






Idiomas
Páginas
Jurídico


DISEÑO Y SÍNTESIS DE UN PÉPTIDO CON Y SIN
RESTRICCIÓN CONFORMACIONAL DE UNA
PROTEÍNA EXPRESADA POR EL GEN Pb-1 DE Oryza
sativa INVOLUCRADA EN EL MECANISMO DE
DEFENSA CONTRA LA PYRICULARIA
DIANA MILENA GÓMEZ MORENO
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS
Facultad de Ciencias y Educación Proyecto Curricular de Licenciatura en Química
Bogotá, D.C. Colombia
2016

II Título de la tesis o trabajo de investigación
DISEÑO Y SÍNTESIS DE UN PÉPTIDO CON Y SIN
RESTRICCIÓN CONFORMACIONAL DE UNA
PROTEÍNA EXPRESADA POR EL GEN Pb-1 DE Oryza
sativa INVOLUCRADA EN EL MECANISMO DE
DEFENSA CONTRA LA PYRICULARIA
DIANA MILENA GÓMEZ MORENO
Trabajo de grado presentado como requisito parcial para optar al título de:
Licenciado en Química
Director: JULIO CESAR CALVO MOZO Qco., Dr.Sc.
Director del Grupo de Investigación de PROTEOMA UD
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS
Facultad de Ciencias y Educación Proyecto Curricular de Licenciatura en Química
Bogotá, D.C. Colombia
2016

III
NOTA DE ACEPTACIÓN
---------------------------------------------------
---------------------------------------------------
---------------------------------------------------
JURADO
---------------------------------------------------
JURADO
---------------------------------------------------
DIRECTOR
Ciudad y Fecha

IV Título de la tesis o trabajo de investigación
DEDICATORIA
Dedico este trabajo en primer lugar a DIOS por darme salud
y vida para culminar un objetivo y un sueño más, por los triunfos
y los momentos difíciles que me han enseñado a valorarlo cada día más.
A mi mamá María Moreno, por la inmensidad de su amor, por la incansable
solicitud de sus cuidados, por enseñarme paso a paso con sabios consejos y profundos valores
el camino de la vida, y que la fe y la constancia pueden hacer de los sueños una realidad.
A mi papá Humberto Gómez por ser mi guía y ejemplo a seguir, por su apoyo incondicional, amor
invaluable y por enseñarme a creer que “no hay cosas imposibles si no personas incapaces”.
A mi hermanito Nicolás Gómez, por ser mi camarada de aventuras, por compartir memorias de
infancia y sueños de adulto, por ser mi confidente y amigo para toda la vida.
A mi abuelito Darío Moreno, por que espero que en lo más alto del cielo se sienta orgulloso de mí.
A mi novio Fernando Varela por enseñarme a creer que el amor existe y que un esfuerzo genuino
supera mi propia imaginación y a mis profesores gracias por su tiempo, su apoyo y por la
sabiduría que me trasmitieron en el desarrollo de mi formación profesional.
“A todas las manos que me han abierto, a las que he visto y a las que no”
Madre Teresa de Calcula

V
AGRADECIMIENTOS
Durante el desarrollo de este trabajo muchas personas se vieron involucradas por tanto
debo agradecer con gran sinceridad, hacer de este proyecto una realidad.
A DIOS por protegerme, guiarme durante todo el camino y darme fuerzas para sortear
obstáculos y ver con alegría un paso más en mi vida.
A mis queridos padres, por ser la columna vertebral de mi crecimiento personal, espiritual
y profesional. A mi hermanito por ser un ejemplo de amistad, esfuerzo y emprendimiento.
A mi abuelito por darme los concejos necesarios en el momento indicado. A mi novio por
ser un ejemplo a seguir como persona y profesional.
A mi mentor y director de tesis, el profesor Julio Cesar Calvo por guiarme, aconsejarme,
enseñarme y permitirme realizar este proyecto a su lado. Por haber creído en mis
capacidades y compartir con migo su gran conocimiento.
A mi guía en el exterior, la profesora Fanny Guzmán y su grupo de investigación en la
Universidad Pontificia Católica de Valparaíso en Chile, por abrirme sus brazos y las puertas
a un mundo nuevo de conocimiento e investigación.
A mi querida y gran amiga Erika Niño, por compartir con migo una gran aventura de
conocimiento, aprendizaje y vivencias donde un “hacer las cosas con amor” marca la
diferencia y por apoyarme en esta etapa tan valiosa.
Al Centro de Relaciones Interinstitucionales (CERI) y al Centro de Investigaciones y
Desarrollo Científico (CIDC) de la Universidad Distrital por permitirme hacer la pasantía
de investigación en el exterior.
A todos GRACIAS.

VI Título de la tesis o trabajo de investigación
RESUMEN
El hongo Pyricularia Oryzae es el agente causante de la piriculariosis, enfermedad de alto
impacto en la producción de arroz (Oryza sativa) en el mundo, causando graves lesiones en
la planta y pérdida parcial o total del grano. La reacción natural de las plantas a especies
patógenas es de corta duración debido a las condiciones bióticas y abióticas del cultivo; sin
embargo, este mecanismo de defensa ha fomentado el estudio y desarrollo de variedades
resistentes, que se emplean para el control de la enfermedad. Con la visión de aportar a la
ampliación del germoplasma en variedades de resistencia de este cereal, la Federación
Nacional de Arroceros de Colombia (1) y los grupos de investigación Biomolc y Proteoma
UD de la Universidad Distrital Francisco José de Caldas, han estudiado genes que confieren
resistencia a la Pyricularia, como es el caso del Gen Pb-1 de Oryza sativa, el cual expresa
una proteína de resistencia involucrada en el mecanismo de defensa ante esta enfermedad.
En este trabajo se seleccionaron los fragmentos LHEDQYAQILQE y DEEEKDNDDN con
tendencia alfa helical de dicha proteína, se sintetizaron las secuencias lineales y sus
correspondientes análogos con restricción conformacional a alfa hélice siguiendo el modelo
de doble dímero (en proceso de patente). Los resultados por cromatografía Liquida de Alta
eficiencia en fase reversa (RP-HPLC), espectrometría de masas (EM) y dicroísmo circular
(CD) muestran que se pudo obtener los productos propuestos y se demostró la utilidad del
sitio de nucleación JAZ (en proceso de patente) para estabilizar fragmentos de proteínas con
tendencia helical
Palabras clave: Síntesis de Péptidos en fase sólida, Pyricularia, Gen Pb-1 de Oryza sativa,
Restricción conformacional, sitio de nucleación.

VII
Contenido
Pág.
RESUMEN ...................................................................................................................... VI
Lista de Ilustraciones........................................................................................................ IX
Lista de Tablas ................................................................................................................. IX
Lista de Figuras ................................................................................................................ X
Lista de Símbolos y abreviaturas ........................................................................................ 1
Introducción ...................................................................................................................... 3
1. Marco teórico .......................................................................................................... 7 1.1 Generalidades del Arroz ...................................................................................... 8 1.2 Enfermedades del Arroz ...................................................................................... 9 1.3 Genes de Resistencia: variedades resistentes ........................................................ 11 1.4 Síntesis de péptidos en fase sólida ...................................................................... 12
1.4.1 Matrices Poliméricas. ............................................................................. 15 1.4.2 Activadores ........................................................................................... 16 1.4.3 Dificultades Sintéticas ............................................................................ 17 1.4.4 Péptidos estructurados ............................................................................ 17 1.4.5 Formas de Presentación Peptídica ............................................................ 19
2. Metodología .......................................................................................................... 22 2.1 Búsqueda de la estructura secundaria y terciaria de una proteína expresada por el Gen
Pb-1 de Oryza sativa, involucrada en el mecanismo de defensa contra la Pyricularia. ......... 22 2.2 Selección de las estructuras de la proteína expresada por el Gen Pb-1 de Oryza
sativa, candidatas para estudio. ..................................................................................... 22 2.3 Diseño y síntesis de un péptido con y sin restricción conformacional que imite un
fragmento de la proteína nativa. .................................................................................... 22 2.4 Síntesis del péptido con Restricción Conformacional ............................................ 23 2.5 Síntesis del péptido con presentación de Dendrímero de doble constructo (DCC): .... 24 2.6 Desalinización de los péptidos crudos por Sephadex ® G-10: ................................ 24 2.7 Caracterización de los compuestos sintetizados: ................................................... 24
2.7.1 Cromatografía Líquida de Alta Eficiencia en Fase Reversa (RP-HPLC) ....... 24 2.7.2 Espectrometría de Masas ........................................................................ 25 2.7.3 Dicroísmo Circular ................................................................................ 25
3. Resultados y Discusión ...................................................................................... 26 3.1 Búsqueda de la estructura secundaria y terciaria de una proteína expresada por el Gen
Pb-1 de Oryza sativa, involucrada en el mecanismo de defensa contra la Pyricularia. .......... 26 3.2 Selección de las estructuras de la proteína expresada por el Gen Pb-1 de Oryza sativa,
candidatas para estudio. ............................................................................................... 27 3.3 Diseño y síntesis de un péptido con y sin restricción conformacional que imite un
fragmento de la proteína nativa. .................................................................................... 29
4. Conclusiones y recomendaciones ...................................................................... 43

VII
I
Título de la tesis o trabajo de investigación
4.1 Conclusiones ................................................................................................... 43 4.2 Recomendaciones ............................................................................................ 43
A. Anexo 1. Tabla de los 20 aminoácidos naturales ....................................................... 44
B. Anexo 2. Solución de Ninhidrina y Azul de Bromofenol ............................................ 45
C. Anexo 3. Grupos Protectores, Resinas y Brazos Espaciadores ................................... 47
Bibliografía ..................................................................................................................... 48

IX
Lista de Ilustraciones
Pág.
Ilustración 1. Morfología de la planta de arroz. Tomado de: “El cultivo del Arroz (Oryza Sativa
L.)” (15). ........................................................................................................................... 9
Ilustración 2. Ciclo de vida de la Pyricularia Oryzae en la planta de arroz (28) ........................ 10
Ilustración 3. Esquema general de la síntesis de péptidos en fase sólida. .................................. 13
Ilustración 4. Mecanismo de desprotección de t-Boc ............................................................. 14
Ilustración 5. Mecanismo de desprotección de Fmoc ............................................................. 14
Ilustración 6. Mecanismo de acople DCC/HOBt ................................................................... 14
Ilustración 7. Estructura de resinas utilizadas como soporte sólido. ......................................... 15
Ilustración 8. Racemización de del carbono terminal de un aminoácido durante el proceso de
acople, a) enolización y b) formación de oxazolonas. ............................................................ 16
Ilustración 9. Estructuras peptídicas; primaria secundaria y terciaria. ....................................... 18
Ilustración 10. La gráfica muestra espectros de dicroísmo circular en el UV lejano asociados con
diversos tipos de estructuras secundarias. Linea sólida: α-hélice, líneas discontinuas largas: hojas β
....................................................................................................................................... 19
Ilustración 11. Representación de las diferentes especies formadas para la síntesis de un polímero
peptídico de cisteínas, cada flecha representa una unidad peptídica; donde n=1 es el monómero
cíclico, n=2 son las especies diméricas que presentan diferente direccionamiento. Se omite las
moléculas cíclicas del dímero, y moléculas de mayor número de unidades. .............................. 20
Ilustración 12. Presentación de un MAP con cuatro unidades peptídicas incorporadas en los grupos
funcionales del amino de la lisina. ....................................................................................... 20
Ilustración 13.Constructos de Doble Dímero (DDC) los cuales contienen cuatro copias de un
mismo péptido. Tomada de la referencia (58) ....................................................................... 21
Ilustración 14. Inserción de ligandos J`y Z` en una secuencia peptídica para dar alfa-hélice (a.) o
“loop” (b.) Proteoma UD. .................................................................................................. 24
Lista de Tablas
Pág.
Tabla 1. Péptidos lineales y sus correspondientes péptidos restringidos sintetizados. ................. 28
Tabla 2. Tendencia de loa aminoácidos a formar estructuras secundarias ................................. 42

X Título de la tesis o trabajo de investigación
Lista de Figuras
Figura 1. Proteína secuenciada de N—C terminal con 1296 aa, con número de acceso BAJ25849.
(9) ................................................................................................................................... 26
Figura 2. Proteína S210084, modelo 1 C-Score = -1.64 S210084_results.tar.bz2, Swiss-
PdbViewer 4.1.0. a) cadena principal de la proteína b) representación de estructura secundaria en
cinta, lamina-β amarillo, α-hélice rojo c) volumen de la proteína superpuesta a la estructura de
cintas d) probabilidad de acceso de cada residuo al solvente, azul menos accesible hasta el rojo más
expuesto. .......................................................................................................................... 27
Figura 3. Péptido lineal (3) (LHEDQYAQILQE; PM: 1484.71 Da). ........................................ 30
Figura 4. Péptido dendrimérico con restricción (4) (JAZ- LHEDQYAQILQE)2-KGC; PM: 3778.2
Da). a) estructura de la secuencia. b) cromatograma RP-HPLC c) espectro de masas MALDI-TOF
....................................................................................................................................... 31
Figura 5. Dicroísmo circular y deconvolución de: a) péptido lineal (3) b) péptido restringido (4) c)
Grafica superpuesta de cada dicroísmo. ................................................................................ 32
Figura 6. Péptido lineal (5) (DEEEKDNDDN; PM: 1222.1 Da), a) estructura de la secuencia. b)
cromatograma RP-HPLC c) espectro de masas ESI ............................................................... 33
Figura 7. Péptido dendrimérico con restricción (6) (JAZ- DEEEKDNDDN)2-KGC; PM: 3249.18
Da), a) estructura de la secuencia. b) cromatograma RP-HPLC c) espectro de masas ................. 34
Figura 8. Dicroísmo circular y deconvolución de: a) péptido lineal y b) péptido restringido c)
Grafica superpuesta de cada dicroísmo. ................................................................................ 35
Figura 9. Péptido lineal (7) (EDQYAQILQEVF; PM: 1482.62 Da), a) estructura de la secuencia. b)
cromatograma RP-HPLC c) espectro de masas ESI ................................................................ 36
Figura 10. Péptido dendrimérico con restricción (r-1) (JAZ- EDQYAQILQEVF)2-KGC; PM:
3770.22 Da), a) estructura de la secuencia. b) cromatograma RP-HPLC c) espectro de masas
MALDI-TOF .................................................................................................................... 37
Figura 11. Dicroísmo circular y deconvolución de: a) péptido lineal y b) péptido restringido c)
Grafica superpuesta de cada dicroísmo. ................................................................................ 38
Figura 12. Péptido lineal (10) (VEDEEEKDND; PM: 1221.16 Da), a) estructura de la secuencia. b)
cromatograma RP-HPLC c) espectro de masas ESI ............................................................... 39
Figura 13. Péptido dendrimérico con restricción (r-2) (JAZ- VEDEEEKDND)2-KGC PM: 3247.3
Da), a) estructura de la secuencia. b) cromatograma RP-HPLC c) espectro de masas MALDI TOF.
....................................................................................................................................... 40
Figura 14. Dicroísmo circular y deconvolución de: a) péptido lineal y b) péptido restringido c)
Grafica superpuesta de cada dicroísmo. ................................................................................ 41

Marco Teórico
Lista de Símbolos y abreviaturas
abreviatura nombre
1-HOBt 1-Hidroxibenzotriazol
ACN Acetonitrilo
DC Dicroísmo Circular
DCC N,N’-diciclohelxilcarbodiimida
DCM Diclorometano
DDC Contructo de doble dendrimero
DIC Diisopropilcarbodiimida
DIEA N,N´-di-isopropiletilamina
DMF N,N´-dimetilformamida
DOTA 2,2′-(Ethylenedioxy)diethanethiol
1,2-Bis(2-mercaptoethoxy)ethane, 3,6-Dioxa-1,8-octane-dithiol
EDT Etanoditiol
9-Fmoc 9-Fluorenilmetiloxicarbonilo
Fmoc-Ala-OH Fmoc-Alanina-OH
Fmoc-Arg(Pbf)-OH Fmoc-Arginina(Pbf)-OH
Fmoc-Asn(Trt)-OH Fmoc-Asparagina(Trt)-OH
Fmoc-Asp(OtBu)-OH Fmoc-Aspártico(OtBu)-OH
Fmoc-Gln(Trt)-OH Fmoc-Glutamina(Trt)-OH
Fmoc-Glu(OtBu)-OH Fmoc-Glutámico(OtBu)-OH
Fmoc-Gly-OH Fmoc-Glicina-OH
Fmoc-His(Trt)-OH Fmoc-Histidina(Trt)-OH
Fmoc-Ile-OH Fmoc-Isoleucina-OH
Fmoc-Leu-OH Fmoc-Leucina-OH
Fmoc-Lys(t-Boc)-OH Fmoc-Lisina(t-Boc)-OH
Fmoc-Lys(Dde)-OH Fmoc-Lisina(Dde)-OH
Fmoc-Met-OH Fmoc-Metionina-OH
Fmoc-Phe-OH Fmoc-Fenilalanina-OH
Fmoc-Pro-OH Fmoc-Prolina-OH
Fmoc-Ser(tBu)-OH Fmoc-Serina(tBu)-OH
Fmoc-Thr(tBu)-OH Fmoc-Treonina(tBu)-OH
Fmoc-Trp(t-Boc)-OH Fmoc-Triptófano(t-Boc)-OH
Fmoc-Trp-OH Fmoc-Triptófano-OH
Fmoc-Tyr(tBu)-OH Fmoc-Tirosina(tBu)-OH
Fmoc-Val-OH Fmoc-Valina-OH
HBTU
N-((1-H-benzotriazol-1-il)-dimetilamino-metilen)-N-
metilmetanaminio
HOBT 1-Hidroxibenzotriazol
HCCA Ácido a-ciano-4-hidroxicinámico
HF Ácido Fluorhídrico

2
IPA Isopropanol
KCN Cianuro de potasio
MALDI-TOF Matrix Assisted Laser Desorption of Ions Time of Flight
MAPs Multiple Antigen Peptides
MeOH Metanol
NaHCO3 Bicarbonato de sodio
NaOH Hidróxido de sodio
NMP 1-metil-2-pirrolidona
OXIMA Oxima
Reactivo de Ellman Ácido ditionitrobenzoico
Ninhidrina 2,2-Dihidroxiindano-1,3-diona
RP-HPLC Reversed phase High performance liquid chromatography
TBTU
Tetrafluoroborato de O-(1H-Benzotriazol-1-il)-N,N,N',N'-
tetrametiluronio
t-Boc Tert-butoxicarbonilo
TFA Ácido trifluoroacético
TFE 2,2,2-Trifluoroetanol
TIS Triisopropil silano
Tritón X100 Polioxietilenoctilfenileter
TCTU
O-(6-Chlorobenzotriazol-1-yl)-N,N,N′,N′-tetramethyluronium
tetrafluoroborate

3
Introducción
El arroz es uno de los productos fundamentales en la dieta alimenticia de la población
mundial por su bajo costo y alto valor nutricional, por esta razón, se ha incrementado
gradualmente la demanda y siembra de este cereal; así mismo, ha aumentado el esfuerzo por
optimizar su producción y métodos de cultivo. La siembra de esta semilla constituye el
ambiente idóneo para el establecimiento y dispersión de numerosas plagas como bacterias,
hongos y virus; afectando negativamente los arrozales, llegando a constituir su control uno
de los principales problemas económicos en el cultivo del arroz (2) (3).
La enfermedad causada por el hongo Pyricularia Oryzae o quemazón del arroz es de las más
perjudiciales en las regiones arroceras con pérdidas de hasta el 90 % de la producción; su
incidencia aumenta en climas húmedos y lluviosos donde se favorece el cultivo, afecta
directamente a las hojas y panícula de la planta dejando inservible la semilla de arroz (4). El
control químico es uno de los métodos más utilizados para combatir esta enfermedad, sin
embargo, aspectos económicos y medioambientales han disminuido su uso.
Por otro lado, las diferentes prácticas de cultivo, como el control de riego, adecuada
fertilización y elección de una variedad resistente, desempeñan un papel importante en el
control de esta enfermedad (5). Como alternativa de control está la búsqueda de tolerancia
a enfermedades, la cual aumenta la variabilidad genética del cultivo y potencia nuevos
mecanismos que interactúan con los agentes patógenos, denotando la importancia en el
mejoramiento de las plantas (6).
De acuerdo con estas consideraciones los grupos de investigación Biomolc y Proteoma UD
de la Universidad Distrital Francisco José de Caldas, en convenio con la Federación
Nacional de Arroceros de Colombia (FEDEARROZ), propuso emplear la estrategia de
síntesis de péptidos en fase sólida por medio de la química Fmoc para diseñar y sintetizar
secuencias peptídicas perteneciente a la proteína expresada por Gen Pb-1 de la especie
Oryza sativa, el cual se ve involucrado en el mecanismo de reacción contra la Pyricularia,
con vistas a su posible empleo en generar anticuerpos antipéptido que identifiquen dicha
secuencia en un extracto de plantas de arroz.

4
Planteamiento del problema
La demanda mundial de arroz, exige que se obtengan variedades resistentes a enfermedades
como la Pyricularia, uno de los agentes más conocidos causantes del añublo o quemazón
del arroz el cual disminuye la producción, rendimiento y calidad del grano, afectando la
mayor parte de las zonas tropicales productoras en el mundo.
Por tanto, es de vital importancia la identificación de proteínas que expresen resistencia a
esta enfermedad, por medio del estudio sintético de fragmentos que imiten la proteína nativa
cuyo enfoque en caso de ser eficiente aminoraría el tiempo de selección de una variedad
determinada y a su vez costos frente a los estudios convencionales. Por consiguiente es de
importancia evaluar si es posible sintetizar un péptido con restricción conformacional en
una presentación de constructo de doble dímero que imite un fragmento estructurado de la
proteína nativa expresada por el Gen Pb-1 de Oryza sativa involucrada en el mecanismo de
defensa contra la Pyricularia.

5
Justificación
La Universidad Distrital, a través de los grupos Biomolc y Proteoma UD formaron un
convenio con la Federación Nacional de Arroceros de Colombia – FEDEARROZ, para
desarrollar el macro proyecto de Cooperación Internacional COL 5024 financiado por la
Agencia Internacional de Energía Atómica (AIEA), que busca ampliar la variedad en
germoplasma con el fin de contar con herramientas para enfrentar los efectos del cambio
climático global que han incidido en la reaparición y/o ampliación de enfermedades tales
como la Pyricularia y la Hoja Blanca, así como el estrés abiótico producido por la sequía
en algunas regiones arroceras del país.
En este macro proyecto, se trabajan cuatro aspectos: la generación de mutantes por
irradiación, el estudio de genes involucrados en la respuesta al estrés biótico y abiótico, el
análisis proteómico de las proteínas relacionadas con la respuestas de la planta al estrés y el
diseño de vías alternas para identificar dichas proteínas. Por tanto la importancia de este
estudio radicó en la síntesis de una secuencia de una proteína expresada por el Gen Pb-1 de
Oryza sativa involucrada en el mecanismo de defensa contra la Pyricularia, por medio de
un constructo complejo de doble dímero que pueda generar anticuerpos antipéptido en
futuros estudios, para reconocer las proteínas que intervienen en la protección de la planta
ante dicha enfermedad.
Hipótesis
Es posible el diseño y síntesis de un péptido con restricción conformacional, que imite
fragmentos de una proteína expresada por el Gen Pb-1 de Oryza sativa, por medio de la
estrategia de síntesis de péptidos en fase sólida extrapolable en la síntesis de moléculas
dendriméricas tipo DDC.

6
Objetivo General
Diseñar y sintetizar un péptido con y sin restricción conformacional de una proteína
expresada por el GEN Pb-1 de Oryza sativa, involucrada en el mecanismo de defensa contra
la Pyricularia.
Objetivos Específicos
- Encontrar la estructura secundaria y terciaria de una de las proteínas expresadas por
el Gen Pb-1 de Oryza sativa, involucrada en el mecanismo de defensa contra la
Pyricularia.
- Seleccionar los fragmentos en la proteína expresada por el Gen Pb-1 de Oryza
sativa, candidatos para el estudio.
- Diseñar y sintetizar un péptido con y sin restricción conformacional que imite un
fragmento de la proteína nativa.
- Caracterizar el producto obtenido.

Marco Teórico
1. Marco teórico
Antecedentes: El arroz es el nombre común para una especie (Oryza sativa) de la familia de
las gramíneas, este cereal es uno de los alimentos más consumidos y el sustento principal
de más de la mitad de la población mundial. Desde tiempos milenarios este grano fue el
alimento primordial en los países asiáticos y hoy es cultivado en los cinco continentes en
diferentes entornos climáticos, ya sea templado, cálido o húmedo, adaptándose a su vez a
todo tipo de terreno seco; estas condiciones han contribuido a la enorme variedad de semillas
de arroz que existen en los distintos mercados del mundo. La evolución en los métodos de
siembra ha provocado alteraciones físicas y biológicas en la planta de arroz generando
cambios que se reflejan en los mecanismos de defensa contra gran variedad de plagas y
enfermedades (7).
De las enfermedades que padece el arroz, la más sobresaliente es la Pyricularia Oryzae,
originada por un hongo microscópico perteneciente a la familia Magnaporthe Oryzae. El
micelio del hongo produce una sustancia tóxica conocida como pericularina la cual inhibe
el crecimiento de los tejidos y los desorganiza, causando un crecimiento parcial o nulo del
grano (8). Por tal razón, organismos internacionales de investigación como la IRRI
(International Rice Research Institute, en Asia), WARDA (West Africa Rice Development
Association, en África), CIAT (International Center for Tropical Agriculture, en América)
y FEDEARROZ (Federación Nacional de Arroceros, en Colombia) estudian y desarrollan
especies genéticamente resistentes a esta enfermedad (1).
En estudios previos se ha caracterizado variedades de este cereal (Oryzae sativa) las cuales
expresan un gen de resistencia durante el mecanismo de reacción frente al hongo del añublo
o quemazón de arroz, denominado gen Pb-1; este gen proveniente del Panicle-Blast-1 se
caracteriza por la durabilidad en el mecanismo de acción contra la enfermedad y ha sido
introducido en variedades de alta calidad para cultivo como gen clonado (9), (10).
FEDEARROZ en convenio con el grupo de investigación de Biología Molecular
(BIOMOLC) de la Universidad Distrital ha estudiado proteínas derivadas de este gen
empleando herramientas bioinformáticas, para seleccionar fragmentos proteicos que
presentan resistencia ante dicha enfermedad (11). Como ejemplo, el uso de estas
herramientas ha permitido determinar zonas proteicas con actividad biológica en contra del
virus de la hoja blanca, otra enfermedad que afecta agresivamente a los cultivos de arroz,
seleccionando regiones con estructuras secundarias definidas, con características
hidrofílicas y ubicadas en la parte externa de la proteína, porque son más accesibles y poseen
alta probabilidad de generar una interacción anticuerpo-antipéptido.

8 Trabajo de investigación
Estas moléculas fueron sintetizadas por medio de la estrategia de Síntesis Química de
Péptidos con y sin restricción conformacional siguiendo el modelo de síntesis
macromolecular por el grupo de PROTEOMA UD (12).
1.1 Generalidades del Arroz
La semilla de arroz proviene de la planta Oryza sativa, considerado como el cereal de mayor
consumo en la población mundial, se cultivan alrededor de 150 millones de hectáreas
anualmente para una producción aproximada de 745 millones de toneladas (13), con un
incremento proyectado del 70% para el año 2025 (14). Los grandes productores de este
grano son: África occidental, Asia suroriental (China, Indonesia, Tailandia y Laos) y
América del sur (Brasil, Colombia, Argentina, Chile, Perú, Venezuela) (15), donde el
principal productor y consumidor es China, con una cifra estimada de 185 millones de
toneladas por año (31% de la producción mundial) (16).
En Suramérica se siembran cerca de 6,7 millones de hectáreas de arroz considerado uno de
los principales sectores en la economía. Colombia ocupa el puesto 24 en el mercado mundial
en producción arrocera, el cual alcanza una producción nacional de 1,5 millones de toneladas
anuales en un área aproximada de 320 mil hectáreas; las principales zonas productoras son
la región caribe, región andina y llanos orientales (17).
Este cereal aporta hasta un 40% del suministro de energía alimenticia según la Organización
de las Naciones Unidas para la Alimentación y la Agricultura (FAO, por sus sigla en inglés,
Food and Agriculture Organization) (18) a su vez es excelente fuente nutricional de
carbohidratos (76%), proteínas (6.5%), fibra (1.9%), agua (12%), minerales (zinc, magnesio
y fosforo) y vitaminas (2%) (tiamina y riboflavina) (19).
Es de crecimiento rápido y gran capacidad productiva, adaptable a diversas condiciones de
clima tropical y suelos; su temperatura óptima de germinación oscila entre 27 y 35 °C. Es
una planta monocotiledónea que proviene de la familia graminácea Oryza sativa, de raíces
fibrosas y delgadas, tallo erguido y cilíndrico, con una altura (30-150 cm) que depende de
las condiciones de cultivo (Ilustración 1) (4). Los tallos se componen de hojas alternas,
largas y planas, las cuales se ramifican en panículas de 20 a 30 cm, cada panícula consta de
entre 50 y 300 espiguillas o flores de las cuales se forma el fruto obtenido en una cariópside
(7).
El manejo de los arrozales es una actividad dinámica donde generalmente se emplean tres
métodos para el cultivo del arroz; trasplante, secano y riego, en los dos últimos la siembra
puede ser mecanizada o por voleo, donde el agricultor esparce manualmente las semillas
(20). El tipo de siembra aumenta o disminuye los niveles nutricionales del arroz, no obstante
el cultivo implementado depende de las facilidades económicas del productor, la variedad
agronómica del grano y el área a cultivar. La siembra por trasplante es un método indirecto
que traspasa la plántula del semillero en el cual germinó al campo definitivo donde se va a
desarrollar, la ventaja es el uso de poca semilla por área de siembra, sin embargo se necesita
de mucha mano de obra lo que influye en altos costos y poco empleo del método (21). El

9
cultivo secano es una técnica más implementada, en la cual se requiere de un suelo fértil,
con alto contenido en minerales, poco uso de productos agroquímicos (3) y empleo
exclusivo de aguas lluvias las cuales pueden ser canalizadas proporcionando un suelo
semihumedo; sin embargo es necesario esperar las lluvias o un riego previo para no afectar
el cultivo, lo que genera mayor inversión en el rubro de las labores.
Ilustración 1. Morfología de la planta de arroz. Tomado de: “El cultivo del Arroz (Oryza Sativa L.)” (15).
Por otro lado, el método de riego requiere una inundación continua lo que favorece el
contenido en minerales, alta presencia de aluminio y la siembra del grano en cualquier época
del año, el rendimiento en este de cultivo es mayor ya que las condiciones le permiten
minimizar la predisposición a enfermedades y no presentan ningún tipo de estrés hídrico lo
que facilita la asimilación de los productos (22). Sin embargo, el arroz como cualquier otra
planta está expuesto a una gran variedad de agentes patógenos que inciden durante todas las
etapas de desarrollo, la mayoría de estos problemas fitosanitarios en el arroz se presentan
en el cultivo secano, favorecido por el ambiente y el manejo del mismo dañando
drásticamente el proceso de labranza al ser el principal obstáculo en la producción arrocera
(21). En Colombia coexisten estos dos últimos sistemas de cultivo mecanizado, siendo el de
riego el más utilizado sin embargo, regiones como los llanos orientales implementan el
cultivo secano favoreciendo la presencia de estas enfermedades (20).
1.2 Enfermedades del Arroz
El arroz es atacado por diversas enfermedades desde el inicio de la siembra hasta poco
antes de la cosecha, las pérdidas que causan son severas, pueden reducir el rendimiento a
valores insignificantes o llevar a la pérdida casi total del cultivo (15). Las enfermedades

10 Trabajo de investigación
van cambiando de acuerdo a la etapa de cultivo, sistema de labranza, densidad de la
siembra, manejo de agua, nutrición, entre otras (23), como consecuencia se genera un
mayor consumo en insumos agroquímicos para su tratamiento, lo que implica un aumento
en los costos de producción que serán trasladados al consumidor final (24). Las
enfermedades de mayor impacto que afectan a nivel mundial el cultivo de arroz son, el
Añublo del Arroz (Pyricularia Oryzae), el Virus de la Hoja Blanca, y el Quemazón de la
Vaina, las cuales pueden ocasionar importantes reducciones en el rendimiento, muerte de
las plantas o influir en la perdida de la calidad del grano (22).
La Pyricularia Oryzae o Añublo del Arroz es de las enfermedades más importantes del
cultivo de arroz por su poder destructivo, esta es ocasionada por el hongo Pyricularia
Grisea u Oryzae (25) el cual produce una sustancia tóxica conocida como pericularina que
inhibe el crecimiento de los tejidos y los desorganiza causando el añublo o quemazón del
cereal (26) los síntomas se ven reflejados principalmente en hojas, tallo y panícula, aunque
puede afectar todas las partes aéreas de la planta visualizadas por medio de puntos color
café verdoso los cuales pueden ser de diferentes tamaños (1-1.5 cm diámetro) según las
condiciones ambientales de la planta, la consecuencia principal de la enfermedad es la
pérdida parcial o total de la panícula, cosechando granos vacíos o parcialmente formados
(27).
Ilustración 2. Ciclo de vida de la Pyricularia Oryzae en la planta de arroz (28)
La reproducción del hongo Pyricularia es asexual y se propaga por medio de esporas
(Ilustración 2), trasmitidas por aire y agua las cuales se posan en nuevas plantas iniciando el
proceso de infección, la espora germinada libera un tubo angosto el cual se adhiere a la
epidermis de la planta, en el extremo del tubo se forma un apresorio u órgano de fijación
cuya función principal es perforar la cutícula y liberar el patógeno (29). La espora se ubica

11
en los nodos de los tallos, hojas y panícula, siempre y cuando las condiciones del cultivo,
como alta humedad relativa (90-92%), temperatura (22-29 °C), concentración de nitrógeno
en agua, nubosidad, presencia de vientos e intensidad lumínica sean favorables (30). Cuando
la espora penetra el vegetal se producen filamentos que obstruyen el sistema vascular de la
planta interrumpiendo la circulación de la savia hacia el tallo y/o panícula, al cabo de 4 o 5
días termina secándose y tornando el color café característico, si la espora llega a afectar la
semilla puede generarse un nuevo foco de enfermedad (24), (31).
Por tanto, es de vital importancia para el agricultor contrarrestar este tipo de enfermedades
y emplear métodos adecuados e integrales que contribuyan al mejoramiento de las defensas
naturales de la planta de arroz, como implementar el riego a bajas temperaturas, disminuir
el contenido de nitrógeno en el abono, purgar el terreno de cosechas previamente infectadas,
y cortar de inmediato las malas hierbas presentes en el sembrado, de igual forma el empleo
de fungicidas también hace parte de la protección de la cosecha siempre y cuando sea
proporcional y adecuado a la especie sembrada, sin ocasionar daño al ambiente (24). Por
consiguiente, una de las prioridades en investigación agrícola es ofrecer a los productores
arroceros variedades resistentes a las enfermedades que se le presentan al momento de
cultivar, en este caso la quemazón de la panícula, lo cual puede representar un control
adecuado (26).
1.3 Genes de Resistencia: variedades resistentes
Las plantas generalmente se defienden de los patógenos de dos formas: 1) características
estructurales que actúan como barreras físicas e inhiben la entrada del patógeno y la
dispersión a través de la planta; 2) reacciones bioquímicas que tienen lugar en las células y
tejidos de la planta que producen sustancias que pueden ser tóxicas para el patógeno (32).
La combinación de estas dos formas de defensa son diferentes para cada sistema hospedero-
patógeno y varían dependiendo de la edad, clase de órgano de la planta, tejido atacado,
condición nutricional y climática (33), con este concepto se busca mejorar la calidad y
rendimiento del cultivo de arroz (34).
Un estudio realizado por el grupo de Takatsuji et al., muestra que los genes de resistencia
que se expresan en la planta de arroz al ser atacada por la Pyricularia son propensos a decaer
debido a la variabilidad patogénica del hongo (9). Sin embargo el gen Pb-1 (Panicle blast
1) derivado de la especie de arroz “indica cultivar-Modan” se caracteriza por la durabilidad
y resistencia ante el hongo (35), otros estudios muestran que la resistencia parcial de la planta
se asocia a la variedad poligénica, aun así el periodo de latencia no contrarresta lo suficiente
la estadía del patógeno, un conjunto de diversos factores ambientales como humedad,
temperatura, tipo de suelo, fertilización, entre otros, pueden influir en la expresión de la
resistencia parcial del gen, ahora bien, en un entorno propicio es posible aplicar una ayuda
a la resistencia de la planta hospedante ante el patógeno (36); Por esta razón, se seleccionó
una proteína de resistencia expresada por este gen, y de esta se seleccionaron fragmentos
peptídicos para ser evaluados y sintetizados en esta investigación.

12 Trabajo de investigación
1.4 Síntesis de péptidos en fase sólida
La síntesis química de péptidos nació en 1901 con la síntesis de dipéptidos desarrollados por
Emil Fischer, posteriormente los investigadores Max Bergmann y Leonidas Zervas en 1932
diseñaron los primeros grupos protectores de los grupos amino (carbobenzoxilo) y de las
cadenas laterales de los aminoácidos, en 1953 el científico Vincent Du Vigneaud sintetizó
por primera vez un péptido funcional, la oxitocina, motivo por el cual fue galardonado con
el premio nobel de química en 1956, un logro importante poco tiempo después fue el diseño
de la síntesis en fase sólida por Bruce Merrifield, por cuyo desarrollo e impacto también le
fue concedido el Premio Nobel de Química en 1984. (37) Esta metodología de síntesis
pronto adquirió gran relevancia y con los avances tecnológicos se empezaron a desarrollar
nuevos soportes sólidos, espaciadores, grupos protectores y estrategias de activación para la
formación del enlace peptídico (38).
La metodología de síntesis de péptidos en fase sólida (SPPS, solid phase peptide synthesis),
consiste en la adición secuencial de alfa-aminoácidos, protegidos en la cadena lateral y su
alfa amino, a un soporte polimérico insoluble que es químicamente inerte y mecánicamente
estable a las condiciones de síntesis (39); esta estrategia sintética permite emplear excesos
del aminoácido y demás reactivos que se eliminan fácilmente por los procesos sencillos de
lavado y filtrado lo cual facilita el proceso de síntesis en comparación a la síntesis en fase
líquida (40) (Ilustración 3). Los grupos protectores acido-lábil t-Boc (tert-butiloxicarbonilo)
o básico-lábil Fmoc (9-fluorenilmetoxicarbonilo) son los más empleados para la protección
alfa-amino y los grupos Benzil (Bzl) y tert-Butil (tBu) respectivamente para la protección
de las cadenas laterales de los aminoácidos que lo requieran (41). El crecimiento de la cadena
peptídica inicia por el extremo carboxílico del aminoácido protegido, el cual se ancla a la
matriz polimérica por medio de un brazo o espaciador funcionalizado que permite la unión,
una vez acoplado el aminoácido se remueve el grupo protector alfa-amino por medio de un
proceso de desprotección (42).
La desprotección del grupo t-Boc y Fmoc se realiza con ácido trifluoroacético (TFA)
(Ilustración 4) y piperidina o piperazina (Ilustración 5), respectivamente; de esta forma el
grupo alfa-amino se encuentra libre y permite la unión al siguiente aminoácido el cual es
previamente activado para formar el enlace peptídico. El proceso de acople (Formación del
enlace amida) requiere la activación del grupo carboxilo del aminoácido entrante usando un
agente de acople de modo que reaccione con el amino terminal del péptido en construcción
(Ilustración 6) (43).
El acople y la desprotección se repite tantas veces como sea necesario hasta completar la
secuencia deseada (38), finalizada la síntesis se remueven los grupos protectores de las
cadenas laterales de los aminoácidos y se desancla el péptido del soporte sólido teniendo en
cuenta la estrategia de síntesis implementada (41). En la estrategia t-Boc se emplea ácido
fuerte como el ácido fluorhídrico (HF) o ácido trifluorometanelsulfonico (TFMSA) y para

13
la estrategia Fmoc se utiliza TFA; en cada uno de estas estrategias químicas se emplea un
coctel de compuestos que dependen de la secuencia peptídica y son adicionados para evitar
o disminuir la formación de reacciones colaterales que son propias del proceso de desanclaje
del péptido (44), (45).
Espaciador
Espaciador
Espaciador
Espaciador
Espaciador
Espaciador
+.
O
NH.OH
R1
P1
T
.
O
NH.X
R1
T
P1
.
O
NH2
X
R1
P1
.
O
NHX
R1
.
O
NH.
R2
P1
.
O
NH.A
R2
T
P2
T
P2
.
O
NHX
R1
.
O
NH
R2
.
O
NH
Rn.
.
O
NH.
Rn+1
P1
P2
Pn
n
T
Pn+1
O
NHX
R1O
NH
R2O
NH
Rn.O
NH2
Rn+1
n
Acople del primer aminoácido a la resina
Remover el grupo protector del -amino
Acople del siguiente aminoácido activado
n ciclos de acople y desprotección
Remoción de todos los grupos protectores y desanclaje del péptido de la resina
Ciclos SPPS
P Grupo protector de la cadena lateral de los aminoácidos
T Grupo protector del -amino de los aminoacidos
Soporte sólido
A Grupo activante
X NH o O
Ilustración 3. Esquema general de la síntesis de péptidos en fase sólida.
14 Trabajo de investigación
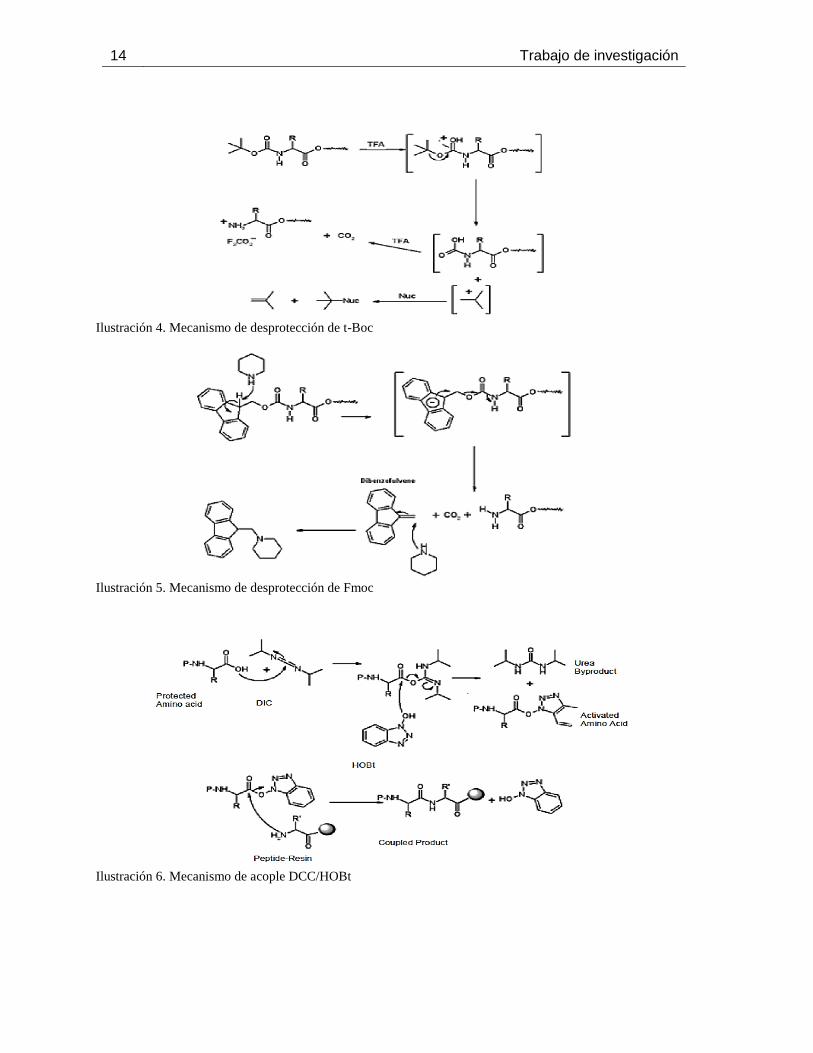
Ilustración 4. Mecanismo de desprotección de t-Boc
Ilustración 5. Mecanismo de desprotección de Fmoc
Ilustración 6. Mecanismo de acople DCC/HOBt

15
1.4.1 Matrices Poliméricas.
La síntesis química de péptidos ha sido la base principal para la preparación de estas
moléculas por más de 50 años, el óptimo resultado de la síntesis depende de la adecuada
elección del soporte sólido, el brazo espaciador “linker”, grupos protectores de los
aminoácidos, metodología de acoplamiento y protocolo de desanclaje del péptido de la
resina. Las características de un buen soporte sólido deben ser; estabilidad física y química
a las condiciones de síntesis, una alta capacidad de hinchamiento en los diferentes solventes
empleados para permitir accesibilidad a los sitios de reacción de los diferentes reactivos,
tener un grado de entrecruzamiento adecuado y de funcionalización aceptable, ser
compatible con los reactivos, longitud y secuencia del péptido deseado. Para la síntesis de
péptidos es favorable el uso de resinas con bajo grado de entrecruzamiento y gran capacidad
de hinchamiento, puesto que la cinética de reacción controlada por la velocidad de difusión,
permite a una resina más hinchada tener tiempos de reacción más cortos y acomodar mejor
a la cadena peptídica en crecimiento (46).
Los soportes basados en poliestiréno, tienen el historial más largo de uso en síntesis de
péptidos en fase sólida, tienen una capacidad de carga significativamente alta y poseen entre
el 1% y 2% de divinilbenceno (DVB) como agente de reticulación; de igual forma otras
matrices con núcleos que incluyen poliacrilato, poliacrilamida y polietilenglicol con igual
rango de reticulación, han sido utilizadas para la síntesis de “péptidos difíciles” (propensos
a agregación), soportan elevadas temperaturas, son insolubles en solventes orgánicos y
solvatadas e hinchadas en solventes apróticos tales como tolueno, dimetil formamida,
diclorometano y 1-metil-2-pirrolidona, sin embargo la capacidad de hinchamiento varia con
respecto al solvente que se expone (45)
Los linkers o brazos funcionalizados, son moléculas que sirven para unir de forma reversible
la cadena peptídica en crecimiento a la resina, a la vez que determinan el grupo funcional y
sirven de grupo protector del C-terminal. La mayoría de los linkers son escindidos por
acidólisis en diferentes concentraciones dependiendo la estrategia química empleada (46)
Ilustración 7. Estructura de resinas utilizadas como soporte sólido.

16 Trabajo de investigación
1.4.2 Activadores
El acople del primer aminoácido a la resina o la formación de enlace amida entre el
aminoácido que ingresa y uno previamente acoplado, requiere la activación del aminoácido
entrante. Esta activación del ácido carboxílico es la base del enlace peptídico, cuya función
principal es formar el acople y mantener la integridad del péptido en crecimiento en especial
el centro estereogénico del carbono alfa del aminoácido. Para esta reacción es necesario
recurrir a agentes de acople, los cuales permiten aumentar la cinética de la reacción puesto
que proporcionan al carbono carbonilo un buen grupo saliente lo que favorece el acople, sin
embargo, también se ve favorecida la formación de oxazolonas y enoles, que son las dos
vias de racemización debidas al aumento de la acidez del protón del carbono alfa el cual es
fácilmente retirado (Ilustración 8) (41).
Ilustración 8. Racemización de del carbono terminal de un aminoácido durante el proceso de acople, a)
enolización y b) formación de oxazolonas.
Los reactivos de acople (activadores) más utilizados se pueden dividir en dos grandes
grupos, las carbodiimidas y triazoles; Las carbodiimidas son los reactivos de acoplamiento
más utilizados en fase sólida, la N´N-diciclohexilcarbodiimida (DCC) fue primera en ser
empleada y hoy día sigue siendo la más popular, sin embargo, la formación del subproducto
diciclohexilurea interviene negativamente en el rendimiento final y a su vez es un potente
alérgeno (47).
Por tal motivo se ha estudiado nuevos reactivos que minimizan la formación de
subproductos no deseados a la vez que disminuyen los niveles de racemización. La mayoría
de estos compuestos son benzotriazoles de las cuales se derivan las sales de fosfonio y
uronio, que en presencia de una base pueden convertir el carboxilato del aminoácido entrante
en una especie activada. (48), Las más empleadas son el Hexafluorofosfato de benzotriazol-
1-il-N-oxi-tris (pirrolidin) fosfonio (PyBOB) derivadas del HOBt. Entre las sales de uronio
se encuentran el hexafluorofosfato de N-oxido de N-((1-H-benzotriazol-1-il)-dimetilamino-
metilen)-N-metilmetanaminio (HBTU), y el tetrafluoroborato de N-oxido de N-((1-H-
benzotriazol-1-il)-dimetilamino-metilen)-N-metilmetanaminio (TBTU) (48).

17
1.4.3 Dificultades Sintéticas
A pesar de los avances en la síntesis química de péptidos en fase sólida y la optimización
del proceso, existen secuencias peptídicas que presentan dificultades sintéticas,
denominados péptidos de secuencia difícil; estas dificultades sintéticas son más frecuentes
en algunos péptidos que otros y ha sido descrito como el problema potencial más serio en la
síntesis de péptidos (49). Estos inconvenientes se presentan en secuencias con alto contenido
de un solo aminoácido, aminoácidos estéricamente impedidos por causa de cadenas laterales
o grupos protectores voluminosos, problemas de hinchamiento de la resina, baja solvatación
del complejo resina-péptido y las interacciones entre aminoácidos que inducen al
plegamiento del péptido, provocando agregaciones inter e intramoleculares de la cadena
peptídica reflejado en la formación de estructuras tipo hojas-β (50), afectando de manera
parcial o total las reacciones de desprotección y acilación; en algunos casos extremos el
complejo péptido-resina se vuelve insoluble y no está disponible para reaccionar.
Las estrategias para aumentar la eficiencia sintética disminuyendo la formación de
agregados y estructuras secundarias se basa en la modificación del entorno químico y la
incorporación de grupos protectores reversibles en la estructura del péptido en construcción.
Todas estas aproximaciones buscan el mejoramiento de la solvatación del complejo resina-
péptido, y así minimizar las dificultades sintéticas. Dentro de estos desarrollos se encuentra
el uso de resinas de composición química especifica tipo polientilenglicol (PEG) o
polietilenglicolacrilamida (PEGA) las cuales permiten mejorar la eficiencia sintética debido
a su gran capacidad de hinchamiento, sumado a esto el uso de resinas de baja sustitución
que disminuye el impedimento estérico entre cadenas peptídicas adyacentes. El uso de
solventes y de activadores que aumentan la velocidad de reacción, sales y detergentes
caotrópicos también han mostrado una exitosa disminución de los fenómenos de agregación
durante el acople de los aminoácidos, detergentes no iónicos, sonicación, aumento de
temperatura (microondas) y procesos de neutralización In-Situ (51).
1.4.4 Péptidos estructurados
Debido a la relación entre secuencia primaria, conformación molecular y función biológica,
estudios como el de Anfinsen permitieron demostrar que la estructura tridimensional de una
proteína está directamente ligada a la secuencia de aminoácidos, puesto que el orden de
nucleótidos en el ADN especifica la secuencia primaria, es decir que un gen contiene toda
la información necesaria para determinar la estructura tridimensional de su producto
proteico (52).
La síntesis de fragmentos peptídicos ha permitido el estudio de moléculas que imiten las
estructuras nativas de las proteínas las cuales están dominadas por los ángulos de torsión
que permiten conformaciones especificas junto con factores externos como enlaces de

18 Trabajo de investigación
hidrógeno e interacciones hidrofóbicas; la estructura hélice alfa es una de ellas, tiene los
ángulos torsionales phi (φ) y psi (ψ) toman valores cercanos a -57° y -47°, respectivamente.
Estas hélices tienen 3,6 residuos por vuelta y por cada aminoácido se produce un incremento
de la hélice de 1,5 Å (traslación por residuo), se caracteriza por que adopta una forma
helicoidal gracias a los puentes de hidrógeno de la cadena principal, los aminoácidos tienen
la cadena lateral orientada hacia el exterior. Para formar esta estructura el grupo carboxilo
de cada aminoácido (n) se une mediante un puente de hidrógeno al grupo amino de otro
aminoácido (n+4), es una de las más estables por que da lugar a un gran número de
interacciones. Otra conformación es la hoja-beta, la cual se forma con enlaces de hidrógeno
entre dos cadenas peptídicas vecinas creando láminas en zig-zag, la disposición puede ser
paralela o antiparalela, puede darse entre dos regiones próximas o distantes del péptido , los
grupos radicales de cada aminoácido sobresalen de la lámina en ambos sentidos, de forma
alterna. También se encuentran en este grupo, los giros β, los cuales se forman por la unión
mediante un puente de hidrógeno del aminoácido n + n+3, también llamados “bucle” o
“loop” (53).
Ilustración 9. Estructuras peptídicas; primaria secundaria y terciaria.
En algunos caso es posible obtener en fragmentos peptídicos estructuras similares a las
encontradas en la proteínas nativas en otros no es posible debido a que la proteína ayuda a
19
estabilizar dichas estructuras, en otros casos es posible el empleo de restricciones
conformacionales que ayudan a estabilizarla (54)
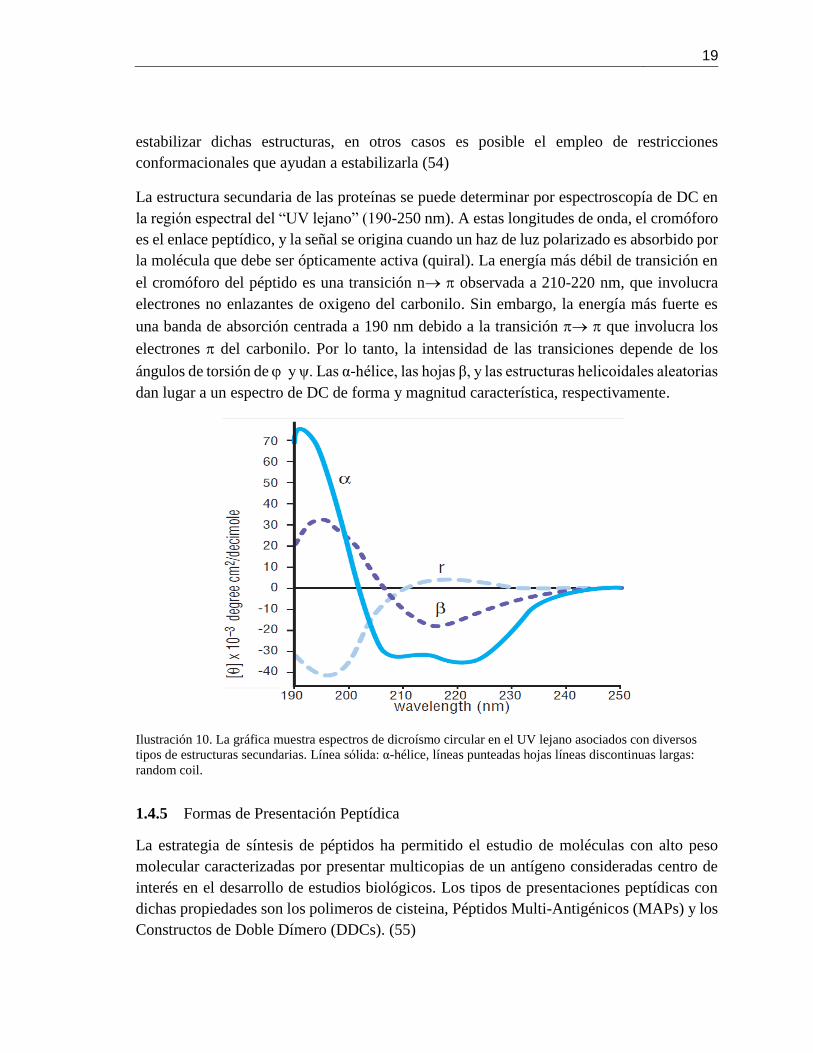
La estructura secundaria de las proteínas se puede determinar por espectroscopía de DC en
la región espectral del “UV lejano” (190-250 nm). A estas longitudes de onda, el cromóforo
es el enlace peptídico, y la señal se origina cuando un haz de luz polarizado es absorbido por
la molécula que debe ser ópticamente activa (quiral). La energía más débil de transición en
el cromóforo del péptido es una transición n observada a 210-220 nm, que involucra
electrones no enlazantes de oxigeno del carbonilo. Sin embargo, la energía más fuerte es
una banda de absorción centrada a 190 nm debido a la transición que involucra los
electrones del carbonilo. Por lo tanto, la intensidad de las transiciones depende de los
ángulos de torsión de y ψ. Las α-hélice, las hojas β, y las estructuras helicoidales aleatorias
dan lugar a un espectro de DC de forma y magnitud característica, respectivamente.
Ilustración 10. La gráfica muestra espectros de dicroísmo circular en el UV lejano asociados con diversos
tipos de estructuras secundarias. Línea sólida: α-hélice, líneas punteadas hojas líneas discontinuas largas:
random coil.
1.4.5 Formas de Presentación Peptídica
La estrategia de síntesis de péptidos ha permitido el estudio de moléculas con alto peso
molecular caracterizadas por presentar multicopias de un antígeno consideradas centro de
interés en el desarrollo de estudios biológicos. Los tipos de presentaciones peptídicas con
dichas propiedades son los polimeros de cisteina, Péptidos Multi-Antigénicos (MAPs) y los
Constructos de Doble Dímero (DDCs). (55)

20 Trabajo de investigación
Los polímeros de cisteína son sintetizados a partir de unidades peptídicas que contienen una
cisteína en el C y N-terminal las cuales son oxidadas por medio del grupo tiol de cada
cisteína dando un enlace disulfuro, que puede unir varias unidades peptídicas, pero esta
primera aproximación es de difícil caracterización debido a las múltiples especies
poliméricas que se pueden formar por medio del grupo tiol y a la falta de direccionamiento
en la formación del enlace disulfuro (Figura 3);
n=1 n=2
Oxidación
C-terminalN-terminal
Ilustración 11. Representación de las diferentes especies formadas para la síntesis de un polímero peptídico de
cisteínas, cada flecha representa una unidad peptídica; donde n=1 es el monómero cíclico, n=2 son las especies
diméricas que presentan diferente direccionamiento. Se omite las moléculas cíclicas del dímero, y moléculas
de mayor número de unidades.
Los MAPs son estructuras que contienen una lisina la cual permite sintetizar los epítopes
por el grupo α y ε amino (Figura 4). Estas estructuras consisten en unidades ramificadas y
una superficie de grupos funcionales donde puede ser sintetizadas (síntesis divergente) o
ancladas unidades peptídicas previamente sintetizadas (síntesis convergente), obteniéndose
moléculas una sola especie; sin embargo, la principal desventaja son los pasos inherentes de
purificación y síntesis (56).
Péptido NH
LYSPéptido NH NH
LYSNHLYSPéptido NH
Péptido NH
Ilustración 12. Presentación de un MAP con cuatro unidades peptídicas incorporadas en los grupos
funcionales del amino de la lisina.
Los DDCs son moléculas con un core de glicina y cisteína, utilizando como punto de partida
para la síntesis de los péptidos los grupos amino α y ε del aminoácido lisina (55), Su
construcción presenta varias ventajas sobre la síntesis de MAPs como lo es una disminución

21
en el impedimento estérico al sintetizar sobre una sola lisina, lo que permite obtener mejores
rendimientos y perfiles cromatográficos, en la obtención de las moléculas sintetizadas.
Finalizada la síntesis se realiza la formación de un puente disulfuro entre dos moléculas
peptídicas por medio del grupo tiol de la cisteína, para producir una plataforma que contiene
cuatro copias del mismo antígeno (57) .
Debido a su tamaño y tipo de estructura esta macromolécula tiene como característica
principal, presentar un antígeno de interés en forma tetramérica en la misma molécula
(Figura9) (58).
Ilustración 13.Constructos de Doble Dímero (DDC) los cuales contienen cuatro copias de un mismo péptido.
Tomada de la referencia (57) (58)

22 Trabajo de investigación
2. Metodología
Este proyecto fue desarrollado con los objetivos propuestos de forma secuencial.
2.1 Búsqueda de la estructura secundaria y terciaria de una proteína expresada por el Gen
Pb-1 de Oryza sativa, involucrada en el mecanismo de defensa contra la Pyricularia.
Mediante el uso de software se buscó un modelo tridimensional de la proteína seleccionada,
el cual aportó datos sobre las estructuras secundarias que se encuentran en la proteína con
lo cual se eligieron los péptidos sintetizados
2.2 Selección de las estructuras de la proteína expresada por el Gen Pb-1 de Oryza sativa,
candidatas para estudio.
A partir de los resultados obtenidos se eligieron las estructuras en función a su estructura
secundaria, hidrofobicidad y ubicación en la proteína, y si la secuencia tenía probabilidad
de alfa hélice se analizaban sus propiedades con el programa Helical Wheel.
2.3 Diseño y síntesis de un péptido con y sin restricción conformacional que imite un
fragmento de la proteína nativa.
Una vez se seleccionó la secuencia se procedió a la síntesis del péptido de C a N terminal
mediante la metodología Fmoc, siguiendo el protocolo que se describe a continuación
0,040 g de resina Fmoc Rink Amida con sustitución 0,59 meq/g, fueron introducidos en una
bolsa de polipropileno con dimensiones de 15 x20 mm, se codificó numéricamente para
identificar la bolsa y así permitir realizar síntesis simultanea de péptidos. La bolsa fue
introducida en el recipiente de reacción, donde se hinchó la resina con 3 mL de
Diclorometano (DCM) (2 x5´), esta matriz polimérica posee un amino protegido con el
grupo Fmoc, el cual fue removido para poder realizar el acople del primer residuo de
aminoácido; (59).
Para esto se adicionó 2 mL de una solución de Piperidina al 20% en Dimetilformamida
(DMF) (2x10´), una vez terminado se lavó con DMF(3X1´), Isopropanol (IPA) (1X1´),
DMF (2X1´) y DCM (2X1´) para retirar remanentes del reactivo y dejar la resina en
condiciones para el acople si es requerido.
Se monitoreó la remoción del grupo protector Fmoc mediante dos reacciones colorimétricas
que determinan la presencia de grupos aminos libres. En el primer método se tomó una
pequeña porción de resina en un ependorf y se adiciono una solución de ninhidrina (3 gotas
de la solución A y 1 gota de la solución B, (ver anexo 2), se calentó por 5 min a 100°C; El
segundo ensayo consistió en adicionar a una porción de resina 5 gotas de la solución de azul
de bromofenol al 1% en DMF, se agito y se observó pasados 3 minutos de reacción; Si el
resultado en las dos pruebas muestran tanto la resina como la solución azul o verdosa indica

23
un ensayo positivo para aminas libres, y se puede proceder a realizar el acople, de lo
contrario (solución y resina incoloras) el ensayo es negativo y se repite el procedimiento de
desprotección.
Seguido a este paso se realizó el acople del primer aminoácido, asi;
5 equivalentes del aminoácido a acoplar (con respecto a la carga de la resina) , 5 equivalentes
del agente de acople (HBTU, TBTU, TCTU, OXIMA, HATU o DIC) y 10 equivalentes de
base N,N-di-isopropiletilenamina (DIPEA) fueron disueltos en 1ml de DMF y adicionados
a la resina, se dejó reaccionar por 1 hora en agitación constante a temperatura ambiente,
después se drenó la solución y se lavó la resina con DMF (3X1´) y DCM (2X1´). Para
verificar que la acilación haya sido efectiva se realizó un ensayo de ninhidrina, si el resultado
es negativo se procede a desproteger el grupo Fmoc del N-terminal como se describe
anteriormente, de lo contrario se repite este paso de reacción variando el agente de acople
hasta obtener la incorporación total del aminoácido. Sucesivamente se realizaron ciclos de
acople y desprotección de acuerdo al número de residuos en la secuencia.
Una vez sintetizada la secuencia se removió el último grupo Fmoc y se procedió a retirar los
grupos protectores de las cadenas laterales de los aminoácidos, en simultaneo con el
desanclaje del péptido de la matriz con una solución de TFA/TIS/H2O (ácido
trifluoroacético/ triisopropilsilano/ agua) en proporciones 95:2,5:2,5 respectivamente para
secuencias que no contienen, Metionina, Cisteina ni Triptofano, en caso contrario se empleó
una solución de TFA/TIS/DOTA/H2O (ácido trifluoroacético/ triisopropilsilanol/ 2,2′-
(etilendioxy)dietanetiol /agua) en proporciones 94:2:1:2 respectivamente, empleandose en
los dos casos un volumen de 2 ml de solución de clivaje por cada 50 mg de resina con un
tiempo de reacción de 3 horas, luego se adiciono éter etílico frio a la solución para obtener
un precipitado el cual fue disuelto, liofilizado y posteriormente analizado por cromatografía
líquida de alta eficiencia en fase reversa y Espectrometría de Masas y dicroísmo Circular.
2.4 Síntesis del péptido con Restricción Conformacional
Para la síntesis del péptido con restricción alfa-helical o “loop” se siguió el diseño propuesto
por el grupo de investigación Proteoma UD. (En proceso de Patente) (60)
El fragmento peptídico con tendencia α-hélice, se sintetizó primero la secuencia sobre la
resina (2.2) y al final se introdujo los ligandos J` y Z` como sitio de nucleación dejando entre
ellos un aminoácido X (X: cualquier aminoácido); si se estabiliza como bucle “loop” se
ubican los ligandos en los extremos de la secuencia; J` como ligando N-terminal y Z` como
ligando C-terminal. Por último se cicla la secuencia llevando a cabo la restricción
conformacional. (En proceso de Patente). (61).

24 Trabajo de investigación
Ilustración 14. Inserción de ligandos J` y Z` en una secuencia peptídica para dar alfa-hélice (a.) o “loop” (b.)
Proteoma UD.
2.5 Síntesis del péptido con presentación de Dendrímero de doble constructo (DCC):
Para la obtención del DDC (tetrámero), se sintetizó inicialmente un core de lisina KGC, y
se continuó con la construcción de las cadenas peptídicas en el α y ε amino de la Lisina,
siguiendo con la metodología presentada en el numeral 2.3 y 2.4. Una vez sintetizadas se
realizó un enlace disulfuro con las cisteínas del C-terminal entre las dos moléculas
diméricas. La reacción se llevó a cabo con una concentración de 10mg/ml en una solución
acuosa de dimetilsulfóxido (DMSO) al 10%, a pH 7,5 (62).
2.6 Desalinización de los péptidos crudos por Sephadex ® G-10:
Los crudos obtenidos fueron desalinizados en una columna cromatográfica Sephadex G 10,
hidratada con agua ultra pura, esta permite la separación de sus moléculas en función a su
tamaño. 10mg de péptido se disolvieron en el menor volumen posible (<500 μl), se pasó a
través de la columna y se recogieron los primeros 4ml de elusión con agua, en donde debería
eluir por completo el péptido.
2.7 Caracterización de los compuestos sintetizados:
2.7.1 Cromatografía Líquida de Alta Eficiencia en Fase Reversa (RP-HPLC)
Las especies obtenidas fueron analizadas por RP-HPLC en una columna (C-18) Water
Corp.XBridgeTM BEH130 equilibrada con acetronitrilo/agua/ácido trifluoroacético. 1mg del
péptido fue disuelto en 1 ml de agua ultra pura (1mg/ml) de allí se tomó una alícuota de 5
uL y se llevó a un volumen de 100 uL (Cf 0.05mg/ml). Se analizó con un gradiente de 0-70
% (v/v) de acetonitrilo en agua con 0,05 % de ácido trifluoroacético, en 10 minutos de
elución a un flujo de 1 ml/mín y la absorbancia medida a 220 nm.
Resina J`XZ` SECUENCIA Resina J` SECUENCIA Z`
J`XZ` SECUENCIA J` SECUENCIA Z`
a. b.
Ciclación

25
2.7.2 Espectrometría de Masas
Para la determinación del peso molecular del péptido se utiliza un espectrómetro Microflex
MALDI-TOF (Bruker Daltonics Inc. MA-USA). Se mezclan 1 µL de una solución matriz
de ácido α-ciano-4-hidroxicinnámico ((HCCA) preparada con una concentración del
10mg/ml en acetonitrilo 50% v/v y ácido fórmico 0.1% v/v) y 1 µL de una solución del
péptido (1mg/mL); se siembra esta mezcla en una placa metálica micro scout, se seca y se
introduce en el espectrómetro para realizar el análisis.
2.7.3 Dicroísmo Circular
1mg del péptido fue disuelto en 1 ml de agua ultra pura (1mg/ml) de allí se tomó una alícuota
de 35 uL y se mezcló con 215 uL de 2,2,2 trifluoroetanol (TFE) al 30 % (v/v) para estabilizar
la estructura del péptido favoreciendo la formación de puentes de hidrógeno intramolecular
y evitando que el péptido reaccione con el agua alterando la estructura, para un volumen
final de 250 ul (Cf 0.14mg/ml).
El espectro CD se obtuvo a 20° C en una celda con un paso óptico de 0,1 cm en un espectro
entre 190 y 250 nm de longitud de onda utilizando un espectrómetro de dicroísmo circular
Jasco (J-815 Jasco Corporation, Japan) con una señal promedio sobre 2 segundos por cada
intervalo de 0,5 nm. Todos los datos de dicroísmo circular fueron expresados como valores
de elipticidad molar.

26 Trabajo de investigación
3. Resultados y Discusión
3.1 Búsqueda de la estructura secundaria y terciaria de una proteína expresada por el Gen Pb-
1 de Oryza sativa, involucrada en el mecanismo de defensa contra la Pyricularia.
Se realizó un estudio bioinformático de la proteína expresada por del gen de resistencia Pb-
1 con el fin de analizar las estructuras secundarias y terciarias de esta proteína, lo cual
permitió la selección de fragmentos de interés para la presente investigación.
Por medio de la base de datos del centro nacional para la información biotecnológica, NCBI
(National Center for Biotechnology Information) se identificó la secuencia completa de la
proteína de resistencia (Grupo Oryza Sativa Japónica), reportada en la literatura (9) y en
estudios previos hechos en la Universidad Distrital (11) luego se filtró información con el
programa de alineamientos de secuencias de tipo local BLAST (Basic Local Alignment
Search Tool) y se obtuvo los siguientes datos.
Es una proteína de resistencia que se compone de 1296 aminoácidos, proveniente del Gen
Pb-1 de la variedad de arroz Oryza sativa Japónica, y esta es la proteína secuenciada (Figura
1)
“1 mtelasgavs sllvvirnea allggvrddv qfikeemesm nsflahlaks apqggehneq 61 vrtwmnqvrl laqdcnncid lylysgnpei hrakgrlrrh lwwaywylrk miarhraavq
121 lcqlkdrard vgerrlrygv evpattkaaa pdaasgyaag ddedeedyeh qlavataghh
181 sarravydpp aledyvnekl lewaeeiptr aiqtlsiaiv apdtdnnevl alaheilvap
241 gyyyrrsimv nvpavhakfl plrpkevlyy ilrelehgeg agsqkqatdq gnpwqgyyni
301 yrdkkkvlrk ikrniekmni ykklekiesd ikheqqksdk qlllqlqkkg vdqvdlhvll
361 qllllqsqqd qvknkaadif klpewndnni mkiarklkkh metdeetkev qieveeettk
421 qgrrgkrged ekqkekgeer keverhkeke eetkeeeqqh kkqeekgeer ekverhekek
481 eeetkeehee kkeregreve deeekdnddn ndgeeeeeee eeddddedpi hlhedqyaqi
541 lqevfpkian sktqqqdkse akqvtktatt tldeerikqm ineakedvfr elrggkpdkn
601 qatcepdvpp dknqatgeha gvmdhngeay fteieqkide ikqefkkqlk ikgivdkikh
661 hlieykikhe lpdecpliil kfddmmdgsr weefrkalsl lecsadalif ttenteqakr
721 ysypprepid yslvglyhyt vleltskhkn edncspkifr dileeceghe fcmkifthal
781 yanpkrsnee lsklhstlqd sqksfdaiak kmfmysyndl pkeyksclly laifpkgqki
841 rrstligrwv aegltlkedw pssvrqanrc fdalirrylv cpvdiggtgk vkscavhdpv
901 hgfittiark qhivetrlsh hlarhfsifn dlrlrssdgi htffqslsrs srvsllkvld
961 legsqcfgvk nqrylkdics kmlllkylsl kkteitqlps einclrelev ldiretkvpa
1021 natvnvlllk lkrllaghid ssprnsgtsv qiphridkmv nievlsnvka qrrddledig
1081 klwqlrklgv vvddkrghlg nllkaisnlh ecirsltiti sttthkdtps npelpdhigs
1141 dlphpkkles lsisgarhlf plliksdnnk lakvtlsstp lnqddlevla klpklqcvrl
1201 qhiscivsel ifkeenfkcl kylliegfnl tnitfedgsa celekmvlss tsiesisgvd
1261 vlpkfkelel nnshvpnevk eavennkrii lkcnep”
Figura 1. Proteína secuenciada de N—C terminal con 1296 aa, con número de acceso BAJ25849. (9)

27
A partir de la secuencia seleccionada se buscó con el programa I-Tasser (Protein Structre
and Function Prediction) (63) la estructura secundaria y terciaria de la proteína de estudio,
el resultado se observa en la figura 2. La secuencia tiene alta presencia en estructuras tipo
hojas beta, sin embargo se encuentran en el interior de la proteína evitando la exposición al
medio, mientras que la proporción de alfa-hélices es menor, pero se ubican en su mayoría
en el exterior permitiendo la accesibilidad del solvente representado en la figura 3-d con el
color verde amarillo. a) b)
Figura 2. Proteína S210084, modelo 1 C-Score = -1.64 S210084_results.tar.bz2, Swiss-PdbViewer 4.1.0. a)
cadena principal de la proteína y la representación de estructura secundaria en cinta, lamina-β amarillo, α-
hélice rojo b) probabilidad de acceso de cada residuo al solvente, azul menos accesible hasta el rojo más
expuesto.
3.2 Selección de las estructuras de la proteína expresada por el Gen Pb-1 de Oryza sativa,
candidatas para estudio.
Con la información obtenida a partir del análisis bioinformático se seleccionaron fragmentos
de regiones anfipáticas y expuestas ya que permiten la accesibilidad e interacción con el
solvente. Sin embargo, para aumentar la posibilidad de generar respuesta antígeno-
anticuerpo, se debe no solo tener en cuenta la estructura primaria, sino en lo posible
fragmentos peptídicos con estructura conformacional definida (estructura secundaria),
puesto que son frecuentemente encontrados en interfaces de proteínas y desempeñan un
papel importante en la interacción receptor-ligando, por este motivo los fragmentos elegidos
presentan características alfa-helical. Estos fueron los péptidos seleccionados.

28 Trabajo de investigación
Tabla 1. Péptidos lineales y sus correspondientes péptidos restringidos sintetizados.
Formula estructural N. Secuencia P. M. Representación helicoidal
(Helical Wheel)1
Primera síntesis
3 LHEDQYAQILQE 1486.61
4 (JAZ- LHEDQYAQILQE)2-KGC 3778.2
5 DEEEKDNDDN 1222.1
6 (JAZ- DEEEKDNDDN)2-KGC 3249.18
Segunda síntesis
1 1Convenciones:
Circulo ---------hidrófilico---------rojo Diamante-------hidrofóbico-------verde al amarillo
Triangulo-------negativo----------azul
Pentágono------positivo-----------azul

29
7 EDQYAQILQEVF 1482.62
r-1 (JAZ- EDQYAQILQEVF)2-KGC 3770.22
10 VEDEEEKDND 1221.16
r-2 (JAZ- VEDEEEKDND)2-KGC 3247.3
3.3 Diseño y síntesis de un péptido con y sin restricción conformacional que imite un
fragmento de la proteína nativa.
Estos fueron los datos obtenidos de las secuencias anteriormente estudiadas, se sintetizaron
por medio de la estrategia química Fmoc de síntesis de péptidos en fase sólida, fueron
caracterizados por RP-HPLC y espectrometría de masas EM, y la conformación de dichos
péptidos fue determinada por medio de Dicroísmo Circular (DC). Todas las secuencias
sintetizadas se encuentran en una región externa de la proteína como se visualiza en la
gráfica de la formula estructural de la tabla uno.
30 Trabajo de investigación
PRIMERA SINTESIS
- (3) LHEDQYAQILQE
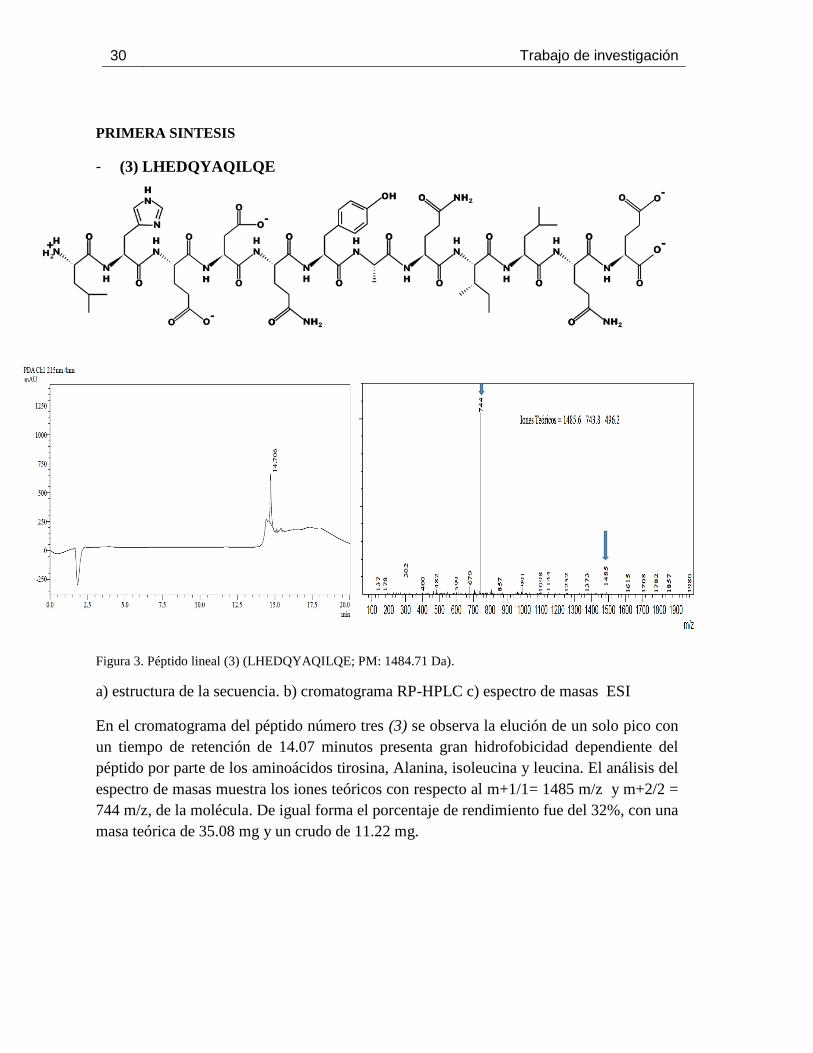
Figura 3. Péptido lineal (3) (LHEDQYAQILQE; PM: 1484.71 Da).
a) estructura de la secuencia. b) cromatograma RP-HPLC c) espectro de masas ESI
En el cromatograma del péptido número tres (3) se observa la elución de un solo pico con
un tiempo de retención de 14.07 minutos presenta gran hidrofobicidad dependiente del
péptido por parte de los aminoácidos tirosina, Alanina, isoleucina y leucina. El análisis del
espectro de masas muestra los iones teóricos con respecto al m+1/1= 1485 m/z y m+2/2 =
744 m/z, de la molécula. De igual forma el porcentaje de rendimiento fue del 32%, con una
masa teórica de 35.08 mg y un crudo de 11.22 mg.

31
- (4) (JAZ- LHEDQYAQILQE)2-KGC
Figura 4. Péptido dendrimérico con restricción (4) (JAZ- LHEDQYAQILQE)2-KGC; PM: 3778.2 Da). a)
estructura de la secuencia. b) cromatograma RP-HPLC c) espectro de masas MALDI-TOF
El péptido número cuatro (4) muestra en el cromatograma la elución de un solo pico con
mayor intensidad sin embargo, el tiempo de retención de 4.39 minutos es menor al lineal,
debido a que aumentan los aminoácidos hidrofílicos en la secuencia como el core de lisina,
aun así la pureza disminuye por la misma amplitud del pico y esto se corrobora con el
análisis del espectro de masas, que muestra la masa de péptido 3778.2 Da, más un aducto
(+23 m/z) que posiblemente es sodio (Na+). De igual forma el porcentaje de rendimiento
fue menor a lo que se esperaba, 6.2%, con una masa teórica de 89.16 mg y un crudo de 5.52
mg
109876543210
1.200
1.100
1.000
900
800
700
600
500
400
300
200
100
0
-100
-200
1,2
25
4,3
92
RT [min]
2480 (154)-281015.DATAmV

32 Trabajo de investigación
Figura 5. Dicroísmo circular y deconvolución de: a) péptido lineal (3) b) péptido restringido (4) c) Grafica
superpuesta de cada dicroísmo.
a) La grafica de dicroísmo del péptido lineal (izquierda) muestra una fuerte tendencia a hoja-
β con un máximo en longitud de onda 190 nm que corresponde a 41723,6 elipticidad molar
y un mínimo marcado en 218 nm con –15562.3 de elipticidad molar por la presencia de
aminoácidos como tirosina e isoleucina los cuales estabilizan este tipo de estructura
secundaria, aunque se encuentren aminoácidos como leucina, glutamina y ácido glutámico
los cuales favorecen la estructura alfa-helical. b) En cuanto al péptido dimérico (derecha) se
observa un cambio en el espectro puesto que disminuye drásticamente la tendencia de hoja
beta conservando el máximo y mínimo característico y aparece un leve porcentaje de α-
hélice, confirmando posiblemente que el sitio de nucleación estabiliza fragmentos con
tendencia alfa-helical. c) En esta última grafica se observa la superposición del espectro
lineal con el dimérico, mostrando la diferencia representativa en estructura conformacional.
-20000
50000
0
20000
40000
190 250200 220 240
Mol. Ellip.
Wavelength [nm]

33
- (5) DEEEKDNDDN
Figura 6. Péptido lineal (5) (DEEEKDNDDN; PM: 1222.1 Da), a) estructura de la secuencia. b)
cromatograma RP-HPLC c) espectro de masas ESI
El péptido lineal número cinco (5) muestra la elución de un solo pico con un tiempo de
retención de 14.55 minutos, con gran hidrofobicidad, a pesar de tener aminoácidos con
carácter hidrofílico como ácido aspártico, ácido glutámico y asparagina. El análisis del
espectro de masas muestra los iones teóricos con respecto al m+1/1= 1221 Da y m+2/2 =
611 Da, de la molécula sintetizada. Así mismo el porcentaje de rendimiento fue del 37%,
con una masa teórica de 28.84 mg y un crudo de 10.84 mg.
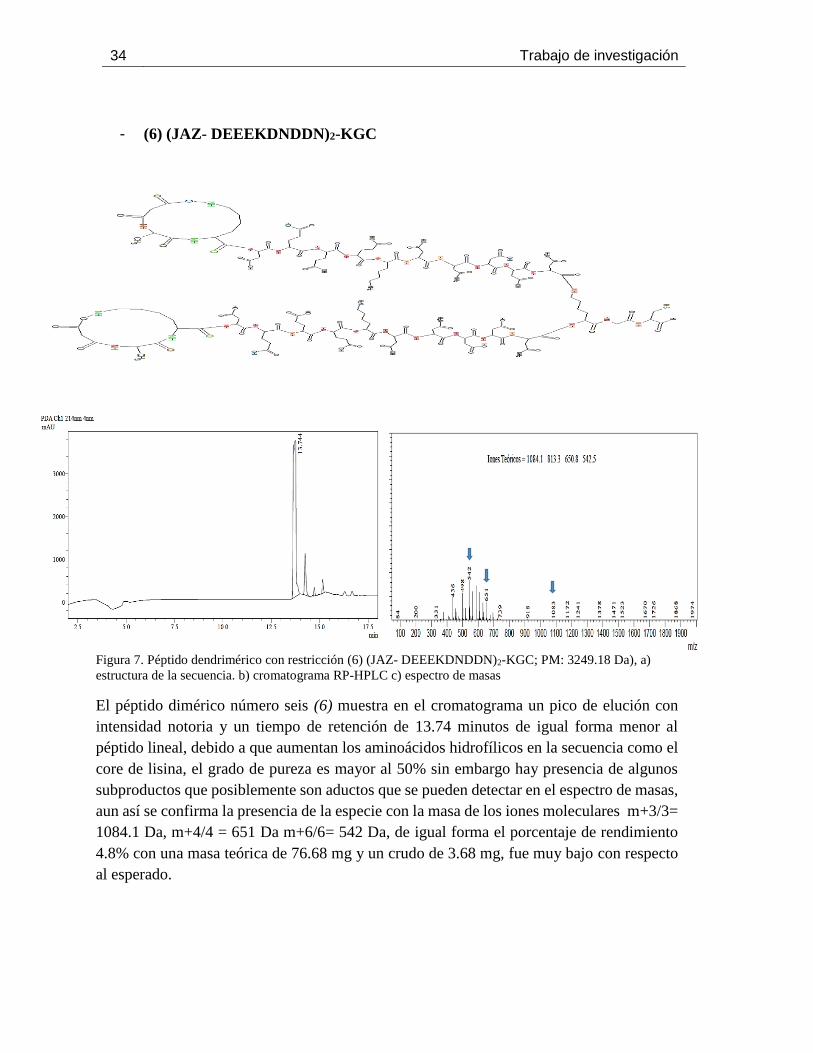
34 Trabajo de investigación
- (6) (JAZ- DEEEKDNDDN)2-KGC
Figura 7. Péptido dendrimérico con restricción (6) (JAZ- DEEEKDNDDN)2-KGC; PM: 3249.18 Da), a)
estructura de la secuencia. b) cromatograma RP-HPLC c) espectro de masas
El péptido dimérico número seis (6) muestra en el cromatograma un pico de elución con
intensidad notoria y un tiempo de retención de 13.74 minutos de igual forma menor al
péptido lineal, debido a que aumentan los aminoácidos hidrofílicos en la secuencia como el
core de lisina, el grado de pureza es mayor al 50% sin embargo hay presencia de algunos
subproductos que posiblemente son aductos que se pueden detectar en el espectro de masas,
aun así se confirma la presencia de la especie con la masa de los iones moleculares m+3/3=
1084.1 Da, m+4/4 = 651 Da m+6/6= 542 Da, de igual forma el porcentaje de rendimiento
4.8% con una masa teórica de 76.68 mg y un crudo de 3.68 mg, fue muy bajo con respecto
al esperado.

35
Figura 8. Dicroísmo circular y deconvolución de: a) péptido lineal y b) péptido restringido c) Grafica
superpuesta de cada dicroísmo.
La grafica de dicroísmo del péptido lineal (izquierda) muestra una tendencia random coil
con un mínimo en 199 nm y con -10427, 1 en elipticidad molar, aunque se esperaba con la
presencia de la secuencia de poliglutámicos una conformación más helical. En cuanto al
péptido dimérico (derecha) se observa un aumento pronunciado de una estructura
determinada, lo cual indica que el punto de nucleación ejerce un posible cambio en la
conformación nativa del fragmento, sin embargo la estructura formada es de tipo hoja-beta
indicados con un mínimo en 217 nm con -17410, 2 de elipticidad molar y un máximo en
190 nm con 21840, 2 elipticidad molar, aun así tiene un leve porcentaje del 16, 9% de alfa-
hélice ya que el máximo de la hoja beta esta en 196 nm y hay alta presencia de glutámicos
que favorecen la estructura alfa-helical sin embargo la conformación del péptido tiende a
estabilizarlo en hoja beta. c) En esta última grafica se observa la superposición del espectro
-20000
30000
-10000
0
10000
20000
190 250200 220 240
Mol. Ellip.
Wavelength [nm]
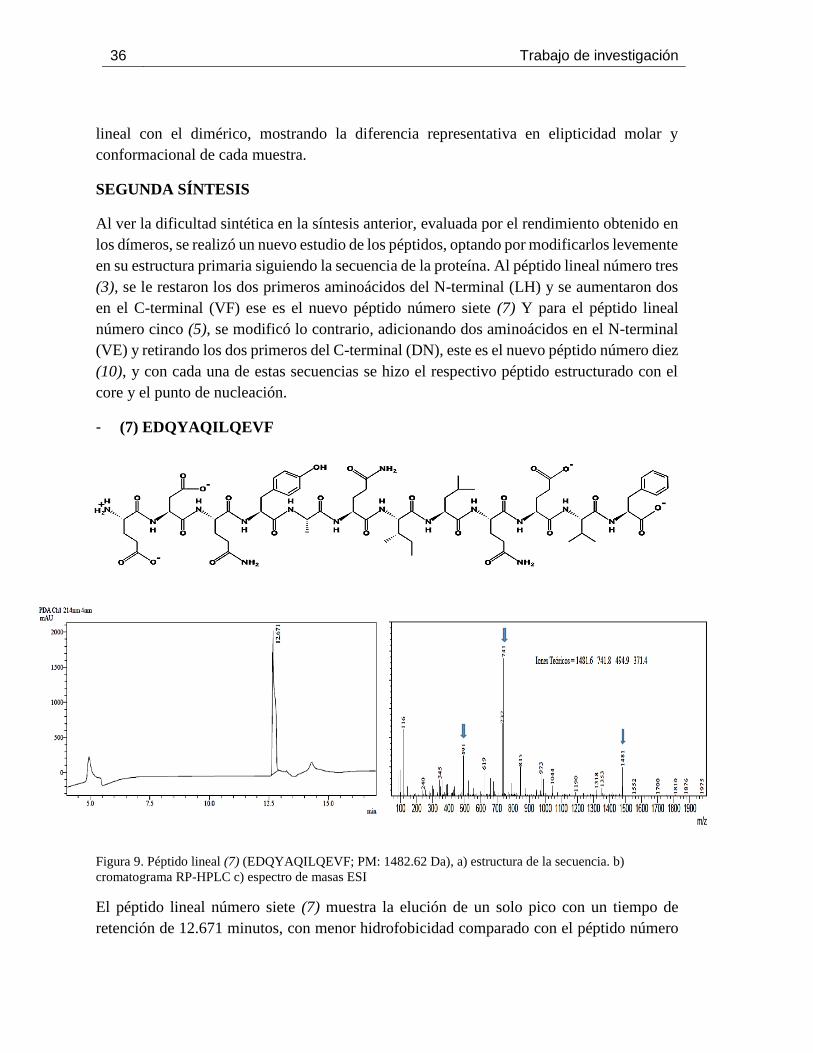
36 Trabajo de investigación
lineal con el dimérico, mostrando la diferencia representativa en elipticidad molar y
conformacional de cada muestra.
SEGUNDA SÍNTESIS
Al ver la dificultad sintética en la síntesis anterior, evaluada por el rendimiento obtenido en
los dímeros, se realizó un nuevo estudio de los péptidos, optando por modificarlos levemente
en su estructura primaria siguiendo la secuencia de la proteína. Al péptido lineal número tres
(3), se le restaron los dos primeros aminoácidos del N-terminal (LH) y se aumentaron dos
en el C-terminal (VF) ese es el nuevo péptido número siete (7) Y para el péptido lineal
número cinco (5), se modificó lo contrario, adicionando dos aminoácidos en el N-terminal
(VE) y retirando los dos primeros del C-terminal (DN), este es el nuevo péptido número diez
(10), y con cada una de estas secuencias se hizo el respectivo péptido estructurado con el
core y el punto de nucleación.
- (7) EDQYAQILQEVF
Figura 9. Péptido lineal (7) (EDQYAQILQEVF; PM: 1482.62 Da), a) estructura de la secuencia. b)
cromatograma RP-HPLC c) espectro de masas ESI
El péptido lineal número siete (7) muestra la elución de un solo pico con un tiempo de
retención de 12.671 minutos, con menor hidrofobicidad comparado con el péptido número
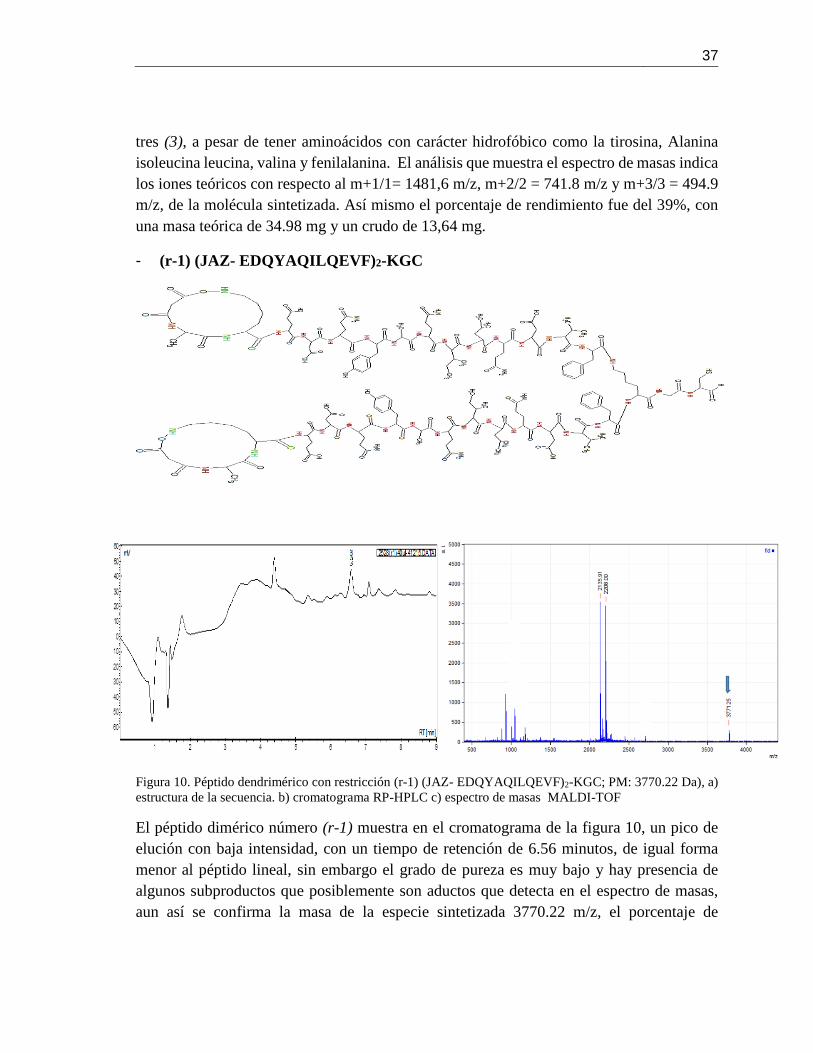
37
tres (3), a pesar de tener aminoácidos con carácter hidrofóbico como la tirosina, Alanina
isoleucina leucina, valina y fenilalanina. El análisis que muestra el espectro de masas indica
los iones teóricos con respecto al m+1/1= 1481,6 m/z, m+2/2 = 741.8 m/z y m+3/3 = 494.9
m/z, de la molécula sintetizada. Así mismo el porcentaje de rendimiento fue del 39%, con
una masa teórica de 34.98 mg y un crudo de 13,64 mg.
- (r-1) (JAZ- EDQYAQILQEVF)2-KGC
Figura 10. Péptido dendrimérico con restricción (r-1) (JAZ- EDQYAQILQEVF)2-KGC; PM: 3770.22 Da), a)
estructura de la secuencia. b) cromatograma RP-HPLC c) espectro de masas MALDI-TOF
El péptido dimérico número (r-1) muestra en el cromatograma de la figura 10, un pico de
elución con baja intensidad, con un tiempo de retención de 6.56 minutos, de igual forma
menor al péptido lineal, sin embargo el grado de pureza es muy bajo y hay presencia de
algunos subproductos que posiblemente son aductos que detecta en el espectro de masas,
aun así se confirma la masa de la especie sintetizada 3770.22 m/z, el porcentaje de

38 Trabajo de investigación
rendimiento 9% es un poco mayor al péptido número cuatro, con una masa teórica de 76.68
mg y un crudo de 3.68 mg, fue muy bajo con respecto al esperado.
Figura 11. Dicroísmo circular y deconvolución de: a) péptido lineal y b) péptido restringido c) Grafica
superpuesta de cada dicroísmo.
La grafica de dicroísmo del péptido lineal (izquierda) muestra una fuerte tendencia a hoja-
β con un máximo en 190 nm y un mínimo marcado en 218 nm por la presencia de
aminoácidos como Tirosina e Isoleucina y el aumento de la Valina y la Fenilalanina los
cuales estabilizan este tipo de estructura secundaria, aunque se encuentren aminoácidos
como leucina, glutamina y ácido glutámico los cuales favorecen la estructura alfa-helical. b)
En cuanto al péptido dimérico (derecha) se observa un cambio notorio en la conformación

39
de la estructura nativa estabilizando la tendencia alfa –helical, confirmando los datos en el
dicroísmo con un máximo de 192 nm y dos mínimos en 209 nm y 221 nm de longitud de
onda verificando una vez más que posiblemente el sitio de nucleación estabiliza e induce
fragmentos con tendencia alfa-helical. c) En esta última grafica se observa la superposición
del espectro lineal con el dimérico, mostrando la diferencia representativa en elipticidad
molar y conformacional de cada muestra.
- (10) VEDEEEKDND
Figura 12. Péptido lineal (10) (VEDEEEKDND; PM: 1221.16 Da), a) estructura de la secuencia. b)
cromatograma RP-HPLC c) espectro de masas ESI
El péptido lineal número diez (10) muestra la elución de un solo pico con un tiempo de
retención de 10.916 minutos el cual indica una pureza alta, con menor hidrofobicidad
comparado con el lineal número cinco (5), debido a los aminoácidos con carácter hidrofílico
como ácido aspártico, ácido glutámico y asparagina que la componen. El análisis que
muestra el espectro de masas indica los iones teóricos con respecto al m+1/1= 1220,5 m/z y
m+2/2 = 611.3 m/z, de la molécula sintetizada. Así mismo el porcentaje de rendimiento fue
mayor que el lineal número (5) del 41%, con una masa teórica de 28.81 mg y un crudo de
11,8 mg.
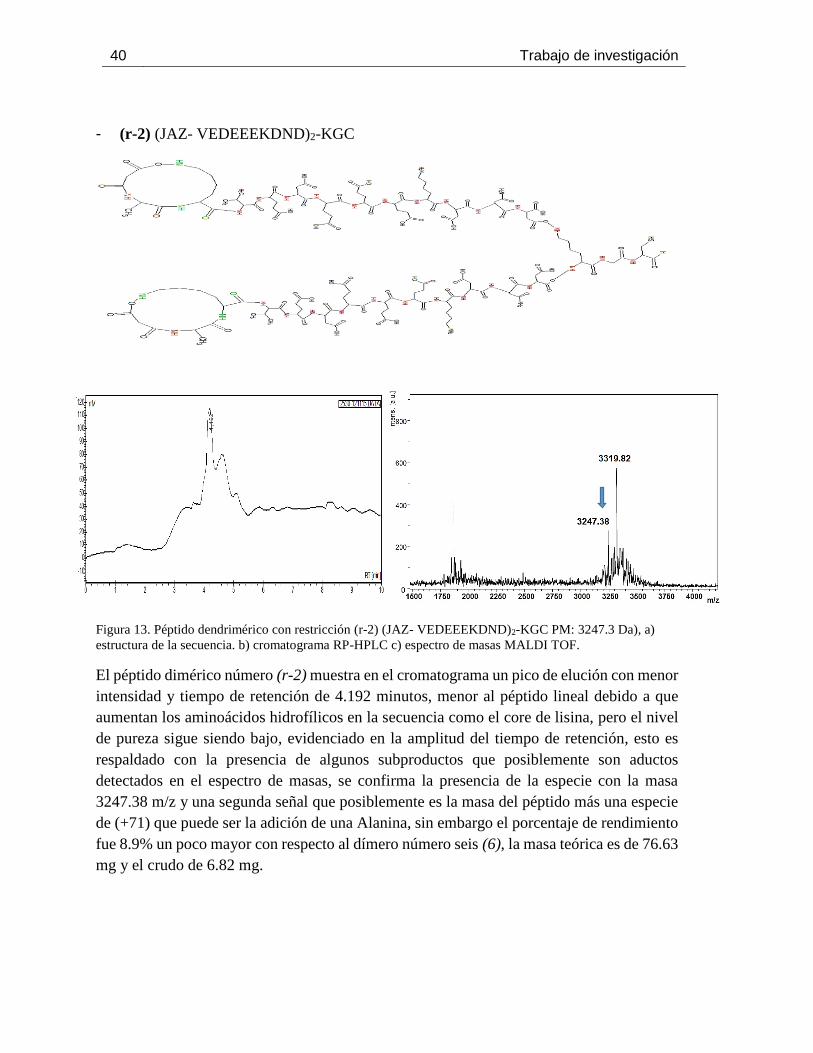
40 Trabajo de investigación
- (r-2) (JAZ- VEDEEEKDND)2-KGC
Figura 13. Péptido dendrimérico con restricción (r-2) (JAZ- VEDEEEKDND)2-KGC PM: 3247.3 Da), a)
estructura de la secuencia. b) cromatograma RP-HPLC c) espectro de masas MALDI TOF.
El péptido dimérico número (r-2) muestra en el cromatograma un pico de elución con menor
intensidad y tiempo de retención de 4.192 minutos, menor al péptido lineal debido a que
aumentan los aminoácidos hidrofílicos en la secuencia como el core de lisina, pero el nivel
de pureza sigue siendo bajo, evidenciado en la amplitud del tiempo de retención, esto es
respaldado con la presencia de algunos subproductos que posiblemente son aductos
detectados en el espectro de masas, se confirma la presencia de la especie con la masa
3247.38 m/z y una segunda señal que posiblemente es la masa del péptido más una especie
de (+71) que puede ser la adición de una Alanina, sin embargo el porcentaje de rendimiento
fue 8.9% un poco mayor con respecto al dímero número seis (6), la masa teórica es de 76.63
mg y el crudo de 6.82 mg.

41
Figura 14. Dicroísmo circular y deconvolución de: a) péptido lineal y b) péptido restringido c) Grafica
superpuesta de cada dicroísmo.
La grafica de dicroísmo del péptido lineal (izquierda) muestra una curva pronunciada con
una leve tendencia alfa helical sin embargo el espectro muestra un mínimo en 216 nm con -
20118, 9 de elipticidad molar y un máximo en 191 nm de 11013, 1 de elipticidad molar que
indican presencia de hoja beta en un 16%, ya que el aminoácido valina, ácido aspártico y
asparagina tienen tendencia a esta conformación. En cuanto al péptido dimérico (derecha)
se observa un cambio pronunciado en la estructura nativa, lo cual indica que el punto de
nucleación ejerce un posible efecto en la conformación del fragmento lineal, mostrando en
el espectro una estructura secundaria alfa helical del 48%, con dos mínimos notables en 223
nm con -18090,9 de elipticidad molar y el otro en 214 nm con -21417,9 elipticidad molar, y
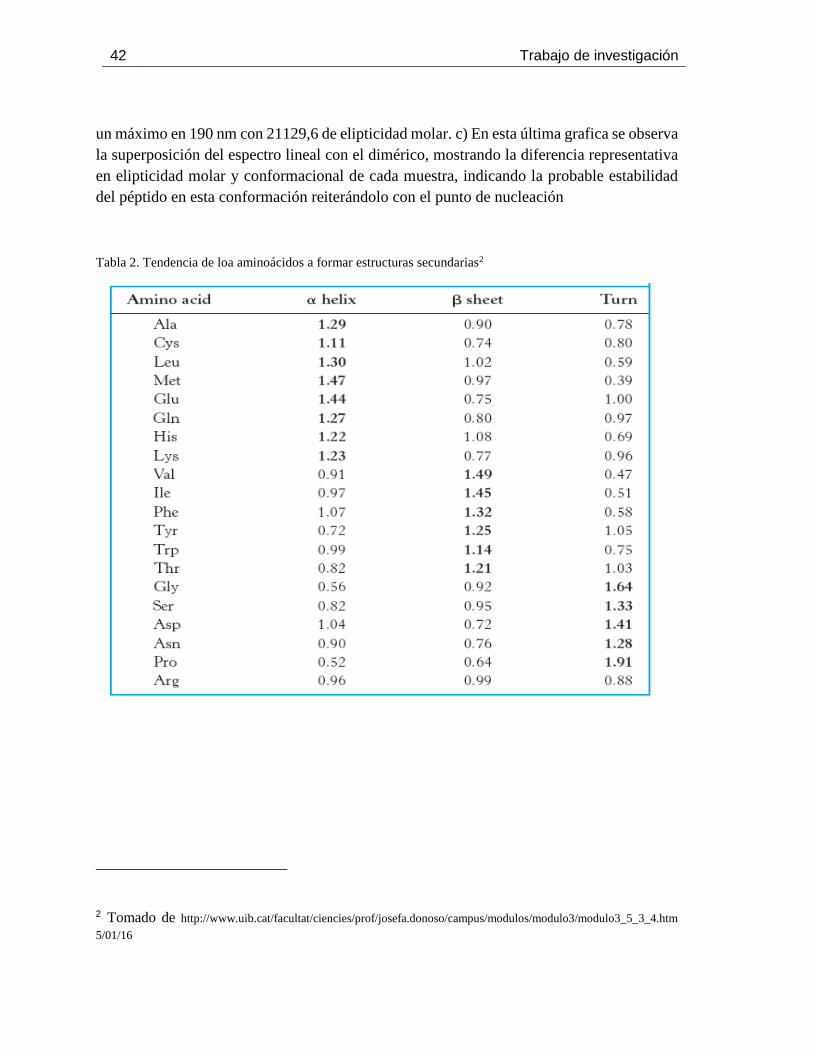
42 Trabajo de investigación
un máximo en 190 nm con 21129,6 de elipticidad molar. c) En esta última grafica se observa
la superposición del espectro lineal con el dimérico, mostrando la diferencia representativa
en elipticidad molar y conformacional de cada muestra, indicando la probable estabilidad
del péptido en esta conformación reiterándolo con el punto de nucleación
Tabla 2. Tendencia de loa aminoácidos a formar estructuras secundarias2
2 Tomado de http://www.uib.cat/facultat/ciencies/prof/josefa.donoso/campus/modulos/modulo3/modulo3_5_3_4.htm
5/01/16

43
4. Conclusiones y recomendaciones
4.1 Conclusiones
1. Todos los productos obtenidos fueron caracterizados por Cromatografía liquida de
alta eficiencia en fase reversa (RP-HPLC) y espectrometría de masas (MS), y la
tendencia conformacional por dicroísmo circular (CD).
2. Se pudo sintetizar el fragmento lineal correspondiente a la secuencia
LHEDQYAQILQE, el cual presentó una tendencia de hoja-β y al sintetizar la misma
secuencia con la restricción conformacional en DDC y el sitio de nucleación
disminuyo la tendencia inicial de su estructura nativa. En la segunda síntesis se pudo
observar que el movimiento de dos aminoácidos cambia notablemente la estructura
y la introducción del punto de nucleación a la secuencia en el constructo
dendrimérico acentúa la tendencia alfa-helical del péptido.
3. El fragmento peptídico con secuencia DEEEKDNDDN, al ser sintetizado mostró
una cierta tendencia de hoja-β y al introducir el punto de nucleación en la secuencia
en una plataforma peptídica, se estabiliza una estructura con cierta tendencia alfa-
helical. Sin embargo en la segunda síntesis del péptido el movimiento de dos
aminoácidos cambia notablemente la estructura y la introducción del punto de
nucleación a la secuencia en el constructo dendrimérico estabiliza notoriamente la
tendencia alfa-helical del péptido.
4. El sitio de nucleación “JAZ” (en proceso de patente) muestra que puede estabilizar
fragmentos con tendencia α-helical en macromoléculas peptídicas tipo DDC.
4.2 Recomendaciones
1. Se propone inocular los péptidos seleccionados en ratones y analizar los sueros
obtenidos en ratones contra los mismos péptidos para verificar su inmunogenicidad,
es decir, para que estas secuencias no solamente puedan reconocer sino producir
anticuerpos.
2. Llevar a cabo ensayos de Elisa con extractos de hoja de la planta de arroz, para
verificar si es posible la identificación de la proteína de estudio por este medio
inmunoenzimático.

44 Trabajo de investigación
A. Anexo 1. Tabla de los 20 aminoácidos naturales
Tomado de:
http://chemwiki.ucdavis.edu/Wikitexts/University_of_California_Davis/UCD_Chem_115_Lab_Manual/Lab_7%3A_Ele
ctrospray_Mass_Spectrometry

45
B. Anexo 2. Solución de Ninhidrina y Azul de
Bromofenol
MONITOREO DEL ACOPLAMIENTO Y DESPROTECCION
El objetivo es verificar mediante una reacción calorimétrica el porcentaje de acople de cada
aminoácido en la síntesis de péptidos en fase sólida.
TEST DE KAISER
1. Preparar las siguientes soluciones:
1.1 Solución 1: Disolver 40 g de fenol p.a. en 10 mL de etanol absoluto y guardar
en un frasco ámbar y en un ambiente fresco).
1.2 Stock de KCN: Disolver 65 mg de KCN en 100 mL de agua.
1.3 Solución 2: Luego tomar 1 mL de esta solución y completar hasta 50 mL con
piridina (recién destilada sobre ninhidrina).
1.4 Solución A: Mezclar las soluciones 1 y 2 en proporción de volúmenes de 1:10 y
agitar. Almacenar en la oscuridad.
1.3 Solución B: Disolver 1.25 g de ninhidrina en 25 mL de etanol absoluto.
2. Colocar una pequeña cantidad de la resina-péptido a ensayar en el respectivo tubo
de ensayo.
3. Adicionar 3 gotas de A y 1 gota de B y agite
4. Realizar un blanco de reactivos.
5. Calentar los tubos de ensayo en un bloque de calentamiento por 3 minutos a 100ºC.
6. Resultados:
a. Resina-péptido y solución azul a verdosa: ensayo positivo.
b. Resina-péptido incolora: ensayo negativo.

46 Trabajo de investigación
AZUL DE BROMOFENOL
1. Preparar la siguiente solución:
1.1 Disolver 0.25 g de azul de bromofenol en 25 mL de DMF.
2. Colocar una pequeña cantidad de resina-péptido en un tubo de microcentrífuga de 0.6 mL.
3. Adicionar 5 gotas de la solución del reactivo, agite y observe.
4. Resultados:
a. Resina-péptido y solución azul a verdosa: ensayo positivo.
b. Resina-péptido incolora: ensayo negativo.

47
C. Anexo 3. Grupos Protectores, Resinas y Brazos
Espaciadores

48 Trabajo de investigación
Bibliografía
1. FEDERACION NACIONAL DE ARROCEROS (FEDEARROZ) FNDA.
FEDEARROZ y Universidad Nacional construyen las bases de la investigación en lineas
transgénicas de arroz. ARROZ. Septiembre-Octubre 2010;58(488).
2. Cárdenas RM, Pérez N, Cristo E, González MC, Fabré L. Estudio sobre el
comportamiento de líneas y variedades de arroz (Oryza sativa Lin.) ante la infección por el
hongo Pyricularia grisea Sacc. Cultivos Tropicales. 2005;26(4):83-7.
3. Pantoja A FA, Correa F, Sanint L, Ramirez A. . MIP en Arroz: Manejo integrado de
plagas; Artrópodos, enfermedades y malezas. Cali Colombia & Caracas Venezuela.: CIAT,
FEDEARROZ & FLAR; 1997.
4. Oltra JRi. Consejeria de agricultura y pesca. EL ARROZ en cultivo ecológico.
Fertilidad de la tierra: revista de agricultura ecológica. 2004(18):32-6.
5. Garcés Fiallos FR, Díaz Coronel TG, Aguirre Calderón ÁJ. SEVERIDAD DE LA
QUEMAZÓN (Pyricularia oryzae Cav.) EN GERMOPLASMA DE ARROZ F1 EN LA
ZONA CENTRAL DEL LITORAL ECUATORIANO. Ciencia y Tecnología (1390-4051).
2012;5(2).
6. Nenínger LH, Hidalgo E, Barrios LM, Pueyo M. Hongos presentes en semillas de
arroz (Oryza sativa L.) en Cuba. Fitosanidad. 2003;7(3):7-11.
7. AGROPECUARIA CVDC. COMISIÓN VERACRUZANA DE
COMERCIALIZACIÓN AGROPECUARIA "MONOGRAFIA DEL ARROZ". Gobierno
del estado de Veracruz-Mexico [Internet]. 2008. Available from:
http://portal.veracruz.gob.mx/pls/portal/docs/PAGE/COVECAINICIO/IMAGENES/ARC
HIVOSPDF/ARCHIVOSDIFUSION/MONOGRAFIA%20ARROZ2010.PDF.
8. Fundora. IBG. ESPECTRO PATOLÓGICO DE LAS PRINCIPALES
ENFERMEDADES DEL CULTIVO DEL ARROZ. Ing Agrónoma, Profesor Asistente,
Departamento de Agricultura Facultad de Agronomía Universidad de Matanzas
9. Hayashi N, Inoue H, Kato T, Funao T, Shirota M, Shimizu T, et al. Durable panicle
blast‐resistance gene Pb1 encodes an atypical CC‐NBS‐LRR protein and was generated by
acquiring a promoter through local genome duplication. The Plant Journal. 2010;64(3):498-
510.
10. Ghatak A, Willocquet L, Savary S, Kumar J. Variability in Aggressiveness of Rice
Blast Isolates Originating from Rice Leaves and Necks: A Case of Pathogen Specialization?
PloS one. 2013;8(6):e66180.
11. EDGAR ANDRES CABALLERO JJDF. CARACTERIZACIÓN DE PROTEÍNAS
EXPRESADAS POR EL GEN Pb1, ASOCIADO A LA RESISTENCIA AL AÑUBLO
DEL ARROZ (Magnaporthe oryzae) EN LAS VARIEDADES DE ARROZ (Oryza sativa)
FEDEARROZ 50, MOCARÍ Y LV 1645 IRRADIADAS CON 60Co. BOGOTÁ, D.C.:
UNIVERSIDAD DISTRITAL FRANCISCO JOSÉ DE CALDAS; 2011.
12. CHURQUE NER. SÍNTESIS, PURIFICACIÓN Y CARACTERIZACIÓN DE UN
FRAGMENTO DE LA PROTEÍNA DE 2RGL: RICE BGLU1 BETA-GLUCOSIDASA DE

49
ORYZA SATIVA [Investigativa]. BOGOTÁ D.C: UNIVERSIDAD DISTRITAL
FRANCISCO JOSE DE CALDAS; 2012.
13. Degiovanni B, Víctor M, Martínez R, César P, Motta O. Producción eco-eficiente
del arroz en América Latina: CIAT; 2010.
14. Zeigler R, Barclay A. The Relevance of Rice. Rice. 2008;1(1):3-10.
15. Cesar M. Guía Técnica El Cultivo de Arroz. Santo Domingo, República Dominicana:
Centro para el Desarrollo Agropecuario y Forestal, Inc. (CEDAF); 2010.
16. PORTADA E. El sector del arroz en los Acuerdos Internacionales y en la
Organización Mundial del Comercio. SG de Comercio Exterior de Productos
Agroalimentarios BOLETÍN ECONÓMICO DE ICE 2007;No 2917.
17. Bondades y Beneficios de Arroz [Internet]. [cited 14/05/2015].
18. FAO FaAO. Rice Market Monitor. Rome- Italy2008.
19. Arregocés O, Rosero M, González J. Guía de estudio: Morfología de la planta de
arroz. Cali: Centro Internacional de Agricultura Tropical. 2005.
20. Induarroz AC. Politica Comercial Para el Arroz2012:[72 p.]. Available from:
http://www.andi.com.co/cai/Documents/ESTUDIO%20FEDESARROLLO%202012.pdf.
21. Ministerio de Agricultura y Desarrollo Rural D. El arroz: Produccion en Colombia.
Boletin Mensual INSUMOS Y FACTORES ASOCIADOS A LA PRODUCCION
AGROPECUARIA. JUNIO 2013 01 - 04 - 2015:36.
22. Agropecuarias IINdI. Analítico: Manual del cultivo del arroz. Guayas, EcuadorFeb.
2007.
23. Degiovanni B, Víctor M, Degiovanni V, Martínez R, César P, Motta O. Producción
eco-eficiente del arroz en América Latina: CIAT; 2010.
24. Fundora. BG. ESPECTRO PATOLÓGICO DE LAS PRINCIPALES
ENFERMEDADES DEL CULTIVO DEL ARROZ. Ing Agrónoma, Profesor Asistente,
Departamento de Agricultura Facultad de
Agronomía Universidad de Matanzas
25. Fernández MG. Piriculariosis y otras enfermedades del Arroz: Cultivos del arroz en
las marismas del Guadalquivir. Agricultura: Revista agropecuaria. 2001(827):358-9.
26. Ou SH. Rice Diseases: Commonwealth Mycological Institute. IRRI; 1985.
27. Prabhu AS, Filippi MC, Castro N. Pathogenic variation among isolates of Pyricularia
oryzae affecting rice, wheat, and grasses in Brazil. Tropical Pest Management.
1992;38(4):367-71.
28. Disponible en: http://www.tecnicoagricola.es/Pyricularia-oryzae-en-arroz/,
consultado 20-02-15
29. Klaubauf S, Tharreau D, Fournier E, Groenewald JZ, Crous PW, de Vries RP, et al.
Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Studies in Mycology.
2014;79(0):85-120.
30. Meneses C, Gutiérrez Y, García R, Antigua P, Gómez S, Correa V, et al. Guía para
el trabajo de campo en el manejo integrado de plagas del arroz. Cuarta ed. La Habana, Cuba:
CIAT & Fondo Latinoamericano para Arroz de Riego (FLAR); 2001.
31. Guimaraes EP. Seleccion recurrente en arroz: CIAT; 1997.
32. Wit PJ. How plants recognize pathogens and defend themselves. Cellular and
Molecular Life Sciences. 2007;64(21):2726-32.

50 Trabajo de investigación
33. Agrios G. How plants defend themselves against pathogens. Plant pathology. 2005.
34. Zhou B, Peng K, Zhaohui C, Wang S, Zhang Q. The defense-responsive genes
showing enhanced and repressed expression after pathogen infection in rice (Oryza sativa
L.). Science in China Series C: Life Sciences. 2002;45(5):449-67.
35. Inoue H, Hayashi N, Matsushita A, Xinqiong L, Nakayama A, Sugano S, et al. Blast
resistance of CC-NB-LRR protein Pb1 is mediated by WRKY45 through protein–protein
interaction. Proceedings of the National Academy of Sciences. 2013;110(23):9577-82.
36. Bonman JM. Durable resistance to rice blast disease-environmental influences.
Euphytica. 1992;63(1-2):115-23.
37. Mitchell AR. R. Bruce Merrifield and Solid-Phase Peptide Synthesis: A Historical
Assessment. Lawrence Livermore National Laboratory,USA. December 5, 2007(UCRL-
JRNL-237037).
38. Guzmán F, Barberis S, Illanes A. Peptide synthesis: chemical or enzymatic2007.
39. Wieland T. The history of peptide chemistry. ChemInform. 1996;27(29).
40. MERRIFIELD B. Concept and Early Development of Solid-Phase Peptide
Synthesis. 1997;289.
41. DOONAN S. Peptides and Proteins. Thomas Graham House SP, editor.
UNIVERSITY OF EAST LONDON: The Royal Society of Chemistry.; 2002.
42. Borgia JA, Fields GB. Chemical synthesis of proteins. Trends in Biotechnology.
2000;18(6):243-51.
43. Fayna Garcı´a-Martı´n PW, Rene´ Steinauer, Simon Coˆ te´, Judit Tulla-Puche,
Fernando Albericio. The Synergy of ChemMatrix Resin1 and Pseudoproline Building
Blocks Renders RANTES, A Complex Aggregated Chemokine*. Biopolymers (Peptide
Science). 29 June 2006;247.
44. Merrifield B. 3 - Solid-Phase Peptide Synthesis. In: Bernd G, editor. Peptides. San
Diego: Academic Press; 1995. p. 93-169.
45. Albericio F. Solid-phase synthesis: a practical guide: CRC Press; 2000.
46. Andreu D, Rivas L. Péptidos en biología y biomedicina: Editorial CSIC-CSIC Press;
1997.
47. Han S-Y, Kim Y-A. Recent development of peptide coupling reagents in organic
synthesis. Tetrahedron. 2004;60(11):2447-67.
48. Benoiton NL. Chemistry of peptide synthesis: CRC Press; 2005.
49. Milton RdL, Milton SC, Adams PA. Prediction of difficult sequences in solid-phase
peptide synthesis. Journal of the American Chemical Society. 1990;112(16):6039-46.
50. Tuschscherer G, Mutter M. Template assisted protein de novo design. Pure and
Applied chemistry. 1996;68(11):2153-62.
51. Epton R. Innovation and perspectives in solid phase synthesis: Intercept; 1992.
52. Anfinsen CB. Principles that govern the folding of protein chains. Science.
1973;181(4096):223-30.
53. Sewald N, Jakubke HD. Peptide synthesis. Peptides: Chemistry and Biology, Second
Edition.175-315.
54. Ross NT, Katt WP, Hamilton AD. Synthetic mimetics of protein secondary structure
domains. Philosophical Transactions of the Royal Society of London A: Mathematical,
Physical and Engineering Sciences. 2010;368(1914):989-1008.

51
55. Sadler KT, James P. Peptide dendrimers: applications and synthesis. Reviews in
Molecular Biotechnology. 2002;90(3–4):195-229.
56. Tam JP. Synthetic peptide vaccine design: synthesis and properties of a high-density
multiple antigenic peptide system. Proceedings of the National Academy of Sciences.
1988;85(15):5409-13.
57. JULIO C. CALVO ZR, CLAUDIA CARVAJAL, ALEXANDER BERNAL,
FRANCISCO CHAVES, JOSE MANUEL LOZANO MORENO, MARY TRUJILLO,
FANNY GUZMAN, MANUEL ELKIN PATARROYO,. Sintesis de Péptidos
Dendriméricos Usando la Estrategia de Doble Dimero. Revista Colombiana De Quimica,
Universidad Nacional de Colombia. 1999;v.28 fasc.1:27 - 34
58. Rivera Z, Granados G, Pinto M, Varón D, Carvajal C, Chaves F, et al. Double dimer
peptide constructs are immunogenic and protective against Plasmodium falciparum in the
experimental Aotus monkey model. The Journal of Peptide Research. 2002;59(2):62-70.
59. Houghten RA. General method for the rapid solid-phase synthesis of large numbers
of peptides: specificity of antigen-antibody interaction at the level of individual amino acids.
Proceedings of the National Academy of Sciences. 1985;82(15):5131-5.
60. Satterthwait A, Chiang L-C, Arrhenius T, Cabezas E, Zavala F, Dyson H, et al. The
conformational restriction of synthetic vaccines for malaria. Bulletin of the World Health
Organization. 1990;68(Suppl):17.
61. Calvo JC, Martínez JC, Patarroyo ME, Satterthwait AC. DE LA PREDICCIÓN AL
ANÁLISIS ANTIGÉNICO: TRAS LAS HUELLAS DE UN EPÍTOPE
CONFORMACIONAL. Revista de la Academia Colombiana de Ciencias Exactas, Físicas
y Naturales. 2003;27(000204769):133-40.
62. CALVO JC. Sintesis y caracterizacion del acido 5,5-dimetoxipentanoico Centro De
Investigaciones Y Desarrollo Cientifico Universidad Distrital Francisco Jose De Caldas
Revista Científica, Colombia. 2005;v. fasc.7:63 - 76.
63. J Yang RY, A Roy, D Xu, J Poisson, Y Zhang. The I-TASSER Suite: Protein
structure and function prediction. Nature Methods. 2015;12:7-8.
Top Related