación biológica de las poblaciones de
camarón durante la veda 2014 en el litoral del Pacífico Mexicano.
SAGARPA. CRIP Mazatlán. México, 96 pp.
INP. 2014b. Dictamen de inicio de veda 2014: análisis de las capturas de camarón en
la temporada 2013-2014 del litoral Pacífico. SAGARPA. CRIP Mazatlán.
México, 67 pp.
Jaquemín P.P. 1976. Estimación de algunos parámetros poblacionales de tres
especies de camarón del Pacífico mexicano. In: Castro-Aguirre J.L. (ed),
Memorias del Simposio sobre Biología y Dinámica Poblacional de
Camarones. Instituto Nacional de Pesca, pp. (2): 169-189.
Jensen A.L. 1996. Beverton and Holt life history invariants result from optimal trade-off
of reproduction and survival. Can. J. Fish. Aquat. Sci. 53: 820-822.
Jones, R. 1974. Assessing the long-term effects of changes in fishing effort and mesh
size from length composition data. ICES C.M. 1974/F:33. 13 pp.
Jones, R. 1981. The use of length composition data in fish stock assessment (with
notes on VPA and cohort analysis). FAO Fish. Circ. No. 734. 55 pp.

232
Kitani H. 1986a. Larval Development of the White Shrimp Litopenaeus stylirostris
Boone Reared in the Laboratory and the Statistical Observation of. Bull. Jpn.
Soc. Sci. Fish. 52(7): 1131-1139.
Kitani H. 1986b. Larval development of the blue shrimp Litopenaeus stylirostris
Stimpson reared in the laboratory. Nippon Suisan Gakk. 52(7): 1121-1130.
Kitani H., Alvarado J.N. 1982. The larval development of the Pacific brown shrimp
Penaeus californiensis Holmes reared in the laboratory. Bull. Jpn. Soc. Sci.
Fish. 48: 375-389.
Kompas, T., Dichmont, C. M., Punt, A. E., Deng, A., Che, T. N., Bishop, J., Gooday, P.,
Ye, Y., Zhou, S. 2010. Maximizing profits and conserving stocks in the
Australian Northern Prawn Fishery. Aus. J. Agric. Res. Econ. 54(3), 281-
299.
Leal, A.G. 1999. Análisis de la variabilidad interanual en el patrón reproductivo y talla
de primera madurez sexual del camarón café Farfantepenaeus
californiensis (Holmes 1900) en el litoral sonorense. Tesis de Licenciatura.
UAS.
Leal-Gaxiola, A., 1999. Análisis de la variabilidad interanual en el patrón reproductivo
y talla de primera madurez sexual del camarón café Farfantepenaeus
californiensis (Holmes, 1900) en el litoral sonorense: 1-64. (Professional
Thesis, Universidad Autónoma de Sinaloa).
Leal-Gaxiola, A., López-Martínez, J., Chávez, E. A., Hernández-Vázquez, S., &
Méndez-Tenorio, F. 2001. Interannual variability of the reproductive period
of the brown shrimp, Farfantepenaeus californiensis (Holmes, 1900)
(Decapoda, Natantia). Crustaceana-IJCR. 74(9), 839-852.
Leslie, P.H. and Davies, H.S. 1939. An attempt to determine the absolute number of
rats on a given area. J Anim. Ecol. 8, 94-113.
Lluch, B. D. 1974. La pesquería de camarón de Altamar en el noroeste. Un análisis
biológico pesquero. Serie Informativa INP/S1: 116 pp.

233
Lluch, B. D. 1977. Diagnóstico, modelo y régimen óptimo de la pesquería de camarón
en altamar en el noroeste de México. Tesis doctoral. IPN-CICIMAR.
Lluch, B. D. 1982. La pesquería del camarón del Pacífico (Diagnosis monográfica de
los conocimientos existentes). IPN-CICIMAR, DGINP-EPES, CICTUS-UAS.
Cocoyoc, Morelos.
Lluch, B.D., Guzmán, S., y R.L. Soto. 1972. Programa-Camarón del Pacifico.
Resultados Parciales de Muestreos en Lagunas Costeras del Noroeste.
Informe Técnico INP/SI:14. Instituto Nacional de Pesca, México.
López-Martínez, J. 2000. Dinámica de la pesquería de camarón café (Penaeus
californiensis) en el litoral sonorense y su relación con algunos parámetros
océano-atmosféricos. Ph.D. thesis, Instituto Politécnico Nacional, Centro
Interdisciplinario de Ciencias Marinas, La Paz, B.C.S., 160 pp.
López-Martınez, J., Arreguın-Sánchez, F., Hernández-Vázquez, S., Garcıa-Juárez, A.
R., Valenzuela-Quiñonez, W. 2003. Interannual variation of growth of the
brown shrimp Farfantepenaeus californiensis and its relation to temperature.
Fish. Res. 61(1), 95-105.
López-Martínez, J., Rábago-Quiroz, C., Nevárez-Martínez, M. O., García-Juárez, A.
R., Rivera-Parra, G., & Chávez-Villalba, J. 2005. Growth, reproduction, and
size at first maturity of blue shrimp, Litopenaeus stylirostris (Stimpson,
1874) along the east coast of the Gulf of California, Mexico. Fish. Res. 71(1),
93-102.
López-Martínez, J., Vazquez, S. H., Valdivia, E. H., Romero, J. R., Chávez, E. A. 2010.
Influencia ambiental en la pesquería de camarón. DE MÉXICO, 115.
MacCall, A.D. 1976. Density dependence of catchability coefficient in the California
Pacific sardine, Sardinops sagax caerulea, purse seine fishery. CalijI Coop.
Oceanic Fish. Invest. Rep. 18, 136-148.

234
MacCall, A.D. 1990. Dynamic Geography of Marine Fish Populations. (Books in
recruitment fishery oceanography). Seattle: Washington Sea Grant
Program. 153 pp.
Macías-Regalado, E., & Calderón-Pérez, A. 1979. Talla de inmigración de postlarvas
de camarón al sistema lagunar Huizáche-Caimanero, Sinaloa,
México.(Crustacea, Decapoda, Penaeus). In: Anales del Instituto de
Ciencias del Mar y Limnología, Universidad Nacional Autónoma, México
(Vol. 6, No. 2, pp. 99-106).
Madrid-Vera, J., Chávez-Herrera, D., Melchor-Aragón, J., Meraz-Sánchez, R. &
Rodríguez-Preciado J.A. 2012. Management for the white shrimp
(Litopenaeus vannamei) from the southeastern Gulf of California through
biomass models analysis. O. J. Mar. Scien.
Magallón, F., Jaquemin, P. 1976. Observaciones biológicas sobre tres especies
comerciales de camarón en las costas de Sinaloa, Mex. In: Castro-Aguirre
J.L. (ed), Memorias del Simposio sobre Biología y Dinámica Poblacional de
Camarones. Instituto Nacional de Pesca, pp. (2): 1-26.
Manetsch, T.J. 1976. Time varying distributed delays and their use in aggregate models
of large systems. IEEE Transactions on Systems, Man and Cybernetics.
8:547-553.
Márquez, T.M. 1976. Captura y esfuerzo en pesquerías de camarón de aguas
interiores (Bahías Guasimas y lobos, Sonora). En: Castro-Aguirre J.L. (ed),
Memorias del Simposio sobre Biología y Dinámica Poblacional de
Camarones. Instituto Nacional de Pesca, pp. (1): 46-56.
Martínez-Aguilar, S., De Anda-Montañez, J. A., & Arreguín-Sánchez, F. 1997.
Densidad y “capturabilidad” de la sardina monterrey, Sardinops sagax
(Pisces: Clupeidae) del Golfo de California, México. Rev. Biol. Trop. 4(3),
45.

235
Martínez-Aguilar, S., Arreguín-Sánchez F., Morales-Bojórquez E. 2005. Natural
mortality and life history stage duration of Pacific sardine (Sardinops
caeruleus) based on gnomonic time divisions. Fish. Res. 71: 103-114.
Martínez-Aguilar, S., de Anda-Montañez, J. A., Arreguín-Sánchez, F., & Cisneros-
Mata, M. Á. 2009. Constant harvest rate for the Pacific sardine (Sardinops
caeruleus) fishery in the Gulf of California based on catchability-at-length
estimations. Fish. Res. 99(2), 74-82.
Martínez-Aguilar, S., Díaz-Uribe J.G., de Anda-Montañez J.A., Cisneros-Mata M.Á.
2010. Natural mortality and life history stage duration for the jumbo squid
(Dosidicus gigas) in the Gulf of California, Mexico, using the gnomonic time
division. Cienc. Pesq. 18: 31-42.
Mathews, C.P. 1981. A review of the North American penaeid fisheries, with particular
reference to Mexico. Kuwait Bull. Mar.Sci. 2: 325-409.
McGurk, M.D. 1986. Natural mortality of marine pelagic fish eggs and larvae: role of
spatial patchiness. MEPS. 34(3), 227-242.
Medina, A. S., & Soto, L. A. 2003. Assessment of the fishing effort level in the shrimp
fisheries of the Central and Southern Gulf of California. NAGA, WorldFish
Center Quarterly, 26(4), 16-20.
Mendoza, J. J., Marcano, L. A., Alio, J. J., & Arocha, F. 2010. Autopsia de la Pesquería
de Arrastre del Oriente de Venezuela: Análisis de los Datos de
Desembarques y Esfuerzo de Pesca. Proceedings of the 62nd Gulf and
Caribbean Fisheries Institute. GCFI. 69-76.
Menz, A., y Bowers, A.B. 1980. Bionomics of Penaeus vannamei Boone and Penaeus
stylirostris Stimpson in a lagoon on the Mexican Pacific coast. Est. Coas.
Mar. Scie. 10(6), 685-697.
Meraz-Sánchez, R., Madrid-Vera, J., & Herrera, D. C. 2013. An Approach to
Assessment to Population of the Brown Shrimp, Farfantepenaeus
californiensis (Holmes, 1900), as a Management Fisheries Tool in the

236
Southeastern Gulf of California. Open Journal of Marine Science. Vol.3
No.1.
Mohammed, H., & Ye, Y. 1999. An analysis of variation in catchability of green tiger
prawn, Penaeus semisulcatus, in waters off Kuwait. Fish. Bull. 97(3),
702Á712.
Morales-Bojórquez, E., López-Martínez, J., & Hernández-Vásquez, S. 2001. Modelo
dinámico de captura y esfuerzo para el camarón café Farfantepenaeus
californiensis (Holmes, 1900) del Golfo de California, México. Cienc. Mar.
27(1), 105-124.
Morales-Bojórquez, E., S. Martínez-Aguilar, F. Arreguín-Sánchez y M. Ramírez-
Rodríguez. 2003. GIM, programa computarizado para estimar la mortalidad
natural y duración de los estadios del ciclo de vida de recursos pesqueros,
basado sobre división de tiempo gnomonico. Centro Regional de
Investigación Pesquera de La Paz del INP, Centro Interdisciplinario de
Ciencias Marinas del IPN. 19p.
Muñoz-Rubí H.A., Chávez-Herrera D. y Villegas-Hernández F. 2012. Evaluación de las
poblaciones de Litopenaeus vannamei, L. stylirostris y Farfantepenaeus
californiensis en el sistema lagunar Huizache-Caimanero, Sinaloa, en 2011.
Informe técnico. INP-CRIP Mazatlán.
Murphy, G. 1965. A solution of the catch equation. J. Fish. Res. Bd Can. 22, 191-202.
Nance, J., Keithly Jr, W., Caillouet Jr, C., Cole, J., Gaidry, W., Gallaway, B. & Travis,
M. 2006. Estimation of effort, maximum sustainable yield, and maximum
economic yield in the shrimp fishery of the Gulf of Mexico. Report to the Gulf
of Mexico Fishery Management Council by The Ad Hoc Shrimp Effort
Working Group.
Navarrete del Proo, A.F. 2007. Análisis de la biología pesquera de camarón rosado
(Farfantepenaeus duorarum, Burkenroad, 1939), en la sonda de Campeche,
Campeche, México. Tesis doctoral, UNAM.

237
NOM-002-SAG/PESC-2013. 2013. NORMA Oficial Mexicana NOM-002-SAG/PESC-
2013, Para ordenar el aprovechamiento de las especies de camarón en
aguas de jurisdicción federal de los Estados Unidos Mexicanos. De:
https://www.gob.mx/conapesca/documentos/normas-oficiales-mexicanas-
pesqueras-y-acuicolas-30453
Núñez, R., Chapa H. y E. Chimal de Nuñez. 1951. La pesca del camarón por artes fijas
en os estados de Sinaloa y Nayarit: III. Datos sobre la producción – Estudio
comparativo de especies y tamaños de camarón que integran la producción
de aguas interiores y oceánicas. Contribuciones técnicas. Instituto de Pesca
del Pacífico A.C. México.
Olguín, M. 1967. Estudio de la biología del camarón café Farfantepenaeus
californiensis Holmes. FAO Fish. Rep. 57(2): 331-356.
Önal, H., McCarl, B. A., Griffin, W. L., Matlock, G., & Clark, J. 1991. A bioeconomic
analysis of the Texas shrimp fishery and its optimal management. Am. J.
Agr. Econ. 73(4), 1161-1170.
Paloheimo, J. E. 1958. A method of estimating natural and fishing mortalities. J. Fish.
Bo. Can. 15(4), 749-758.
Pauly, D. 1980. On the interrelationships between natural mortality, growth parameters,
and mean environmental temperature in 175 fish stocks. Journal Cons. 39:
175-192.
Pauly, D. 1981. The relationship between gill surface area and growth performance in
fishsh: a generalization of von Bertalanffy’s theory of growth.
Meeresforschung. 28, 251-282.
Pearse, P.H. 1980. Regulation of fishing effort. FAO. Fish. Tech. Pap. (197): 82 pp.
Pella, J.J.y P.K. Tomlinson, 1969 A generalized stock production model. Bull.I-ATTC,
13(3):421–52

238
Pérez-Farfante, I. 1988. Illustrated key to penaeoid shrimps of commerce in the
Americas. US Department of Commerce, National Oceanic and
Atmospheric Administration, National Marine Fisheries Service.
Pérez, E., & Defeo, O. 2003. Time-space variation in the catchability coefficient as a
function of catch unit of effort in Heterocarpus reedi (decapoda, pandalidae)
in north-central Chile. Interciencia. 28(3), 178-181.
Poli, C. R., & Calderón-Pérez, J. A. 1985. Efecto de los cambios hidrológicos en la
boca del río Baluarte sobre la inmigración de las postlarvas de Penaeus
vannamei Boone y P. stylirostris Stimpson al sistema lagunar Huizache-
Caimanero, Sin. México. An. Inst. Cienc. del Mar y Limnol., Universidad
Nacional Autónoma de México, 14(1), 29-44.
Pope, J.G. 1972. An investigation of accuracy of Virtual Population Analysis. Znt.
Comm. NW Atl. Fish. Res. Bull. 9, 65-74.
Pope, J.G. and Shepherd, J.G. 1985. A comparison of the performance of various
methods for tuning VPA using effort data. J. Cons. int. Explor Mer. 42, 129-
51.
Prahl, H.V., Gardeazabal M. 1977. Commercial culture of the blue shrimp, Litopenaeus
stylirostris Stimpson in Colombia. In: 8th Annual World Mariculture Society
Meeting, San Jose, Costa Rica, 9-13 January 1977. San Jose, Costa Rica.
pp. 9-13.
Prince, J. D. 1992. Using a spatial model to explore the dynamics of an exploited stock
of the abalone Haliotis rubra. Abalone of the world: biology, fisheries and
culture, 305-317.
Ramírez-Rodríguez, M., & Arreguín-Sánchez, F. 2003. Spawning stock-recruitment
relationships of pink shrimp Farfantepenaeus duorarum in the southern Gulf
of Mexico. Bull. Mar. Scienc. 72(1), 123-133.
Ramírez-Rodríguez, M., Arreguín-Sánchez F. 2003. Life history stage duration and
natural mortality for the pink shrimp Farfantepenaeus duorarum

239
(Burkenroad, 1939) in the southern Gulf of Mexico, using the gnomonic
model for time division. Fish. Res. 60: 45–51.
Ramos-Cruz, S. 2000. Composición por tallas, edad y crecimiento de Litopenaeus
vannamei (Natantia: Penaeidae), en la laguna Mar Muerto, Oaxaca-
Chiapas, México. Rev. Biol. Trop. 48(4), 873-882.
Ramos-Cruz, S. 2013. Evaluación de la pesquería artesanal de camarón en el sistema
lagunar La Pampita-Joya-Buenavista, Chiapas, México. Cienc. Pesq., 21(2),
5-11.
Ramos-Cruz, S., Sánchez-Meraz B., Carrasco-Ayuso F., Cervantes-Hernández, P.
2006. Estimación de la tasa de mortalidad natural de Farfantepenaeus
californiensis (Holmes, 1900) y Litopenaeus stylirostris (Boone, 1931) en la
zona costera del Golfo de Tehuantepec, México. Rev. Biol. Mar. Oceanogr.
41: 221-229.
Renfro, W.C. 1964. Life history stages of Gulf of Mexico brown shrimp. U. S. Fish and
Wildl. Serv., Cir. 183: 94-98.
Ricker, W.E. 1958. Handbook of computations for biological statistics offish
populations. Bull. Fish. Res. Bd. Can. 119: 1-300.
Ricker, W.E. 1975. Computation and interpretation of biological statistics of fish
populations. Bull. Fish. Res. Board Can. 191: 382.
Ricker, W.E., 1975 Computation and interpretation of biological statistics of fish
population. Bull.Fish.Res.Board Can. (191):382 p.
Rikhter, V.A., Efanov, V.N. 1976. On one of the approaches to estimation of natural
mortality of fish populations. ICNAF Res. Doc. 76/VI/8: 1-12 .
Rivera, G., Soto, L. A., Ugarte, I. H. S., & Naranjo, E. J. 2009. Assessment of an
artisanal shrimp fishery of Litopenaeus vannamei in a lagoon-estuarine
system based on the concept of maximum sustainable yield. Rev. Biol. Mar.
Ocean. 44(3), 635-646.

240
Rodríguez de la Cruz, M.C. 1976. Sinopsis biológica de las especies del género
Penaeus del Pacífico Mexicano. In: Castro-Aguirre J.L. (ed), Memorias del
Simposio sobre Biología y Dinámica Poblacional de Camarones. Instituto
Nacional de Pesca, pp. (2): 280-314.
Rodríguez de la Cruz, M.C. 1981. Aspectos pesqueros del camarón de alta mar en el
Pacífico Mexicano. Cien. Pesq. 1: 1-20.
Rodríguez de la Cruz, M.C., Rosales J.F. 1973. Sinopsis de Penaeus (Melicertus)
californiensis, Holmes. INO- CPPG. Serie técnica no 2.
Rodríguez de la Cruz, M.C., Rosales J.F. 1976. El camarón del noroeste de México.
INP Serie información. INP/SI: i48.
Rodríguez de la Cruz, M.C., Chávez E.A. 1994. La pesquería de camarón en alta mar.
Pacífico de México. In: XXX Aniversario del INP, Pesquerías Relevantes de
México. Secretaria de Pesca (ed). SEMARNAT, México. pp.11-37.
Romero-Moreno, G.E. 2009. Manejo de ecosistemas aplicado a la pesquería del
camarón. Tesis de Maestría. Universidad Nacional Autónoma de México.
115 p.
Romero-Sedano, J. C., E. A. Aragón-Noriega, M. M. Manzano-Sarabia, C. A. Salinas-
Zavala, A. R. García-Juárez. 2004. Reproductive period of brown shrimp
Farfantepenaeus californiensis (Holmes, 1900) in the Agiabampo coastal
lagoon system, Sonora/Sinaloa, México. Cienc. Mar. 30(3):465–475.
Rosas-Cota, J. A., García-Tirado, V. M., Gonzalez Camacho, J. R. 1996. Análisis de
la pesquería de camarón de altamar en San Felipe, BC durante la
temporada de pesca 1995-1996. Boletín CRIP Ensenada INP-SEMARNAP,
2, 23-30.
Ross-Terrazas, M.A. 1988. Evaluación poblacional de dos especies de camarón que
sostienen la pesquería de altamar en el norte y centro del Golfo de
California. B.S. thesis, Universidad Autónoma de Baja California Sur, Marine
Biology Department, La Paz, B.C.S., 70 pp.

241
Royer, J., Périès P., Robin J. P. 2002. Stock assessments of English Channel loliginid
squids: updated depletion methods and new analytical methods. J. Cons.
59: 445-457.
Rumrill, S.S. 1990. Natural mortality of marine invertebrate larvae. Ophelia. 32: 163-
198.
Sáenz-Martínez, P.G., Lluch-Belda, D. 1990. Análisis de una temporada de pesca de
camarón azul en alta mar. Serie documentos de trabajo año II, No. 28,
Instituto Nacional de la Pesca. 19 pp.
Sainsbury, K.J. 1980. Effect of individual variability on the von Bertalanffy growth
equation. Can. J. Fish. Aquat. Sci. 37: 241-247.
Salgado, B.R. (1976). Efecto d ela temperatura sobre la migración de Penaeus
stulirostris en el Estero Sta. Cruz, Sonora, durante 1975. In: Castro-Aguirre
J.L. (ed), Memorias del Simposio sobre Biología y Dinámica Poblacional de
Camarones. Instituto Nacional de Pesca. (1) pp. 372-374.
Schaefer, M.B., 1954 Some aspects of the dynamics of populations, important to the
management of the commercial marine fisheries. Bull.I-ATTC/Bol.CIAT,
1(2):56 p.
Schafer, H. 1971. Advances in Pacific shrimp culture, G.C.F.I. Proc. (23): 133-138.
Seijo, J. C., Defeo, O., & Salas, S. 1997. Bioeconomía pesquera. Teoría, modelación
y manejo. FAO: Documento Técnico de Pesca, (368).
Seijo, J.C. 1986. Comprenhensive simulation model of a tropical demersal fishery: red
grouper (Epinephelus morio) od the Yucatan continental Shaelf. Ph.D.
Dissertation, Michigan State University. 210pp.
Sepúlveda-Medina, A. 1976. Crecimiento y mortalidad de camarón blanco (Penaeus
vannamei Boone) en el sistema lagunar Huizache-Caimaero, Sin. durante
la temporada 1974-1975. En: Memorias del Simposio sobre Biología y
Dinámica de Poblaciones de Camarones. 1977. Tomo I: 1-12.

242
Sepúlveda-Medina, A. 1981. Estimación de la mortalidad natural y por pesca del
camarón blanco Litopenaeus stylirostris en el sistema lagunar Huizache–
Caimanero, Sin. Durante la temporada 76–77. Cienc. Pesq. 1: 71-90.
Sepúlveda Medina, A. 1991. Análisis biológico pesquero de los camarones peneidos
comerciales en el Pacifico mexicano durante el periodo de veda 1974-1983.
M. Sc. thesis, Universidad Nacional Autónoma de México, México, 154 pp.
Sepúlveda Medina, A. 1999. Dinámica poblacional de los peneidos comerciales en el
alto, centro golfo de California, Topolobampo y costa occidental de la Baja
California en el litoral del pacifico mexicano. Ph.D. thesis, Instituto
Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas, La Paz,
B.C.S., 147 pp.
Sepúlveda Medina, A., Soto L.A. 2001. Patterns of growth rates in the populations of
brown and blue shrimp (Farfantepenaeus californiensis, Holmes 1900;
Litopenaeus stylirostris , Stimpson 1871) in the northern Gulf of California.
Rev. Invest. Mar. 22(3):205-218.
Shepherd, J. G. 1987. Towards a method for short-term forecasting of catch rates
based on length composition. Length-based methods in fisheries research,
13. International Center for Aquatic Resources Management (ICLARM),
Manila, Philippines; and KISR, Kuwait, 167776.
Sierra-Rodríguez, P. y Reyna-Cabrera, 1. E. 1993. Análisis de la pesquería del
camarón en el Golfo de Tehuantepec (con énfasis en la veda del camarón
de lagunas costeras). Informe técnico interno. Centro Regional de
Investigación Pesquera, Salina Cruz, Oaxaca. 40 p.
Sierra, P., C. Acosta, J. A. García, A. R. García, A. Liedo, J. M. Melchor, S. Ramos, A.
Rosas, M. P. Toledo, and E. Zárate. 2000. Camarón del Océano Pacífico.
119-158. In Cisneros-Mata, M. A. Sustentabilidad y pesca responsable en
México, evaluación y manejo 1999–2000. Instituto Nacional de Pesca,
SEMARNAP, México. 1046 pp.

243
Silliman, R. P. 1943. Studies on the Pacific pilchard or sardine (Sardinops caerulea).
6. Thermal and diurnal changes in the vertical distribution of eggs and
larvae. Spec. Scient. Rep., US Fish Wildl. Serv.,(22), 1-17.
Sissenwine, M.P. 1978. Is MSY an adequate foundation for optimum yield?. Fisheries.
3(6), 22-42.
Soto-López, R. 1973. Complemento al informe de los Muestreos de Camarón en los
esteros del sur de Sinaloa temporada 1973. Informe técnico. INP/SI:19.
Tabash Blanco, F. A. 2007. Explotación de la pesquería de arrastre de camarón
durante el período 1991-1999 en el Golfo de Nicoya, Costa Rica. Rev. Biol.
Trop. 55(1), 207-218.
Thompson, W.F. y F.A. Bell, 1934 Biological statistics of the Pacific halibut fishery. Part
2. Effects of changes in intensity upon total yield and yield per unit of gear.
Rep.Int.Pac.Halibut Comm., (8):49 p.
Thorson, G. 1966. Some factors influencing the recruitment and establishment of
marine benthic communities. Neth. J. Sea Res. 3(2): 267-293.
Tian, X., Leung, P., & Hochman, E. 1993. Shrimp growth functions and their economic
implications. Aquac. Eng. 12(2), 81-96.
Torres-Acuña, I. 2008. Bioenergética de larvas zoea de camarón blanco Litopenaeus
stylirostris alimentadas con diferentes raciones de microalga
Phaeodactylum tricornutum. B.S. thesis, Universidad Michoacana de San
Nicolas de Hidalgo, Michoacán, 54 pp.
Valles-Jiménez, R., Cruz, P., & Perez-Enriquez, R. (2004). Population genetic structure
of Pacific white shrimp (Litopenaeus vannamei) from Mexico to Panama:
microsatellite DNA variation. Mar. Biotec. 6(5), 475-484.
Vázquez, F.M. 2011. Variaciones interanuales en la abundancia de postlarvas de los
generos Litopenaeus y Farfantepenaeus en la desembocadura del rio
presidio y boca del puerto de Mazatlán Sinaloa 2007, 2008 y 2009. Tesis de
Licenciatura. UAS.

244
Vetter, E.F. 1988. Estimation of natural mortality in fish stocks: a review. Fish. Bull. 86:
25-43.
Ye, Y. 2000. Is recruitment related to spawning stock in penaeid shrimp fisheries?.
ICES Journal of Marine Science: Journal du Conseil, 57(4), 1103-1109. Ye,
Y. (2000). Is recruitment related to spawning stock in penaeid shrimp
fisheries?. ICES J. Mar. Sci: J. C. 57(4), 1103-1109.


















