
DETERMINACIÓN DE LA VIDA DE ANAQUEL DE...
72
Transcript of DETERMINACIÓN DE LA VIDA DE ANAQUEL DE...


DETERMINACIÓN DE LA VIDA DE ANAQUEL DE ALIMENTOS CON ALTO CONTENIODO DE CARBOHIDRATOS
Belmares J.L., Amaya Guerra, C.A. Espinoza Mata, A., Núñez González M. A., Báez González,
J.G. Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León. Av. Pedro de Alba, Ciudad Universitaria. C.P. 66451. San Nicolás de los Garza, N.L. México. * [email protected] RESUMEN: La vida de anaquel de productos alimenticios ha sido calculada por métodos diversos entre los que destacan el análisis sensorial y la observación de su deterioro fisicoquímico al final de un período de almacenamiento. Las pruebas de laboratorio permiten acelerar el proceso de alteración, mediante la aplicación de calor en períodos de tiempo establecidos y bajo condiciones cuidadosamente supervisadas, por lo cual para determinar el tiempo de vida útil de caramelo, mermelada de fresa y miel de abeja (con porcentaje alto de carbohidratos), así como de cacahuate, fríjol cocido y salchicha (con porcentaje alto de proteínas) se sometieron a temperaturas superiores a las de almacenamiento tradicional. Así, los primeros alimentos se expusieron a 35°C, 45°C y 55°C, mientras que los segundos a 8°C, 13°C y 18ºC; La calidad de las muestras almacenadas fue monitoreada por un panel sensorial semientrenado; y mediante análisis de pérdida de agua, viscosidad, pH, e índice de peróxido. Las cinéticas de deterioro de los alimentos con alta concentración de carbohidratos nos indicaron un valor promedio de Q10 de 1.58 para el caramelo, de 2 para mermelada y de 1.75 para miel de abeja, lo que nos indica una vida útil de 70 días, 32 días y 42 días, respectivamente, a una temperatura de 25ºC. En el caso de los alimentos con alta concentración de proteínas las cinéticas de deterioro nos indicaron un valor promedio de Q10 de 1.41 para el cacahuate, de 2.05 para salchicha y de 2.10 para el fríjol cocido, en cuyo caso les corresponde una vida útil de 56 días, 20 días y 24 días, respectivamente, a 25ºC para el primero y a 4ºC para los últimos dos. En el caramelo, el análisis sensorial fue significativo para predecir la vida útil; en la mermelada de fresa y miel de abeja, el análisis sensorial y la viscosidad son parámetros que se pueden emplear para determinar la VA. La evaluación sensorial y el índice de peróxido son dos parámetros significativos para determinar la vida útil en cacahuate tostado; en salchicha y fríjol cocido el pH y el índice de peróxido son dos parámetros relevantes que pueden usarse para determinar la vida útil. ABSTRACT: The life of shelf of nutritious products has been calculated by diverse methods among those that highlight the sensorial analysis and the observation from its physiochemical deterioration to the end of a period of storage. The laboratory tests allow to accelerate the alteration process, by means of the application of heat in established periods of time and I lower carefully supervised conditions, reason why to determine the time of useful life of candy, strawberry marmalade and bee honey (with high percentage of carbohydrates), as well as of peanut, cooked fríjol and sausage (with high percentage of proteins) they underwent superior temperatures to those of traditional storage. This way, the first foods were exposed at 35°C, 45°C and 55°C, while the seconds at 8°C, 13°C and 18ºC; The quality of the stored samples was monitoreada for a panel sensorial semientrenado; and by means of analysis of loss of water, viscosity, pH, and index of peroxide. The kinetic of deterioration of the foods with high concentration of carbohydrates indicated us a value average of Q10 1.58 for the candy, of 2 for marmalade and of 1.75 for bee honey, what indicates us an useful life of 70 days, 32 days and 42 days, respectively, to a temperature of 25ºC. In the case of the foods with high concentration of proteins the kinetic of deterioration indicated us a value average of Q10 1.41 for the peanut, of 2.05 for sausage and of 2.10 for the cooked fríjol, in which case it

corresponds them an useful life of 56 days, 20 days and 24 days, respectively, at 25ºC for the first one and at 4ºC for the last ones two. In the candy, the sensorial analysis was significant to predict the useful life; in the strawberry marmalade and bee honey, the sensorial analysis and the viscosity they are parameters that can be used to determine the VA. The sensorial evaluation and the index of peroxide are two significant parameters to determine the useful life in toasted peanut; in sausage and cooked fríjol the pH and the index of peroxide are two outstanding parameters that can be used to determine the useful life. Palabras clave: Anaquel, carbohidratos, sensorial. INTRODUCCIÓN En el desarrollo de cualquier producto alimenticio incluyendo la reformulación, el cambio del empaquetado o la condición de almacenamiento/ distribución, un aspecto importante que hay que conocer, es la vida de anaquel (VA). La vida útil de un producto alimenticio es vital para su éxito en el mercado y por lo menos debe exceder el tiempo mínimo requerido para la distribución del procesador al consumidor. Los encargados de calificar el producto también necesitan datos confiables de ésta para colocar los productos y establecer la marca de fábrica. La determinación periódica de la VA ayuda a proporcionar seguridad de que el producto restante será constante en un cierto plazo con respecto a su calidad (Fritsch, 1997). Dentro de los factores que contribuyen en la búsqueda para mejorar la vida útil de los alimentos se incluye su disponibilidad para el consumo a lo largo de todo el año con la máxima frescura y calidad, así como la continua globalización de los sistemas de distribución, que ha puesto la presión en el sector alimenticio de asegurar tiempos de estabilidad y de almacenaje de estante para que los productos viajen más y más lejos de su lugar de origen. La vida de anaquel de productos alimenticios ha sido calculada por métodos diversos entre los que destacan el análisis sensorial y la observación de su deterioro al final de un período de almacenamiento. En los últimos 20 años se ha desarrollado un interés importante en la implementación de técnicas que permitan predecir la vida de anaquel de alimentos con períodos largos de vida útil; actualmente existen suficientes bases teóricas que sustenten y validen la aplicación de diversos métodos. Desafortunadamente, la predicción de la vida de anaquel de cualquier producto alimenticio después de su manufactura resulta difícil de obtener, debido a la carencia de información disponible sobre las condiciones ambientales durante su distribución y en algunas ocasiones también sobre el lugar de almacenamiento. Sin embargo, algunos fabricantes de productos alimenticios manufacturados graban en sus empaques datos que le permiten conocer al cliente la caducidad o vida de anaquel aproximada del producto al momento de su compra (Jamieson, 1974). Los tres grupos constitutivos principales en los alimentos: carbohidratos, grasas y proteínas, son materias complejas construidas con unidades químicas sencillas: azúcares, ácidos grasos y aminoácidos. La cantidad y clase de estas materias constitutivas presentes en un alimento dado determinarán la naturaleza y el valor nutritivo del mismo y, además, pueden determinar la tendencia del alimento a experimentar cambios químicos deteriorantes después de la elaboración y durante el almacenamiento (Jamieson, 1974). METODOLOGÍA Selección y preparación de las muestras

Se seleccionaron 6 tipos de alimentos para este estudio. Tres de ellos correspondientes a miel de abeja, caramelo y mermelada de fresa representantes del grupo de alimentos con alta concentración de carbohidratos. Los otros tres son los de alto valor proteico y se incluyeron cacahuate, salchicha y fríjol cocido. A partir de una muestra de mayor de cada alimento, en el caso de miel de abeja se realizaron 56 submuestras de 170 g para determinar viscosidad y 27 de 90 g para análisis sensorial. Para el caramelo se realizaron 60 submuestras de 12.5 g para determinar pérdida de agua y otras 36 de 80 g para llevar a cabo el análisis sensorial. En el caso de la mermelada se estudiaron 56 submuestras de 170 g para determinar viscosidad, 54 de 90 g para análisis sensorial y 51 de 100 g para determinar la pérdida de agua. Para los alimentos de naturaleza proteica como el cacahuate se trabajaron con 39 submuestras de 100 g cada una para la determinación de peróxidos y otras 33 submuestras con la misma cantidad para el análisis sensorial. En el caso de la salchicha se determinó el índice de peróxidos en 28 submuestras de 100 g cada una, las cuales primeramente pasaron por un proceso de quitar humedad (secado) para realizar la extracción de la grasa usando el método Soxhlet y así poder cuantificar el índice de peróxidos y en 42 submuestras de 90 g se llevó a cabo la evaluación organoléptica y la determinación de pH; mientras que en el fríjol cocido se utilizaron 84 bolsas cada una con 490 g, las cuales a su vez fueron sometidas de igual manera que en la salchicha a un proceso de secado y así poder realizar la extracción de la grasa por el método Soxhlet y posteriormente con está grasa determinar el índice de peróxidos, también se utilizaron 39 submuestras de 90 g para su evaluación organoléptica y determinación de pH.
B) Temperaturas de almacenamiento El fundamento de la predicción de vida de anaquel por extrapolación o interpolación, se basa en ensayos a altas temperaturas, para lo cual se necesitaban al menos 3 temperaturas de experimentación por encima de las temperaturas registradas en almacén; se decidió conveniente usar temperaturas de ensayo altas para predecir por extrapolación, de tal manera que las muestras con alto contenido de carbohidratos y la de cacahuate tostado se almacenarán a temperaturas de 35, 45 y 55ºC, en tres diferentes incubadoras ajustadas a dicho parámetro, cada semana se analizaron por triplicado cada muestra para cada una de las pruebas establecidas. Los alimentos proteicos como salchicha y fríjol cocido se almacenaron a temperaturas de 8, 13 y 18ºC, durante el tiempo que duraron estables según la carga microbiana y los cambios fisicoquímicos que se presentaron.
C) Parámetros para determinar vida de anaquel
Análisis sensorial La calidad sensorial fue evaluada por 6 panelistas semientrenados para identificar los cambios organolépticos originados en cada temperatura a la que fueron sometidos cada uno de los alimentos. Ésta se realizó empleando una prueba triangular, la cual pertenece a un método analítico por diferenciación, cuyo objetivo fue determinar si existe diferencia sensorialmente perceptible entre dos muestras, comparando tres muestras a la vez, de las cuales dos son iguales entre sí (sometidas a las temperaturas ya mencionadas, según sea el caso) y la otra diferente mantenida a temperatura adecuada de anaquel que sirvió como control. Las muestras se codificaron en 3 grupos (A, B y C) y cada grupo con 3 números diferentes. Éste análisis sensorial

se evaluó en cada temperatura de almacenaje de los alimentos con el propósito de observar en que días por efecto de las temperaturas se aceleran más rápido las reacciones y éstas se hacen presentes en el deterioro de la calidad sensorial de los alimentos y fue aplicada como lo muestra la siguiente tabla. Tabla 1. Aplicación de la Prueba Triangular en los alimentos con alto contenido de carbohidratos y con alto contenido de proteínas. Alimento Temperatura de
almacenaje Días para la determinación de la evaluación sensorial
Duración del análisis sensorial en días
Miel de abeja 35ºC 7 28
Miel de abeja 45ºC 7 21 Miel de abeja 55ºC 7 14 Caramelo 35ºC 7 35 Caramelo 45ºC 7 28 Caramelo 55ºC 7 14 Mermelada 35ºC 4 32 Mermelada 45ºC 4 28 Mermelada 55ºC 4 16 Cacahuate 35ºC 7 35 Cacahuate 45ºC 7 28 Cacahuate 55ºC 7 21 Salchicha 8ºC 4 8 Salchicha 13ºC 4 8 Salchicha 18ºC 4 8 Fríjol 8ºC 4 12 Fríjol 13ºC 4 8 Fríjol 18ºC 4 8 Determinación de pérdida de agua. Esta prueba se llevó a cabo cada 4 días en el caramelo y la mermelada de fresa, en el primero se determinó por 32 días a 35ºC, 32 días a 45ºC y 16 días a 55ºC, mientras que en el segundo se analizó durante 28, 24 y 16 días a temperaturas de 35, 45 y 55ºC respectivamente. La pérdida de agua, se determinó por diferencia de peso de la muestra misma que se procesó después de extraerla de su temperatura de incubación, se dejó temperar por 30 minutos a temperatura ambiente. Se pesó el recipiente con la muestra misma que se vació en una toalla absorbente de peso conocido y ésta se colocó en una balanza analítica para dejar reposar por 6 min al cabo de los cuales se extrajo; se colocó una nueva toalla en la balanza y se vació el alimento de la toalla anterior a ésta para determinar el peso nuevamente. La diferencia de pesos nos indicó el agua que la muestra liberó. Determinación de viscosidad. La medición de la viscosidad se realizó a la mermelada de fresa y a la miel de abeja cada 7 días, la cual se cuantificó durante 28 días a 35ºC; 21 días a 45ºC y 14 días a 55ºC en cada alimento. La prueba se realizó utilizando el aparato Rheometer Brookfield DV-III Ultra, el cual consta con un sistema de cono-plato y se determinó en unidades de medición de centipoise (Pascal) Determinación de pH Se realizó cada 2 días en las muestras de salchicha y fríjol cocido, las lecturas de la salchicha fueron por 10, 8 y 8 días a temperaturas de 8, 13 y 18ºC respectivamente, mientras que en el fríjol

se determinaron durante 12, 6 y 6 días a temperaturas de 8,13 y 18ºC; para lo cual se pesaron 10 g de muestra que serán homogeneizados con 90 ml de agua destilada pH 7 y la mezcla obtenida se almacenó en refrigeración durante 1 hora para posteriormente realizar la lectura a temperatura de 25ºC en potenciómetro Corning 530. Determinación de índice de peróxidos. Se realizó cada 5 días en la salchicha y fríjol cocido, el primer alimento se analizó durante 10 días correspondiéndole las temperaturas de 8, 13 y 18ºC; en el segundo la determinación fue por 15, 10 y 10 días a temperaturas de 8, 13 y 18ºC, respectivamente. Otro alimento en el que se aplicó ésta prueba es el cacahuate, en donde se cuantificó cada 7 días durante 35, 28 y 21 días a temperaturas de 35, 45 y 55ºC, respectivamente. La prueba del índice de peróxidos fue determinado por el método de la AOAC.
D) Cálculos para predecir la vida de anaquel
a) Determinación del valor Q10 La expresión Q10 ( coeficiente de temperatura) empleada para expresar la razón entre índices de reacción de dos o más temperaturas separadas exactamente por 10ºC y que se puede aplicar convenientemente para que defina los efectos de la temperatura en las diversas reacciones químicas que pueden tener lugar en alimentos (Jamieson, 1974), se realizó en todas las muestras para conocer la velocidad de reacción al deterioro en relación con el incremento de la temperatura; para ello se efectuaron extrapolaciones de acuerdo a los días en que se presentó el deterioro fisicoquímico en los alimentos sometidos a las temperaturas y períodos establecidos. Este valor se obtuvo una vez realizado el análisis estadístico (ANOVA) el cual nos indicó si existe o no una alta diferencia significativa entre las medias de cada una de las determinaciones en las pruebas de pérdida de agua, viscosidad, pH e índice de peróxidos. El valor de Q10 se obtuvo comparando el resultado promedio con respecto a los días de las determinaciones y las temperaturas a las que fueron sometidos los alimentos, y se realizó dividiendo Q10/ Q10. b) Determinación del valor D Nos indica el tiempo necesario para el deterioro de los alimentos en su composición química para que su valor D descienda un ciclo logarítmico (reducción de la décima parte) según las temperaturas de almacenamiento. Se llevó a cabo en base a la formula siguiente log Cf / Co = T / D, donde Cf es la concentración final, Co es la concentración inicial, T son los días o tiempo de vida útil y D es la variable a calcular. c) Determinación del valor Z Éste valor es el número de grados centígrados que es preciso elevar la temperatura del alimento para que su valor D descienda un ciclo logarítmico (se reduzca la décima parte). Se llevó a cabo en base a la formula siguiente log D / D0 = T0-T / Z, donde D0 y D son valores obtenidos de D, y en donde Z expresa la dependencia que tienen de la temperatura los cambios enzimáticos, físicos y bioquímicos.
E) Análisis estadístico Todos los tratamientos incluyeron 3 repeticiones que fueron expresadas en el valor medio de cada observación durante ciertos días de almacenaje. Se realizó usando el paquete Statistical Package for the Social Sciences (SPSS), en el cual se aplicó un análisis de varianza (ANOVA) y prueba de Tukey para el caso de los resultados obtenidos en las determinaciones de pérdida de agua, viscosidad, pH, peróxidos y para la evaluación sensorial se utilizó la prueba de chi-cuadrada.

RESULTADOS Y DISCUSIÓN A) Alimentos con alto contenido de carbohidratos 1. Caramelo Análisis sensorial Al determinar la evaluación sensorial mediante la prueba triangular, los 6 panelistas lograron detectar cambios en los atributos, éstos fueron principalmente relacionados con la disminución de los sabores fresa y ácido; además de la presencia del fenómeno de la sinéresis y de cristalización de azúcares. Después de obtener las afirmaciones y comentarios por parte de los jueces, se realizó un análisis estadístico, con un análisis de Chi-cuadrada (Tabla 2), donde se muestra que existe una alta dependencia significativa en relación a las evaluaciones sensoriales a 35 y 45ºC durante 35 y 21 días, respectivamente, mientras que a 55ºC durante 14 días no se presentó tal efecto. El deterioro organoléptico de acuerdo al tiempo (días) y temperaturas de almacenamiento indicó un incremento en los cambios de los atributos con un valor promedio de Q10 de 1.58. Considerando que la literatura marca una vida útil de los caramelos entre 1 y 2 años, dependiendo de su composición química nuestra muestra, según los datos registrados en las evaluaciones sensoriales en cada una de las temperaturas de almacenaje, tendría una vida útil de 70 días a 25ºC, lo que nos parece poco tomando en consideración su composición y que es un alimento que presenta una baja aw. Esto nos conduce a pensar que los caramelos debieron haberse procesado en su empaque normal y distribuidos en menor cantidad en los frascos, ya que al analizar las submuestras almacenadas a las mencionadas temperaturas, éstas presentaban el fenómeno sinéresis y se pegaban entre si, efecto que detectaron los evaluadores, sobre todo cuando fueron comparados con el control. Tabla 2. Resultado del análisis por Chi-cuadrada de la evaluación sensorial de caramelo. Chi-
cuadrada
Alimento Temperaturas (ºC) Días (Vida útil) Q10 N Valor Sig *
Caramelo
35 35 90 13.962 0.007 ***
45 21 1.58 72 13.784 0.003 ***
55 14 36 0.450 0.502 *
* Valores de Sig > 0.05 indican que no existe una dependencia significativa. ** Valores de Sig ≤ 0.05 indican que existe una dependencia significativa. *** Valores de Sig ≤ 0.01 indican una alta dependencia significativa. Pérdida agua
El comportamiento del caramelo con respecto a la pérdida de agua (g) mostró el efecto de las temperaturas sobre los caramelos durante el tiempo de almacenamiento. Los resultados promedios obtenidos de la pérdida de agua expresados en gramos (Figura 1) se aprecian variaciones en cuanto a la pérdida y ganancia de humedad, los cuales se determinaron cada 4 días durante 32 días a temperaturas de 35º y 45ºC y por 16 días a 55ºC. Esto es lógico debido a que se trabajo con submuestras escogidas al azar, lo cual comprobamos con el análisis estadístico (Tabla 3), donde se muestra que no se obtuvo una varianza (F) y por consiguiente no hay ninguna significancia entre las medias de cada temperatura durante los días de almacenamiento.

-0.2333
-0.1333
0.4000
-0.1667 -0.1667
0.0000
0.1667
-0.1000-0.0667
-0.1000
-0.3333
0.0000
0.1667
-0.0667
0.1334
-0.0333-0.03330.0000
0.2000
-0.0333
-0.4000
-0.3000
-0.2000
-0.1000
0.0000
0.1000
0.2000
0.3000
0.4000
0.5000
0 4 8 12 16 20 24 28 32 36
Pérdida agua (g)
Días
35º C
45º C
55º C
Figura 1. Representación del comportamiento de la pérdida de agua (g) del caramelo frente al tiempo a diversas temperaturas. Tabla 3. Resultado estadístico correspondiente a la pérdida de agua del caramelo. Alimento Tratamiento Temperaturas
(ºC)
Días
(vida útil)
n Media (*) F Sig *
Caramelo
Pérdida Agua
35 32 24 -0.029
45 32 24 -0.1875 - -
55 16 12 0.0334
* Valores de Sig > 0.05 indican que no existe una diferencia significativa. ** Valores de Sig ≤ 0.05 indican que existe una diferencia significativa. *** Valores de Sig ≤ 0.01 indican una alta diferencia significativa. a-d Las medias son diferentes, prueba Tukey. 2. Mermelada de fresa
Análisis sensorial
Las aseveraciones de los evaluadores confirmaron que entre más alta es la temperatura de almacenaje el período de vida útil es más corto; esto lo podemos afirmar ya que los cambios sensoriales durante las temperaturas de almacenaje a 35, 45 y 55ºC empezaron a manifestarse a los 16, 8 y 4 días, respectivamente. Las reacciones que se presentaron por efecto de las temperaturas y que fueron detectadas por los panelistas fueron: sinéresis, reacción de Maillard, caramelización, disminución en los sabores dulce y fresa así como pérdida en la consistencia. El incremento de los cambios (Q10) en la calidad de la mermelada de fresa lo representamos con un valor de 2. (Tabla 4) nos indica en relación a la prueba Chi-cuadrada que existe una alta diferencia significativa a 35 y 45ºC, mientras que a 55ºC solo se presentó diferencia significativa, ésta se refiere a las afirmaciones correctas e incorrectas en la prueba triangular que se realizó en cada temperatura. La vida media de una mermelada es de aproximadamente un año y medio, en nuestro caso y, tomando en consideración los cambios presentados en cada temperatura de almacenamiento, la vida útil a 25ºC será de 32 días, lo que nos parece muy poco, tomando en

cuenta de que la mermelada es un alimento de larga durabilidad y además de que tiene un bajo aw, por lo tanto el manejo o la distribución equitativa en los envases de plástico que nosotros le dimos no fue el apropiado ya que ésta se homogeneizo al distribuir en los frascos y esta manipulación pudo haber interferido en la modificación de algunas propiedades reológicas del alimento, las cuales sufrieron un daño considerable además de la consecuente aparición de la reacción de Maillard y otros cambios percibidos por los panelistas. Por lo que se sugiere que al realizar el método para determinar vida de anaquel en mermeladas, se utilicen muestras enteras, no extraídas; o como lo hemos mencionado, no trabajar con submuestras. Tabla 4. Resultado del análisis por Chi-cuadrada de la evaluación sensorial de mermelada de fresa.
Chi-
cuadrada
Alimento Temperaturas (º C) Días (vida útil) Q10 n Valor Sig *
Mermelada
35 16 144 30.341 0.000 ***
45 8 2 126 30.989 0.000 ***
55 4 72 9.391 0.025 **
* Valores de Sig > 0.05 indican que no existe una dependencia significativa. ** Valores de Sig ≤ 0.05 indican que existe una dependencia significativa. *** Valores de Sig ≤ 0.01 indican una alta dependencia significativa. b) Pérdida agua
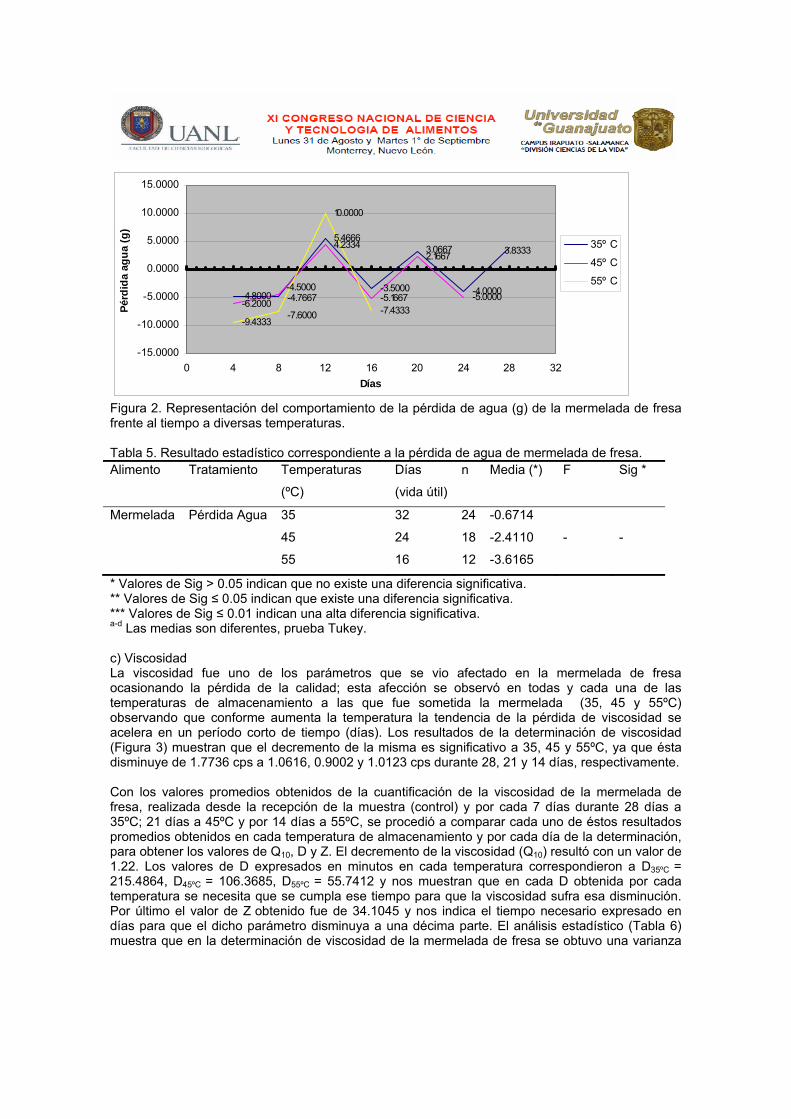
Con respecto a la pérdida de agua en la mermelada de fresa llevada a cabo cada 4 días durante 32 días a 35°C, por 28 días a 45°C y 16 a 55°C se obtuvieron promedios de este valor que nos indicaron variaciones en cuanto a la ganancia y a la pérdida de humedad lo que nos muestra que hay diferencias provocadas por el uso de submuestras al azar (Figura 2); con estos resultados promedios se realizó un análisis de estadístico (Tabla 5), en donde no se obtuvo una varianza (F) ni la significancia de las medias de las temperaturas en los días de almacenamiento.
-4.8000
5.4666
-3.5000 -4.0000-6.2000
4.2334
-5.1667
2.1667
-5.0000
-9.4333
10.0000
-7.4333-4.7667
3.0667 3.8333
-4.5000
-7.6000
-15.0000
-10.0000
-5.0000
0.0000
5.0000
10.0000
15.0000
0 4 8 12 16 20 24 28 32Días
Pérd
ida
agua
(g)
35º C
45º C
55º C
Figura 2. Representación del comportamiento de la pérdida de agua (g) de la mermelada de fresa frente al tiempo a diversas temperaturas. Tabla 5. Resultado estadístico correspondiente a la pérdida de agua de mermelada de fresa. Alimento Tratamiento Temperaturas
(ºC)
Días
(vida útil)
n Media (*) F Sig *
Mermelada
Pérdida Agua
35 32 24 -0.6714
45 24 18 -2.4110 - -
55 16 12 -3.6165
* Valores de Sig > 0.05 indican que no existe una diferencia significativa. ** Valores de Sig ≤ 0.05 indican que existe una diferencia significativa. *** Valores de Sig ≤ 0.01 indican una alta diferencia significativa. a-d Las medias son diferentes, prueba Tukey. c) Viscosidad La viscosidad fue uno de los parámetros que se vio afectado en la mermelada de fresa ocasionando la pérdida de la calidad; esta afección se observó en todas y cada una de las temperaturas de almacenamiento a las que fue sometida la mermelada (35, 45 y 55ºC) observando que conforme aumenta la temperatura la tendencia de la pérdida de viscosidad se acelera en un período corto de tiempo (días). Los resultados de la determinación de viscosidad (Figura 3) muestran que el decremento de la misma es significativo a 35, 45 y 55ºC, ya que ésta disminuye de 1.7736 cps a 1.0616, 0.9002 y 1.0123 cps durante 28, 21 y 14 días, respectivamente. Con los valores promedios obtenidos de la cuantificación de la viscosidad de la mermelada de fresa, realizada desde la recepción de la muestra (control) y por cada 7 días durante 28 días a 35ºC; 21 días a 45ºC y por 14 días a 55ºC, se procedió a comparar cada uno de éstos resultados promedios obtenidos en cada temperatura de almacenamiento y por cada día de la determinación, para obtener los valores de Q10, D y Z. El decremento de la viscosidad (Q10) resultó con un valor de 1.22. Los valores de D expresados en minutos en cada temperatura correspondieron a D35ºC = 215.4864, D45ºC = 106.3685, D55ºC = 55.7412 y nos muestran que en cada D obtenida por cada temperatura se necesita que se cumpla ese tiempo para que la viscosidad sufra esa disminución. Por último el valor de Z obtenido fue de 34.1045 y nos indica el tiempo necesario expresado en días para que el dicho parámetro disminuya a una décima parte. El análisis estadístico (Tabla 6) muestra que en la determinación de viscosidad de la mermelada de fresa se obtuvo una varianza
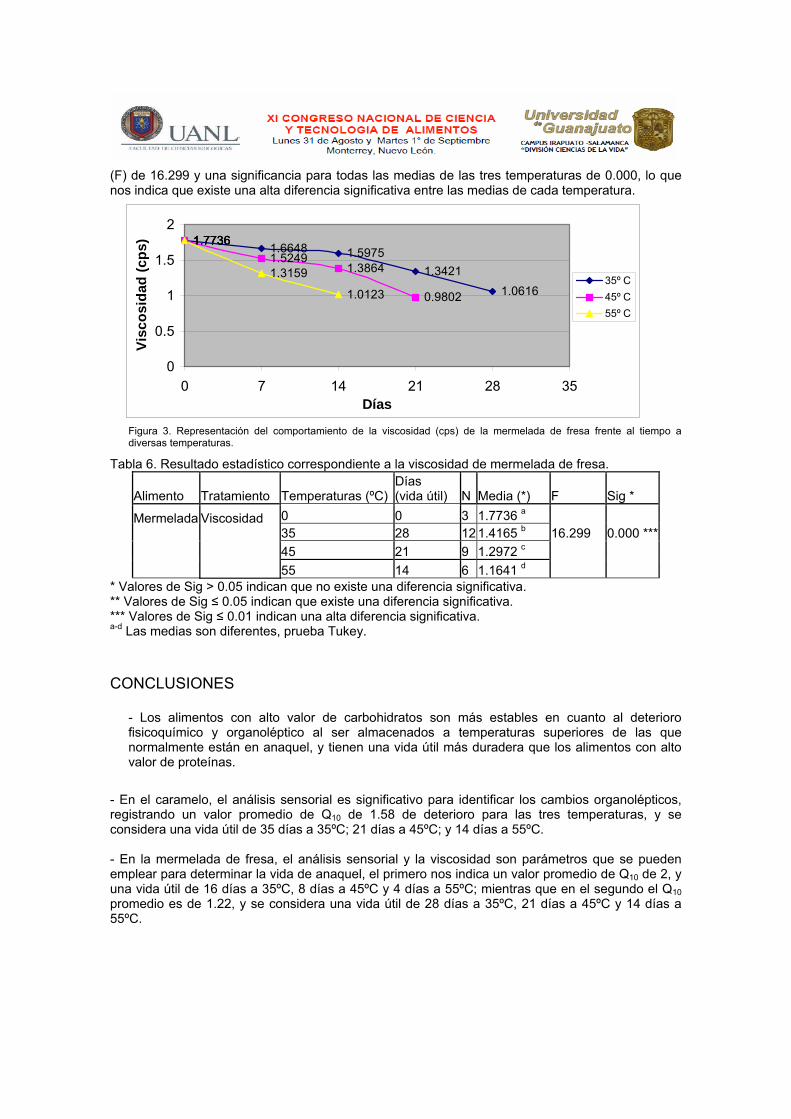
1.7736 1.6648 1.59751.3421
1.0616
1.77361.5249
1.3864
0.9802
1.7736
1.3159
1.0123
0
0.5
1
1.5
2
0 7 14 21 28 35Días
Visc
osid
ad (c
ps)
35º C45º C55º C
(F) de 16.299 y una significancia para todas las medias de las tres temperaturas de 0.000, lo que nos indica que existe una alta diferencia significativa entre las medias de cada temperatura.
Figura 3. Representación del comportamiento de la viscosidad (cps) de la mermelada de fresa frente al tiempo a diversas temperaturas.
Tabla 6. Resultado estadístico correspondiente a la viscosidad de mermelada de fresa.
Alimento Tratamiento Temperaturas (ºC)Días (vida útil) N Media (*) F Sig *
Mermelada
Viscosidad
0 0 3 1.7736 a 35 28 12 1.4165 b 16.299 0.000 ***
45 21 9 1.2972 c
55 14 6 1.1641 d
* Valores de Sig > 0.05 indican que no existe una diferencia significativa. ** Valores de Sig ≤ 0.05 indican que existe una diferencia significativa. *** Valores de Sig ≤ 0.01 indican una alta diferencia significativa. a-d Las medias son diferentes, prueba Tukey. CONCLUSIONES
- Los alimentos con alto valor de carbohidratos son más estables en cuanto al deterioro fisicoquímico y organoléptico al ser almacenados a temperaturas superiores de las que normalmente están en anaquel, y tienen una vida útil más duradera que los alimentos con alto valor de proteínas.
- En el caramelo, el análisis sensorial es significativo para identificar los cambios organolépticos, registrando un valor promedio de Q10 de 1.58 de deterioro para las tres temperaturas, y se considera una vida útil de 35 días a 35ºC; 21 días a 45ºC; y 14 días a 55ºC. - En la mermelada de fresa, el análisis sensorial y la viscosidad son parámetros que se pueden emplear para determinar la vida de anaquel, el primero nos indica un valor promedio de Q10 de 2, y una vida útil de 16 días a 35ºC, 8 días a 45ºC y 4 días a 55ºC; mientras que en el segundo el Q10 promedio es de 1.22, y se considera una vida útil de 28 días a 35ºC, 21 días a 45ºC y 14 días a 55ºC.

- Los mismos parámetros que en el punto anterior, se consideraron para determinar la vida útil en miel de abeja. El análisis sensorial nos indicó un incremento en el deterioro con un valor promedio de Q10 calculado de 1.75, y una vida útil de 21 días a 35º C, 14 días a 45ºC y 7 días a 55ºC; mientras que en la viscosidad valor promedio de Q10 obtenido fue 1.28, y se supone una vida útil de 28 días a 35ºC, 21 días a 45ºC y 14 días a 55ºC. - La evaluación sensorial y el índice de peróxido son dos parámetros significativos para determinar la vida útil en cacahuate tostado, el primero nos muestra un valor promedio de Q10 de 1.41, con una vida útil de 28 días a 35º C, 21 días a 45ºC y 14 días a 55ºC; el segundo nos reveló un valor promedio de Q10 de 1.90, y se presume una vida útil de 35 días a 35ºC, 28 días a 45ºC y 21 días a 55ºC. - En la salchicha el pH y el índice de peróxido son dos parámetros relevantes que pueden usarse para determinar la vida útil. El pH nos indica un valor promedio de Q10 de 2.05, y muestra una vida media de 10 días a 8ºC, 8 días a 13 y 6 días a 18ºC. Por su parte el índice de peróxido, su alteración nos representó un valor promedio de Q10 de 4.31 y una vida útil de 10 días a 8, 13 y 18ºC, respectivamente. - En el fríjol cocido el pH y el índice de peróxido también son parámetros importantes que pueden usarse para determinar la vida útil. El pH nos indica un valor promedio de Q10 de 2.10, y muestra una vida media de 12 días a 8ºC, 6 días a 13ºC y 6 días a 18ºC. Por su parte el índice de peróxido, su incremento nos representa un valor promedio de Q10 de 3.09 y una vida útil de 15 días a 8, y de 10 días 13 y 18ºC, respectivamente. - Los datos de vida útil obtenidos en cada temperatura de tratamiento de los alimentos pueden considerarse para realizar extrapolaciones para saber la vida útil a una temperatura deseada. - Los alimentos con alta concentración de carbohidratos tienen un incremento en las reacciones por cada temperatura de almacenaje representado con un valor promedio aproximado de Q10 = 1.56. - Los alimentos con alta concentración de proteínas y lípidos tienen un incremento en las reacciones por cada temperatura de almacenaje representado con un valor promedio aproximado de Q10 = 2.47. REFERENCIAS Badui Dergal Salvador. Química de los Alimentos. 1999. Longman de México Editores, S. A. de C. V. pp. 45, 69-84, 125, 170-179 Baker G. L., C. A. Sims, D. A. Gorbet, T. H. Sanders, S. F. O'Keefe. 2002. Storage Water Activity Effect on Oxidation and Sensory Properties of High-Oleic Peanuts. Journal of Food Science 67:1600 Bate, E. C., T. N. Morris, 1952. Food Science and Preservation of Foods Cambridge: University Press, pp 319. Biradar U. S., D. K. Dev, y U. M. Ingle. 1985. Shelf-Life Extension of Pedha by Packaging”, Journal of Food Science 50:51-55. Brennan, J. G., J. R. Butters, N. D. Cowell. 1998. Las operaciones de la ingeniería de los alimentos. Editorial Acribia, S. A. Zaragoza, España. pp. 473-482 Cardelli, C., T. P. Labuza, 1988. Application of Weibull Hazard Analysis to the Determination of the Shelf Life of Roasted and Grounded Coffee, Food Science and Nutrition 77:123 Cardelli, C., T. P. Labuza. 1988. “Predicting Algorithms for Oxygen Uptake and Shelf Life of Dry Foods and the Application to Coffee”, Food Science and Nutrition 78:83

Cavia M. M., M. A. Fernández- Muiño, E. Gömez-Alonso, M. J. Montes-Pérez, J. F. Huidobro, M. T. Sancho. 2002. Evolution of fructose and glucose in honey over one year: influence of induced granulation. Journal of Food Chemistry 78:157-161. Cassab Manal, Imad Toufeili. 2006. Identification of some selected rancidity indices at end of shelf life of sesame seed oil. Food Chemistry 9:7 Cegielska R. R., Pikul J. 2004. Sodium lactate addition on the quality and shelf life of refrigerated sliced poultry sausage packaged in air or nitrogen atmosphere. J Food Prot. 67(3):601-606. Deak T., E. K. Heaton, Y. C. Hung, l. R. Beuchat. 1987. Extending the Shelf Life of Fresh Sweet Corn by Shrink-Wrapping, Refrigeration, and Irradiation, Journal of Food Science 52:1625-1631. Del Nobile M. A., Buonocore G. G, Limbo S, Fava P. 2003. Shelf Life Prediction of Cereal-based Dry Foods Packed in Moisture-sensitive Films, J Food Sci 68(4):1292-1300. Escudero A., E. Asensio, J. Cacho, V. Ferreira. 2002. Sensory and Chemicals changes of young white wines stored under oxygen. An assessment of the role played by aldehydes and some other important odorants, Journal of Food Chemistry 77: 325-331. Eyring, H. 1936. Viscosity, plasticity and diffusion as example of absolute reaction rates. J. Chem. Phys. 4:283 Fennema Owen R. 1996. Química de los Alimentos. Editorial Acribia, S. A. Zaragoza, España. pp. 82-153 Fritsch C. W., C. N. Hofland, Z. M. Vickers. 1997. Shelf Life of Sunflower Kernels, Journal of Food Science 62:425-429. Hart, F. L. 1998. Análisis Moderno de los Alimentos. Editorial Interamericana. México, D. F. pp. 145-151 Hodge, J. E. 1953. Chemistry of browning reactions in models systems. Journal of agricultural and Food Chemistry 1:928. Inglett, G. E. 1975. Fabricated foods. The avi publishing Wesport Conn. pp. 99-105 Jamieson, Michael, Peter Jobber. 1974. Manejo de los Alimentos. Vól. 1 Ecología del almacenamiento, Editorial Pax-México D. F., pp. 105-120, 146-149 Karbancioglu F. Guler. 2004. Effect of storage temperature on the shelf life of compound chocolate. Food Chemistry 17:6 Labuza, T. P., Riboh D. 1982. The Search for Shelf Life. An update on continued efforts in understanding practical strategies for determining and testing the shelf life of food products” Food Technol. 36:66-74 Labuza, T. P. S. Altmarch, M. 1982. Kinetics of browning and protein loss in egg noodles under constant and square wave temperature distribution. Cereal Chem. 59:142 Lee, C. M., A. V. A. Resurrección. 2003. A comparison of Weibull Hazard method and consumer acceptance in determining the shelf life of roasted peanuts stored at different temperatures and water activities. Journal of Sensory Studies 19:433 Lee S. Y., J. M. Krochta. 2002. Accelerated shelf life testing of whey-protein-coated peanuts analyzed by static headspace gas chromatography. J Agric Food Chem. 27;50(7):2022-8. Lee Y.-U., Y.-M. Kang, J.-B. Eun, E. Y. Jang, and J.-S. Kum. 2005. Changes of physical and chemical characteristics and sensory properties in instant rice soup mix during storage. Food Sci. & Technol. 54:25 Maftoonazad Neda, Hosahalli S. Ramaswamy, 2006. Canada Kinetics of quality changes associated with coated and non-coated avocados stored at different temperatures. Fruit & Vegetable Products 16:03. Meilgaard, M. Civille, G. Carr, B. 1991, CRC. Press. Boca Ratón, FL. pp. 60, 106-110 Oellingrath I. M., Erik Slinde. (1988). “Sensory Evaluation of Rancidity and Off-Flavor in Frozen Stored Meat Loaves Fortified with Blood”, Journal of Food Science 53:967-968. O`Connor S. R. E., R. Roberts, A. L. Ford, S. M. Nottingham. (1994). Shelf Life of Minimally Processed Honeydew, Kiwifruit, Papaya, Pineapple and Cantaulope, Journal of Food Science 59:1203-1206 Pedrero, D. y Pangborn, R. M. 1997. Evaluación Sensorial de los Alimentos. Métodos analíticos. Longman de México Editores, S. A. de C. V. Alambra Mexicana. pp. 77-105

Pfeiffer Christian, D´Aujourd´hui Marcel, Walter Juris, Nuessli Jeannette and Escher Felix. 1999. Optimizing Food Packaging and Shelf Life, Journal Food Technology 53:52-57. Potter Norman, 1995. La ciencia de los alimentos. Editorial Harla, México, D. F., pp. 41-62, 141-169. Rehman Z., Habib Farzana, S. I. Zafar. 2002. Nutritional changes in maize (Zea mays) during storage at three temperatures. Journal of Food Chemistry 77:197-201. Reynolds, T.M. (1963) Adv. Food Res., 12, 1-52 Schamberger, G. P., T. P. Labuza. 2005. Inhibition of the Maillard reaction in extended shelf life milk. Food Science & Nutrition 31:7 Wolfrom, M.L. & Rooney, C.S. 1953, J. Amer. Chem. Soc. 75: 5435-3436 Yettella Ramesh Reddy, 2006. Post-harvest treatment effects on crown-cut broccoli shelf life. Fruit & Vegetable Products 78:7

XI CONGRESO NACIONAL DE CIENCIA
Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre
Monterrey, Nuevo León.
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
CUANTIFICACIÓN DE COMPUESTOS ANTINUTRICIONALES PRESENTES EN PLANTAS COMESTIBLES
González Labastida, Fa*., Márquez Téllez, Ga., Zúñiga Pérez, Ca., Alanís, G. Ea., Valadez Vega M. Ca.
aUniversidad Autónoma del Estado de Hidalgo, Instituto de Ciencias de la Salud, Área Académica de Medicina.
Carretera a la Concepción s/n Tilcuautla Hgo, México. Teléfono y fax 7717172000, ext. 5117. Mail: [email protected].
RESUMEN
El consumo de plantas exóticas es cada vez más común en algunas regiones de Hidalgo, sin embargo se desconocen los compuestos antinutricionales que dichos alimentos puedan tener. En esta investigación se realizó el análisis químico proximal y la cuantificacón de compuestos antinutricionales presentes en las semillas de Retama (Retama monosperma) y Lupinus (Lupinus sp.). para la cual se determinó el contenido compuestos nutricionales tales como proteínas, cenizas, lípidos y carbohidratos; así como el de algunos compuestos antinutricionales que interfieren en la calidad nutricional de dichos vegetales, tales como saponinas, glucósidos cianogénicos, lectinas y taninos. Los resultados mostraron que la retama tiene mayor contenido de carbohidratos totales, cenizas, saponinas, taninos y lectinas en comparación con lupinus; mientras que para el contenido de proteínas el lupinus presentó mayor cantidad; por otro lado no se observó la presencia de glucósidos cianogénicos en ninguna de las plantas estudiadas.
ABSTRACT
The consume of exotic plants is becoming increasingly in some parts of Hidalgo, however, are unknown the antinutritional compounds that such foods may it have. This research was conducted to known the chemical proximal analysis and to quantify the antinutritional compounds present in the seeds of Retama (retama monosperma) and Lupinus (Lupinus sp). Which was determined the nutritional compounds such as protein, ash, lipids and carbohydrates, as well as some antinutritional factors that interfere with the nutritional quality of these plants, such as saponins, cyanogenic glycosides, lectins and tannins. The results showed that retama seeds has a higher content of total carbohydrates, ash, saponins, tannins and lectins compared with lupinus, while the protein content lupinus was highest than retama, on the other hand was not observed the presence of cyanogenic glycosides in any of the plants studied.
Palabras clave: Antinutrientes, lupinus, retama

XI CONGRESO NACIONAL DE CIENCIA
Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre
Monterrey, Nuevo León.
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
INTRODUCCIÓN
La población muchas veces consume plantas comestibles “tadicionales” por que creen que son saludables y desconocen los compuestos antinutricionales que dichos alimentos puedan tener, debido a que existe muy poca información acercade las características químicas, físicas, bioquímicas, toxicológicas y nutricionales.
El término antinutriente se refiere a aquellos compuestos que interfieren negativamente, en mayor o menor grado, en la absorción y metabolismo de sustancias nutritivas; algunos de los compuestos antinutricionales son: Glucósidos cianogénicos, saponinas, lectinas y taninos. Se conoce que la inhalación de HCN puede ocasionar algunos efectos tales como dolor de cabeza, somnolencia, vértigo, ritmo cardíaco rápido y débil, respiración acelerada, enrojecimiento facial, náusea y vómito (Hernández, 2007-2008); mientras que la presencia de taninos en la dieta puede formar un complejo tanino-proteína lo cual ocasiona que las proteínas sean menos digerible, inhibición de enzimas digestivas y un mal funcionamiento del tracto digestivo (Tavera 2007); así mismo la ingesta de saponinas puede ocasionar daños en las mucosas digestivas que se manifiestan en vómitos, dolor de estómago, hemorragias, mareo y úlceras y una vez que pasan a la sangre pueden producir daños en los riñones e hígado y afectar al sistema nervioso pudiendo producir un paro cardiorespiratorio; por otra parte las lectinas esta relacionada con el bajo valor nutricional de las leguminosas, se ha observado que su adición en la dieta de animales, puede causar una severa disminución del crecimiento y en ocasiones la muerte (Liener, 1989).
Por lo que en esta investigación se tuvo el objetivo de determinar la presencia de compuestos tóxicos naturales en semillas de retama y lupinus.
METODOLOGÍA
Se estudiaron dos leguminosas de grano (lupinus y retama) colectadas en el municipio de Mineral del Monte se secaron en estufa a 40 °C, se separaron las semillas de la vaina y sometieron a molienda hasta pasar por malla No 60.
Se realizó el análisis químico proximal determinando el contenido de proteínas por el método oficial Kjeldahl de la AOAC 991.20 (2002), para lo cual se utilizó el equipo de digestión, destilación y neutralización, marca Gerhardt modelo Vapodest

XI CONGRESO NACIONAL DE CIENCIA
Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre
Monterrey, Nuevo León.
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
50 (GMBH & Co, Alemania), y el factor de conversión 6.25 para obtener el porcentaje de proteína.
Para determinar el porcentaje de cenizas se utilizó el método de incineración oficial de la AOAC 923.03 (2002) para el cual fue necesario emplear una mufla Thermolyne F 1500 modelo FD 1535M (Barnstead thermolyne, USA).
El contenido de lípidos se realizó por la técnica de extracción de Soxhlet, empleando el método oficial de la AOAC 920.35 (2002), finalmente se calculó el contenido de carbohidratos totales sumando los porcentajes obtenidos de lípidos, cenizas y proteína, restando el resultado al 100%.
La determinación de saponinas se baso en su capacidad hemolítica sobre los eritrocitos; para lo cual se realizó una extracción metanólica. Las saponinas extraídas fueron disueltas en solución salina (0.9%), se agregó una suspensión de eritrocitos tripsinizados, empleando el método de microtitulación (Conrath, 1972; Jaffé et al., 1974).
Las lectinas fueron extraídas de la harina de las semillas empleando solución amortiguadora de fosfatos salinos (PBS 10 mM, pH 7.4), el extracto fue separado por centrifugación y a partir de este se determinó la actividad hemaglutinante de lectinas empleando el método de diluciones seriadas empleando una suspensión de eritrocitos humanos al 2% propuesta por jaffé en 1980, en donde se identifica la ultima dilución que presenta hemaglutinación; así mismo se determinó en contenido de proteína soluble mediante el método de Bradford empleando albumina bovina como estándar. La presencia de lectinas se reportó como actividad especifica de lectinas en 50 uL de solución
Para los glucósidos cianogénicos se utilizo el método de la AOAC (1975) que se basa en la reacción de Guignard, para poder cuantificar el HCN total que potencialmente puede ser liberado se hace uso de una hidrólisis enzimática del glucósido cianogénico.
El ensayo de taninos se realizó mediante extracción con metanol, empleando vainillina para su detección de acuerdo a la técnica propuesta por Price et al. (1978) realizando una curva de calibración empleando catequina (Sigma Chemical Co, St Louis, MO, USA) como estándar, reportándose los resultados como miligramos equivalentes de catequina por 100 g de muestra.
RESULTADOS Y DISCUSIÓN
XI CONGRESO NACIONAL DE CIENCIA
Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre
Monterrey, Nuevo León.
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
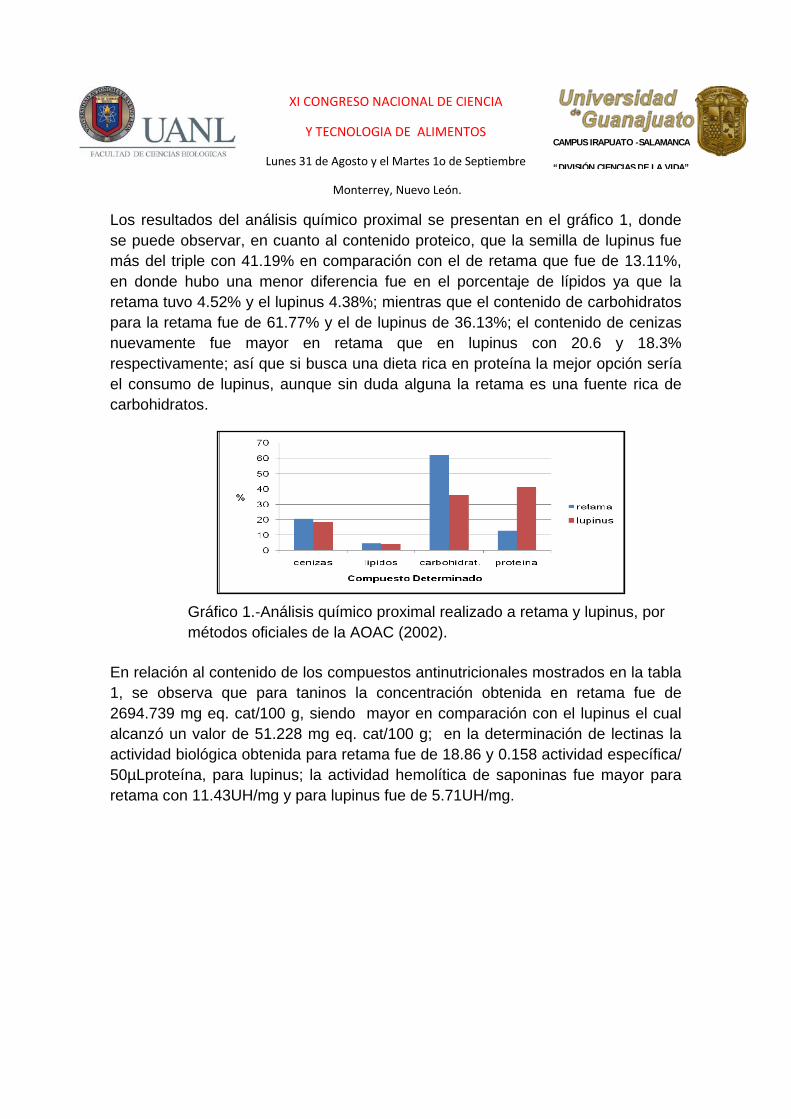
Los resultados del análisis químico proximal se presentan en el gráfico 1, donde se puede observar, en cuanto al contenido proteico, que la semilla de lupinus fue más del triple con 41.19% en comparación con el de retama que fue de 13.11%, en donde hubo una menor diferencia fue en el porcentaje de lípidos ya que la retama tuvo 4.52% y el lupinus 4.38%; mientras que el contenido de carbohidratos para la retama fue de 61.77% y el de lupinus de 36.13%; el contenido de cenizas nuevamente fue mayor en retama que en lupinus con 20.6 y 18.3% respectivamente; así que si busca una dieta rica en proteína la mejor opción sería el consumo de lupinus, aunque sin duda alguna la retama es una fuente rica de carbohidratos.
En relación al contenido de los compuestos antinutricionales mostrados en la tabla 1, se observa que para taninos la concentración obtenida en retama fue de 2694.739 mg eq. cat/100 g, siendo mayor en comparación con el lupinus el cual alcanzó un valor de 51.228 mg eq. cat/100 g; en la determinación de lectinas la actividad biológica obtenida para retama fue de 18.86 y 0.158 actividad específica/ 50µLproteína, para lupinus; la actividad hemolítica de saponinas fue mayor para retama con 11.43UH/mg y para lupinus fue de 5.71UH/mg.
Gráfico 1.-Análisis químico proximal realizado a retama y lupinus, por métodos oficiales de la AOAC (2002).

XI CONGRESO NACIONAL DE CIENCIA
Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre
Monterrey, Nuevo León.
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
El lupinus utilizado para esta investigación no presentó una elevada concentración de saponinas, en comparación con lo reportado para el lupinus angustifolius por Mercedes Muzquiz (1993) quien reportó un valor mayor de este compuesto (270.1mg /kg); sin embargo también reportó que lupinus albus no presentó este tipo de antinutriente, al igual que Fuertes Ruitón (1998) quien al realizar un estudio con Lupinus ballianus C.P. reportó como negativa la prueba para determinar saponinas, mientras que para taninos a pesar de no reportar una concentración específica afirma que el análisis fue positivo, aunque en este estudio la retama presentó una concentración de 11.43UH/mg, ésta no fue muy alta en comparación con la de lupinus angustifolius ni con una especie de frijol (winged) que presentó 320 UH/mg reportada por Nget-Hong Tan (1983).
Comparando el contenido de proteína en leguminosas reportado por mercedes muzquiz (2005) la retama del presente estudio resulto un poco por abajo del rango, mientras que el lupinus estuvo por encima de este; ya que dicho rango es de 15-40% de proteína para leguminosas, grupo de alimentos al que pertenecen ambas muestras de este estudio, por otra parte el contenido graso de las leguminosas también fue reportado en un rango de 1-7%, lo cual indica que el contenido de lípidos obtenido en este estudio para retama y lupinus se encuentran dentro del rango aceptable de las leguminosas.
CONCLUSIÓN
Las semillas de lupinus y retama contienen una cantidad aceptable de nutrientes, sin embargo la presencia de compuestos antinutricionales pueden ser causa de daño a la salud de los consumidores de estas semillas si no son procesadas física o químicamente antes previo a su consumo para su inactivación.
Muestra retama lupinus Taninos (mg eq.
Cat./100g) 2694.739 51.228
Lectinas (act. Especif.) 18.86 0.158
Saponinas (UH/mg muestra) 11.43 5.71
Glucósidos cianogénicos ND ND
Tabla 1.- resultados de compuestos antinutricionales. ND-No detectado

XI CONGRESO NACIONAL DE CIENCIA
Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre
Monterrey, Nuevo León.
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
REFERENCIAS
Badui Dergal Salvador, 2006. Química de los alimentos, 4ta edición. Editorial Pearson Education, México. Capitulo 7. Págs. 401-439.
Flores Oscar, Ibrahim Muhammand, Kass Donald, Andrade Hernán, 1998. Elefecto de los taninos de especies leñosas forrajeras sobre la utilización denitrógeno por bovinos. Revista Agroforestería en las Américas. Disponible en: http://www.virtualcentre.org/silvopastoral/documentos/x6325S00.htm#Resumen
Fuertes Ruitón César M., Roque Alcarraz Mirtha y Tristan Vidalón Maribel, 1998. Flavonoides y Alcaloides de Lupinus Ballianus C.P. Smith con Actividad Antibacteriana y Antifúngica. Instituto de Química Orgánica Aplicada a la Farmacia, UNMSM. Facultad de Farmacia y Bioquímica. Disponible en: http://sisbib.unmsm.edu.pe/BVRevistas/ciencia/v01_n2/flavonoides.htm
González Gómez J.C., Ayala Burgos A. y Gutiérrez Vázquez E., 2006. Determinación de fenoles totales y taninos condensados en especies arbóreas con potencial forrajero de la Región de Tierra Caliente Michoacán, México. Disponible en: http://www.cipav.org.co/lrrd/lrrd18/11/guti18152.htm
Hernández Jesús, 2007-2008, Toxinas naturales de origen vegetal, Facultad de Medicina Universidad Autónoma de Madrid, disponible en: http://www.uam.es/departamentos/medicina/farmacologia/especifica/ToxAlim/ToxAlim_L9.pdf
Muzquiz Mercedes, 2005. Impacto Positivo del Consumo de Legumbres en la Salud Humana. Grain Legumes Integrated Project Dissemination Event Madrid. Disponible en: http://www.grainlegumes.com/fckeditor/aepfiles/File/GLIP/Madrid%202005/Slides04-Muzquiz.pdf
Muñoz Tavera G. S.; Gutiérrez Avella D. M., 2007. Rastreo de Taninos en 14 Malezas Usadas como Forraje en el Estado de Querétaro. Facultad de Química Universidad Autónoma de Querétaro. Disponible en: http://www.uaq.mx/investigacion/difusion/veranos/memorias-2007/43_6UAQMunozTavera.pdf

XI CONGRESO NACIONAL DE CIENCIA
Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre
Monterrey, Nuevo León.
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
Muzquiz Mercedes, 1993. The Saponin Content and Composition of Sweet and Bitter Lupin Seed. Departamento de Producción y Tecnología de Alimentos, Madrid España.
Nget-Hong Tan,1983. Winged Bean (Psophocarpus tetragonolobus) Tannin Level, Phytate Content and Hemagglutinating Activity. Department of Biochemistry University of Malaya.
Sharon N. y Lis H. 1989 Lectins. Chapman and Hall.

CITOTÓXICIDAD DE PLOMO y CADMIO EN CÉLULAS EPITELIALES DE INTESTINO DELGADO DE RATA MACHO-Wistar
Reyes Toral, Na*, Zúñiga Pérez Ca., Sánchez Gutiérrez Ma.,
Valadez Vega M. Ca.
Universidad Autónoma del Estado de Hidalgo, Instituto de Ciencias de la Salud, Área Académica de Medicina. Carretera a la Concepción s/n, C.P. 42160, Tilcuautla Hidalgo, México. Teléfono y fax 7717172000, ext. 5117. Mail*:
[email protected] Resumen: Los metales que se encuentran en alimentos, deben su presencia a diferentes causas, que van desde su obtención o cultivo, hasta su industrialización y distribución. En esta investigación se realizó un estudio para determinar el efecto citotóxico de cadmio y plomo sobre células epiteliales de intestino delgado de ratas. Se emplearon fragmentos de intestino, los cuales fueron incubados con diversas concentraciones de los metales; se realizó un frotis y se midió el daño que causaban a las células intestinales. En otro ensayo, se aislaron las células epiteliales empleando tripsina; las células aisladas se incubaron con los metales bajo estudio y se determinó el número de células que sufrieron daño. Los resultados mostraron que el plomo y cadmio causan efecto citotóxico sobre las células epiteliales, el cual es dependiente de la concentración del metal; siendo mayor el efecto causado por el plomo; así mismo, se observó que el daño es mucho mayor cuando las células son liberadas del intestino debido a que en este proceso las células están más expuestas a los metales. ABSTRACT: Heavy metals may enter the human body through food, water, air, or absorption through the skin when they come in contact with humans in agriculture and in manufacturing, pharmaceutical and industrial. This research was realized to evaluate the citotoxicity effect of cadmium and lead on epithelial cells from small intestine of rats. Intestine fragments were incubated in metals solutions. The intestinal cells spread on microscope slides and measure the effect of the metal damage on intestine cells. In other assay the epithelial cells were isolated employing trypsin; the isolated cells were incubated with metal solutions, after incubation was determinate the number of cells damage. The results shown that cadmium and lead cause cytotoxic effect on the epithelial cells, which is metal concentration depending. The lead caused mayor effect than cadmium; the effect was higher when the cells were liberated from the intestine because the cells are more exposed to the metal. Palabras clave: Cadmio, Plomo, citotoxicidad, células epiteliales.

INTRODUCCIÓN Los metales son materiales que se encuentran presentes de manera natural en el medio ambiente y desde la antigüedad han desempeñado un papel importante en el desarrollo de las civilizaciones. Sin embargo, cuando los metales son utilizados de manera irracional terminan por causar daño a la salud. Una fuente importante de contaminación por metales pesados son los alimentos; en la mayoría de los casos, los alimentos actúan como vehículos de los metales los cuales a menudo son contaminantes presentes en el medio ambiente o deben su presencia a diferentes causas, que van desde su obtención o cultivo de los alimentos hasta su industrialización y distribución (Wienk et al., 1999) Los metales pesados llegan a ser tóxicos cuando no son metabolizados por el organismo y se acumulan en los tejidos dependiendo, sobre todo, de la concentración, tal es el caso del envenenamiento por cadmio que produce osteoporosis, enfisema pulmonar, cáncer de pulmón, cáncer de próstata, hipertensión, diversas cardiopatías y retraso en la habilidad verbal de los niños (Nordberg, 1988); sin embargo, existen metales como el plomo que son utilizados en la industrialización de los alimentos a pesar de tener conocimiento sobre el daño toxico que causa, por ejemplo, este metal es utilizado en la fabricación de latas y de soldadura. El plomo puede ser lixiviado a partir de la soldadura a los alimentos almacenados en la lata, también se utiliza en la elaboración de cerámicas y esmaltes que están hechos o revestidos con pinturas a base de plomo (Valle and Lucas, 2000). El objetivo del presente trabajo fue evaluar el efecto citotóxico del plomo y del cadmio en células epiteliales de intestino delgado de ratas Wistar macho. METODOLOGÍA Se emplearon ratas Wistar macho con previo ayuno de 12 h y agua ad libitum. Se extrajo el intestino delgado, se fragmentó (Sotelo et al, 1980) y parte de este fue empleado para liberar las células epiteliales intestino empleando tripsina bovina al 0.25% según el método de Culling et al. (1973). Los fragmentos de intestino se incubaron con soluciones de cadmio o plomo a diferentes concentraciones en lapsos de 15 min, posteriormente se realizó un frotis y mediante la técnica de exclusión se determinó la viabilidad al microscopio óptico empleando azul de tripán (0.05%). Para el estudio de citotoxicidad en células libres se tomó una alícuota de las células liberadas y se incubaron con las soluciones de plomo o cadmio durante 15 min, posteriormente se tomó una alícuota, se colocó sobre una cámara de Neuvauer, se tiñó con azul de tripán y se realizó el conteo de células vivas al microscopio óptico. RESULTADOS Y DISCUSIONES

( )( )
En la Figura 1 se muestra el efecto citotóxico del cadmio en las células epiteliales del intestino delgado; observándose que en las células libres (línea en azul) y las células de los frotis (línea en rojo) la viabilidad disminuye al aumentar la concentración del metal, sin embargo, en el caso de las células libres, el efecto que ejerce este metal es mayor que cuando se realiza el estudio en frotis. Al calcular la dosis inhibitoria 50 (DI50) para el estudio de las células en frotis se obtuvo una concentración de 1079.91µg/mL y para células libres la concentración fue de 171.06 µg/mL; el efecto observado se debe a que en el estudio de frotis al ser expuestas al cadmio, las células se encuentran más protegidas por las condiciones de su entorno natural que es el intestino, mientras que las células libres se encuentran desprotegidas y más expuestas al metal.
Figura 1. Estudio de citotoxicidad del cadmio en células epiteliales.
La línea azul representa el estudio realizado con células libres según el método de Culling et al. (1973), la línea roja indica el estudio con en fragmentos de intestino, en donde se realiza un frotis para determinar la viabilidad celular de acuerdo a la técnica de (Sotelo et al., 1980). La línea en negro indica la concentración a la cual la viabilidad disminuye en un 50% (DI50). Estudios realizados por Marcano et al., 2005 indicó que el efecto citotóxico causado por el cadmio en células normales de hígado de ratón albino, fue debido a intoxicación crónica probablemente como consecuencia del estrés oxidativo y manifestado con alteraciones en la actividad biosintética y en el metabolismo de lípidos, haciéndose más severo al aumento de concentración, pudiendo conllevar a una necrosis celular; de igual manera Beman (2006) reportó que la exposición a CdCl2 causó la inhibición de la actividad en la glutatión peroxidasa. Estudios In vitro han demostrado que el cadmio causa efecto citotóxico en líneas celulares de hepatoma, dañando principalmente a la membrana celular (Fotakis and Timbrell, 2006). Los resultados obtenidos en el presente trabajo mostraron que para células epiteliales de intestino se necesita de mayores concentraciones de cadmio para causar daño citotóxico, en comparación a células hepáticas reportadas por otros investigadores (Beman, 2006; Fotakis and Timbrell, 2006; Marcano et al., 2005.
( )( )

. En la Figura 2 se muestran los resultados del efecto citotóxico causado por el plomo a las células epiteliales del intestino delgado, observándose diferencias entre los estudios de células epiteliales libres y las células en frotis; en el caso de las células libres, la viabilidad disminuye más rápidamente que en las células de los frotis; debido a que el plomo causa un mayor daño a las células libres que se encuentran fuera de su entorno natural mientras que, las células de los frotis se encuentran protegidas por el intestino al ser expuestas al metal. Siendo la DI50 para células libres de 8.04 µg/ml y para frotis de 91.03 µg/ml.
Figura 2. Estudio de citotoxicidad del plomo sobre células epiteliales.
La línea azul ( )( ) representa el estudio realizado con células libres según el método de Culling et al. (1973), la línea roja indica el estudio con en fragmentos de intestino, en donde se realiza un frotis para determinar la viabilidad celular de acuerdo a la técnica de (Sotelo et al., 1980). La línea en negro indica la concentración a la cual la viabilidad disminuye en un 50% (DI50). Estudios realizados In vivo con humanos e In vitro con ratas por Jung-Hun et al. (2009) demostraron que el plomo puede causar efecto citotóxico sobre eritrocitos y provocar actividad procoagulante, además de causar trombos en las ratas; también la exposición a plomo causó estrés celular sobre fibroblastos de acuerdo a Beman (2006). Otro estudio realizado con ratones macho-Swiss mostró que el acetato de plomo provocó estrés oxidativo en células germinales Acharya (2003). De acuerdo al estudio realizado en la presente investigación, en comparación con el estudio realizado con fibroblastos se observa que en células epiteliales necesitan una menor concentraciones de plomo para observar un efecto citotóxico que en fibroplastos (Beman, 2006); sin embargo cuando se emplean eritrocitos y células germinales las concentraciones utilizadas de plomo para causar daño fueron más bajas que para células epiteliales (Acharya, 2003; Jung-Hun et al., 2009). Estas diferencias se deben al tipo de células empleadas para cada estudio, siendo los hepatocitos, células germinales y células epiteliales más vulnerables a la exposición del plomo.
( )( )

CONCLUSIONES El plomo presentó mayor daño citotóxico a las células epiteliales que el cadmio, siendo más severo el daño en las células cuando estas se encuentran fuera del tejido, ya que dentro de esté mostraron mayor resistencia. La ingesta constante de alimentos con elevados contenidos de plomo y cadmio pueden ser perjudiciales para nuestra salud ya que se ha demostrado que causan daño en las células del intestino delgado en donde se lleva acabo la absorción de nutrientes y otras sustancias. REFERENCIAS
1. Ali Beman Zaree Mahmodabady, Mehdi Saberi, Hossein Eimani, Jila Pyrzad,
Reza Rezaee Sharifabady, Autumn 2006 .Cytotoxic and Oxidative Stress Caused by Cadmium and Lead on Human Skin Fibroblast Cells. Biochemistry Department, Molecular Biology. Yakhteh Medical Journal, 8, (3):176.
2. Culling, C. F. A., Reid, P. E., Trueman, L. S. y Dunn, W. Y. 1973. A simple
method for isolation of viable epithelial cells of the gastrointestinal tract. Proc Soc. Exp. Biol. Med.
3. Cruz Vallejo, V. L. y Morales Ramírez, Cinética de inducción de daño en el
ADN por la exposición de leucocitos de ratón in vivo a MNU. 2000, Revista electrónica salud pública y nutrición. Depto. Biol. ININ. AP18-1027 México, D.F. 2:15-21.
4. George Fotakis, John A. Timbrell, 2006. In vitro cytotoxicity assays:
Comparison of LDH, neutral red,MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. London SE1 8WA 171–177.
5. Greger, J.L. 1985. Aluminum content of the american diet. Food Technol.
39(5):73. 6. Lindner, L. 1995 Toxicología de alimentos. Editorial. Arribia. Zaragoza,
España. (2): 163-166. 7. Louis W. Chang, 1993. Toxicology of metals. Lewis Publishers, New York.1:15-
20.

8. Marcano Letty, Faria, Clarisa de R, Carruyo,Ingrid, Montil Xiomara (Junio 2006) Cadmum citotoxicity in mice hepatocytes and implicatios on tropcal environments. Revista de biología tropical. 54(2) 257-263.
9. Sotelo, A. Arteaga, M.E., Frías, M. I. y González-Garza, M. T. 1980. Cytotoxic
effect of two legumes on epithelial cells of the small intestine Qual Plant Foods Human Nutritional.30(2):79-85.
10. U.R. Acharya, S. Acharya and M. Mishra, 2003. Lead Acetate Induced
Cytotoxicity in Male Germinal Cells of Swiss Mice. Department of Zoology, Berhampur University, Berhampur, 760007, Orissa, India. 41, 291–294.

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
CITOTÓXICIDAD DE EXTRACTOS PROTEICOS DE FRIJOL TEPARY Y LECTINA PURA DE FRIJOL PHA-E EN CÉLULAS EPITELIALES DE
INTESTINO DELGADO DE RATA MACHO-Wistar.
Reyes Toral Na, Sánchez Lozada L Y.a*, Islas Islas V.a, Sánchez Gutiérrez M.a, Valadez Vega MC.a
aUniversidad Autónoma del Estado de Hidalgo, Instituto de Ciencias de la Salud, Área Académica de Medicina. Carretera a la Concepción s/n, CP 42160, Tilcuautla, Hidalgo,
México. Teléfono y fax 7717172000, ext. 5117. *Mail: [email protected].
RESUMEN: Las leguminosas como el frijol son una fuente rica y económica de proteínas carbohidratos, minerales y fibra dietética; ocupa un lugar importante en la alimentación de muchos países, sin embargo se conoce que contiene algunos compuestos que pueden disminuir su calidad nutricional, entre los que se encuentran las lectinas, los cuales han sido ampliamente estudiadas debido a su efecto toxico. Por esta razón se propuso evaluar el efecto citotóxico de extractos proteicos y lectinas puras de frijol en células epiteliales de intestino delgado de rata macho Wistar. Se emplearon fragmentos de intestino, los cuales fueron incubados con diversas concentraciones de extractos o lectinas puras; se realizó un frotis y se midió el daño que causaban a las células intestinales. En otro ensayo, se aislaron las células epiteliales empleando tripsina; las células aisladas se incubaron con extractos o lectinas puras y se determinó el número de células que sufrieron daño, resultados indican que tanto para extractos como lectina pura para los estudios de frotis y de células libres hay una disminución en la viabilidad al aumentar la concentración, siendo mayor el efecto citotóxico cuando las células se encuentran libres, siendo mayor el efecto causado por las lectinas puras en ambos estudios. ABSTRACT: Legumes as the bean is a rich and economic source of proteins, carbohydrates, minerals and dietetic fiber; its occupy an important place in the feeding of many countries, nevertheless is known that it contains some compounds that can diminish their nutritional quality, between them are the lectins, which has been widely studied due to their toxic effect. Therefore the task was evaluate the cytotoxic effect of protein extracts and pure lectins of common bean on epithelial cells of small intestine of Wistar male rat. Intestine fragments were incubated extracts or pure lectins solutions. The intestinal cells spread on microscope slides and measure the effect of the metal damage on intestine cells. In other assay the epithelial cells were isolated employing trypsin; the isolated cells were incubated with extracts or pure lectins solutions, after incubation was determinate the number of cells damage. The results indicate that as the extracts as pure lectins, in the two studies, slides and free cells, the viability diminish when concentration increasing, being greater the cytotoxic effect when the cells are free; being greater the effect caused by the pure lectins in both studies. Palabra Clave: Lectina, Extracto proteico, Células Epiteliales.

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
INTRODUCCIÓN Entendemos por leguminosas a todos los granos secos que vienen de vainas, como lo son: el frijol, haba, lenteja, soya y garbanzos, sus granos contienen entre 50% y el 70% de hidratos de carbono solubles, bajo contenido en grasas (1-2%), alto porcentaje de proteínas, en general superior al 20%, fibras en un 8% y sustancias minerales, próximas al 3%, destacando su alto contenido en calcio y hierro (FAO/OMS/ONU. 1985), también contienen ciertos componentes que dificultan la digestibilidad y causan trastornos fisiológicos que limitan su consumo, conocidos como compuestos antinutricionales siendo muchos de ellos eliminados por la cocción (Antunes and Sgarbieri, 1980). El frijol es una fuente de alimentación de gran importancia en la dieta de la población de bajos recursos económicos, contiene 22% de proteínas de alta digestibilidad, es de alto valor energético, contiene alrededor de 70% de carbohidratos totales y aporta cantidades importantes de minerales (Ca, Mg, Fe), Vitaminas A, B1, B2, C. (Azevedo et al, 2003). Así mismo se conoce que contiene compuestos antifisiológicos como las lectinas, taninos, saponinas, inhibidores de tripsina los cuales causan daño a células, tejidos y órganos de animales y humanos cuando su alimentación contiene dichos compuestos en su forma activa. Por dicha razón se propuso evaluar el efecto citotóxico de extractos proteicos de frijol tépari y flor de mayo así como lectina puras de frijol común sobre células epiteliales de intestino delgado de rata wistar. MÉTODOLOGIA
Obtención de extractos proteicos Se emplearon semillas de frijol de las cuales se obtuvo una harina la cual se sometió a una extracción de las proteínas con solución reguladora fosfato salino(PBS) 0.01M, pH 7.4, en relación 1:10(W/V) se agitó durante 16h a 4ºC, se centrifugaron obteniendo el sobrenadante el cual fue dializado contra agua desionizada con cuatro cambios por 24h a 4°C y posteriormente fueron liofilizados para ser utilizados en los ensayos. Obtención de fragmentos intestinales y células epiteliales Se utilizaron ratas macho Wistar con previo ayuno de 12 h y agua ad libitum. Los animales fueron sacrificados en cámara de gases con cloroformo, se colocaron en forma ventral y se les realizo una laparotomía, se extrajo el intestino delgado, se corto en segmentos de 2 cm los cuales fueron invertidos en aplicadores de madera estéril se lavaron con PBS con estreptomisina al 1.5% a continuación con solución salina isotónica posteriormente fueron colocados en Medio de cultivo completo (Medio Mc Coy 5A Modificado, suero fetal bovino al 7%, estreptomicina 1.5% Gibco) dichos fragmentó fueron utilizados para el estudio de viabilidad por el método de frotis y para la liberación de células.

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
Para la liberación de células libres fragmentos de intestino fueron incubados por 1h a 37°C en tripsina bovina al 0.25% (Fluka); las células liberadas fueron lavadas mediante centrifugación con medio Mc Coy 5ª modificado y finalmente fueron resuspendidas en medio de cultivo completo. Estudios de Viabilidad Celular por el método de frotis Se tomaron trozos de intestino y se incubaron por lapsos de 15min a 37°C con extractos proteicos o lectina pura a diferentes concentraciones, posteriormente se realizaron frotis con el trozo de intestino en un porta objetos y mediante la técnica de exclusión empleando azul tripán al 0.05% se determino la viabilidad al microscopio óptico, reportando los resultados en porcentaje de viabilidad. Estudio de Viabilidad de Células Libres De la suspensión celular previamente obtenida se colocaron en tubos alícuotas de 1.4x108 células/mL, a las cuales se le agregaron extracto proteico o lectina pura a diferentes concentraciones se incubaron por 15 min a 37°C; transcurrido la incubación se tiñeron con azul tripán y se contaron al microscopio óptico en cámara de Neuvauer, Reportando resultados en porcentaje de viabilidad. RESULTADOS Y DISCUSIONES
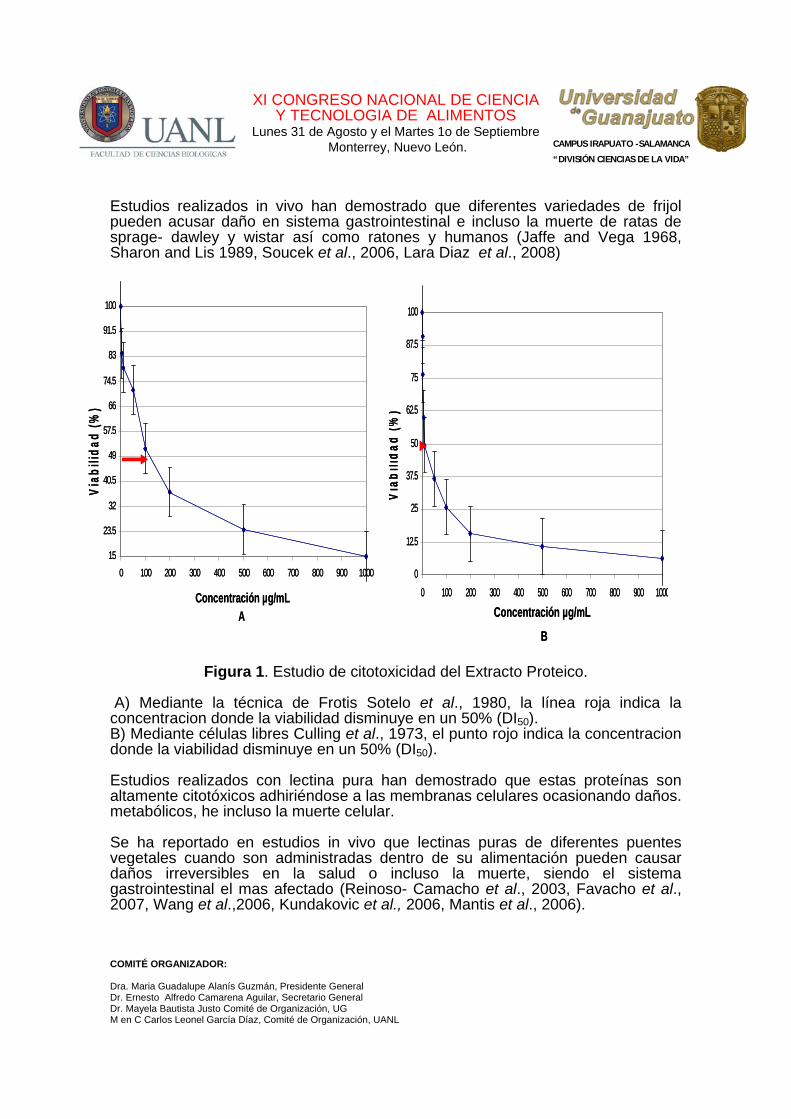
En la Figura 1 se observa el efecto citotóxico del extracto proteico en las células epiteliales del intestino delgado; observándose que tanto en el estudio de células libres (figura 1ª) como para el estudio en frotis (figura 2ª) la viabilidad disminuye al aumentar la concentración del extracto, sin embargo, en el caso de las células libres, el efecto que ejerce es mayor que cuando se realiza el estudio en frotis. Al calcular la DI50 para el estudio de las células en frotis se obtuvo un valor de 100 µg/ml, mientras que para células libres la concentración inhibitoria 50 fue de 10 µg/ml; el efecto observado se debe a que en el estudio de frotis al ser expuestas al extracto, las células se encuentran más protegidas por las condiciones de su entorno natural que es el intestino, mientras que las células libres se encuentran desprotegidas y más expuestas al extracto proteico. El efecto observado se puede deber a que en los extractos proteicos contengan lectinas, las cuales tienen la capacidad de unirse a carbohidratos específicos en la membrana celular alterando funciones celulares; tales como disminuyendo la absorción de nutrientes, metabolismos e inclusive ruptura de membrana celular. Estudios realizados en células epiteliales de rata wistar han demostrado que extractos proteicos de diferentes variedades de frijol causan daño citotóxico en estas células (Valadez et al., 2005). Estudio realizado en células epiteliales por Sotelo et al., en 1980 demostraron que semillas de leguminosas pueden causar daño en células.
XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
Estudios realizados in vivo han demostrado que diferentes variedades de frijol pueden acusar daño en sistema gastrointestinal e incluso la muerte de ratas de sprage- dawley y wistar así como ratones y humanos (Jaffe and Vega 1968, Sharon and Lis 1989, Soucek et al., 2006, Lara Diaz et al., 2008)
Figura 1. Estudio de citotoxicidad del Extracto Proteico.
A) Mediante la técnica de Frotis Sotelo et al., 1980, la línea roja indica la concentracion donde la viabilidad disminuye en un 50% (DI50). B) Mediante células libres Culling et al., 1973, el punto rojo indica la concentracion donde la viabilidad disminuye en un 50% (DI50). Estudios realizados con lectina pura han demostrado que estas proteínas son altamente citotóxicos adhiriéndose a las membranas celulares ocasionando daños. metabólicos, he incluso la muerte celular. Se ha reportado en estudios in vivo que lectinas puras de diferentes puentes vegetales cuando son administradas dentro de su alimentación pueden causar daños irreversibles en la salud o incluso la muerte, siendo el sistema gastrointestinal el mas afectado (Reinoso- Camacho et al., 2003, Favacho et al., 2007, Wang et al.,2006, Kundakovic et al., 2006, Mantis et al., 2006).
15
23.5
32
40.5
49
57.5
66
74.5
83
91.5
100
0 100 200 300 400 500 600 700 800 900 1000
Viab
ilida
d (%
)
Concentración µg/mLA
15
23.5
32
40.5
49
57.5
66
74.5
83
91.5
100
0 100 200 300 400 500 600 700 800 900 1000
Viab
ilida
d (%
)
Concentración µg/mLA
0
12.5
25
37.5
50
62.5
75
87.5
100
0 100 200 300 400 500 600 700 800 900 1000
Viab
ilida
d (%
)
Concentración µg/mL
B
0
12.5
25
37.5
50
62.5
75
87.5
100
0 100 200 300 400 500 600 700 800 900 1000
Viab
ilida
d (%
)
Concentración µg/mL
B

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
En la figura 2 se muestran los resultados del efecto citotóxico causado por la lectina PHA-E a las células epiteliales del intestino delgado, observándose diferencias entre los estudios de células epiteliales libres y las células en frotis; en el primer caso (2ª), la viabilidad disminuye más rápidamente que en las células de los libres; debido a que la lectina causa un mayor daño a las células libres ya que no se encuentran protegidas por el intestino al ser expuestas a la lectina. Siendo la DI50 para células libres de 4 µg/ml y para frotis de 10µg/ml.
Gráfica 2. Estudio de citotoxicidad del Lectina PHA-E sobre células epiteliales.
A) Mediante la técnica de Frotis Sotelo et al., 1980, la línea roja indica la concentración donde la viabilidad disminuye en un 50% (DI50). B) Mediante células libres Culling et al., 1973, el punto rojo indica la concentración donde la viabilidad disminuye en un 50% (DI50). CONCLUSIONES Se conoce que las semillas de frijol presentan compuestos antinutricionales por lo que un extracto proteico también presentan proteínas toxicas tales como las lectinas las cuales en gran medida son causantes de daños citotóxicos. Al comparar el efecto del extracto y la lectina, se observó que la lectina causa mayor daño a las células epiteliales libres ya que son las susceptibles al efecto citotóxico debido a que se encuentran fuera de su ambiente natural. REFERENCIAS Antunes, P.L. y Sgarbieri, V.C. 1980. Effect of heat treatment on the toxicity and nutritive value of dry bean (Phaseolus vulgaris) proteins. J. Agric. Food Chem. 28: 935-39.
0
10
20
30
40
50
60
70
80
90
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280 300
Concentración µg/mLA
Viab
ilida
d (%
)
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 10 100 200 300
Concentración µg/mLB
Via
bilid
ad (%
)
0
10
20
30
40
50
60
70
80
90
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280 300
Concentración µg/mLA
Viab
ilida
d (%
)
0
10
20
30
40
50
60
70
80
90
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280 300
Concentración µg/mLA
Viab
ilida
d (%
)
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 10 100 200 300
Concentración µg/mLB
Via
bilid
ad (%
)
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 10 100 200 300
Concentración µg/mLB
Via
bilid
ad (%
)
0
10
20
30
40
50
60
70
80
90
100
0 2 4 6 10 100 200 300
Concentración µg/mLB
Via
bilid
ad (%
)

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
Azevedo, L., Gomes, J. C., Stringheta, P. C., Gontijo, A. M, Padovani C. R., Ribeiro L. R., Salvadori, D. M. 2003. Black bean (Phaseolus vulgaris L.) as a protective agent against DNA damage in mice. Food Chem Toxico., 41:1671-6.
Culling, C F A, Reid P E., Trueman L. C. and Dunn W Y. 1973 a simple method for isolation of viable epithelial cells of the gastrointestinal tract. Proc Soc Exp. Biol. Med. De Mejia EG, Guzmán-Maldonado SH, Acosta-Gallegos, JA, Reynoso- Camacho R,, Ramirez- Rodriguez E, Ponz – Hernandez, JL, Gonzalez – Chavira M, Catellanos JZ and Kelly JD 2003 Effect of cultivar and growing location on the trypsin inhibitors, tannins, and lectins of common beans grown in the semiarid highlands of Mexico J Agric Food Chem 51: 5962-5966. Elvira Gonzalez de Mejia, Maria del Carmen Valadez Vega, Rosalia Reynoso Camacho y Guadalupe Loarca piña 2005 Tannis, Trypsin Inhibitors and Lectin cytotoxicity in Tepary and Common beans. FAO/OMS/ONU. 1985. Necesidades de energía y proteínas. OMS., Ginebra. Serie de Informes técnicos Nº 724. Favacho AR, Cintra EA, Coelho LC, Linhares MI 2007 In vitro activity evaluation of Parkia pendula seed lectin against human cytomegalovirus and herpes vírus 6. Jaffé, W. G. y Vega C. 1968. Hea labile growth inhibition factors in bean (Phseolos vulgaris) . American Journal Nutritional. Kundakovic T, Stanojkovic T, Juranic Z, Kovacevic N 2006 Cytotoxicity in vitro of naphthazarin derivatives from Onosma arenaria. Lara-Diaz VJ, Gaytán-Ramos AA, Dávalos- Balderas AJ, Santos-Guzman J, Mata- Cárdenas BD, Vargas-Villarreal J, Barbosa-Quintana A, Sanson M, López- Reyes AG, Moreno- Cuevas. Mantis NJ, McGuinness CR, Sonuvi O, Edwards G, Farrant SA 2006 Immunoglobulin A antibodies against ricin A and B Subunits protect epithelial cells from ricin intoxication. Sharon, N. y Lis, H. 189. Lectins. Ed. Chapman and Hall LTd. Sotelo A, Arteaga Me, Frias Hi Gonzalez Garza Mt 1980. Cytotixic effect of two legunes on epithelial cells of the small intestine qual plant Foods HumNutr 30; 79-85. Soucek J, Skvor J, Poucková P, Matousek J, Slavik T, Matousek J 2006 Mung bean sprout ( Phaseolus aureus) nuclease and its biological and antitumor effects.
Wang BZ, Zou WG, Liu WY 2006 The lower cytotoxicity of cinnamomin ( a type II RIP ) is due to its B- Chain.

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”

PEPTIDOS CON ACTIVIDAD ANTIHIPERTENSIVA OBTENIDOS A PARTIR DE AISLADOS PROTEICOS DE GARBANZO (CICER ARIETINUM L)
Carrasco-Castilla, J.a*, Jiménez-Martínez, C.a y Dávila-Ortíz, G.a
a Escuela Nacional de Ciencias Biológicas, Departamento de Graduados en Alimentos
Instituto Politécnico Nacional, Prol. Carpio, Esq. Plan de Ayala, S/N. Col. Casco de Santo Tomás. Del. Miguel Hidalgo. C.P. 11340. México, D. F. Tel.: (+52) 55 57296000 Ext.
62462 * [email protected]
RESUMEN: La hipertensión arterial es uno de los padecimientos que ocupa cada vez mayor atención debido al gran número de personas afectadas y sus graves consecuencias en riñón, corazón, cerebro y retina, principalmente. Numerosos estudios in vitro e in vivo, sugieren que el consumo de péptidos con actividad antihipertensiva representa una alternativa como tratamiento en personas que padecen dicha enfermedad. En el presente trabajo se investiga el uso de la proteína de garbanzo (Cicer arietinum L.) como fuente de estos biopéptidos. Para tal fin se obtuvo un aislado proteico de garbanzo con una concentración de 73.75g/100g, y se obtuvo un rendimiento del 43%. Posteriormente, el aislado proteico se hidrolizó de forma individual con enzimas digestivas (pepsina, tripsina y quimotripsina) y se evaluó el efecto antihipertensivo de los hidrolizados, con la técnica in vitro de Hayakari et al, 1978. El grado de hidrólisis obtenido, después del tratamiento enzimático de los aislados, fue de mayor a menor, la pepsina, tripsina y quimotripsina. El hidrolizado de proteína de garbanzo tratado con tripsina durante 3h mostró mayor efecto inhibitorio de la ECA-I (IC50 0.613 mg/mL). Los resultados obtenidos permiten sugerir que las proteínas de garbanzo presentan péptidos con efecto antihipertensivo. ABSTRACT: Arterial hypertension is one of the illnesses that demands major attention due to the great number of affected people and its serious complications in kidney, heart, brain and retina, mainly. Numerous in vitro and in vivo studies suggest that the consumption of peptides with antihypertensive activity represents an alternative treatment in patients who suffer this disease. In this study, we investigated the use of the protein of chickpea (Cicer arietinum L.) as source of bioactive peptides. There was obtained a chickpea protein isolates with a concentration of 73.75g/100g and a yield of 43 %. After that, the isolates were hydrolysed by an individual way with digestive enzymes (pepsin, trypsin and chymotrypsin) and the antihypertensive effect of the hydrolysates was determined according to the colorimetric method of Hayakari et al, 1978. The obtained hydrolysis degree after the enzymatic treatment of the concentrates was since higher with pepsin, followed with trypsin and chymotrypsin. The protein hydrolysates of chickpea treated with trypsin during 3h showed major inhibitory effect (IC50 0.613 mg/mL) of the angiotensin-I converting enzyme activity (ACE). These results show that chickpea protein hydrolysates obtained by treatment with pepsin, trypsin and chymotrypsin are a source of peptides with ACE inhibitory activity. Palabras clave: Actividad antihipertensiva, Hidrolizados proteicos de garbanzo, Péptidos bioactivos.

INTRODUCCIÓN La hipertensión arterial sistémica (HAS) es una enfermedad crónica, controlable que se caracteriza por un aumento en la presión arterial sistólica (PS) mayor de 140 mmHg y/o de la presión arterial diastólica (PD) igual o mayor a 90 mmHg. La HAS es un importante problema de salud pública por su estrecha asociación con el desarrollo de las enfermedades cardiovasculares y cerebrovasculares. La prevalencia de HAS en México es de 30.7% en adultos mayores de 20 años, sin embargo 2 de cada 3 hipertensos desconoce su enfermedad, de éstos poco más de la mitad reciben tratamiento y sólo el 14.6% logran su control tensional (Velazquez-Monroy et al, 2000). El tratamiento consiste básicamente en medidas de tipo dietéticas, pero cuando no es suficiente, entonces se opta por iniciar un tratamiento farmacológico. Sin embargo, su ingesta puede presentar efectos colaterales en el paciente, por lo cual, el uso de péptidos bioactivos representa una alternativa como tratamiento. Los péptidos bioactivos se definen como secuencias de aminoácidos inactivas de pequeño tamaño (entre dos y quince residuos) en el interior de la proteína precursora, pero que ejercen propiedades biológicas al liberarse por procesos de hidrólisis enzimática (Vioque et al, 2000). Todos los alimentos pueden aportar péptidos bioactivos. Además de la leche humana y de vaca, que son los más estudiados, se han aislado péptidos a partir de hidrolizados enzimáticos de proteínas muy diversas como las de de sardina, huevo, maíz, soja, etc (Yamamoto, 1997). Los péptidos con actividad antihipertensiva, inhiben la enzima convertidora de angiotensina I (ECA-I). Esta enzima degrada la angiotensina I, y libera el octapéptido angiotensina II, el cual es un potente vasoconstrictor y provoca un aumento en la presión arterial actuando sobre receptores específicos. Se ha encontrado que los péptidos bioactivos disminuyen la presión en ratas hipertensas (Matsufuji et al, 1995) y en personas hipertensas (Matsui et al, 1995). Además de prevenir la hipertensión, en otros estudios se ha encontrado que los péptidos inhibidores de la ECA pueden ser utilizados en el tratamiento de falla cardiaca crónica y del infarto del miocardio (Pitt, 1997). METODOLOGÍA Materiales Las semillas secas de garbanzo (Cicer arietinum L. var. Kabuli) fueron adquiridas en la Central de Abastos de la Ciudad de México. Las enzimas tripsina, pepsina, quimotripsina, la enzima convertidora de la angiotensina-I (ECA-I) y el sustrato sintético N-Hipuril-l-Histidil-l-Leucina (HHL) son puras (Sigma Co.) El marcador de peso molecular para la electroforesis fue de BioRad. Los demás reactivos fueron de grado analítico. Análisis Químico Proximal

La determinación de humedad (Método 925.09), ceniza (Método 923.03), proteína cruda (Método 954.01), fibra total dietaria (Método 985.29) y lípidos (Método 920.39) para la harina y el aislado proteico se realizó con los métodos oficiales de la AOAC, 1995, por triplicado. Obtención del Aislado. Se preparó una suspensión 1:10 p/v de la harina de garbanzo y se ajustó el pH a 9 con NaOH 0.1N y se agitó durante 30 min a temperatura ambiente. Después de centrifugar a 10000 rpm, 30 min a 4 oC, se ajustó el pH del sobrenadante al punto isoeléctrico (pI 4.3) y precipitado formado se recuperó por centrifugación en las condiciones mencionadas. El precipitado se liofilizó para usos posteriores. Hidrólisis Enzimática Una solución al 4% del aislado proteico de garbanzo fue hidrolizado en los tiempos 0, 0.5, 1.0, 1.5, 2.0, 2.5 y 3.0 h con tres enzimas puras: quimotripsina, tripsina y pepsina. La temperatura fue de 37 oC, para las tres enzimas y la relación enzima- sustrato manejada fue de 0.01. El pH de la solución se ajustó al valor óptimo de la acción enzimática (tripsina y quimotripsina pH 8.0, pepsina pH 2.0). La hidrólisis fue controlada registrando el valor del gasto de ácido o álcali para mantener el pH constante. La inactivación de las enzimas se realizó elevando la temperatura a ebullición durante 5 min, en el caso de la pepsina, previamente se elevo el pH a 8.0. Después de la inactivación se procedió a centrifugar las muestras a 10 000 rpm por 30 min, y se recuperó el sobrenadante en el cual se encuentra teóricamente la proteína soluble. Electroforesis La electroforesis se realizó utilizando gel de poliacrilamida con dodecil sulfato de sodio (SDS-PAGE) utilizando un gel separador al 13 % de poliacrilamida con un gel concentrador al 5% de poliacrilamida, en un equipo Minin-Protean 3 de Bio-Rad. La muestra de aislado proteico (1mg de proteína) se solubilizó en 500 µL de agua desionizada con el mismo volumen del regulador de muestras (0.1 M Tris-HCl, pH 6.8, 2% SDS, 5% de 2. Mercaptoetanol y Azul de bromofenol al 0.02%). Las muestras de los hidrolizados se mezclaron en una relación 1:1 (v/v) con el regulador de muestra. El gel se tiñó con una solución conteniendo 40 % de metanol, 7 % de ácido acético y Azul de Comassie R-250 (0.125%). Poder de Inhibición de la ECA de los Hidrolizados La actividad de la enzima convertidora de la angiotensina I (ECA-I) se determinó utilizando el método colorimétrico de Hayakari et al (1978)7. El cual se basa en la reacción entre el ácido hipúrico y el cloruro de cianuro. La mezcla de reacción contenía 100 mU/mL de ECA-I, el péptido inhibidor derivado de los hidrolizados, regulador de fosfato de potasio 40 µmol, regulador NaCl 300 µmol y el sustrato sintético HHL al 0.3 %. Esta mezcla se incubó a 45 min con agitación a 37 oC y la reacción se detuvo adicionando la solución de cloruro de cianuro (TT) en dioxano al 3% (p/v) y regulador de fosfato de potasio 0.2 M. La absorbancia a 382 nm fue determinada en el sobrenadante al someter la mezcla de reacción a centrifugación a 10 000 rpm por 10 min a 4oC. El poder inhibitorio IC50 es la concentración de péptido en mg de proteína/ml requeridos para inhibir el 50% de la actividad de la

Hidrólisis enzimática
0
1
2
3
4
5
6
7
0 0,5 1 1,5 2 2,5 3 3,5
Tiempo (horas)
ml
de
Na
OH
o H
Cl0
,1 N
tripsinaquimiotripsinapepsina
ECA-I en las condiciones, y se determinó por regresión lineal del porciento de inhibición de la ECA-I versus la concentración del péptido. RESULTADOS Y DISCUSIÓN Análisis Proximal del Aislado Proteico El contenido de proteína se incrementó de 18.48 g/100g en la harina de garbanzo a 73.75 g/100g en el concentrado proteico. Sin embargo, el contenido de lípidos en el concentrado (14.85 g/100g) representa un valor alto, por lo cual se recomienda usar harina desengrasada (Sánchez-Vioque et al, 1999). El rendimiento de la proteína recuperada fue del 43 % (Tabla 1).
Tabla 1. Análisis químico proximal de la harina y del concentrado proteico de garbanzo (Cicer arietinum L) (g/100g de muestra) Análisis
Harina Concentrado
Humedad Cenizas Proteína Lípidos Fibra total dietaria Carbohidratosa
8.32 ± 0.42 3.15 ± 0.3 18.48 ± 0.94 6.04 ± 0.30 3.0 ± 0.15 61.01 ± 3.05
8.68 ± 0.43 2.40 ± 0.12 73.75 ± 3.69 14.85 ± 0.74 0.0 0.32 ± 0.02
El factor para el porciento de proteína fue de 6.25 Las determinaciones se realizaron por triplicado. Los resultados muestran el promedio y su correspondiente desviación estándar a Calculado por diferencia. Figura 1 Cinética de hidrólisis de los concentrados proteicos tratados con pepsina
(∆), tripsina ( ) y quimotripsina ( ) Hidrólisis enzimática La Figura 1 muestra que se obtuvo una mayor hidrólisis del concentrado proteico de garbanzo utilizando la enzima pepsina. La hidrólisis alcanzada con el tratamiento con tripsina y quimotripsina es parecida siendo ligeramente mayor la tripsina. Este comportamiento resulta congruente al analizar la proporción de

M C 0.0 0.5 1.0 1.5 2.0 2.5 3.0M C 0.0 0.5 1.0 1.5 2.0 2.5 3.0KDa
25015010075
50
37
25
20
15
M C 0.5 1.0 1.5 2.0 2.5 3.0 0.0M C 0.5 1.0 1.5 2.0 2.5 3.0 0.0KDa
75
50
37
25
20
15
100150250
M C 0.0 0.5 1.0 1.5 2.0 2.5 3.0M C 0.0 0.5 1.0 1.5 2.0 2.5 3.0KDa
75
50
37
25
20
15
100150250
6,04
14,36
0
1
2
3
4
5
6
7
0 0,5 1 1,5 2 2,5 3 3,5
Tiempo (horas)
Gra
do d
e hi
dról
isis
, ml H
Cl 0
,1N
0
2
4
6
8
10
12
14
16
% In
hibi
ción
de
la E
CA
-I
% Inhibición de ECA Grado de hidrólisis40,96
24,78
14,00
0
0,5
1
1,5
2
2,5
3
3,5
0 0,5 1 1,5 2 2,5 3 3,5
Tiempo (horas)
Gra
do d
e hi
dról
isis
, ml N
aOH
0,1
N
0
5
10
15
20
25
30
35
40
45
% In
hibi
ción
de
la E
CA
-I
% Inhibición de la ECA Curva de hidrólisis
7,84
15,03
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
2
0 0,5 1 1,5 2 2,5 3 3,5
Tiempo (horas)
Gra
do d
e H
idró
lisis
, ml N
aOH
0,1
N
0
2
4
6
8
10
12
14
16
% In
hibi
ción
de
la E
CA
% Inhibición de ECA Grado de Hidrólisis
aminoácidos para los cuales cada enzima es específica en la proteína de garbanzo. Figura 2. Perfil electroforético del aislado proteico de garbanzo tratado con a) Pepsina, b) Tripsina, c) Quimotripsina. Los pesos moleculares del marcador (Carril M) se encuentran de lado izquierdo. Carril C= aislado proteico de garbanzo sin tratamiento, Carril 0.0-3.0= tiempo de hidrólisis en h. Figura 3. Cinética de hidrólisis de los hidrolizados proteicos de garbanzo (Cicer arietinum L) y la Capacidad Inhibitoria (porciento de Inhibición) obtenidos por el tratamiento con pepsina (a), tripsina (b) y quimotripsina (c).
a) b) c)
a)b)
c)

El perfil electroforético (Figura 2) de los hidrolizados obtenidos concuerda con los resultados de la Figura1. El aislado proteico de garbanzo fue más susceptible a la degradación con pepsina (Figura 2a). En la primer h, las bandas de 96, 70 y las de rango del 46-55 kDa, desaparecieron. Los perfiles del aislado proteico tratado con tripsina y quimotripsina se muestran en las Figuras 2b y 2c, respectivamente. Las bandas de 30-96 kDa son hidrolizadas rápidamente con la tripsina en comparación con la quimotripsina, sin embargo la degradación de las bandas después de la primera h no continua. El perfil de los hidrolizados obtenidos con la quimotripsina muestra que fue menos efectiva que la tripsina y la pepsina en degradar las bandas de alto peso molecular. Actividad antihipertensiva. La actividad antihipertensiva calculada para los hidrolizados proteicos de garbanzo obtenidos, muestra que el hidrolizado que fue tratado con tripsina durante 3 h presentó mayor poder inhibitorio (IC50= 0.613 mg/ml). En la Figura 3 se presentan los resultados de la actividad inhibitoria de la ECA-I de los hidrolizados de concentrados proteicos de garbanzo relacionándolos con la cinética de hidrólisis. Para la pepsina (Figura 3a) se observó que al aumentar la hidrólisis aumentó también el porciento de inhibición, siendo el valor máximo de 14.36 % a las 3 h. En comparación con la tripsina (Figura 3b), el porciento de inhibición a las 0.5 h fue de 24.78%, pero disminuyó a las 2 h a 14 % y a las 3 h aumentó nuevamente a 40.96%, demostrando que no existe una relación directa entre el grado de hidrólisis y la actividad antihipertensiva. Los resultados de los hidrolizados obtenidos con la quimotripsina (Figura 3c), muestran un porciento de Inhibición máximo de 15.03%, valor cercano al encontrado con la pepsina en el mismo tiempo (3h), a pesar de que con la pepsina se hidrolizó más la proteína que con la quimotripsina. CONCLUSIONES Los resultados encontrados en este trabajo demuestran que los hidrolizados del aislado proteico de garbanzo tratados con las enzimas digestivas pepsina, tripsina y quimotripsina, presentan péptidos con capacidad de inhibir la enzima convertidora de angiotensina-I. El hidrolizado con mayor poder antihipertensivo encontrado fue el tratado con tripsina durante 3h, y presentó un 40.96 % de inhibición. Los hidrolizados con actividad antihipertensiva podrían ser usados en la elaboración de dietas específicas como alternativa en el tratamiento de pacientes hipertensos. REFERENCIAS AOAC. Oficial Methods of Analysis; (1995); 16th edition. Association of Oficial Analytical Chemist. Washington, D.C.; USA. Hayakari, M., Kondo, Y., y Izumi, H.; (1978); A rapid and simple spectrophotometric assay of angiotensin-converting enzyme. Analitical Biochemistry; 84. 361-369

Matsufuji, H., Matsui, T., y Ohshige, S.; Kawasaki, T.; Osajima, K.; y Osajima, Y.; (1995); Antihypertensive effects of angiotensina fragments in SHR; Bioscience Biotechnology Biochemistry; 59, 1938-1401. Matsui, T., Li, C.H., Tanaka, T., Maki, T., Osajima, Y., y Matsumoto, K.; (2000); Depressor effect of wheat germ hydrolysate and its novel angiotensin I-converting enzyme inhibitory peptide, Ile-Val-Tyr, and the metabolism in rat and human plasma;. Biological Pharmaceutical Bulletin; 23, 427-431. Pitt, B.; (1997); ACE inhibitors in heart failure: prospects and limitations; Cardiovascular Drug Therapy; 11, 285–290. Sánchez-Vioque, R., Clemente A., Vioque, J., Bautista J., y Millán, F.; (1999); Protein isolates from chickpea (Cicer arietinum L.): chemical composition, functional properties and protein characterization; Food Chemistry ; 64, 237-243. Velázquez Monroy O, Rosas Peralta M, Lara Esqueda A, Pastelín Hernández G, Grupo ENSA 2000, Attie F, Tapia Conyer R. Hipertensión arterial en México: Resultados de la Encuesta Nacional de Salud (ENSA) 2000. Arch Cardiol Mex 2002;72(1):71-84. Vioque, J.; Sánchez-Vioque, R.; Clemente, A.; Pedroche, J.; Yust, M. M., y Millán, F.; (2000); “Péptidos bioactivos en proteínas de reserva”; Grasas y aceites; Vol. 51. Fasc.5; 361-365. Yamamoto, N.; (1997) Antihypertensive peptides derived from food protein. Biopolymers, 43:129-43.

OBTENCIÓN DE PÉPTIDOS BIOACTIVOS CON ACTIVIDAD ANTIHIPERTENSIVA APARTIR DE SEMILLA DE FRÍJOL (Phaseolus vulgaris
var. Jamapa).
Hernández-Alvarez, A. Ja*, Carrasco-Castilla, Ja, Dávila-Ortíz, G.a, Jiménez-Martínez, Ca.
a Escuela Nacional de Ciencias Biológicas-Instituto Politécnico Nacional, Prol. Carpio, Esq. Plan de Ayala, S/N. Col. Casco de Santo Tomás. Del. Miguel Hidalgo. C.P. 11340. México, D. F. Tel.: (+52)
55 57296000 Ext. 62462. *[email protected]
RESUMEN: El Fríjol (Phaseolus vulgaris) var. Jamapa es una leguminosa típica de México. La importancia que el fríjol tiene en la dieta de nuestro país es indiscutible, hoy representa 15% de los alimentos que ingieren habitantes de zonas rurales y urbanas. De la semilla de fríjol Jamapa (21% de proteína), por precipitación isoeléctrica se obtuvo un concentrado proteico (74% proteína), se utilizaron dos enzimas comerciales (AlcalaseMR (AL) y FlavourzymeMR (FL)) para la producción de un aislado proteico. El perfil electroforético (SDS-PAGE) revelo para la muestra tratada con FL la degradación de las cadenas ácidas y básicas de la proteína tipo 11S, por otro lado la muestra tratada con AL revelo una banda tenue que posiblemente corresponde a la cadena ácida de la proteína tipo 11S, misma que desapareció completamente a los 45 min de hidrólisis. Los valores de mejor actividad inhibitoria de la ECA fueron con los hidrolizados proteicos obtenidos con AL y FL a los 15 min y 30 min con 0.231 y 0.344 mg/mL respectivamente. Los resultados anteriores sugieren que el hidrolizado proteico de P. vulgaris var. Jamapa preparado con dichas enzimas puede ser utilizado para la elaboración de alimentos funcionales con actividad antihipertensiva. ABSTRACT: The Bean (P. vulgaris) var. Jamapa is a typical Mexican legume. The importance that the bean has had in the diet of our country is undeniable, today it represents 15% of the food intake that inhabitants of rural and urban zones consume. Of the seed of bean Jamapa (21% of protein), by isoelectric precipitation it was obtained a protein concentrate (74% protein), which was used as starting material for the production of an enzymatic hydrolysate using two commercial enzymes (AlcalaseMR (AL) and FlavourzymeMR (FL)). The electrophoresis (SDS-PAGE) revealed in the protein profile for the sample treated with FL the degradation of the acid and basic chains of the 11S like protein, furthermore the protein profile of the sample treated with AL reveals a band with less intensity of the acid chains of the 11S like protein. The best ACE inhibitory activities obtained with the protein hydrolysates treated with AL and FL were at 15 min and 30 min with an IC50 value of 0.231 and 0.344 mg/mL respectively. The previous results suggest that the protein hydrolysate of P. vulgaris var. Jamapa prepared with the above mentioned enzymes could be used for the production of functional foods with antihypertensive activity. Palabras clave: Actividad Inhibitoria de la ECA, Concentrado Proteico, Grado de Hidrólisis. INTRODUCCIÓN Las proteínas, determinan las propiedades físicas y sensoriales de muchos alimentos. Así, la consistencia y textura de la carne, leche, queso o pan, dependen en gran medida de la naturaleza de las proteínas que los constituyen. Pero también en los alimentos elaborados con una presencia menor de proteínas pueden jugar también un papel muy importante, influyendo en propiedades

funcionales, como la absorción de agua o aceite o la formación de emulsiones, geles y espumas (Zayas, 1997). Las proteínas también constituyen un aporte nutricional importante, representando una fuente de energía, nitrógeno y aminoácidos esenciales. Así mismo cubren necesidades fundamentales como la alimentación y satisfacer los sentidos desde el punto de vista funcional y sensorial, pueden funcionar como agentes que previenen enfermedades, ya que son una excelente materia prima para la obtención de péptidos bioactivos (Clemente et. al., 1999). Los péptidos bioactivos son cadenas de aminoácidos relativamente pequeñas de 2 a 15 residuos, que se encuentran inactivos dentro de la proteína precursora pero que pueden ser liberados, durante la digestión del alimento al organismo del individuo ó por un procesado (hidrólisis enzimática o fermentación) previo del mismo (Vioque et. al., 2000) los cuales pueden ejercer diversas funciones metabólicas. Las leguminosas, son importantes nutritiva y económicamente por su presencia en los alimentos de millones de personas de todo el mundo, ya que contienen proteínas (el doble o triple de la mayoría de los cereales) y son una valiosa fuente de energía (De Mejía, 2003). En los países en vías de desarrollo, las leguminosas contribuyen con el 10% de las proteínas diarias y el 5% del aporte energético. El fríjol común (Phaseolus vulgaris) es un alimento ampliamente consumido en diversas regiones de Latinoamérica y África, y posee buen valor nutricional, proporcionando proteínas (20-28%), carbohidratos (15-60%), vitaminas del complejo B y minerales. En años recientes ha recibido mayor atención debido a su carácter nutracéutico, ya que proporciona algunos compuestos que juegan un papel importante en la prevención de enfermedades crónico degenerativas (De Mejía, 2003). México, como centro de origen del fríjol posee una gran variedad de ellos y se considera a los silvestres y enmalezados fuente de recursos genéticos debido a su mayor diversidad. Sin embargo, son escasos los estudios enfocados a conocer el potencial nutricional y nutracéutico de estos materiales (Espinosa et al., 2006). Debido a todo lo anterior, se hace necesario seguir buscando nuevas fuentes ricas en proteínas tal como el fríjol (P. vulgaris var. Jamapa) y evaluar sus fracciones peptídicas, obtenidas mediante la hidrólisis enzimática de un aislado proteínico, así como conocer su actividad antihipertensiva tanto in vitro como en pruebas clínicas en humanos. METODOLOGIA Materiales Se utilizaron granos de fríjol jamapa (P. vulgaris var. Jamapa) cosecha 2005, donados por la Dra. Carmen Jacinto Hdéz, de CEVAMEX, Campus Texcoco. Todos los reactivos utilizados en los análisis son de marcas registradas como Sigma y J.T. Baker (Phillisburg, N.J, USA). Las enzimas fueron adquiridas de los laboratorios Sigma (Sigma Chemical Co., St. Louis, MO, USA); Merck (Darmstadt, Germany) y Novo (Copenhague, Denmark). Composición proximal de la harina y concentrado

Se determinaron de acuerdo a los procedimientos oficiales descritos por la AOAC, (1997). Obtención del Concentrado proteico Se utilizó el método de Pedroche et. al. (2002). Se realizó una suspensión de harina en agua destilada en una relación 10:200 p/v (harina/agua), se ajustó el pH a 9.5 con una solución de NaOH 1N y se dejó en agitación por 30 min con un agitador mecánico (Caframo RZ-1) a 400 rpm y a una temperatura de 40-50°C. Posteriormente se centrifugó a 5000 x g por 30 min, se desecho el precipitado y se ajustó el sobrenadante a un pH de 4.5 con HCl 1 N, posteriormente se centrifugó a 10,000 rpm por 30 min, y se desecho el sobrenadante para recuperar el precipitado, el cual se seco a -47°C y 13x10-3 mbar en un liofilizador Labconco. Determinación de Compuestos Fenólicos Los compuestos fenólicos totales expresados como mg eq.(+)-catequina/100g de semilla, se cuantificaron de acuerdo al método de Deshpande y Cheryan (1987). Los autores sugieren que estos equivalentes de (+)-catequina en fríjol incluyen flavonoides mono y poliméricos, así como flavonoles y dihidrocalconas. La extracción de compuestos fenólicos en P. vulgaris var. Jamapa y en el concentrado obtenido con y sin testa se llevó a cabo por el método descrito por Deshpande y Cheryan (1985 y 1987), y se realizó como sigue: 1 g de muestra molida, se le adicionaron 10 mL de metanol acidificado (1% HCl), se mantuvo en agitación durante 16-20 h a 30 °C. A 1 mL de la solución se le adicionaron 2.5 mL de reactivo de vainillina al 1% en metanol (fresca) y 2.5 mL de HCl al 8 % en metanol, agregando 1-1 para llevar a cabo la reacción, después se colocó a 30 °C por 20 min e inmediatamente se leyó su absorbancia a 500 nm en un espectrofotómetro Perkin Elmer Lambda 2S. La concentración se calculó con base a una curva de calibración de (+)-catequina (0,0.25, 0.50, 0.75 y 1 mg). Hidrólisis Enzimática Para la hidrólisis del concentrado proteínico se utilizó el método descrito por Pedroche et. al. (2002), para el cuál se empleó un diseño de bloques aleatorios, donde los bloques fueron las enzimas comerciales empleadas (AlcalaseMR 2.4 L FG y FlavuorzymeMR 500 MG) (Novo Nordisk, Bagsvaerd, Denmark), el factor que se evaluó fue el tiempo de reacción (0, 15, 30, 45 y 60 min.) y la variable de respuesta fue el grado de hidrólisis. Se realizo una curva de hidrólisis, la cuál se obtuvo mediante la técnica de pH-stat usando los siguientes parámetros de hidrólisis: concentración de sustrato 4%; enzima/sustrato en una proporción 0.3 UA/g-1 de AlcalaseMR y 50 UAPL/g-1 de FlavourzymeMR; a pH 7 para la AlcalaseMR y 8 para Flavourzyme MR; a una temperatura de 50°C. La hidrólisis se llevó a cabo en un vaso de 200 mL para hidrólisis, equipado con un agitador mágnetico, termómetro y un eléctrodo para medir pH. La hidrólisis fue detenida por acidificación a pH 4. El hidrolizado fue centrifugado a 10,000 rpm por 30 min., para eliminar el sustrato insoluble y la enzima residual. El material centrifugado se liofilizó y almacenó en congelación a -20°C. Electroforesis en gel de Poliacrilamida

La electroforesis se realizó utilizando gel de poliacrilamida con dodecil sulfato de sodio (SDS-PAGE), un gel separador al 13% de poliacrilamida y un gel concentrador al 5% de poliacrilamida, en un equipo Minin-Protean3 de Bio-Rad. La muestra de concentrado proteíco (1mg de proteína) se solubilizó en 500 µL de agua desionizada con el mismo volumen del regulador de muestras (0.1M Tris-HCL, pH 6.8, 2% SDS, 5% de 2-Mercaptoetanol y Azul de bromofenol al 0.02%). Las muestras de los hidrolizados de 50 µL, se mezclaron con el regulador de muestra al mismo volumen y todas las muestras se agitaron en un vortex para posteriormente calentarlas a ebullición por 5 min. Se aplicaron 15 µL tanto para la muestra como para el marcador en cada pozo del gel. La separación electroforética se realizo a corriente constante a 110 V durante 90 min. Posteriormente el gel se colocó en una solución de ácido tricloroácetico al 12.5% por 30 min. El gel se tiñó con una solución conteniendo 40% de metanol, 7% de ácido acético y Azul de Comassie R-250 (0.125%) durante toda la noche. El proceso de desteñido consistió, en lavar el gel en una solución de etanol al 50% por 15 min tres veces, después se aplicó la solución de etanol-ácido acético-agua (30-7-63, v/v) por 30 minutos y finalmente se lavó con un solución metanol-ácido acético-agua (25-10-65, v/v) por 30 min. El PM de las proteínas se determinó mediante una gráfica en la cual se traza la movilidad relativa de estándares de referencia contra el logaritmo del PM, y con la ecuación de la línea recta se interpola el valor correspondiente de la movilidad relativa de las fracciones proteicas analizadas. Evaluación in vitro de la actividad inhibitoria de la ECA de las fracciones peptídicas La actividad inhibitoria de la ECA fue determinada de acuerdo al método de Hayakari et. al. (1978). Dicha actividad se le determino a la fracciones peptídicas obtenidas. Estas fracciones están en forma líquida. La hidrólisis de la ECA con HHL (Hippuryl-L-Histidyl-L-Leucina) produce ácido hipúrico e his-leu. Este método se basa en la reacción colorimétrica del ácido hipúrico con TT (2, 4,6-tricloro-triasina). Cada mezcla de 0.5 mL a analizar tuvo los siguientes componentes a la concentración final indicada, regulador de fosfato de potasio 100mM; cloruro de sodio 600mM; HHL 3mM y 2mg/mL de enzima. Esta mezcla fue incubada a 37°C por 15 min. La reacción se detuvo por la adición de 1.5 mL de TT (3% w/v) en dioxano y 3 mL de regulador de fosfatos (0.2 M) a pH 8.3. Posteriormente se centrifugo a 10,000 x g por 10 min. Del sobrenadante se obtuvo la absorbancia la cuál fue medida a 382 nm. El valor de IC50 es definido como la concentración de péptidos en 1mg de proteína/mL -1 requerido para producir el 50% de la inhibición de la ECA en las condiciones descritas anteriormente y se determino por análisis de regresión el porcentaje (%) contra la concentración de péptidos. RESULTADOS Y DISCUSIÓN Obtención del Concentrado proteico Para la obtención del concentrado proteínico se utilizaron 500 g de harina de fríjol P. vulgaris var. Jamapa, con un contenido de proteína verdadera de 21.17% (b.s.). Posteriormente de 500 g de harina se consiguió un rendimiento de 58 g de concentrado proteínico ya liofilizado, con un contenido de proteína verdadera de

73.6%, el cuál es similar a los valores encontrados por Chel-Guerrero et. al., (2002) en harina y concentrado de P. lunatus (25.5 y 72.0 % respectivamente, de proteína cruda). De todo el proceso se logro una eficiencia de recuperación de proteína verdadera de 40.32 %. Las proteínas en su superficie siempre poseen una carga eléctrica que depende del pH, de los electrolitos presentes y del disolvente. Composición Proximal de la Harina y el Aislado La caracterización proximal de la harina (HP) y el concentrado proteínico de P. vulgaris var. Jamapa (CPPJ) se presenta en el Cuadro 1 y se tuvo una concentración de proteína cruda de 21.17% y 73.6% respectivamente. Los datos obtenidos de los materiales en estudio, fueron semejantes a los reportados por Betancur-Ancona et. al. (2004) ya que en la harina y en el concentrado proteínico de P.lunatus reportan 25.50 y 71.13% respectivamente. Cuadro 1. Composición proximal de la harina (HP) y concentrado proteínico de P. vulgaris var. Jamapa (CPPJ).
Componentes (%)
HarinaP. vulgaris
Concentrado P. vulgaris
Humedad 9.44 ± 0.50 8.34 ± 0.50Proteína 21.17 ± 2.00 73.60 ± 2.10Grasa 2.30 ± 0.50 4.24 ± 0.50Fibra 5.40 ± 0.80 0.25 ± 0.20Cenizas 4.20 ± 0.20 2.32 ± 0.30Carbohidratos* 57.44 ± 0.50 11.25 ± 0.50
*Calculados por diferencia Los resultados representan la media de tres determinaciones independiente ± DE El contenido de grasa encontrado en la HP y CPPJ en estudio fue de 2.3 y 4.24% respectivamente, los cuales son similares a los reportados por Betancur-Ancona et. al. (2004) con 0.65% para la harina y 0.68% para el concentrado proteínico de P. lunatus. Cabe mencionar que el alto contenido de grasa encontrado en el CPPJ pudiera ser debido a que durante la extracción del concentrado se usaron pH’s alcalinos (pH 9.5 con NaOH), por que la grasa pudo haber interactuado con el álcali pudiendo saponificar las grasas principalmente de naturaleza polar, y éstas a su vez interactuar con las proteínas del concentrado y al momento de llevar las proteínas al punto isoeléctrico para su precipitación, las grasas precipitaron junto con las proteínas, tal como lo menciona Sánchez-Vioque et. al. (1999) para el chícharo (Cicer arietinum L.) y Chel-Guerrero et. al. (2002) para el concentrado proteínico de C. ensiformis. Sin embargo, está saponificación probablemente es depediente de la composición de ácidos grasos de la leguminosa. Concentración de Compuestos Fenólicos A partir de los resultados obtenidos podemos observar claramente que existe una pequeña diferencia entre la concentración de compuestos fenólicos del CPPJ de

P. vulgaris var. Jamapa con testa y sin testa, para el CPPJ sin testa se obtuvo una concentración aproximada de compuestos fenólicos de 1.61 mg de (+)-catequina/g de semilla, mientras que para el CPPJ con testa se obtuvo una concentración aproximada de compuestos fenólicos de 5.72 mg de (+)-catequina/g de semilla; la concentración de compuestos fenólicos debió haber aumentado debido a que durante la extracción del concentrado se utilizaron pH’s alcalinos y ácidos, y el grupo fenol (un anillo aromático unido a al menos un grupo funcional hidroxilo) pudo haber interactuado con el álcali o el ácido; mientras que los taninos son considerados derivados del catecol (estos pueden formar polímeros y estar condensados), por lo que se generan polímeros de alto peso molecular (flobáfenos), generalmente estos son compuestos de elevado peso molecular y contienen un número elevado de grupos hidroxilo capaces de formar uniones efectivas entre proteínas y otras macromoléculas (Bartolomé et al., 1997). Las proantocianidinas han sido detectadas en diferentes variedades de fríjol común en una concentración aproximada de 9.4-37.8 mg equivalentes de (+).catequina/g, lo cuales se encuentran en gran proporción en la testa (De Mejía, 2003). Electroforesis en Gel de Poliacrilamida SDS-PAGE Los perfiles electroforéticos de los hidrolizados proteícos de P. vulgaris var. Jamapa tratados por medio de un diseño de bloques aleatorios con las enzimas comerciales AlcalaseMR y FlavourzymeMR (Figura 1), muestran la degradación del concentrado proteico a moléculas de menor peso molecular y eventualmente a péptidos. Figura 1. Perfil electroforético de los hidrolizados de P. vulgaris var. Jamapa con las enzimas comerciales AlcalaseMR y FlavourzymeMR a diferentes tiempos de hidrólisis (15 – 60 min). M= marcador, C= concentrado proteico, a= Albuminas 100 kDa, b= Subunidad 7S (75 kDa), c= Subunidad 11S (60.15 kDa),d= Cadenas Ácidas (37.2 kDa), e= Cadenas Básicas (26.19kDa). (Yust et al, 2003) El SDS-PAGE del concentrado muestra un perfil con la presencia de complejos proteicos cuyos pesos moleculares se encuentran en el intervalo de 157 a 17.84

kDa, se observa así también las bandas que corresponden a las cadenas α y β de la proteína tipo 11S, liberadas por la acción del 2-mercaptoetanol que es un agente reductor de puentes disulfuro. Las bandas de 60.15 kDa y 75 kDa aparentemente corresponden a las subunidades que conforman a la globulina tipo 11S y 7S respectivamente. También se observa la presencia de una banda de aproximadamente 100 kDa, la cual podría equivaler a las albúminas (Yust et al, 2002). En el perfil se muestra que para el caso de la hidrólisis con ambas enzimas, estas presentan una degradación efectiva, el perfil que se muestra para los hidrolizados con FlavourzymeMR desde los 15 min se observa una banda con un peso molecular aproximado a 37.23 kDa la cuál corresponde posiblemente a las cadenas ácidas de la proteína tipo 11S (PM aproximado de 35 kDa). Es importante mencionar que esta banda va disminuyendo conforme avanza la reacción, se muestra que a los 60 min la banda de las cadenas ácidas se torna muy tenue; ya que tanto la subunidad tipo 7S y 11S desparecen casi de inmediato que se inicia la hidrólisis; también se observa una banda con un peso molecular aproximado de 17.8 kDa, la cuál coincide con las cadenas básicas de la proteína tipo 11S (PM aproximado de 20 kDa), aunque esta banda se degrada casi en su totalidad a los 60 min, alrededor de los 45 min se observa que aparece una banda con un peso molecular de 5.29 kDa, obteniéndose péptidos de muy bajo peso molecular, los cuales pueden presentar sin duda actividad antihipertensiva (Pedroche et al., 2002). El perfil que se muestra para la hidrólisis con AlcalaseMR presenta una banda con peso molecular de 37.23 kDa la cual pertenece a las cadenas ácidas de la proteína tipo 11S, aunque esta banda se observa muy tenue en comparación con la banda obtenida para FlavourzymeMR, degradándose completamente a los 45 min de hidrólisis (Clemente et al., 1999). En cuanto a las cadenas básicas de la proteína tipo 11S solamente se visualizan para los 15 min y se tornan muy tenues a los 30 min, ya que posteriormente la banda desaparece en su totalidad. Evaluación in vitro de la actividad inhibitoria de la ECA Se determinó la capacidad inhibitoria de la Enzima Convertidora de Angiotensina I, de los hidrolizados obtenidos a diferentes tiempos de hidrólisis. La actividad inhibitoria de la ECA mostrada por los hidrolizados proteicos obtenidos con las enzimas comerciales a diferentes tiempos de reacción, sugiere que los péptidos liberados de las proteínas intactas de la leguminosa por la hidrólisis enzimática, fueron los responsables de la inhibición de la ECA (Cuadro 3). Cuadro 3. Actividad inhibitoria in vitro de los hidrolizados proteicos sobre la ECA.
Tiempo de reacción (min)
Frijol Jamapa IC50 (mg/mL)AlcalaseMR FlavourzymeMR
15 0.231 0.70030 0.312 0.34445 0.463 0.75760 0.380 0.417
Las pruebas de inhibición de la ECA de los hidrolizados proteicos de P. vulgaris var. Jamapa con AlcalaseMR mostró una mayor actividad inhibitoria de la ECA a los 15 min de reacción con un IC50 = 0.231 mg/mL. Por otro lado, el hidrolizado de P.

vulgaris var. Jamapa con FlavourzymeMR tuvo una mayor actividad inhibitoria de la ECA a los 30 min de reacción con un IC50= 0.344 mg/mL, mientras que los demás tiempos de reacción oscilaron entre IC50= 0.417 a 0.757 mg/mL, para los péptidos bioactivos con actividad antihipertensiva, el intervalo que se maneja varía desde 0.025 a 2.4 mg/mL. Los valores antes mencionados fueron similares a los reportados por Hong et al., (2005) en el frijol mungo (P. radiatus L.) a las 2 h de reacción con AlcalaseMR con una IC50= 0.64 mg/mL. También cabe mencionar que diversos investigadores han reportado diversos valores de IC50 en un intervalo entre 0.20 y 246.7 mg/mL (Hong et al, 2005, Kim et al., 1990). Debido a lo anterior sugiere que los valores encontrados en este trabajo se encuentran dentro de los intervalos de inhibición reportados por diversos investigadores con otras materias primas tanto de origen vegetal como animal. CONCLUSIONES La hidrólisis extensiva de las semillas de fríjol Phaseolus vulgaris var. Jamapa utilizando las proteasas comerciales AlcalaseMR y FlavourzymeMR, nos proveen de un hidrolizado rico en péptidos bioactivos con efectos fisiológicos directos, como lo es la actividad antihipertensiva. Los hidrolizados proteicos obtenidos en esta investigación no solamente muestran mejoras en las características nutricionales y funcionales, sino también contienen péptidos bioactivos los cuales incrementan el valor del producto y amplían el campo de aplicaciones de las proteínas de fríjol, por ejemplo en la fortificación de alimentos líquidos ó bebidas energéticas. Otras investigaciones son necesarias para examinar si los péptidos obtenidos exhiben actividad antihipertensiva in vivo; así como también el aislamiento y la caracterización de los péptidos responsables de dicha función. REFERENCIAS A.O.A.C. 1995. Official methods of analysis of the Association of Analytical Chemist, 15th Ed. Washington D.C. Betancur-Ancona, D.; Gallegos-Tintoré, S., and Chel-Guerrero, L. 2004. Wet-Fractionation of Phaseolus lunatus Seeds: Partial Characterization of Starch and Protein. J. Sci Food Agric, 84(10): 1193-1201. Chel-Guerrero, L.; Pérez-Flores, V.; Betancur-Ancona, D., and Dávila-Ortiz G. 2002. Functional Properties of Flours and Protein Isolates from Phaseolus lunatus and Canavalia ensiformis Seeds. J. Agric. Food Chem, 52: 5895-5901. Clemente, A.; Vioque, J. and Millán, F. 1999. Hidrolizados Proteicos de Origen Vegetal. Departamento de Fisiología y Tecnología de Productos Vegetales. Instituto de la Grasa, 2(6): 25-32. De Mejía, E.; Guzmán-Maldonado, S.H.; Acosta-Gallegos, J.A.; Reynoso-Camacho, R.; Ramírez-Rodríguez, E.; Pons-Hernádez, J.L.; González-Chavira, M. M.; Castellanos, J.Z.; Nelly, J.D. 2003. Effect of cultivar and growing location on the trypsin inhibitors, tannins and lectins of common beans (Phaseolus vulgaris L.) grown in semiarid highlands of Mexico. J. Agric. Food Chem., 51, 5956-5966.

Desphande S.S. and Cheryan M. 1987 .Determination of phenolic compounds of dry beans using vanillin, redox and precipitation assay. J. Food Sci, 52 (2): 332-334. Espinosa, G. L.C. 2006. Hidrólisis Enzimática del Concentrado Proteínico de Vigna unguiculata para su Incorporación a un Alimento para Régimen Especial. Tesis de Maestría. Facultad de Ingeniería Química de la Universidad Autónoma d Yucatán. pp 40-42. Hayakari, M.; Kondo, Y., and Izumi, H. 1978. A Rapid and Simple Espectrophotometric Assay of Angiotensin-Converting Enzyme. Analytical Biochemistry, 84: 361-369. Hong, L. G.; Wei, L. G.; Liu, H., and Hui, S. Y. 2005. Mung-Bean Protein Hydrolysates Obtained with Alcalase Exhibit Angiotensin I-Converting Enzyme Inhibitory Activity. Food Sci. Tech Int, 11(4): 281-287. Kim, Y. S.; Park, W. S. P., and Rhee, C. K. 1990. Functional Properties of Proteolytic Enzyme Modified Soy Protein Isolate. Journal of Agricultural and Food Chem, 38(3): 651-656. Pedroche, J.; Yust, M. M.; Girón-Calle, J.; Alaiz, M. Millán, F., and Vioque, J. 2002. Utilization of Chickpea Protein Isolates for Production of Peptides with Angiotensin I-Converting Enzyme (ACE)-Inhibitory Activity. J. Sci. Food Agric, 82: 960-965. Sánchez-Vioque, R.; Clemente, A.; Vioque, J.; Bautista, J., and Millán, F. 1999. Protein Isolates from Chickpea (Cicer arietinum L.): Chemical Composition, Functional Properties and Protein Characterization. Food Chem, 64: 237-243. Vioque, J.; Sánchez-Vioque, R.; Clemente, A.; Pedroche, J.; Yust, M. M., y Millán, F. 2000. Péptidos Bioactivos en Proteínas de Reserva. Grasas y Aceites, 51(5): 361-365.

LACTOSUERO Y SU PROBLEMÁTICA EN EL MEDIO AMBIENTE
Guerrero Rodríguez W. J.a, Gómez Aldapa C. A.a*, González Ramírez C. A.a, Castro
Rosas J.a
aCentro de Investigaciones Químicas, ICBI, Universidad Autónoma del Estado de Hidalgo. Carretera Pachuca-Tulancingo, Km. 4.5. C.P. 42070 Pachuca, Hidalgo. México
RESUMEN: El lactosuero es un subproducto que se genera en la industria láctea, sus principales componentes son la lactosa, proteínas y algunos minerales. En la zona lechera del municipio de Acatlán, Hidalgo, existe un gran número de industrias procesadoras de lácteos, en su mayoría utilizan sistemas rudimentarios de producción. Este subproducto tiene poco valor comercial para los productores, una pequeña parte es utilizada en la alimentación del ganado, mientras que el volumen restante es desechado en los sistemas de alcantarillado municipal sin ningún tipo de tratamiento o bien es arrojado al medio ambiente. El desecho constante de este subproducto ha generado afectación en los cuerpos receptores de agua y terrenos de cultivo. ABSTRACT: Whey is a byproduct of the dairy industry in which the principal components are lactose, proteins and mineral. In Acatlán, Hidalgo, there are many dairy industries, the majority using rudimentary systems of production. This byproduct has a little commercial value for producers, a small part is used in cattle feed, while the remainder is discarded in the municipal sewer system without any treatment or it is thrown into the environment. The continuous discharge of whey onto water and land can produce serious pollution problems. Palabras clave: Lactosuero, lácteos, contaminación. INTRODUCCIÓN La industria alimenticia es considerada una de las más contaminantes a nivel mundial, esto debido a que produce residuales con altas cargas orgánicas, las cuales son de difícil degradación en el medio ambiente. El lactosuero es un subproducto que se genera en la industria láctea, sus principales componentes son la lactosa, proteínas y algunos minerales. Existen dos tipos de lactosuero, el lactosuero dulce y el lactosuero ácido, el primero se obtiene en la elaboración de lácteos en donde se utiliza la coagulación enzimática con un pH cercano a 5.6. El lactosuero ácido se produce cuando la coagulación es por acidificación a un pH de 5.1 o menor (Arrojo et al., 2003). Aproximadamente 47% de los 115 millones de toneladas de lactosuero producido anualmente a nivel mundial son desechados al medio ambiente. Esto representa una pérdida de una fuente de alimentación y causa serios problemas de contaminación, ya que el lactosuero contiene una alta cargo orgánica con valores de DBO de 40 000- 60 000 mg/L y DQO 50 000-80 000 mg/L. (Ben-Hassan and Ghaly, 1994; Fournier et al., 1993). Más del 90% de la DBO y DQO del lactosuero proviene de la lactosa
(Kisaalita et al., 1990; Malaspina et al., 1996). La composición típica de un lactosuero se observa en la tabla 1 (Saddoud et al., 2007).
PARÁMETRO MUESTRA DQO (g/L) 68.6 ± 3.3 DBO5(g/L) 37.71 ± 2.84 SST (g/L) 1.35 ± 0.06
Lactosa (g/L) 45.9 ± 0.88 Proteínas (g/L) 2.71 ± 0.05
pH 4.9 ± 0.27 NTK (g/L) 1.12 ± 0.01 FT(g/L) 0.5 ±1.8 x 10 -3
Tabla 1. Composición química del suero lácteo. Las industrias productoras de lácteos pequeñas, no pueden aprovechar las tecnologías de revalorización del lactosuero, como la recuperación de proteínas y lactosa, o el secado por aspersión, debido a sus altos costos. Los tratamientos fisicoquímicos o biológicos son los más viables para estas empresas (Gavala et al., 1999). Así, el problema de la disposición del lactosuero ha ido empeorando. La continua descarga de este sobproducto en la tierra pone en peligro la estructura física y química de los suelos, reduciendo los rendimientos de cultivos y provocando serios problemas de contaminación de agua subterránea (Ben-Hassan and Ghaly, 1994). En la zona lechera del municipio de Acatlán, Hidalgo, existe un gran número de industrias procesadoras de lácteos, en su mayoría utilizan sistemas rudimentarios para la producción de los mismos, los volúmenes de procesamiento de leche de bovino varía desde los 4000 hasta los 30 mil litros, aproximadamente. Más de la mitad de la leche que se procesa en las industrias es utilizada en la producción de queso tipo Oaxaca, mientras que el volumen restante se utiliza en la elaboración de crema y queso tipo: panela, manchego, botanero y tenate. El lactosuero representa cerca del 85 al 95% del volumen de leche usada en la trasformación de los productos lácteos. Este subproducto tiene poco valor comercial para los productores, una pequeña parte es utilizada en la alimentación del ganado, mientras que el volumen restante es desechado en los sistemas de alcantarillado municipal sin ningún tipo de tratamiento o bien es arrojado al medio ambiente, generándose consecuencias nocivas para los cuerpos de agua, los cuales son utilizados en los sistemas de riego, por lo que los terrenos de cultivo igualmente se ven afectados. En la figura 1 se muestran las localidades con mayor producción de lactosuero.

Figura 1. Localidades con mayor producción de suero. Fuente COEDE 2008.
El objetivo de este estudio es dar a conocer la afectación de un cuerpo de agua por la descarga de suero lácteo proveniente de diferentes industrias lácteas. METODOLOGÍA Área de estudio El área de estudio se encuentra localizada en el sur del municipio de Acatlán. Sus ubicaciones se encuentran en la tabla 2 y figura 2. La presa “El sabino” recibe los residuales de lactosuero provenientes de las localidades de: Los Migueles, Metepec primero, Metepec segundo, Chauténco y Agustín Olvera.
Tabla 2. Ubicación geográfica de los sitios de estudio.
LOCALIDAD COORDENADAS GEOGRÁFICAS
ALTITUD (m)
Presa el Sabino 20°09´08.33´´ N 98°23´57.37´´ O 2124 Los Migueles 20°09´43.93´´ N 98°24´23.82´´ O 2128 Metepec Primero 20°09´01.24´´ N 98°26´17.25´´ O 2168 Metepec Segundo 20°09´37.28´´ N 98°25´03.24´´ O 2129 Agustín Olvera 20°07´28.01´´ N 98°25´12.90´´ O 2156 Chauténco 20°08´26.60´´ N 98°26´22.02´´ O 2155

Figura 2. Mapa de ubicación de las áreas de estudio. Fuente INEGI 2009.
Obtención de datos Se realizo un recorrido de campo a través del cual se identificaron los puntos de descarga de lactosuero. El recorrido consistió en hacer una visita a la presa, se observo que un pequeño arroyo descarga lactosuero en la parte noroeste del cuerpo receptor de agua (Figura 3). Una vez identificado este punto se realizo un recorrido hacia el nacimiento del arroyo. De esta manera se identificaron los puntos de descarga (Figura 4). Una vez identificados estos puntos, se procedió a realizar una encuesta a empresas productoras de lácteos, en donde los datos principales a obtener fueron los volúmenes de lactosuero generados por cada empresa y la disposición de los mismos.

Figura 3. Descarga de suero en la presa el Sabino.
Figura 4. Descarga de suero en arroyo.
RESULTADOS Y DISCUSIÓN Se identificaron 11 industrias procesadoras de lácteos a lo largo de 6 km del arroyo. El volumen de leche procesada por estas industrias es de 145, 500 litros diarios. La leche que se procesa proviene del municipio de Acatlán, Actopan, Valle de Tulancingo, Valle del Mezquital, Tizayuca, Veracruz, Puebla, Jalisco, Querétaro, Aguascalientes y Guanajuato. El producto principal que se obtiene es el queso tipo Oaxaca, seguido de crema y requesón. En menores cantidades se obtiene queso tipo panela, botanero, morral, manchego y tenate. El 72.72% de las empresas encuestadas se encuentran registradas ante el Consejo Estatal de Ecología. El 90% labora los 7 días a la semana. Son industrias pequeñas, ya que en promedio generan 12 fuentes de empleo. En total se producen

aproximadamente 122,000 litros/día de lactosuero, el cual se genera después de la obtención del requesón, por lo que se considera un suero del tipo ácido, un 39.35% del volumen de lactosuero es utilizado en la alimentación del ganado mientras que el volumen restante es desechado al medio ambiente. CONCLUSIONES El lactosuero es un subproducto con alto valor nutricional, sin embargo en el municipio de Acatlán no es aprovechado adecuadamente debido a que las empresas generadoras son pequeñas industrias, las cuales no cuentan con los recursos necesarios para invertir en tecnologías de aprovechamiento de este subproducto. Debido a esta situación, el lactosuero es desechado al medio ambiente, ya que esto no genera ningún costo monetario. El desecho constante del mismo ha provocado la afectación en los cuerpos receptores de agua y terrenos de cultivo. El descontento social es un problema constante, ya que el lactosuero no solo afecta suelo y agua, sino que afecta al ambiente en general debido a los olores molestos que se generan cuando el subproducto se encuentra a cielo abierto. REFERENCIAS Arrojo B, Omil F, Garrido JM, Méndez R. 2003. Combinación de un filtro anaerobio y un sistema SBR para el tratamiento de las aguas generadas en un laboratorio de análisis de productos lácteos. Afinidad 60(506):344-354. Ben-Hassan RM, Ghaly AE. 1994. Continuous propagation of Kluyveromyces fragilis in cheese whey for pollution potential reduction. Appl. Biochem. Biotechnol. 47:89–105. COEDE. Consejo Estatal de Ecología en el estado de Hidalgo. Fournier D, Schwitzguebel JP, Peringer P. 1993. Effect of different heterogeneous inocula in acidogenic fermentation of whey permeate. Biotechnol. Lett. 15:627–632. Gavala HN, Skiadas IV, Lyberatos G. 1999. On the performance of a centralised digestion facility receiving seasonal agroindustrial wastewaters. Water Sci. Technol. 40:339–346. Instituto Nacional de Estadistica y Geografía. (online). Disponible en: http://mapserver.inegi.org.mx/map/visortoDx/visor.html. Kisaalita WS, Lo KV, Pinder K L. 1990. Influence of whey protein on continuous acidogenic degradation of lactose. Biotechnol. Bioeng. 36:642–645. Malaspina F, Cellamare CM, Tilche A. 1996. Anaerobic treatment of cheese-whey with a downflow-upflow hybrid reactor. Bioresource Technology 55:131–139. Saddoud A, Hassaïri I, Sami S. 2007. Anaerobic membrane reactor with phase separation for the treatment of cheese whey. Bioresource Technology 98:2102-2108.

RELACIÓN ENTRE LA VIABILIDAD DE BACTERIAS ÁCIDO LÁCTICAS ENCAPSULADAS Y LA TEXTURA DE DIFERENTES HIDROCOLOIDES
UTILIZADOS COMO MATERIAL ENCAPSULANTE
Ariza, T. J.a,*, Pérez-Chabela M. L.b y Totosaus, A.a
a Laboratorio de Alimentos, Tecnológico de Estudios Superiores de Ecatepec, Av. Tecnológico s/n, Col. Valle de Anáhuac, Ecatepec, Estado de México, C. P. 55210.
b Departamento de Biotecnología, Universidad Autónoma Metropolitana, Iztapalapa, Av.
San Rafael Atlixco No. 186, Col. Vicentina, México D.F., C. P. 09340.
* [email protected] RESUMEN: El objetivo de este trabajo fue evaluar la viabilidad de cuatro bacterias ácido lácticas (Enterococcus faecium, Lactobacillus plantarum, Aerococcus viridans y Pediococcus pentosaceus) encapsuladas con tres mezclas de hidrocoloides: Alginato de Sodio (AGL), Alginato/Gelana (AGL) y Alginato/κ-Carragenina/Algarrobo (AKL) y su efecto sobre la textura. Se determinaron los parámetros dureza, cohesividad, resorteo y resiliencia (vía un Análisis del Perfil de Textura TPA), y la ductilidad y esfuerzo aparente (prueba de Penetración) para las tres formulaciones de gel. La viabilidad de las cepas se determinó por conteo en placa y por microscopía de fluorescencia mediante los tintes SYTO9 y Yoduro de Propidio. De acuerdo al TPA en los parámetros de cohesividad, resorteo y elasticidad aparente entre las tres formulaciones de geles no hubo diferencia significativa (p<0.05). El gel de ALG fue el más duro, mientras que la mezcla con otro hidrocoloide decremento la dureza del gel. El gel de AGL fue el más dúctil y resiliente de los tres. Las cepas pierden viabilidad en ALG como material encapsulante, sin embargo, conservan su viabilidad hasta después de un mes de elaboradas cuando son encapsuladas en AKL, mismo comportamiento que se observó en la microscopía de fluorescencia. ABSTRACT: The objective of this work was to evaluate the existing relation between the viability of lactic acid bacteria (E. faecium, L. plantarum, A. viridams y P. pentosaceus) and the textural properties of the three gelling hydrocolloids Sodium Alginate (ALG), Alginate/Gellan (AGL) and Alginate/κ-Carrageenan/Locust Bean Gum (AKL) used as encapsulating materials. Hardness, cohesiveness, springiness and resilience (through a Textural Profile Analysis, TPA), and ductility and apparent elasticity (Penetration test) were determined in the three formulations. Plating count technique was used in order to determine strain viability, also fluorescent microscopy was used with an appropriate mixture of the SYTO9 and Propidium iodide stains. Parameters of cohesiveness, springiness and apparent elasticity were not significantly different (p<0.05) between the three formulations, according to the TPA. The hardest gel was formed with ALG, in contrast a gelling hydrocolloids mixture decreases gel hardness. The AGL gel was the most ductile and resilient. Strain viability was affected using ALG as coating material; nevertheless, viability is preserved when AKL is used. The same behaviour was observed by Fluorescent Microscopy. Palabras clave: Microencapsulación, hidrocoloides gelificantes, viabilidad de BAL.

INTRODUCCIÓN Las bacterias acido lácticas (BAL) son un grupo de microorganismos que por lo general se asocian con el tracto gastrointestinal humano; se ha reportado que éstas pueden reprimir el crecimiento de patógenos y estabilizar el sistema digestivo (Kim et al., 2008); así como reducir el colesterol, incrementar la tolerancia a la lactosa y prevenir el riesgo de contraer cáncer (O’Riordan et al., 2001). Algunas de estas bacterias poseen características que les proporcionan un efecto probiótico, sin embargo su viabilidad se puede ver afectada debido a las características adversas del tracto gastrointestinal. La definición del término probiótico con el paso del tiempo ha sido modificado en varias ocasiones, actualmente se ha definido el término probiótico como los “microorganismos viables que llegan al intestino en cantidades suficientes como para ejercer un efecto benéfico en el organismo” (Vrese and Schrezenmeir, 2008); según la Federación Internacional de Lácteos se recomiendan niveles de 107 UFC/g para productos con este tipo de bacterias (Ouwehand and Salminen, 1998). Para que una bacteria sea considerada un probiótico debe presentar actividad probiótica documentada, ser tolerante al ácido láctico, no perder viabilidad durante el almacenamiento, sobrevivir el paso a través de la parte superior del tracto intestinal y ser resistente a los ácidos biliares (Knorr, 1998). Como una manera de proteger a las BAL se han realizado numerosos estudios en los cuales se provee a las bacterias de un recubrimiento inocuo conocido como matriz o membrana con lo que se pretende reducir la pérdida de células viables, proceso que se conoce como: Microencapsulación (Pimentel et al., 2009). Diferentes materiales se han utilizado con esta finalidad como son algunos carbohidratos, gomas, lípidos, celulosas y proteínas. Las gomas, entre las que se encuentran el alginato, almidón, κ-carragenina, algarrobo, xantana, gelana y gelatina han demostrado ser capaces de retener células viables, protegerlas de condiciones adversas e incrementar su supervivencia; así como diversos métodos de encapsulación entre los que se encuentran secado por aspersión, emulsificación, extrusión, coacervación, inclusión, centrífugación, suspensión y liofilización (Muthukumarasamy et al., 2006). El alginato es uno de los materiales más utilizados en la encapsulación de microorganismos. Este compuesto ha mostrado la capacidad de incrementar la supervivencia de bacterias probióticas en un rango entre 80-90% (Mandal et al., 2006). Por lo que, el objetivo de este trabajo fue evaluar la relación existente entre la viabilidad de las bacterias ácido lácticas termorresistentes: Enterococcus faecium, Lactobacillus plantarum, Aerococcus viridams y Pediococcus pentosaceus; y las propiedades de tres mezclas de hidrocoloides gelificados: AGL (Alginato de Sodio), AGL (Alginato/Gelana) y AKL (Alginato/κ-Carragenina/Algarrobo) utilizados como materiales encapsulantes.
METODOLOGÍA Preparación de los geles Se utilizaron tres formulaciones de mezclas de hidrocoloides para formar los geles (Cuadro 1). Los componentes fueron disueltos en agua destilada, manteniendo agitación constante y llevando la solución hasta una temperatura máxima de 60°C. Una vez realizada la mezcla se moldearon las soluciones y para la formación final del gel se agregó 2% de CH3COOH.
Cuadro 1. Formulaciones utilizadas en la elaboración de los geles. Mezcla de hidrocoloides ALG AGL AKL
Células centrifugadas (%) 0.05 0.05 0.05 Gelana (%) - 0.1 -
KCl (%) - - 0.1 κ-Carragenina (%) - - 0.3
Algarrobo (%) - - 0.3 Alginato (%) 1.0 1.0 1.0 CaCO3 (%) 0.4 0.4 0.4
Análisis de Perfil de Textura (TPA) Los geles se moldearon en una funda sintética de 20mm de diámetro y se rebanaron cada 20mm de longitud. El TPA fue realizado en un Analizador de Textura TAXT2i (Texture Technologies Corp.). Los parámetros determinados a partir del TPA fueron dureza, cohesividad, resorteo y resiliencia (Pons and Fiszman, 1996). Prueba por penetración Para medir la resistencia del gel a la fractura por punción, los geles se moldearon dentro de tubos de ensaye y fueron penetrados por un vástago de 8mm de diámetro. Esta prueba se realizó en un Analizador de Textura TAXT2i (Texture Technologies Corp.) obteniéndose parámetros de Ductilidad y Elasticidad Aparente (Hickson et al., 1982). Cepas bacterianas Las bacterias ácido lácticas (BAL) utilizadas fueron: Enterococcus faecium, Lactobacillus plantarum, Aerococcus viridans y Pediococcus pentosaceus las cuales en trabajos previos se ha comprobado su potencial probiótico. Encapsulación de las BAL Se utilizó la técnica de emulsión que consiste en preparar una solución acuosa de las mezclas de hidrocoloides (Cuadro 1), mientras que por separado se prepara la fase oleosa con 200mL de Aceite de Vegetal. Con agitación constante, se vertió la
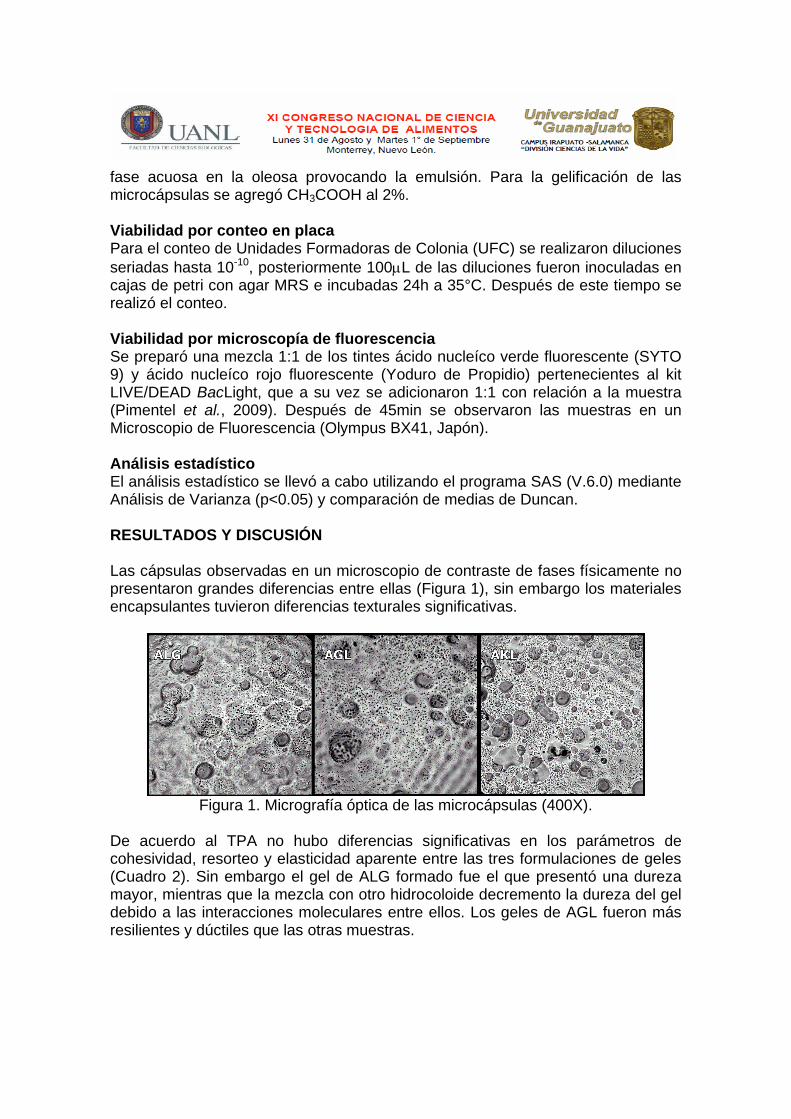
fase acuosa en la oleosa provocando la emulsión. Para la gelificación de las microcápsulas se agregó CH3COOH al 2%. Viabilidad por conteo en placa Para el conteo de Unidades Formadoras de Colonia (UFC) se realizaron diluciones seriadas hasta 10-10, posteriormente 100µL de las diluciones fueron inoculadas en cajas de petri con agar MRS e incubadas 24h a 35°C. Después de este tiempo se realizó el conteo. Viabilidad por microscopía de fluorescencia Se preparó una mezcla 1:1 de los tintes ácido nucleíco verde fluorescente (SYTO 9) y ácido nucleíco rojo fluorescente (Yoduro de Propidio) pertenecientes al kit LIVE/DEAD BacLight, que a su vez se adicionaron 1:1 con relación a la muestra (Pimentel et al., 2009). Después de 45min se observaron las muestras en un Microscopio de Fluorescencia (Olympus BX41, Japón). Análisis estadístico El análisis estadístico se llevó a cabo utilizando el programa SAS (V.6.0) mediante Análisis de Varianza (p<0.05) y comparación de medias de Duncan. RESULTADOS Y DISCUSIÓN Las cápsulas observadas en un microscopio de contraste de fases físicamente no presentaron grandes diferencias entre ellas (Figura 1), sin embargo los materiales encapsulantes tuvieron diferencias texturales significativas.
Figura 1. Micrografía óptica de las microcápsulas (400X).
De acuerdo al TPA no hubo diferencias significativas en los parámetros de cohesividad, resorteo y elasticidad aparente entre las tres formulaciones de geles (Cuadro 2). Sin embargo el gel de ALG formado fue el que presentó una dureza mayor, mientras que la mezcla con otro hidrocoloide decremento la dureza del gel debido a las interacciones moleculares entre ellos. Los geles de AGL fueron más resilientes y dúctiles que las otras muestras.

Cuadro 2. Parámetros texturales de los geles utilizados. PARÁMETRO ALG AGL AKL
Dureza (N) 3.91±0.92AB 4.77±2.01A 3.00±0.05B Cohesividad 3.00±0.12A 2.89±0.44A 2.86±0.07A
Resorteo (mm) 1.00±0.00A 1.03±0.05A 1.03±0.04A Resiliencia 0.11±0.01B 0.12±0.02A 0.07±0.00C Ductilidad 0.47±0.01B 0.48±0.01A 0.48±0.01AB
Elasticidad aparente (N/cm2) 2.64±1.08A 2.85±1.87A 5.53±2.96A A,B,C Medias con misma letra en el mismo renglón no son significativamente diferentes (P<0.05)
Las cepas de A. viridans y P. pentosaceus presentaron una mayor viabilidad que E. faecium y L. plantarum. En cuanto al material encapsulante las BAL encapsuladas en AGL y AKL presentaron una viabilidad aceptable aún después de un mes de almacenamiento a 4°C, siendo ALG el material que redujo la viabilidad de las cepas hasta en 5 ciclos logarítmicos.
Figura 2. Viabilidad en placa de las BAL encapsuladas.
La viabilidad por microscopía de fluorescencia confirma los resultados obtenidos en placa, siendo el ALG un material que reduce la viabilidad de las células y AKL el material que mejor la conserva (Figura 3).

Figura 3. Microfotografía de fluorescencia (400X).
CONCLUSIONES Los materiales que forman la matriz de las microcápsulas tienen un marcado efecto sobre la viabilidad de las BAL debido a sus diferentes propiedades texturales resultado de la interacción de los hidrocoloides que las forman, en especial la formulación de AKL. A. viridans y P. pentosaceus fueron las cepas encapsuladas con mayor viabilidad en placa. REFERENCIAS Hickson D, Dill C, Morgan R, Sweat V, Suter V and Carpenter Z. 1982. Rheological
properties of two heat-induced proteins gel. Journal of Food Science. 47:783-791. Kim SJ, Cho S, Kim SH, Song O, Shin I, Cha D and Park H. 2008. Effect of
microencapsulation on viability and other characteristics in Lactobacillus acidophilus ATCC 43121. LWT. 41:493-500.
Knorr D. 1998. Technology aspects related to microorganisms in functional foods. Trends in Food Science & Technology. 9:295-306.
Mandal S, Puniya AK and Singh K. 2006. Effect of alginate concentrations on survival of microencapsulated Lactobacillus casei NCDC-298. International Dairy Journal. 16:1190-1195.
Muthukumarasamy P, Allan P and Holley R. 2006. Stability of Lactobacillus reuteri in Different Types of Microcapsules. Journal of Food Science. 71(1):20-24.
O’Riordan K, Andrews D, Buckle K and Conway P. 2001. Evaluation of microencapsulation of a Bifidobacterium strain with starch as an approach to prolonging viability during storage. Journal of Applied Microbiology. 91:1059-1066.
Ouwehand AC and Salminen SJ. 1998. The health effects of cultured milk products with viable and non-viable bacteria. International Dairy Journal. 8(9):749-758.
Pimentel DJ, Campos RG, Lobato C, Pedroza R y Vernon EJ. Encapsulation of Lactobacillus rhamnosus in double emulsions formulated with sweet whey as emulsifier and survival in simulated gastrointestinal conditions. Food Research International. 42:292-297.
Pons M and Fiszman S. 1996. Instrumental Texture Profile Analysis with particular reference to gelled systems. Journal of Texture Studies. 27:597-624.
Vrese M and Schrezenmeir J. 2008. Probiotics, Prebiotics, and Synbiotics. Adv Biochem Engin/Biotechnol. 111:1-66.

MARCADOR BIOLOGICO DE LA CALIDAD DE LA CARNE BOVINA: PURIFICACION, CARACTERIZACION Y LOCALIZACION CELULAR DE LA
ANTITROMBINA III
Herrera Méndez C. H.a,*, Ruiz Aguilar G. M. de la L.a, Arroyo Figueroa G.a, Saavedra Medina L. F.a, Trujillo Santoyo A. D.a
a Departamento de Ingeniería Agroindustrial, División de Ciencias de la Salud e Ingenierías, Campus Celaya-Salvatierra, Universidad de Guanajuato, Privada de Arteaga
s/n, Colonia Centro, C.P. 38900, Salvatierra, Guanajuato, México.
* [email protected]; [email protected]. RESUMEN: En el presente estudio, la Antitrombina III (AT-III) fue purificada de músculo esquelético de bovino, utilizando cuatro etapas cromatográficas incluyendo sucesivamente Sephadex G100, Q-Sepharosa y EMD-DEAE 650. La identidad de la serpina purificada fue confirmada utilizando diferentes técnicas incluyendo secuenciación N-Terminal, western-blot revelado con un anticuerpo policlonal dirigido contra la AT-III humana y con el mapa peptídico por espectrometría de masas Maldi-tof. Todos los resultados demostraron que la serpina purificada del músculo es sin duda la antitrombina-III. De acuerdo a las velocidades de asociación determinadas experimentalmente, la AT-III muscular es un potente inhibidor de la trombina, una actividad inhibitoria que mostró ser dependiente de la presencia (kass = 2.3 x107 M−1 · s−1) o ausencia (kass = 1.8 x105 M−1 · s−1) de heparina. La tripsina (kass = 5 x105 M−1 · s−1), quimotripsina (kass = 6.8 x104 M−1 · s−1) y plasmina (kass = 1.7 x104 M−1 · s−1) también son inhibidas por la AT-III muscular. Asimismo fue demostrado por titulación que una molécula de trombina o tripsina interactúa con una molécula de AT-III (SI=1). Investigaciones inmunohistoquímicas demostraron que la AT-III es esencialmente intracelular en todos los músculos de bovino estudiados: Diaphragma pedialis, Rectus abdominis y Transversus abdominis. ABSTRACT: In the present study, antithrombin III (AT-III) was purified from skeletal bovine muscle using a four-step chromatography procedure including successively Sephadex G100, Q-Sepharose and EMD-DEAE 650. Identity of the purified serpin was assessed using different approaches including N-terminal sequencing, western blot revealed with a polyclonal antibody raised against human AT-III and Maldi-tof mass spectrometry peptide map. All results demonstrated that the purified muscle serpin is undoubtedly antithrombin III. According to the experimental association rates determined, muscle AT-III is a potent inhibitor of thrombin, an inhibitory activity shown to be dependent on the presence (kass = 2.3 x107 M−1 · s−1) or absence (kass = 1.8 x105 M−1 · s−1) of heparin. Trypsin (kass = 5 x105 M−1 · s−1), chymotrypsin (kass = 6.8 x104 M−1 · s−1) and plasmin (kass = 1.7 x104 M−1 · s−1) are also inhibited by muscle AT-III. As demonstrated by titration, one molecule of thrombin or trypsin interact with one molecule of AT-III (SI=1). Immunohistochemical investigations demonstrate that AT-III is essentially intracellular in all bovine muscles studied: Diaphragma pedialis, Rectus abdominis and Transversus abdominis muscles.
Carne, Terneza, Serpina

INTRODUCCIÓN La terneza es el atributo de calidad más importante de la carne. Entre diferentes variables de orden biológico y fisicoquímico, in vivo los inhibidores de serina peptidasas (SERPINAS) como la antitrombina III (AT-III), han sido los mejores indicadores de la terneza de la carne de bovino (Zamora et al., 2005). En la sangre, la actividad de la trombina está controlada por la AT-III, un inhibidor especifico. La trombina extravascular está regulada, y la AT-III es un candidato potencial. La enzima en las células musculares se localiza en la parte externa de la membrana del plasma en la unión neuromuscular y su regulación es en dicha unión (Liu et al., 1994). En bovino, su presencia es desconocida. La AT-III es hallada en fibras musculares de ratón sugiriendo que esta serpina se expresa en células musculares (Businaro et al., 1995). A pesar que la AT-III debe estar expresada en el músculo esquelético de bovino, esta proteína nunca ha sido purificada y caracterizada en dicho tejido. El objetivo del presente trabajo es purificar y caracterizar la AT-III del músculo esquelético de bovino; marcador biológico de la calidad de la carne. METODOLOGÍA La purificación de la AT-III fue realizada de un extracto de músculo Diafragma pedialis de bovino. El proceso implica cinco etapas cromatográficas. Se evaluó: la actividad inhibitoria contra la tripsina en las fracciones recolectadas, la titración de dicha enzima y la estequiometría de la interacción enzima/inhibidor. Se midió las constantes de asociación y electroforesis en gel de poliacrilamida fue realizada. La estabilidad al pH del inhibidor purificado fue analizada en un rango de pH (2-12). La estabilidad a la temperatura fue determinada entre 40-100 °C. La secuencia N-Terminal de la AT-III fue realizada con un secuenciador Applied Biosystems 477A. El análisis de espectrometría de masas MALDI-TOF fue realizado con la tripsina, enzima de digestión. La inmunolocalización de la AT-III fue efectuada en cortes transversales de varios músculos utilizando un anticuerpo policlonal dirigido contra la AT-III humana. RESULTADOS Y DISCUSIÓN La identidad de la serpina purificada fue establecida por diferentes métodos analíticos. El análisis SDS-PAGE de la AT-III purificada mostró una banda con un peso molecular estimado de 58 kDa (Fig. 1, línea b). La especificidad del anticuerpo policlonal dirigido contra la AT-III humana fue analizada con extracto crudo de músculo y de serpina purificada. Como se muestra en la Fig. 2a, después del SDS-PAGE, el anticuerpo revela una banda en el extracto bruto (línea 1) migrando similarmente la AT-III purificada (línea 2). En condiciones no desnaturalizantes, el anticuerpo reconoce una banda en el extracto crudo (Fig. 2b, línea 1) migrado como la AT-III purificada (Fig. 2b, línea 2). Estos descubrimientos sugieren una especificidad alta del anticuerpo, el cual fue utilizado en la localización de la AT-III en el tejido muscular. La espectrometría de masas identifica esta proteína como la AT-III.

Figura 1. El inhibidor fue migrado en condiciones desnaturalizantes en un gel de
poliacrilamida a 12.5% y revelado por coloración en plata (a) Marcadores de peso molecular, (b) AT-III purificada
Figura 2. Análisis por Wester-blot con el anticuerpo policlonal contra la AT-III
humana. (2a, línea 1): extracto bruto; (2a, línea 2): AT-III purificada. En condiciones no desnaturalizanrtes (2b, línea 1): extracto bruto; (2b, línea 2): AT-III purificada
El tratamiento de la AT-III por 15 min. a temperaturas entre 40 y 100 °C confirma la baja estabilidad térmica de la AT-III bovina (Fig. 3a). La AT-III bovina es relativamente estable solo a pH 10 (Fig. 3b). Figura 3. Actividad de la AT-III (a) calentada 15 min. a temperaturas entre
40-100 °C. (b) preincubada por 1h a diferentes pH entre 2-12.

El patrón de actividad inhibitoria de la AT-III de músculo y las constantes de velocidad de asociación (Tabla 1) indican que todas las peptidasas son sensibles a la acción inhibitoria de esta serpina.
Tabla 1. Constantes de velocidad de asociación (kass) para serina peptidasas
La AT-III fue inmunolocalizada entre las membranas del plasma y las miofibrillas donde está altamente concentrada, sin embargo existe una intensidad baja de fluorescencia dentro de las miofibrillas en el centro de las fibras musculares indicando que la AT-III del músculo es intracelular pero con una localización periférica preferencial (Fig. 4B). No fue detectada fluorescencia en la muestra de control en la cual el anticuerpo primario fue omitido (Fig. 4A). Figura 4. Inmunolocalización de la AT-III muscular en el músculo Diafragma
pedialis de bovino. La inmunolocalización fue realizada utilizando un antisuero de conejo diluido 1/50 revelado con IgG de cabra contra conejo marcado con FITC (isotiocianato fluorescente) en una dilución 1/500. (A) Muestra control incubada solo con el anticuerpo secundario. (B) Muestra tratada con los anticuerpos primario y secundario (aumento x 1200).
Investigaciones han indicado que la trombina es un regulador importante del desarrollo del músculo (Suidan et al., 1996; Citron et al., 1997; Glazner et al.,
Peptidasa kass(M-1, s-1)
Tripsina 5 x 105
Quimotripsina 6.8 x 104
Plasmina 1.7 x 104
Trombina 1.8 x 105

1997; De Niese et al., 2002). Muchas respuestas celulares son mediadas por la trombina (De Niese et al., 2002). En el músculo, esta serina peptidasa ejerce asimismo un número de acciones claves en su desarrollo y reparación. Por lo tanto su actividad debe estar estrictamente regulada. Una serie de investigaciones realizadas con el objetivo de entender mejor las funciones de la trombina en el músculo esquelético fueron basadas en el uso de inhibidores de trombina. Sin embargo, la AT-III nunca ha sido utilizada, sugiriendo que este inhibidor no se expresa en las células musculares. Businaro et al. (1995) mostró que la AT-III está presente dentro de todas las fibras musculares del músculo esquelético de ratón, un descubrimiento que apoya la expresión de esta proteína en células musculares. Experimentos preliminares y apoyando estos descubrimientos, la presencia de la AT-III fue detectada por western-blot en extractos crudos de músculo esquelético. Por lo que nosotros decidimos purificar y caracterizar AT-III del músculo esquelético de bovino. CONCLUSIONES La AT-III nunca antes había sido purificada del tejido muscular y esta es la primera vez que un procedimiento de purificación es propuesto. El presente trabajo debe ser considerado como una nueva vía de información en los estudios sobre el rol de relación trombina/antitrombina en el desarrollo del músculo y su influencia en la terneza de la carne, la característica organoléptica más buscada por los consumidores. REFERENCIAS Businaro R., Nori SL, Toesca A, De Renzis G, Ortolani F, De Santis E and
Fumagalli L. 1995. Immunohistochemical detection of three serum protease inhibitors in mouse skeletal muscle by confocal laser scanning microscopy. Ital. J. Anat Embryol. 100 Suppl 1, 123-130.
Citron BA, Smirnova IV, Zoubine MN, and Festoff BW. 1997. Quantitative PCR
analysis reveals novel expression of prothrombin mRNA and regulation of its level in developing mouse muscle. Thromb. Res., 87, 303-313.
De Niese MR, Chinni C, Pike RN, Bottomley SP, Mackie EJ. 2002. Dissection of
protease-activated receptor-1-dependent and -independent responses to thrombin in skeletal myoblasts. Exp Cell Res. 274, 149-156.
Glazner GW, Yadav K, Fitzgerald S, Coven E, Brenneman DE and Nelson PG.
1997. Cholinergic stimulation increases thrombin activity and gene expression in cultured mouse muscle. Dev. Brain Res. 99, 148–154.
Liu Y, Fields RD, Festoff BW and Nelson, PG. 1994. Proteolytic action of thrombin
is required for electrical activity-dependent synapse reduction. Proc. Natl. Acad. Sci. U. S. A. 91, 10300–10304.

Suidan HS, Niclou SP, Dreessen J, Beltraminelli N and Monard D. 1996. The thrombin receptor is present in myoblasts and its expression is repressed upon fusion. J. Biol. Chem. 271, 29162–29169.
Zamora F. Aubry L, Sayd T, Lepetit J, Lebert A, Sentandreu M and Ouali A. 2005.
Serine peptidase inhibitors, the best predictor of beef ageing amongst a large set of quantitative variables. Meat Sci., ,71, 730-742.

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
POTENCIAL DE PRODUCCIÓN DE PROPOLEO EN UN APIARIO UBICADO EN EL MUNICIPIO DE IRAPUATO, GTO.
Bucio Villalobos C.M. a,*, Estrada Ibarra J.L. a, Martínez Jaime O.A. b, Torres Morales J.J. b
a Departamento de Agronomía, División Ciencias de la Vida, Universidad de Guanajuato,
Ex–Hacienda El Copal, Km. 9 Carretera Irapuato-Silao, C.P. 36821, Irapuato, Gto., México
b Universidad de Guanajuato, Irapuato, Gto. México
* [email protected] RESUMEN: La miel es el principal producto obtenido en una explotación apícola, sin embargo el propóleo también puede ser cosechado. Para conocer la potencialidad de la zona de Guanajuato, se realizó el presente trabajo con el objetivo de evaluar la producción de propóleo cosechado con la técnica de raspado, a lo largo de un año en un apiario ubicado en el municipio de Irapuato, Guanajuato. El propóleo fue cosechado cada 14 días durante 12 meses consecutivos, raspándose donde éste era localizado en la tapa, el alza colocada inmediatamente debajo de ella y la cámara de cría. La producción por colmena varió desde 87.2 hasta 1.6 gr de propóleo total al año, siendo el promedio general de 35.0, obteniéndose la producción más alta de agosto a octubre, época de lluvias en que abundan las plantas que pueden proporcionar el propóleo a las abejas, y continuando (aunque con un 36.7 % de menor producción) durante noviembre a enero, con suficiente producción para que las abejas propolizaran aberturas en sus colmenas para protegerse del frío. Durante los meses de febrero a julio se tuvieron las menores producciones. La producción general obtenida fue considerada baja, por lo que se sugiere el uso de trampas especiales. ABSTRACT: Honey is the main product obtained from apiary nevertheless propolis also can be harvested. In order to know the potentiality of Guanajuato region, present research was realized with objective of evaluating the production of propolis harvested with the scraped technique, throughout a year in an apiary located in Irapuato, Guanajuato. Propolis was harvested every 14 days during 12 months consecutive. Propolis was scraping from cover, honey box and camera when it was located. The production by beehive varied from 87.2 to 1.6 gr of total propolis by year, being a general average of 35.0, obtaining the highest production from august to october, during of rain season in which the plants abound that can provide propolis to the bees, and continuing (although with a 36.7 % of smaller production) from november to January, with sufficient production so that the bees close with propolis the openings in their beehives to protect of cold. From february to july the smaller productions were had. Propolis production obtained was considered low, reason why the use of special traps is suggested. Palabras clave: Abejas, propóleo, cosecha en campo. INTRODUCCIÓN Aún con la explotación moderna de las abejas, la actividad apícola en México, presenta poca diversificación en los productos cosechados, ya que sólo se limita a la recolección de miel como producto primario, sin considerar la posibilidad de

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
obtención de otros productos (Quijano et al., 2006). Es posible obtener de una explotación apícola productos como jalea real, cera, polen, veneno, propóleo, abejas reina, núcleos de abejas y extracto de larva (Gris, 2004). De entre ellos, el propóleo ha sido objeto de intensas investigaciones químicas y farmacológicas en los últimos 40 años, las cuales han probado su benéfica actividad biológica (Rosado et al., 2006). Los principales países productores de propóleo son China, Brasil, Argentina, Cuba, Chile, Uruguay y Canadá, mientras que los países que principalmente captan dicha producción son Dinamarca, Francia, Alemania, Hungría, Ucrania y Estados Unidos de Norteamérica (Martínez et al., 2004). En México, la zona sureste comprendida por los estados de Tabasco, Veracruz, Campeche, Yucatán y Quintana Roo, cuenta con 691,000 colmenas en explotación, produciendo 27,875 toneladas de miel, 1,326 toneladas de cera, 7.5 toneladas de jalea real, 13 toneladas de polen y 6 toneladas de propóleo por año (Centro de Estudios Agropecuarios, 2001); para el resto de las zonas mexicanas donde se explotan abejas, no existen estadísticas salvo las referentes a la producción de miel, siendo el caso del estado de Guanajuato donde se carece de información sobre la potencialidad de la zona para la producción de propóleo de abeja. En base a esto, el objetivo del presente trabajo fue evaluar la producción de propóleo cosechado con la técnica de raspado, a lo largo de un año en un apiario ubicado en el municipio de Irapuato, Guanajuato. METODOLOGÍA La recolección de propóleo se realizó en 12 de las colmenas de un apiario ubicado en la comunidad de “El Copal” del Municipio de Irapuato, Gto., utilizando para ello la técnica de raspado. El propóleo fue cosechado cada 14 días durante 12 meses consecutivos, raspándose donde éste era localizado en la tapa, el alza colocada inmediatamente debajo de ella y la cámara de cría. La primera cosecha fue descartada y sirvió para estandarizar las colmenas. Una vez cosechado, el propóleo fue colocado en pequeñas charolas y almacenado bajo congelación hasta su pesado. RESULTADOS Y DISCUSIÓN Se observaron diferencias estadísticas en la producción de propóleo entre las diferentes colmenas, cuyos resultados se ilustran en la Figura 1. La producción por colmena varió desde 87.2 hasta 1.6 gr de propóleo total al año, siendo el promedio general de 35.0 gr, el cual resultó menor que el obtenido por Ramírez (1992) de 111.6 gr. Diversas condiciones pueden afectar la producción de propóleo, como son la conducta propolizadora de las abejas, el vigor de la colonia, la disponibilidad de plantas con resinas, entre otras (Martínez et al., 2006). Durante un año completo se hicieron colectas catorcenales de propóleo, dando un total de 27 fechas, cuya información es mostrada en la Figura 2. Se observaron dos épocas distintivas, la primera (de baja producción, menor de 1 gr) fue de febrero a julio, mientras que la segunda (de alta producción, mayor de 1 gr) fue de agosto a enero. La producción más alta de propóleo coincidió con la época de lluvias (agosto a octubre), época en que abundan las plantas que pueden proporcionar el propóleo a las abejas, pero continuó alta (aunque con un 36.7 % menor a los meses de agosto a octubre) durante los meses más fríos (noviembre a enero), con suficiente producción para que las abejas propolizaran las aberturas

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
en sus colmenas protegiéndose con ello del frío. Durante los meses de febrero a julio se tuvieron las menores producciones de propóleo, meses en que las temperaturas fueron más altas y estuvieron fuera del período de lluvias.
0
20
40
60
80
100
1 2 3 4 5 6 7 8 9 10 11
gr.
Figura 1. Producción total anual de propóleo por colmena.
0
0.5
1
1.5
2
2.5
3
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27
Fecha
Prod
ucci
ón p
rom
edio
(g)
Figura 2. Producción catorcenal de propóleo (del 1° de abril para la fecha 1, hasta
el 31 de marzo para la fecha 27).

XI CONGRESO NACIONAL DE CIENCIA Y TECNOLOGIA DE ALIMENTOS
Lunes 31 de Agosto y el Martes 1o de Septiembre Monterrey, Nuevo León.
COMITÉ ORGANIZADOR: Dra. Maria Guadalupe Alanís Guzmán, Presidente General Dr. Ernesto Alfredo Camarena Aguilar, Secretario General Dr. Mayela Bautista Justo Comité de Organización, UG M en C Carlos Leonel García Díaz, Comité de Organización, UANL
CAMPUS IRAPUATO -SALAMANCA
“DIVISIÓN CIENCIAS DE LA VIDA”
CONCLUSIONES La producción de propóleo obtenida durante el período y lugar de este estudio fue baja, lo que sugiere que la zona no es altamente productora. Sin embargo se sugiere evaluar trampas especiales para incrementar la producción, las cuales podrían principalmente ser colocadas sobre las colmenas durante los meses de agosto a enero. REFERENCIAS Centro de Estudios Agropecuarios. 2001. Apicultura. Ed. Grupo Iberoamericana: México, 108p. Gris VAG. 2004. El polen y la cera, alternativas de producción en la apicultura. Revista Imagen Veterinaria 4:31-40. Martínez PLR, Delgado HMA, Rojas HN y Casillas PR. 2004. El propóleo y las técnicas para su colecta. Memorias del XVIII Seminario Americano de Apicultura. Villahermosa, Tab. México. Quijano CE, Delgado HM, Mena RG y Pérez MI. 2006. Obtención del aceite esencial del propóleo que se produce en el sureste de México. Memorias del XX Seminario Americano de Apicultura. Querétaro, Qro. México. Ramírez VR. 1992. Evaluación de la producción de propóleo en un apiario de San Lorenzo Atemoaya, Xochimilco, DF. Tesis de licenciatura. Facultad de Medicina Veterinaria y Zootecnia. Universidad Nacional Autónoma de México. p23. Rosado PA, Martínez PLR, Rojas HN, Cuesta RO, Campo FM, y Montes de Oca PR. 2006. Características químicas de propóleos del estado de Campeche, México. Memorias del XX Seminario Americano de apicultura. Querétaro, Qro. México.


















