Digestion de Carbohidratos - … Discutir las bases bioquímicas de la digestión y absorción de...
101
Digestion de Carbohidratos
-
Upload
duongthuan -
Category
Documents
-
view
218 -
download
3
Transcript of Digestion de Carbohidratos - … Discutir las bases bioquímicas de la digestión y absorción de...

Digestion de
Carbohidratos

Objetivos:
Discutir las bases bioquímicas de la digestión y absorción de los hidratos de carbono
Describir las etapas y secuencia de reacciones de las vías metabólicas de los carbohidratos.
Explicar los mecanismos de regulación hormonal del metabolismo de los hidratos de carbono
Reconocer alteraciones bioquímicas básicas del metabolismo de los hidratos de carbono

Resultado de Aprendizaje:
Identificar los distintos procesos del metabolismo de los hidratos de carbono en condiciones normales y
anormales

Metabolismo
de los hidratos
de carbono

DIGESTION Y ABSORCION DE CARBOHIDRATOS
La mayor parte de los CHO se encuentran naturalmente en forma de:
almidón lactosa
glucógeno glucosa
celulosa fructosa
En la dieta occidental el 60% de las calorías totales derivan de los CHO, antes que estos sean completamente digeridos sus compuestos tienen que degradarse hasta monosacáridos.
Las enzimas que los degradan son glucosidasas o
glucogenasas.

Funciones de la Digestión de CH
Degradar CH complejos hasta CH simples.
Permite la absorción de los productos resultantes.
Comienza en la boca por acción de la amilasa salival.
Continúan hacia el intestino delgado, donde se distiende la pared duodenal, estimula la secreción de 2 Hh:
colecistoquinina
la secretina.
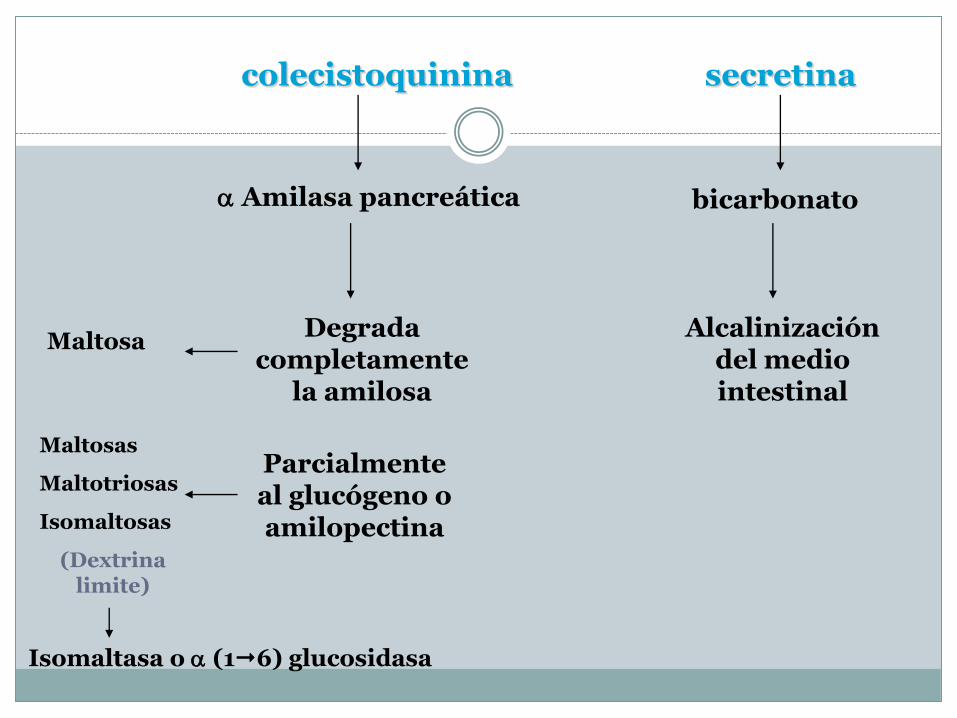
colecistoquinina secretina
Amilasa pancreática bicarbonato
Alcalinización del medio intestinal
Degrada completamente
la amilosa
Parcialmente al glucógeno o amilopectina
Maltosa
Maltosas
Maltotriosas
Isomaltosas
(Dextrina limite)
Isomaltasa o (16) glucosidasa
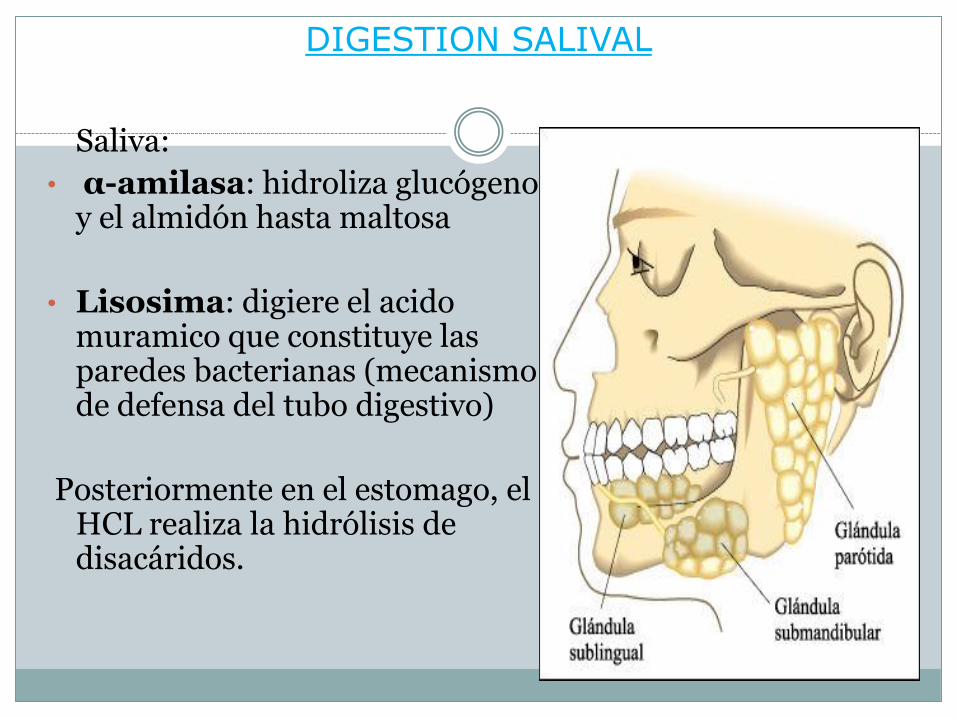
DIGESTION SALIVAL
Saliva:
• α-amilasa: hidroliza glucógeno y el almidón hasta maltosa
• Lisosima: digiere el acido muramico que constituye las paredes bacterianas (mecanismo de defensa del tubo digestivo)
Posteriormente en el estomago, el HCL realiza la hidrólisis de disacáridos.
DIGESTION PANCREATICA
La secreción pancreática contiene:
• α-amilasa pancreática:
• hidroliza los enlaces α-1,4, el polisacárido se transforma en una mezcla de oligosacáridos lineales:
• Maltosa
• Isomaltosa
• Maltotriosa
• α-dextrinas.
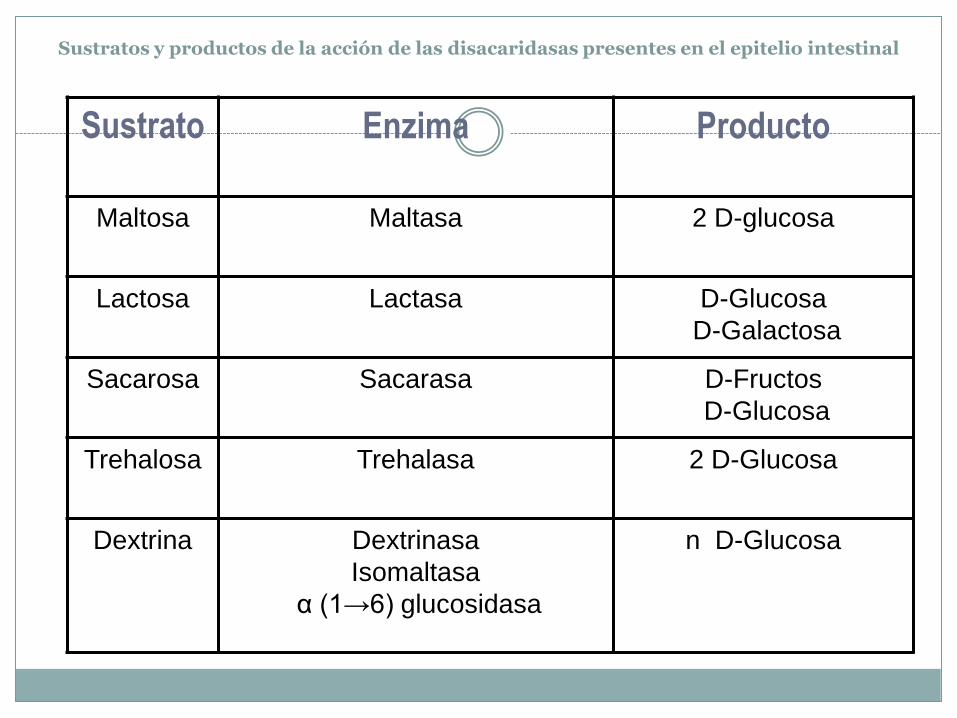
Sustratos y productos de la acción de las disacaridasas presentes en el epitelio intestinal
Sustrato Enzima Producto
Maltosa Maltasa 2 D-glucosa
Lactosa Lactasa D-Glucosa
D-Galactosa
Sacarosa Sacarasa D-Fructos
D-Glucosa
Trehalosa Trehalasa 2 D-Glucosa
Dextrina Dextrinasa
Isomaltasa
α (1→6) glucosidasa
n D-Glucosa

ABSORCION DE LOS MONOSACARIDOS
Difusión Pasiva a través de poros
Difusión Facilitada por Transportadores
Transporte Activo
Pinocitocis
Exocitosis
Endocitosis

DEFECTOS DE LA DIGESTION Y ABSORCION DE CHO
1. Mala absorción:
Imposibilidad de absorber correctamente los CHO
Causas:
Deficiencia enzimática inducida por una enfermedad. El defecto más común es la deficiencia de lactasa.
También puede haber una deficiencia de sacarasa e isomaltasa (con síntomas iguales a lod de la deficiencia de lactasa)

Sistemas de transporte en la membrana epitelial:
1. El transportador de Glucosa (GLUT)
2. El cotransporte Glc/Na+
GLUT:
Son prot con 12 dominios transmembrana(regiones de la prott que atraviesan la membrana).
Existen por lo menos 5 GLUT en el organismo.
Es un transporte facilitado
Son transportadores pasivos del tipo uniporte
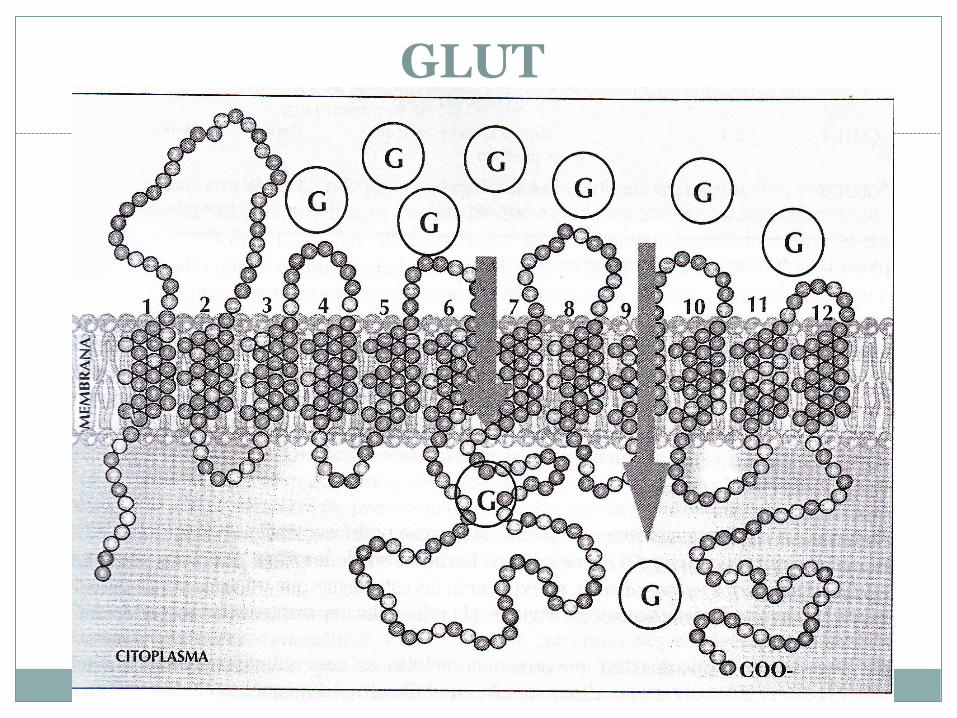
GLUT
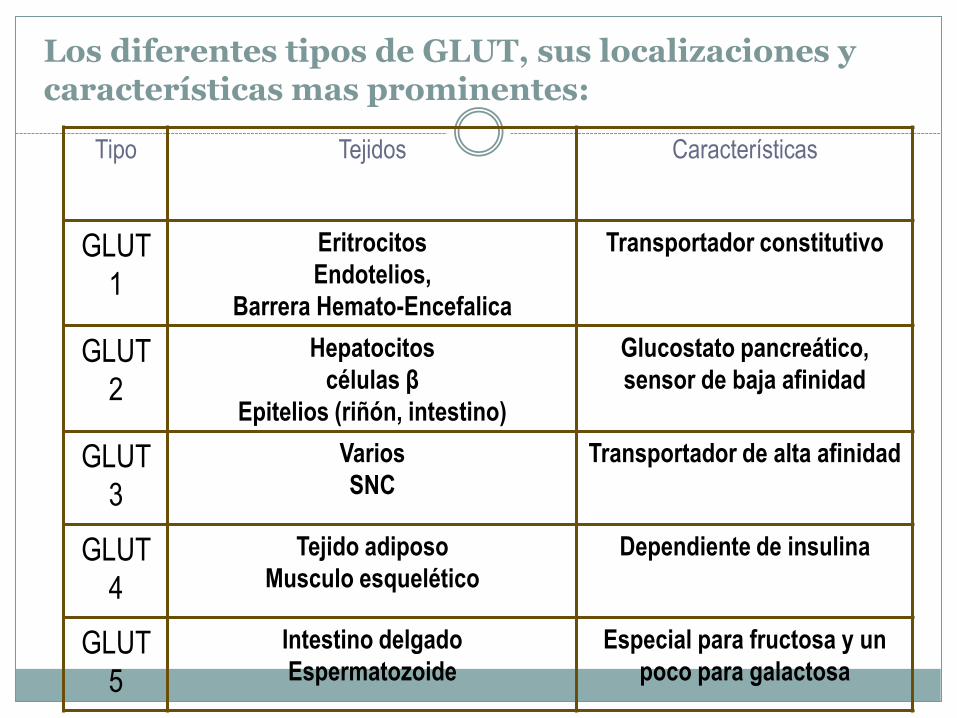
Los diferentes tipos de GLUT, sus localizaciones y características mas prominentes:
Tipo Tejidos Características
GLUT
1
Eritrocitos
Endotelios,
Barrera Hemato-Encefalica
Transportador constitutivo
GLUT
2
Hepatocitos
células β
Epitelios (riñón, intestino)
Glucostato pancreático,
sensor de baja afinidad
GLUT
3
Varios
SNC
Transportador de alta afinidad
GLUT
4
Tejido adiposo
Musculo esquelético
Dependiente de insulina
GLUT
5
Intestino delgado
Espermatozoide
Especial para fructosa y un
poco para galactosa
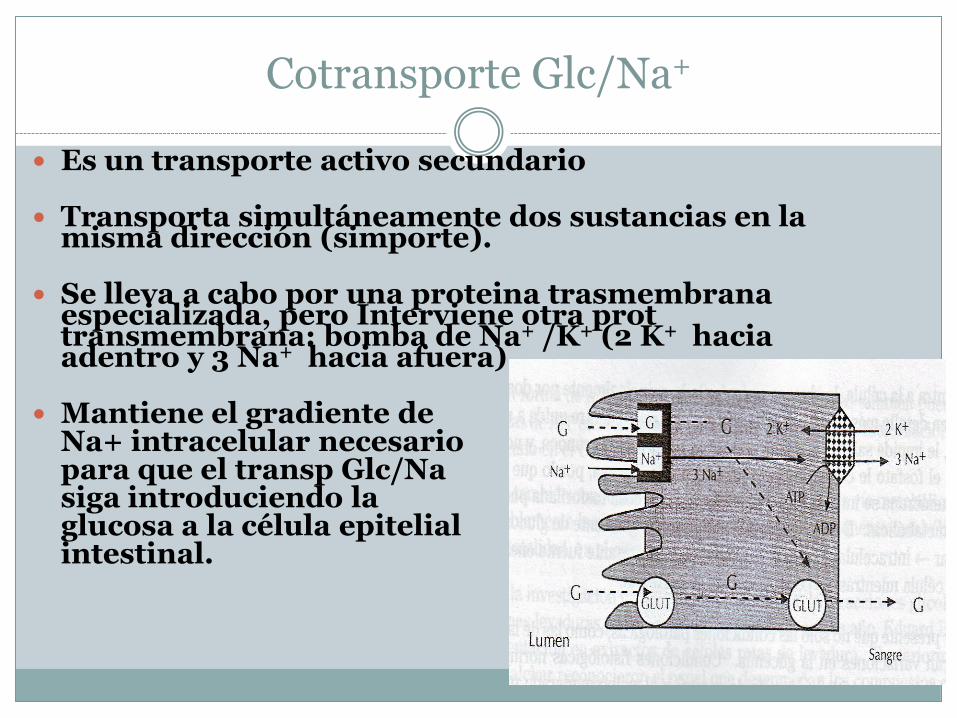
Cotransporte Glc/Na+
Es un transporte activo secundario
Transporta simultáneamente dos sustancias en la misma dirección (simporte).
Se lleva a cabo por una proteina trasmembranaespecializada, pero Interviene otra prottransmembrana: bomba de Na+ /K+ (2 K+ hacia adentro y 3 Na+ hacia afuera)
Mantiene el gradiente de Na+ intracelular necesariopara que el transp Glc/Nasiga introduciendo la glucosa a la célula epitelial intestinal.

METABOLISMO DE CH
Glicemia:
[Glucosa] en sangre cuyo valor normal es 60-90 mg/100 ml (metabolismo normal).
Depende de: alimentación, actividad celular, entrada y salida de Glucosa en la sangre.
En el interior de la celula la Glc es fosforilada:
- El GLUT no la reconoce
- Mayor polaridad a la Glc

Glucolisis
Catabolismo de los Hidratos de Carbono:

Glucolisis
Procede de las palabras griegas que significan
Glico= “dulce” Lisis= “romper”.
Es la ruta por medio de la cual los azucares de 6 carbonos se rompen, dando lugar a un compuesto de 3 carbonos, el piruvato.
Reacción neta de glucólisis:
Glucosa + 2 ADP + 2 piruvato + 2 ATP
2 NAD(+) + 2P + 2 NADH + 2 H(+)
+ 2 H20

La conversión de glucosa a piruvato se acompaña no solo
por la síntesis de ATP sino también por la reducción deNAD+ a NADH en la etapa de la gliceraldehido 3-fosfatodeshidrogenasa.
La enzima sacarasa, fijada sobre la superficie intestinal mediante una cadena de 30 aminoácidos, cataliza la hidrólisis de la sacarosa ingerida a glucosa más fructosa (la fructosa es metabolizada por el hígado)

LA LACTOSA, COMO LA SACAROSA, SE PUEDE CATABOLIZAR VIA GLUCOLISIS.
Disacárido lactosa fuente importante de energía para la crianza de los mamíferos, incluyendo los bebes humanos.
Se requieren 4 enzimas para convertir la galactosa en glucosa 6-fosfato para que entre en la vía glucolítica y después sea metabolizada a piruvato.
La intolerancia a la lactosa que resulta de la deficiencia en lactasa es una situación muy común en el ser humano adulto.

Glucólisis
La Glucosa es degradada para dar origen a dos moléculas de piruvato.
Objetivo principal: almacenar energía en forma de ATP y de NADH
Es la 1º vía metabólica que se dilucido. Es netamente catabólica Se lleva a cabo en el citoplasma celular Después de la formación de piruvato, puede ocurrir la
glucólisis aeróbica, la fermentación láctica o la fermentación alcohólica.
Se compone de 10 reacciones consecutivas, distribuidas en dos fases: preparatoria (1-5)
y retributiva (6-10).

Glucolisis
Se dan 3 tipos de transformaciones qq:
1. Degradación del esqueleto de carbono de
la Glc para formar piruvato
2. Fosforilación del ADP para formar ATP (a
nivel de sustrato)
3. Transferencia de un H+ al NAD+ para
formar NADH
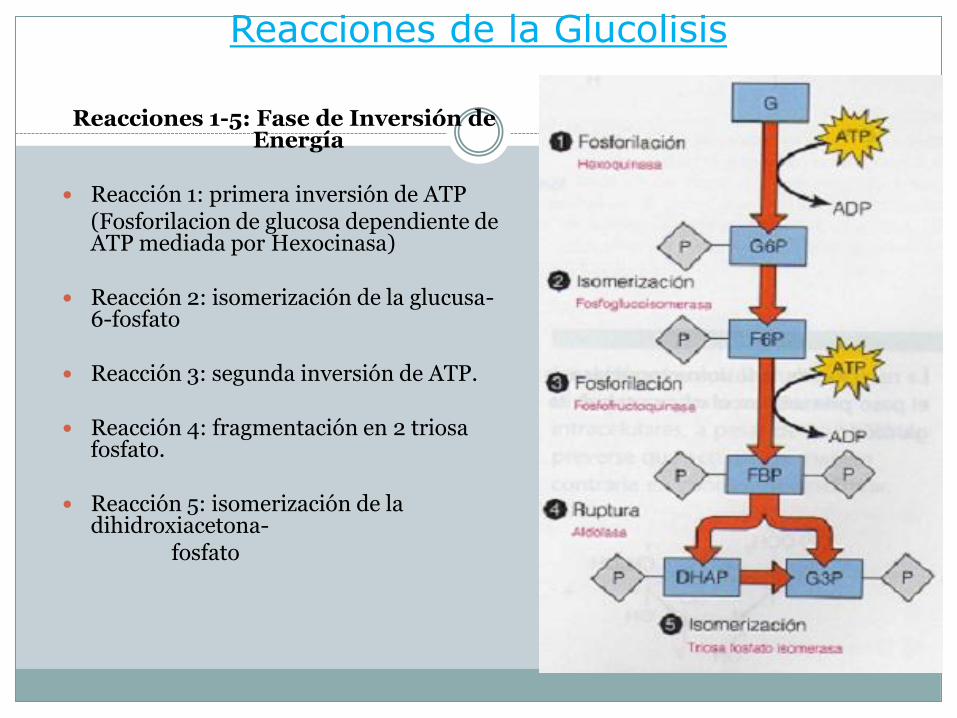
Reacciones de la Glucolisis
Reacciones 1-5: Fase de Inversión de Energía
Reacción 1: primera inversión de ATP (Fosforilacion de glucosa dependiente de ATP mediada por Hexocinasa)
Reacción 2: isomerización de la glucusa-6-fosfato
Reacción 3: segunda inversión de ATP.
Reacción 4: fragmentación en 2 triosa fosfato.
Reacción 5: isomerización de la dihidroxiacetona-
fosfato

Factor regulador clave:
La concentración del producto: glucosa 6-fosfato, (inhibe alostéricamente la hexocinasa)
La enzima hexoquinasa por su baja especificidad permite la fosforilacion de varios azucares hexosas, entre ellos la fructosa y la manosa.
El segundo sustrato para hexocinasa (complejo Mg 2+ - ATP).
El ATP aislado es un potente inhibidor competitivo de la hexocinasa.
La participación esencial del Mg 2+ es mantener protegidos las cargas negativas del fosfato, permitiendo que el fosforo sea más accesible.
Primera reaccion:

Una cinasa es una enzima que transfiere grupos fosforilo entre el ATP y un metabolito, el cual sirve como aceptor del fosforilo se identifica con el prefijo del nombre de la cinasa.
Se ha demostrado la existencia de cuatro isoenzimas:
Isoenzima I:
Su actividad no depende de la insulina
Casi siempre esta saturada por el sustrato
Se encuentra diversos tejidos: cerebro, hígado, riñon y pulmon.

Isoenzima II:
En musculo esquelético y tejido cardiaco, e hígado
Su actividad se aumenta por la insulina.
Isoenzima III:
se encuentra en menor cantidad que las anteriores, en la mayoría de los tejidos.
Isoenzima IV (glucocinasa)
se encuentra en el hígado de muchas especies
es regulada por la insulina.

Se ha demostrado presencia de 2 isoenzimas A2 Y B2:
A2: esta en tejidos con importante metabolismo anaeróbico, como el musculo esquelético blanco.
B2: predomina en el tejido aeróbico, como es el caso del musculo rojo y el hígado.
Este mecanismo de reacción involucra una catálisis general acido-basicacon las siguientes etapas:
Un acido, en el grupo ε-amino de la lisina cataliza la abertura del anillo.
Una base, acepta el proton acidico del carbono 2, para formar un cis-enediol.
El proton es reemplazado en el carbono 1.
La enzima requiere como cofactor el Mg 2+, y sus inhibidores son EDTA y 2-deoxi-glucosa-6-fosfato.
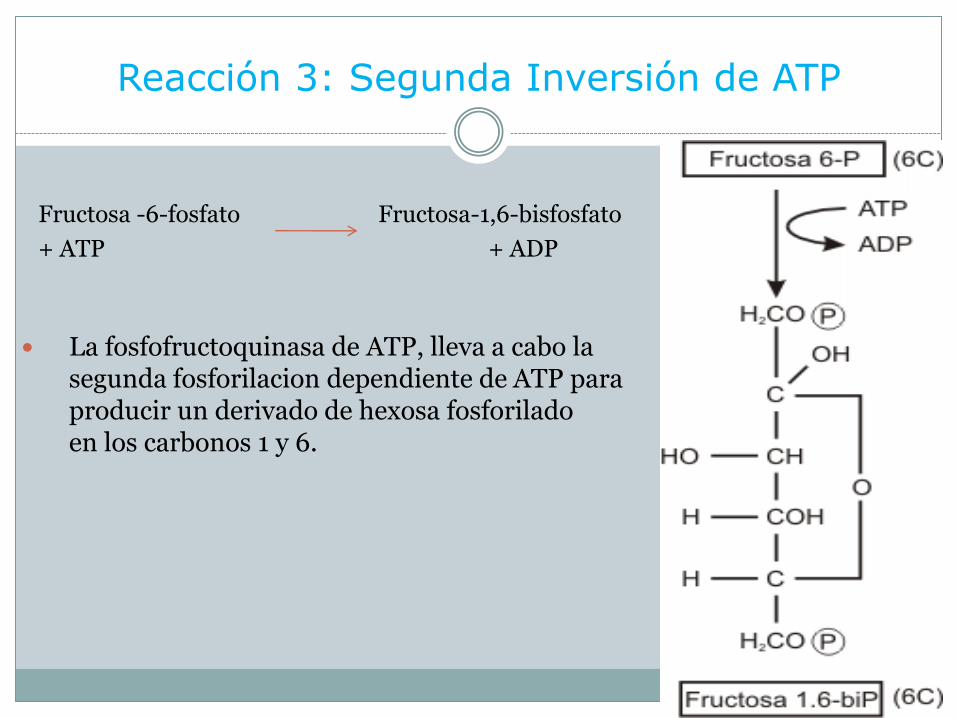
Reacción 3: Segunda Inversión de ATP
Fructosa -6-fosfato Fructosa-1,6-bisfosfato
+ ATP + ADP
La fosfofructoquinasa de ATP, lleva a cabo la segunda fosforilacion dependiente de ATP para producir un derivado de hexosa fosforiladoen los carbonos 1 y 6.

El producto, fructosa-1,6-bisfosfato, (Bisfosfato: cuando dos fosfatos están separados entre si; al estar unidos a diferentes carbonos en la misma molécula).
Esta reacción es irreversible in vivo, y es importante porque la fosfofructoquinasaconstituye el lugar principal de regulación del flujo de carbono a través de la glucolisis.

Al ser uno de los sitios claves de regulación de la glucolisis se han demostrado varios inhibidores y activadores:
Inhibidores alostericos:
ATP es un inhibidor alosterico que actúa incrementando la Km de la enzima para la F-6-P.
citrato
Acidos grasos de cadena larga
Los activadores alostericos:
(Pi),
AMP,
AMPc,
ADP
F-6-P.
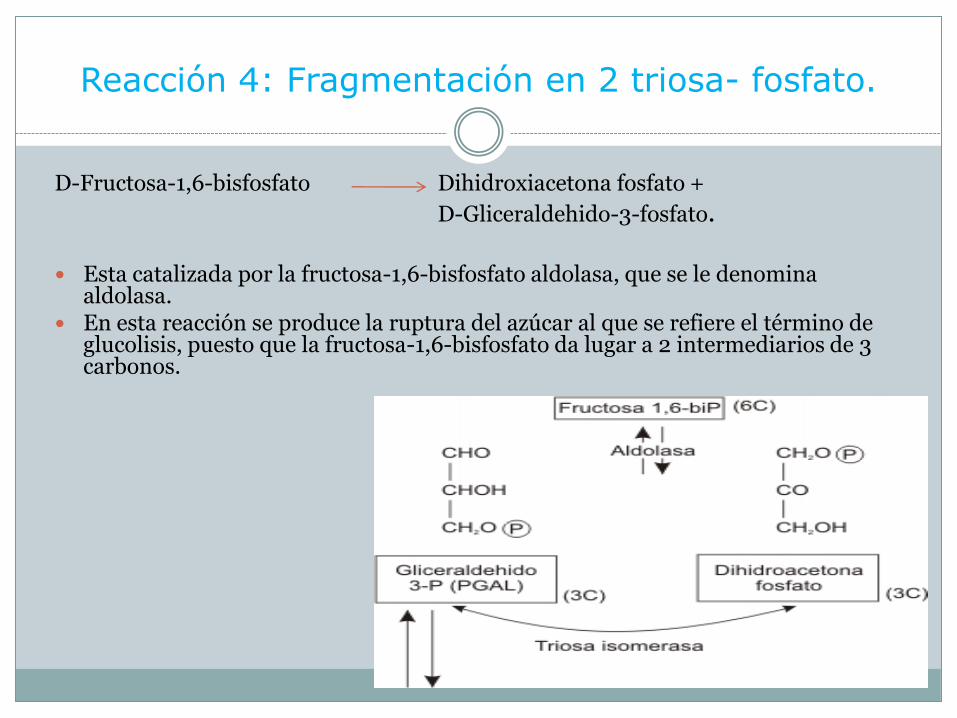
Reacción 4: Fragmentación en 2 triosa- fosfato.
D-Fructosa-1,6-bisfosfato Dihidroxiacetona fosfato +
D-Gliceraldehido-3-fosfato.
Esta catalizada por la fructosa-1,6-bisfosfato aldolasa, que se le denomina aldolasa.
En esta reacción se produce la ruptura del azúcar al que se refiere el término de glucolisis, puesto que la fructosa-1,6-bisfosfato da lugar a 2 intermediarios de 3 carbonos.

Un evento clave es la formación de una base de Schiff entre un residuo de lisina de la enzima y el grupo carbonilo del sustrato.
La base Schiff ancla el sustrato a la enzima y proporciona un átomo de nitrógeno cargado positivamente que sirve como un vertedero de electrones en las etapas subsiguientes.
La reacomodación de los electrones se inicia por el anión triolato de un residuo desprotonado de cisterna de la enzima, y culmina en la ruptura del primer producto, el gliceraldehido 3-fosfato y la formación de una enamina.
Después de la tautomerización de la enamina, la hidrólisis de la basa de Schiff libera la dihidroxiacetona fosfato.

Hay 2 tipos de aldolasas:
las aldolasas tipo I: que se encuentran en células animales. Las aldolasas tipo II: que son dependientes del Zn 2+ y se hallan
en bacterias y hongos.
En los vertebrados la isoenzima tipo I presenta 3 tipos de variantes : A,B y C.
Tipo I A: es la más eficaz en la glucolisis, se encuentra en el musculo, y es predominante en embriones de mamíferos.
Tipo I B: es importante para el metabolismo de la fructosa, y en la glucogénesis. Se ha detectado en el hígado, riñon e intestino delgado.
Tipo I C: se presenta en tejido nervioso, asi como en embriones de aves.
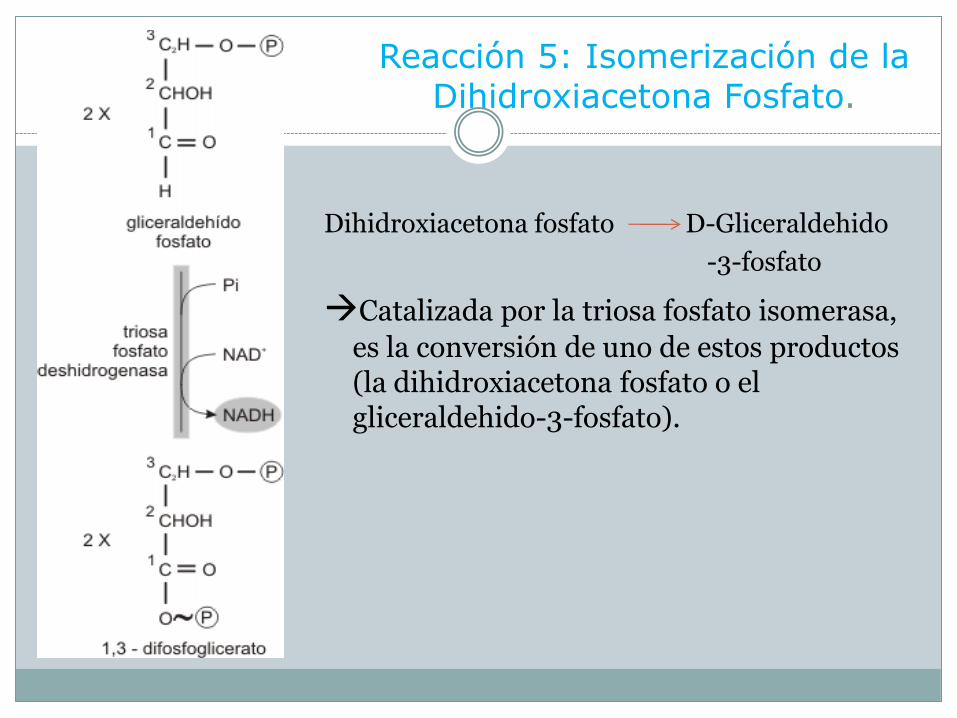
Reacción 5: Isomerización de la Dihidroxiacetona Fosfato.
Dihidroxiacetona fosfato D-Gliceraldehido
-3-fosfato
Catalizada por la triosa fosfato isomerasa,
es la conversión de uno de estos productos (la dihidroxiacetona fosfato o el gliceraldehido-3-fosfato).

La isomerización de la dihidroxiacetona fosfato se produce a través de un intermediario enediol. En este punto la glucolisis ha gastado 2 moléculas de ATP y ha convertido una hexosa en 2 moléculas de gliceraldehido-
3-fosfato.
No utiliza cofactores o iones metálicos. El mecanismo de reacción es similar al de la fosfohexosa isomerasa. Es una reacción reversible.
La deficiencia de esta enzima produce anemia hemolítica crónica no esferocitica, crecimiento retrasado y trastornos musculares y neurológicos.
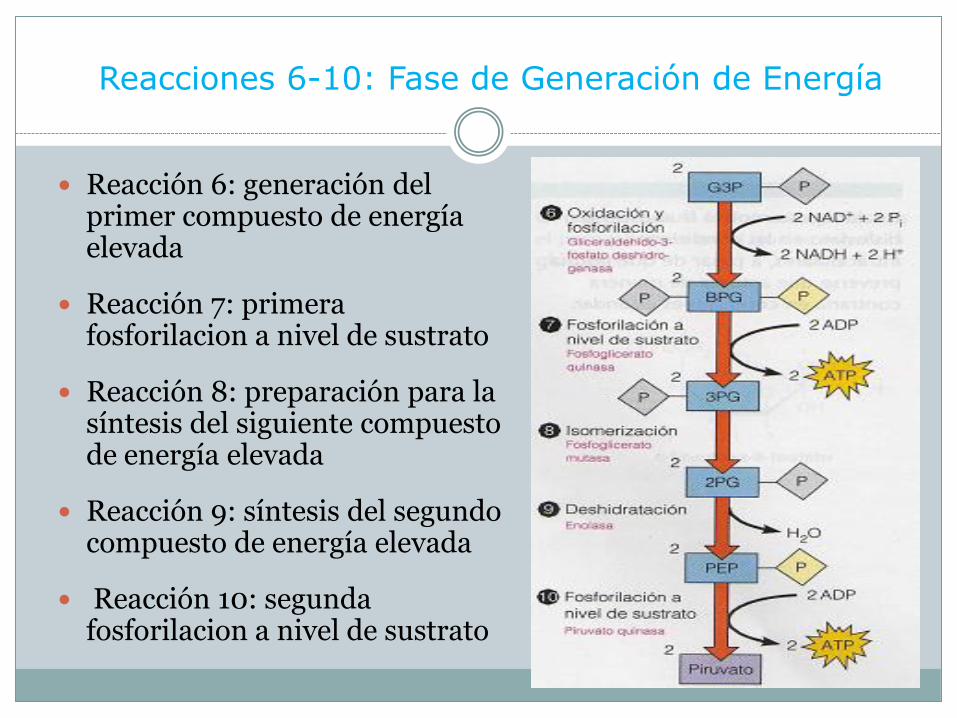
Reacciones 6-10: Fase de Generación de Energía
Reacción 6: generación del primer compuesto de energía elevada
Reacción 7: primera fosforilacion a nivel de sustrato
Reacción 8: preparación para la síntesis del siguiente compuesto de energía elevada
Reacción 9: síntesis del segundo compuesto de energía elevada
Reacción 10: segunda fosforilacion a nivel de sustrato
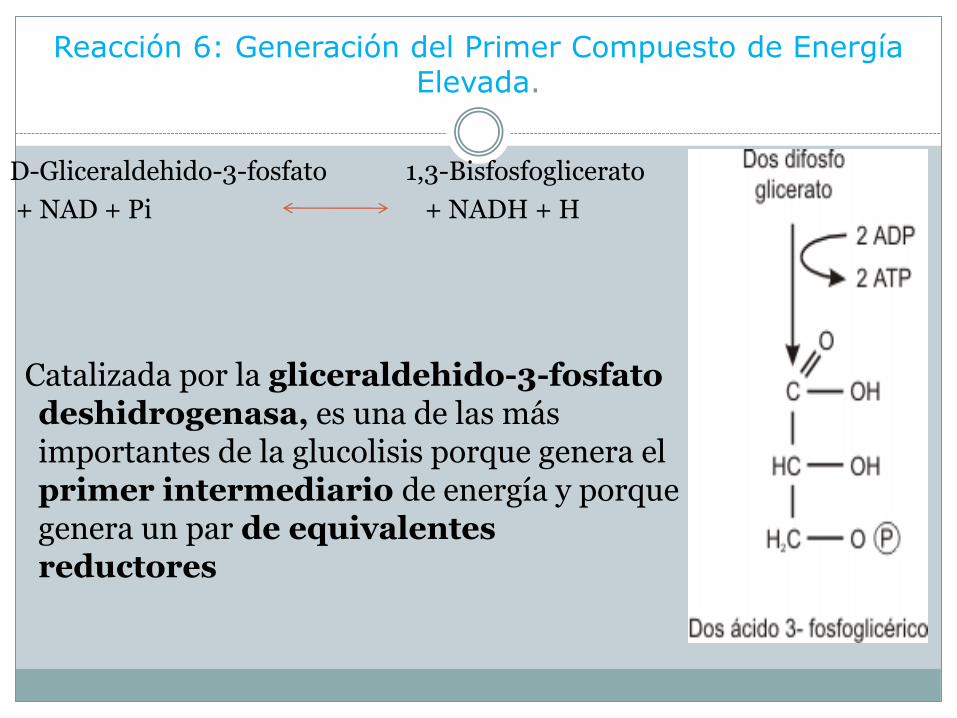
Reacción 6: Generación del Primer Compuesto de Energía Elevada.
D-Gliceraldehido-3-fosfato 1,3-Bisfosfoglicerato
+ NAD + Pi + NADH + H
Catalizada por la gliceraldehido-3-fosfato deshidrogenasa, es una de las más importantes de la glucolisis porque genera el primer intermediario de energía y porque genera un par de equivalentes reductores

se puede ilustrar en cinco etapas.
Etapa 1: el grupo –S(-) de un residuo de cisterna ataca al grupo carbonilo del gliceraldehido 3-fosfato, dando como resultado la formación de un tiohemiacetal unido covalentemente.
Etapa 2: se remueve un ion hidruro del C-1 del tiohemiacetal por NAD(+).
Etapa 3: Los productos se disocia la NADH de la enzima y es remplazada por NAD (+).

Etapa 4: el fosfato se fija en el sitio activo y ataca al grupo carbonilo del intermediario tioacil-enzima.
Etapa final: el producto 1,3 bisfosfoglicerato se disocia del sitio activo de la enzima, terminando el ciclo catalítico.
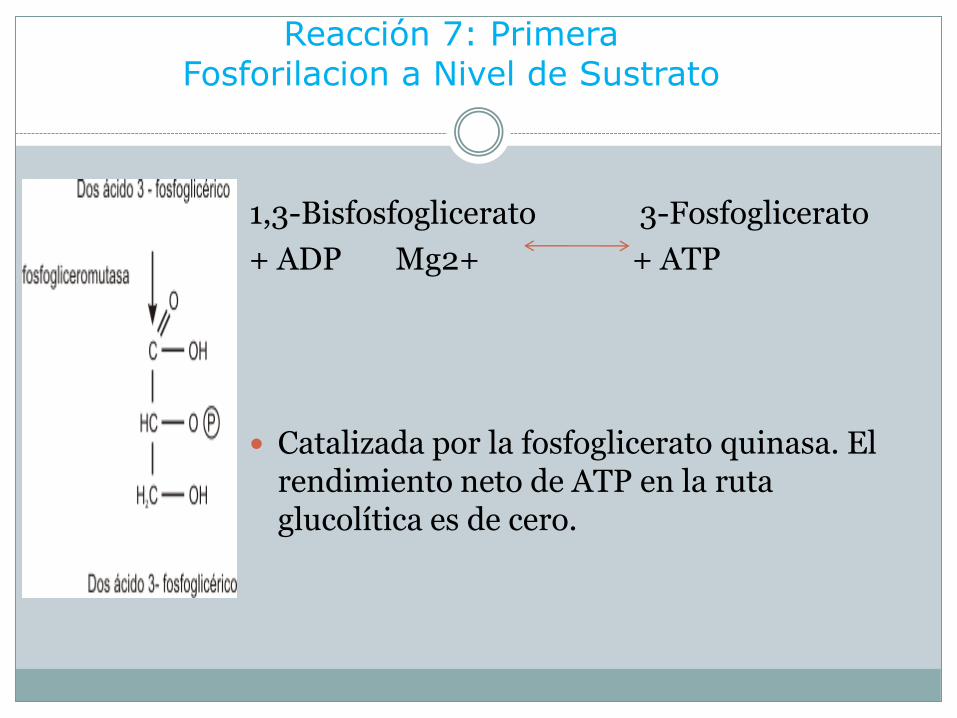
Reacción 7: Primera Fosforilacion a Nivel de Sustrato
1,3-Bisfosfoglicerato 3-Fosfoglicerato
+ ADP Mg2+ + ATP
Catalizada por la fosfoglicerato quinasa. El rendimiento neto de ATP en la ruta glucolítica es de cero.

Es una fosforilacion a nivel de sustrato, o sea que se forma una molécula de ATP a partir de una de ADP, sin intervención de cadena respiratoria.
La reacción requiere de Mg2+, o Mn y ADP.
Sus inhibidores son el Ac. di,tio,bis-nitrobenzoico y EDTA.
La actividad de la enzima está en función de relación ATP/ADP. (Enfermedad que causa la sustitución genética del aminoácido asparagina por treonina).
Reacción 8: Preparación para la Síntesis del Siguiente Compuesto de Energía Elevada.
Mg2+
3-Fofoglicerato 2-Fosfoglicerato
La activación del 3-fosfoglicerato se inicia con una isomerización catalizada por la fosfoglicerato mutasa, para esta reacción se requiere Mg 2+.
En el primer paso de la reacción se transfiere el fosfato desde la enzima al sustrato para formar un intermediario, el 2, 3-bisfosfoglicerato.
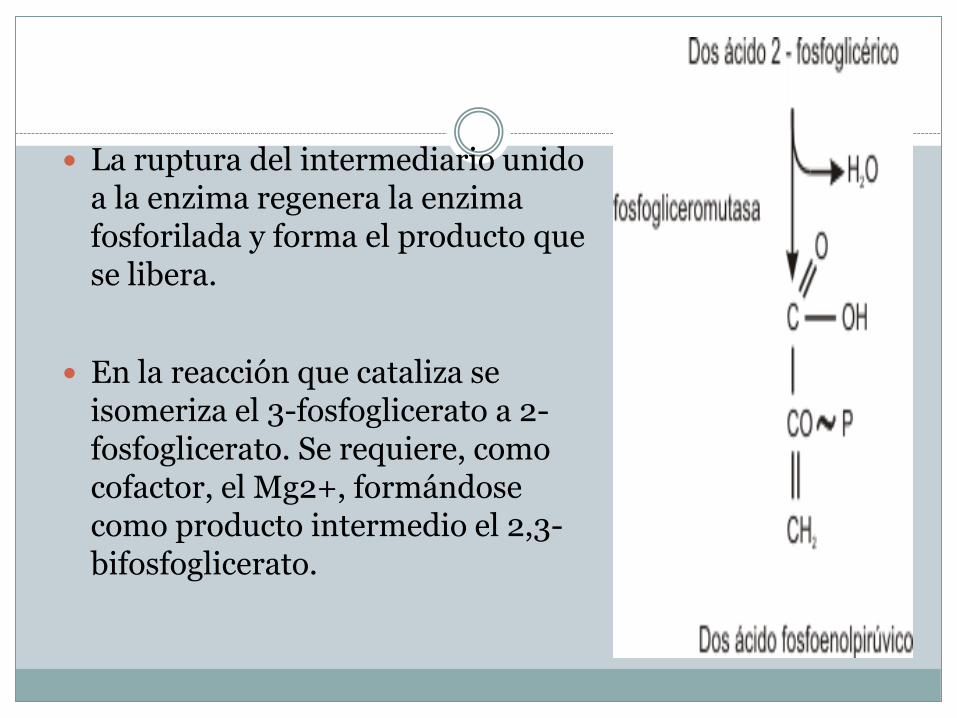
La ruptura del intermediario unido a la enzima regenera la enzima fosforilada y forma el producto que se libera.
En la reacción que cataliza se isomeriza el 3-fosfoglicerato a 2-fosfoglicerato. Se requiere, como cofactor, el Mg2+, formándose como producto intermedio el 2,3-bifosfoglicerato.

Reacción 9: Síntesis del Segundo Compuesto de Energía Elevada
Mg2+
2-Fosfoglicerato Fosfoenolpiruvato + H20
Catalizada por la enolasa, genera otro compuesto de energía súper-elevada, el fosfoenolpiruvato, que participa en la segunda fosforilacion a nivel de sustrato de la glucolisis.
La reacción consiste en una deshidratación simple, y su efecto consiste en aumentar enormemente la energía libre de hidrólisis del enlace fosfato.
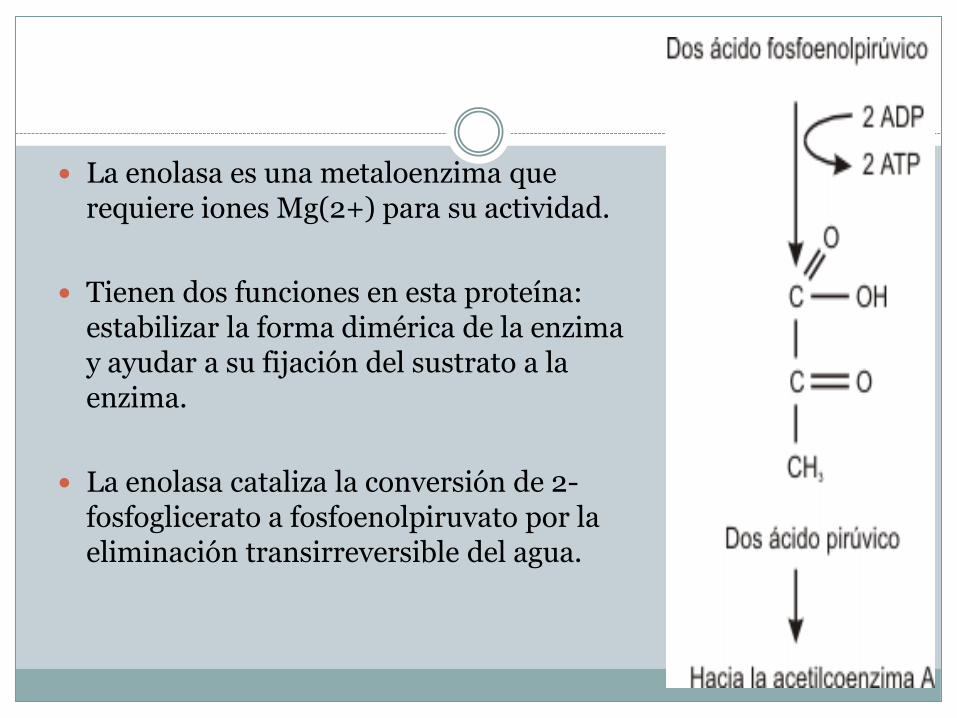
La enolasa es una metaloenzima que requiere iones Mg(2+) para su actividad.
Tienen dos funciones en esta proteína: estabilizar la forma dimérica de la enzima y ayudar a su fijación del sustrato a la enzima.
La enolasa cataliza la conversión de 2-fosfoglicerato a fosfoenolpiruvato por la eliminación transirreversible del agua.

Reacción 10: Segunda Fosforilacion a Nivel de
Sustrato
Mg2+
Fosfoenolpiruvato + H+ ADP Piruvato + ATP
K+
Catalizada por la piruvato quinasa, el fosfoenolpiruvato transfiere su grupo fosforilo al ADP en otra fosforilacion a nivel de sustrato.
La enzima requiere Mg2+ y K+.
El aumento de la piruvato quinasa incrementa la velocidad de generación de energía mediante la glucolisis.
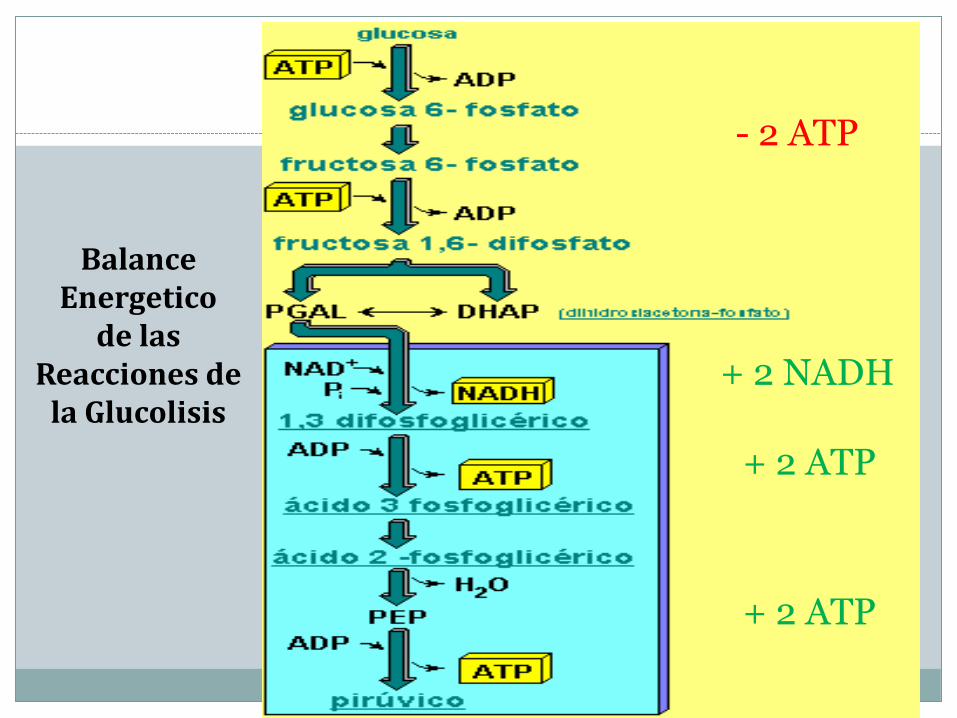
Balance Energetico
de las Reacciones de
la Glucolisis
- 2 ATP
+ 2 ATP
+ 2 NADH
+ 2 ATP

Regulacion de la Glicolisis
El flujo de glucosa a la glicolisos debe ser constantemente regulado paramantener niveles constantes de ATP e intermediarios metabolicos parabiosintesis.
Los ajustes necesarios se realizan por la interrelacion entre: Consumo de ATP Regeneracion del NADH Regulacion Alosterica de las enzimas glicoliticas: hexocinasa, PFK-1 y
piruvato cinasa Variacion segundo a segundo de concentracion de metabolitos que reflejan
el balance celular entre la produccion y consumo de ATP En una escala a mas largo plazo, es regulada a nivel hormonal: Insulina,
glucagon, epinefrina y cortisol. A largo plazo por la expresion genetica de las enzimas glicoliticas
Tarea: papel de hormonas reguladoras de la glicolisis

Catabolismo de la glucosa en tejidos cancerosos
La velocidad del consumo de Glucosa y la glicolisis se encuentraaumentada 10 veces en los tumores solidos comparando con tejidos no cancerosos. Las celulas tumorales comunmente se encuentran en estado de hipoxia (limitada disponibilidad de oxigeno), ya que inicialmente no poseen una red capilar queles supla oxigeno.
Como resultado las celulas cancerosas dependen de la glicolisis anaerobia para producir la mayoria de su ATP.
Utilizan mas glucosa que las celulas normales, produciendopiruvato y convirtiendolo en lactato para reciclar el NADH
La mayor tasa de glicolisis tambien es resultado del menornumero de mitocondrias en las celulas cancerosas (menorcantidad de ATP generado por fosforilacion oxidativa)

Adicionalmente, algunas celulas tumoralessobreproducen enzimas glicoliticas, como una isozima de la hexocinasa sensible a inhibicion por retroalimentacionde G-6-P lo que monopoliza el ATP mitocondrial para la conversion de glucosa a G-6-P obligando a la celula a un proceso continuado de glicolisis
El factor de transcripcion inducido por hipoxia (HIF-1) es una proteina que estimula la sintesis de por lo menos8 enzimas glucoliticas a nivel de ARN, lo que le da al tumor la capacidad de sobrevivir anaerobicamente hastaque se forma una red vascular capaz de suplir suficienteoxigeno
Catabolismo de la glucosa en tejidos cancerosos

Metabolismo de Hidratos de Carbono
MET DEL GLICOGENO: GLUCOGÉNESIS Y GLUCOGENÓLISIS
GLUCONEOGÉNESIS
RUTA DE LAS PENTOSAS FOSFATO.


Metabolismo del
Glucogeno

Metabolismo del Glucógeno
La síntesis y degradación del glucógeno están reguladas cuidadosamente para que pueda disponerse de suficiente glucosa para las necesidades energéticas del organismo.
La glucogénesis y la glucogenólisis están controladas principalmente por tres hormonas:
Insulina
Glucagón
Adrenalina.

Glucogénesis
La glucogénesis es la ruta anabólica por la que tiene lugar la síntesis de glucógeno (también llamado glicógeno) a partir de un precursor más simple, la glucosa.
Se lleva a cabo principalmente en el hígado, y en menor medida en el músculo.

• La glucogénesis es estimulada por la hormona insulina, secretada por las células β de los islotes de Langerhans del páncreas.
• Es inhibida por su contra reguladora, la hormona glucagón, secretada por las células α de los islotes de Langerhans del páncreas, que estimula la ruta catabólica llamada glucogenólisis para degradar el glucógeno almacenado y transformarlo en glucosa y así aumentar la glicemia
Glucogénesis

La glucogénesis se realiza en el hígado, músculos y otras zonas orgánicas.
En el hígado se puede producir a partir de la glucosa, e indirectamente (mediante interconversión a glucosa) de la fructosa, galactosa y también de los metabolitos capaces de sintetizar glucosa.
Glucogénesis

Síntesis de glucógeno
La síntesis de glucógeno se produce tras una comida, cuando la concentración sanguínea de glucosa es elevada.
La síntesis de glucógeno se cree que se inicia por la transferencia de glucosa desde la UDP-glucosa a un residuo específico de tirosina en una proteína denominada glucogenina.

Reacciones de la Glucogénesis(síntesis de glucógeno)
Síntesis de glucosa 1 fosfato
Síntesis de UDP glucosa
Síntesis de glicógeno a partir de UDP-glucosa
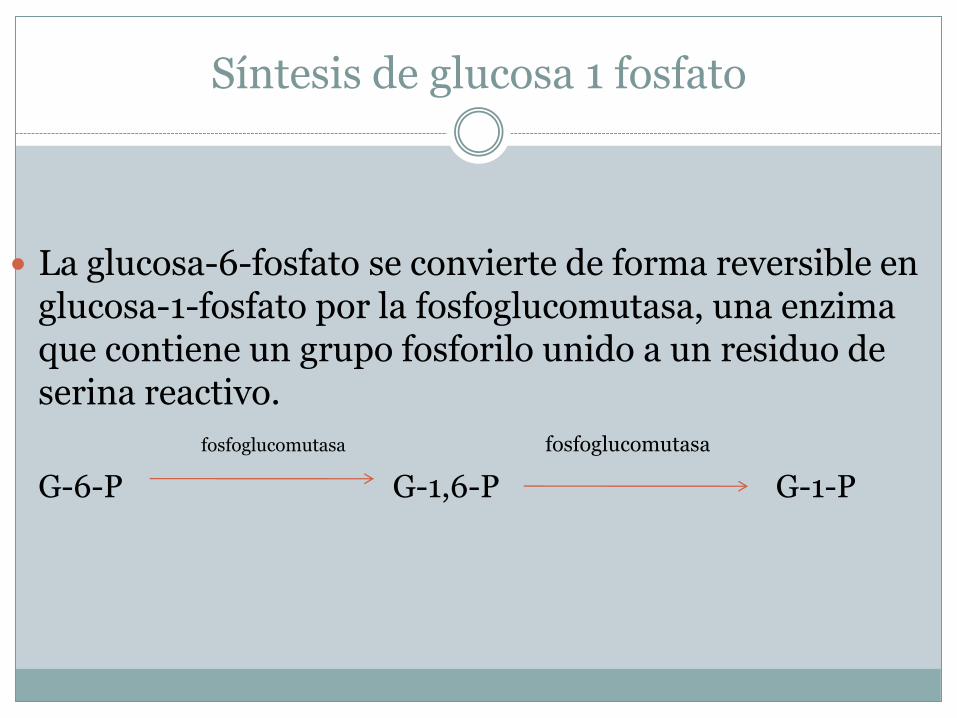
Síntesis de glucosa 1 fosfato
La glucosa-6-fosfato se convierte de forma reversible en glucosa-1-fosfato por la fosfoglucomutasa, una enzima que contiene un grupo fosforilo unido a un residuo de serina reactivo.
fosfoglucomutasa fosfoglucomutasa
G-6-P G-1,6-P G-1-P

Síntesis de UDP glucosa
La síntesis de nucleótidos-azúcar es una reacción común que precede la transferencia de azúcar y a los procesos de polimerización.
La uridina bifosfato glucosa (UDP glucosa) es mas reactiva que la glucosa y se mantiene de forma más segura en el lugar activo de las enzimas que catalizan las reacciones de transferencia.
Debido a q la UDP glucosa contiene dos enlaces fosforilo es una molécula muy energética.

Síntesis de glicógeno a partir deUDP-glucosa
Esta reacción requiere de dos enzimas:
▫ Glicógeno sintasa: cataliza la transferencia del grupo glucosilo de la UDP-glucosa a los extremos no reductores del glucógeno.
▫ Amilo alfa (1,4 1,6) glucosil transferasa(enzima ramificante): crea los enlaces α (1,6) para las ramificaciones de la molécula.
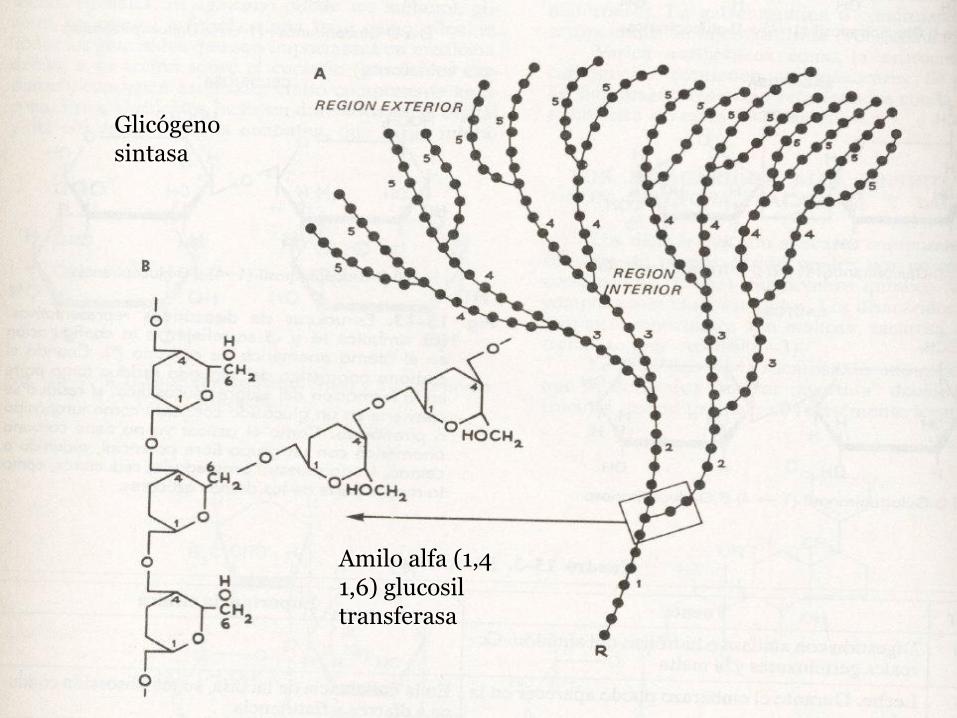
Amilo alfa (1,4 1,6) glucosiltransferasa
Glicógeno sintasa

Glucogenólisis(degradación de glucógeno)
Este proceso ocurre fundamentalmente en el hígado y músculo.
En musculo:
este tejido carece de enzima glucosa 6 fosfatasa, no puede liberar la glucosa al exterior de la célula y por consecuencia, no influye en la glucemia
la degradación de glucosa produce glucosa-6 fosfato, la cual se incorpora como metabolito de la glucólisis.
En hígado:
esta enzima si esta presente y puede aportar glucosa al torrente sanguíneo y así responder a las necesidades metabólicas.

La glucogenólisis requiere de tres enzimas:
• fosforilasa (glucogeno fosforilasa):
cataliza la fosforolisis del glicógeno (ruptura de un enlace por la adición de un fosfato), para producir glucosa 1 fosfato.
• desramificante:
remueve las ramificaciones del glicógeno, lo cual permite la continua acción de la fosforicasa.
También es capaz de hidrolizar el enlace alfa(1,6) liberando glucosa. Por consecuencia 90% del glicógeno se transforma en glucosa 1,6 fosfato y el 10% en glucosa.

fosfoglucomutasa:
convierte la glucosa 1 fosfato en glucosa 6 fosfato para que ingrese al proceso de glucólisis en el músculo y serán desfosforilada en el hígado para producir glucosa.
La glucogenólisis requiere de tres enzimas:

La degradación del glicógeno requiere de las dos reacciones siguientes:
1) Eliminación de la glucosa de los extremos no reductores del glicógeno:
Utilizando fosfato inorgánico la glicógeno fosforilasarompe los enlaces alfa (1,4) de las ramificaciones externas del glicógeno para dar glucosa 1 fosfato.
El glicógeno fosforilasa se detiene cuando llega a 4 residuos de glucosa hasta el punto de ramificación.

2) Hidrólisis de los enlaces glucosídicos alfa (1,6) en los puntos de ramificación del glicógeno.
La amilo alfa (1,6) glucosidasa que también se denomina enzima desramificante comienza a eliminar los puntos de ramificación alfa (1,6) transfiriendo los tres residuos de glucosa más externos de los cuatro unidos al punto de ramificación a un extremo no cercano.
La degradación del glicógeno requiere de las dos reacciones siguientes:

Regulación del metabolismo del glicógeno
El metabolismo del glicógeno está regulado de forma cuidadosa para evitar el derroche de energía.
Tanto la síntesis como la degradación están controladas mediante un mecanismo complejo con la participación de la insulina, el glucagón y la adrenalina.

Estas hormonas inician procesos que controlan varios conjuntos de enzimas.
La unión del glucagón a las células hepáticas estimula la glucogenólisis e inhibe la glucogénesis.
Al caer la concentración sanguínea de glucosa, horas después de una comida, el glucagón asegura la liberación de glucosa al torrente sanguíneo.
Regulación del metabolismo del glicógeno

Gluconeogénesis
Reacción anabólica que consiste en la formación de moléculas nuevas de glucosa a partir de precursores que no son hidratos de carbono y se produce principalmente en el hígado.

Estos precursores son:
El Lactato (que se forma en el músculo y los eritrocitos)
El Piruvato
El Glicerol (que se produce en la degradación de los TAG)
Intermedios del ciclo de Krebs
Determinados α-cetoácidos (moléculas derivadas de aminoácidos.)

La síntesis de Glucosa a partir del Piruvato es el proceso contrario a la glucólisis.
En determinadas situaciones como acidosis e inanición el riñón también puede formar glucosa.
Gluconeogénesis

Importancia
Entre comidas se mantienen las concentraciones sanguíneas adecuadas de glucosa por la hidrólisis del glucógeno hepático.
Cuando se agota el glucógeno hepático (ej. Alimentación con muchas grasas, ayuno prolongado o ejercicio excesivo) la ruta de la gluconeogénesisproporciona al organismo la glucosa necesaria ( los eritrocitos y el cerebro dependen exclusivamente de la glucosa como fuente de energía).

Reacciones de la gluconeogénesis (reacciones de circunvalación)
La secuencia de reacciones de la gluconeogénesis es en gran medida lo inverso de la glucólisis.
Dentro de la gluconeogénesis se dan reacciones alternativas catalizadas por enzimas diferentes, dado que dentro de la glucólisis existen tres reacciones que son irreversibles, las reacciones catalizadas por las enzimas:
hexoquinasa (Rxn 1: P de Glucosa)
la PFK-1 (Rxn 3: P de Fructosa 6P)
Piruvato quinasa (Rxn 10: PEP a Piruvato)

Al contrario que las reacciones de la glucólisis, que solo tienen lugar en el citoplasma, varias reacciones de la gluconeogénesis tienen lugar dentro de compartimentos celulares:
mitocondria:
las reacciones catalizadas por las enzimas piruvato carboxilasa
retículo endoplásmico:
las reacciones catalizadas por la glucosa-6-fosfatasa.
Reacciones de la gluconeogénesis (reacciones de circunvalación)

1) Sintesis PEP y Conversion del Oxaloacetato en Malato
2) Conversión de la fructosa-1,6-bisfosfato en
fructosa-6-fosfato
3) Formación de glucosa a partir de glucosa 6 fosfato
Reacciones de la gluconeogénesis(reacciones de circunvalación)
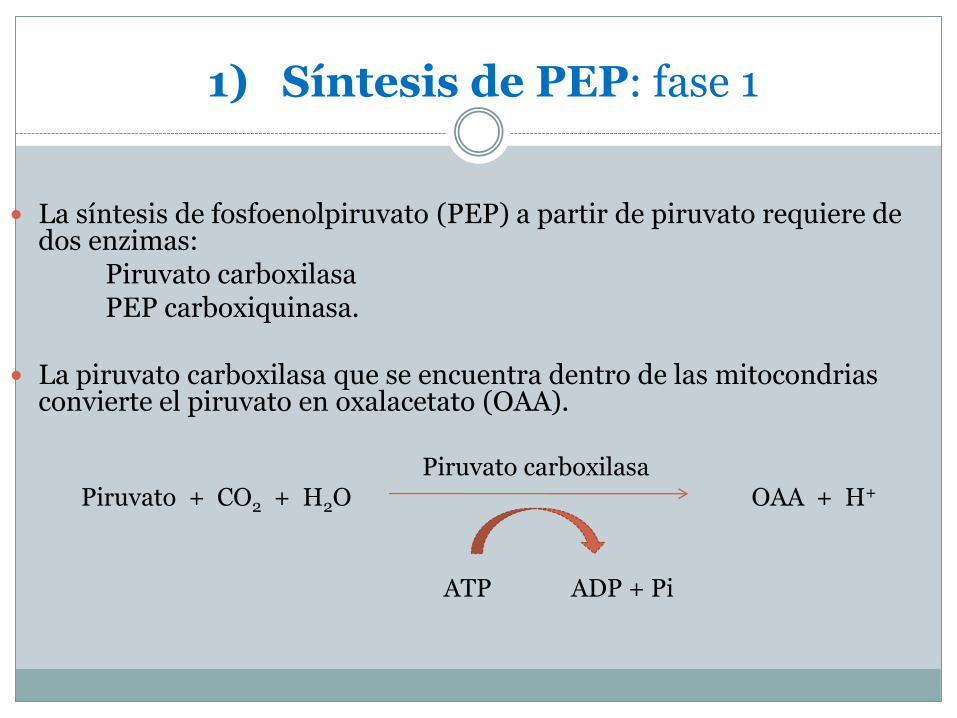
1) Síntesis de PEP: fase 1
La síntesis de fosfoenolpiruvato (PEP) a partir de piruvato requiere de dos enzimas:
Piruvato carboxilasaPEP carboxiquinasa.
La piruvato carboxilasa que se encuentra dentro de las mitocondrias convierte el piruvato en oxalacetato (OAA).
Piruvato carboxilasaPiruvato + CO2 + H2O OAA + H+
ATP ADP + Pi

Síntesis de PEP: fase 2
El oxalacetato se descarboxila y se fosforila por la PEP carboxiquinasa en una reacción impulsada por la hidrólisis de la guanosina trifosfato (GTP).
carboxiquinasa
OAA + PEP PEP + CO2
GTP GDP

La PEP carboxiquinasa se encuentra dentro de las mitocondrias de algunas especies y en el citoplasma de otras.
En el ser humano esta actividad enzimática se encuentra en ambos compartimientos.
Debido a que la membrana mitocondrial interna es impermeable al OAA, las células que carecen de PEP carboxiquinasa mitocondrial transfieren el OAA al citoplasma utilizando la lanzadera del malato.

La lanzadera del malato.
En este proceso, el OAA se convierte en malato por la malato deshidrogenasa mitocondrial.
OAA + NADH + H+ malato deshidrogenasa Malato + NADH+
Tras el transporte del malato hacia el citoplasma a través de la membrana mitocondrial, la reacción inversa (la conversión del malato a OAA) es catalizada por la malato deshidrogenasa citoplasmática.

2) Conversión de la fructosa-1,6-bisfosfato en fructosa-6-fosfato
La reacción irreversible de la glucólisis catalizada por la PFK-1 se circunvala por la fructosa-1,6-bisfosfatasa.
La fructosa-1,6-bifosfatasa cataliza la conversión de fructosa 1,6 bisfosfato a fructosa-6-fosfato.
Se le encuentra en el hígado, riñones y el músculo esquelético.

Conversión de la fructosa-1,6-bisfosfato en fructosa-6-fosfato
Esta reacción exergónica (que libera energía) es también irreversible en las condiciones celulares, el ATP reducido a ADP por la PFK-1 en la glucólisis no se regenera.
fructosa-1,6-bisfosfatasaFructosa-1,6-
bisfosfato + H2O
fructosa-6-fosfato + Pi

3) Formación de glucosa a partir de glucosa 6 fosfato
La glucosa 6 fosfatasa que solo se encuentra en el hígado y el riñón cataliza la hidrólisis irreversible de la glucosa 6 fosfato para formar glucosa y Pi.
Glucosa-6-fosfato glucosa 6 fosfatasa Glucosa + Pi
+H2O
A continuación la glucosa se libera al torrente sanguíneo

Cada una de las reacciones anteriores esta emparejada con una reacción opuesta irreversible en la glucólisis.
Cada conjunto de estas relaciones de pareja se denomina ciclo de sustrato.
La gluconeogénesis es un proceso que consume energía.
En lugar de generar ATP (como la glucólisis), la gluconeogénesis requiere la hidrólisis de seis enlaces fosfato de energía elevada.
Reacciones de la Gluconeogénesis(reacciones de circunvalación)

Regulación de la Gluconeogénesis
Los cambios en la disponibilidad de los sustratos son la causa de la mayor parte de las alteraciones en el metabolismo.
Hay tres mecanismos encargados de regular la actividad de las enzimas:
Cambios en la rapidez de la síntesis enzimática
Modificación covalente mediante fosforilaciónreversible
Efectos alostéricos

Cambios en la rapidez de la síntesis enzimática
Las enzimas que intervienen en el uso de la glucosa se vuelven más activas cuando hay exceso de glucosa y en estas condiciones, las enzimas a las que se debe la gluconeogénesis tienen baja actividad.
Ej:
La insulina secretada en respuesta al incremento de glucosa en la sangre intensifica la síntesis de las enzimas importantes en la glucólisis.

Modificación covalente mediante fosforilación reversible
El glucagón y la adrenalina, hormonas a las que se debe la disminución de glucosa en la sangre inhiben la glucólisis y estimulan la gluconeogénesis en el hígado mediante el aumento de la concentración de AMPc.
Este activa a su vez a la proteincinasa dependiente de AMPc, lo cual da lugar a la fosforilación y desactivación de la piruvato quinasa.

Efectos Alostéricos
La piruvato carboxilasa requiere el Acetil CoA como un activador alostérico en la gluconeogénesis.
La activación de la piruvato carboxilasa y la inhibición de la piruvato deshidrogenasa a causa del Acetil CoA explican la acción de la oxidación de AG que no afecta la oxidación de piruvato y estimula la gluconeogénesis.

Ruta de las Pentosas Fosfato

Ruta de las pentosas fosfato
Es otra ruta metabólica de la oxidación de la glucosa en la que no se genera ATP.
Sus productos principales son:
NADPH (agente reductor que se requiere en procesos anabólicos)
Ribosa 5 fosfato (componente estructural de nucleótidos y ácidos nucleicos)

Las rutas de las pentosas fosfato se producen en citoplasma en dos fases:
Fase Oxidativa: 3 reacciones
Fase no oxidativa: 2 reacciones
Ruta de las pentosas fosfato
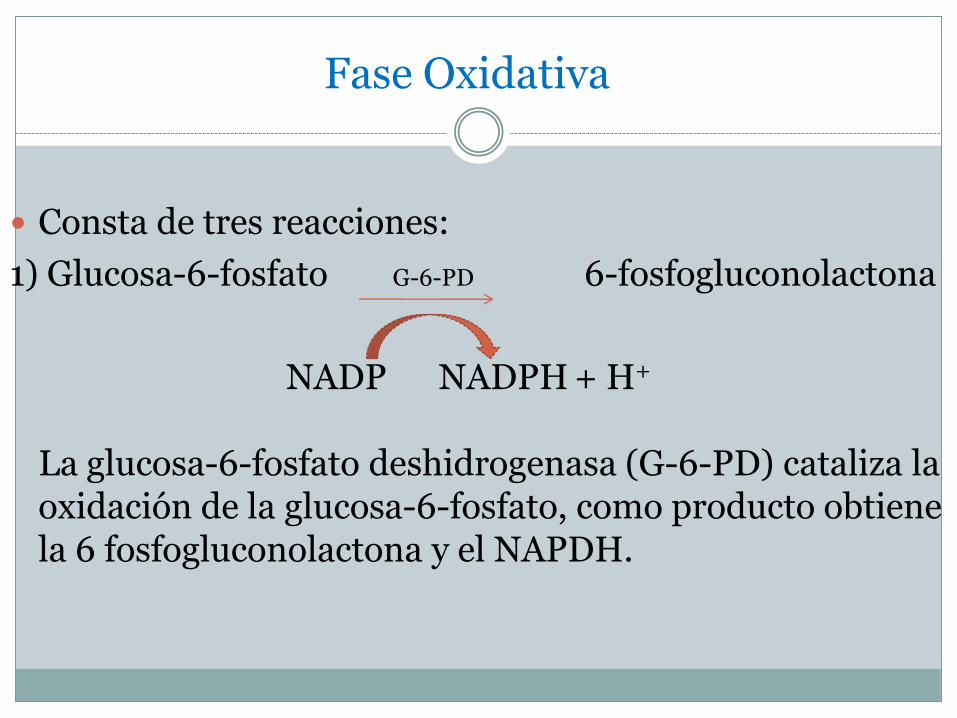
Consta de tres reacciones:
1) Glucosa-6-fosfato G-6-PD 6-fosfogluconolactona
NADP NADPH + H+
La glucosa-6-fosfato deshidrogenasa (G-6-PD) cataliza la oxidación de la glucosa-6-fosfato, como producto obtiene la 6 fosfogluconolactona y el NAPDH.
Fase Oxidativa
Fase oxidativa
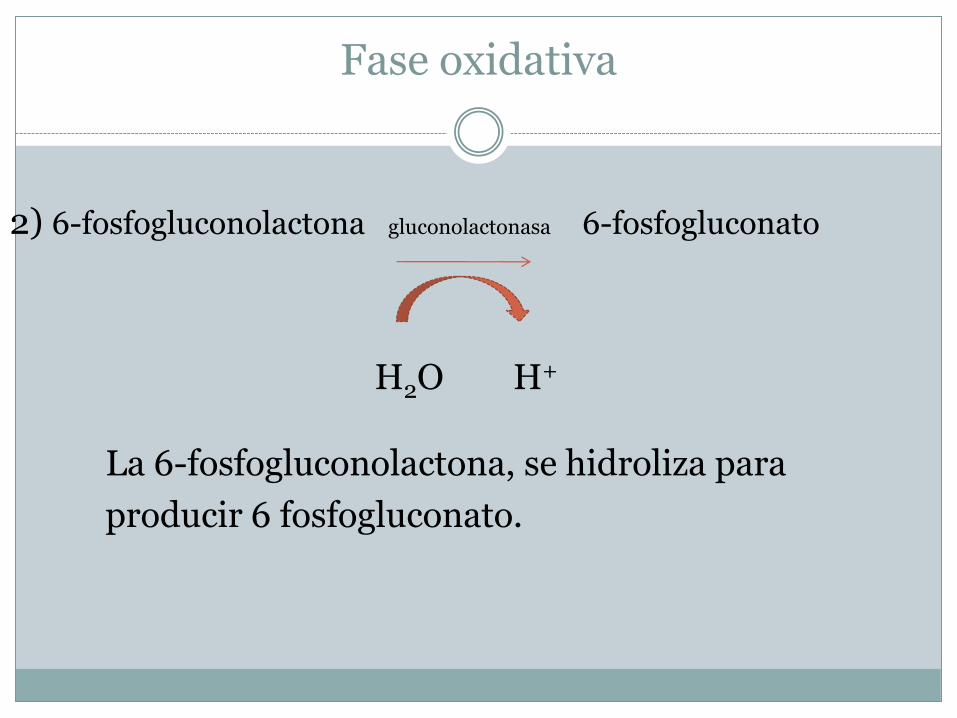
2) 6-fosfogluconolactona gluconolactonasa 6-fosfogluconato
H2O H+
La 6-fosfogluconolactona, se hidroliza para
producir 6 fosfogluconato.
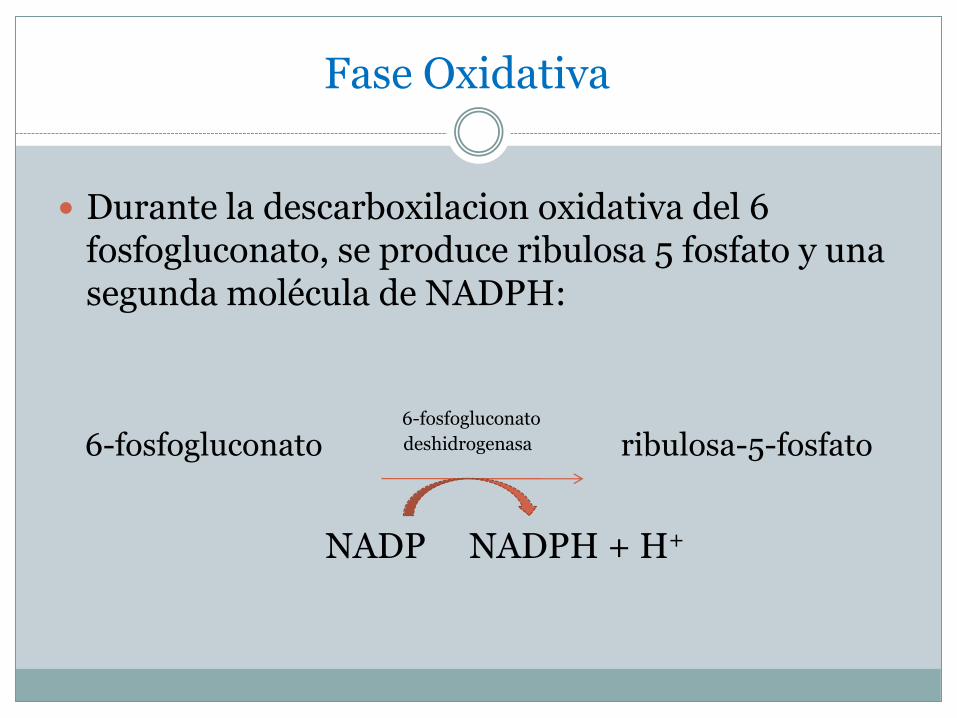
Durante la descarboxilacion oxidativa del 6 fosfogluconato, se produce ribulosa 5 fosfato y una segunda molécula de NADPH:
6-fosfogluconato ribulosa-5-fosfato
NADP NADPH + H+
6-fosfogluconato
deshidrogenasa
Fase Oxidativa

Comienza con la conversión de la ribulosa-5-fosfato en ribosa-5-fosfato por la ribulosa-5-fosfato isomerasa, o en la xilulosa-5-fosfato por la ribulosa-5-fosfato epimerasa
ribosa-5-fosfato
Ribulosa 5-fosfato
xilulosa-5fosfato
ribulosa 5-fosfato
isomerasa
Fase No Oxidativa
ribulosa 5-fosfato
epimerasa

Dos reacciones están catalizadas por transcetolasa:
• En la primera reacción, la enzima transfiere una unidad de dos carbonos desde la xilulosa-5-fosfato a la ribosa-5-fosfato produciendo Gliceraldehido-3-fosfato y sedoheptulosa-7-fosfato.
ribosa-5-fosfato Gliceraldehido-3-fosfato
+ +xilulosa-5fosfato sedoheptulosa-7-
fosfato.
Fase No Oxidativa
Trnscetolasa

En la segunda reacción, una unidad de dos carbonos de otra molécula de xilulosa-5-fosfato se transfiere a la eritrosa-4-fosfato y forman Gliceraldehido-3-fosfato y fructosa-6-fosfato.
eritrosa-4-fosfato Gliceraldehido-3-fosfato
+ +xilulosa-5fosfato Fructosa-6-fosfato.
Fase No Oxidativa
Trnscetolasa

La ruta de las pentosas fosfato está regulada de forma que satisfaga los requerimientos momentáneos de NADPH y ribosa-5-fosfato.
La fase oxidativa es muy activa en células como eritrocitos y hepatocitos, en las que las demandas de NADPH son elevadas.

Estas reacciones proporcionan una cantidad substancial de NADPH que se requiere para los procesos reductores (síntesis de lípidos) y los mecanismos antioxidantes.
Esta ruta es más activa en las células en las que se sintetizan cantidades relativamente grandes de lípidos:
tejido adiposo glándula mamaria
corteza suprarrenal el hígado.